Enhancing Hydrogen Production from Chlorella sp. Biomass by Pre-Hydrolysis with Simultaneous Saccharification and Fermentation (PSSF)


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae and Inoculum
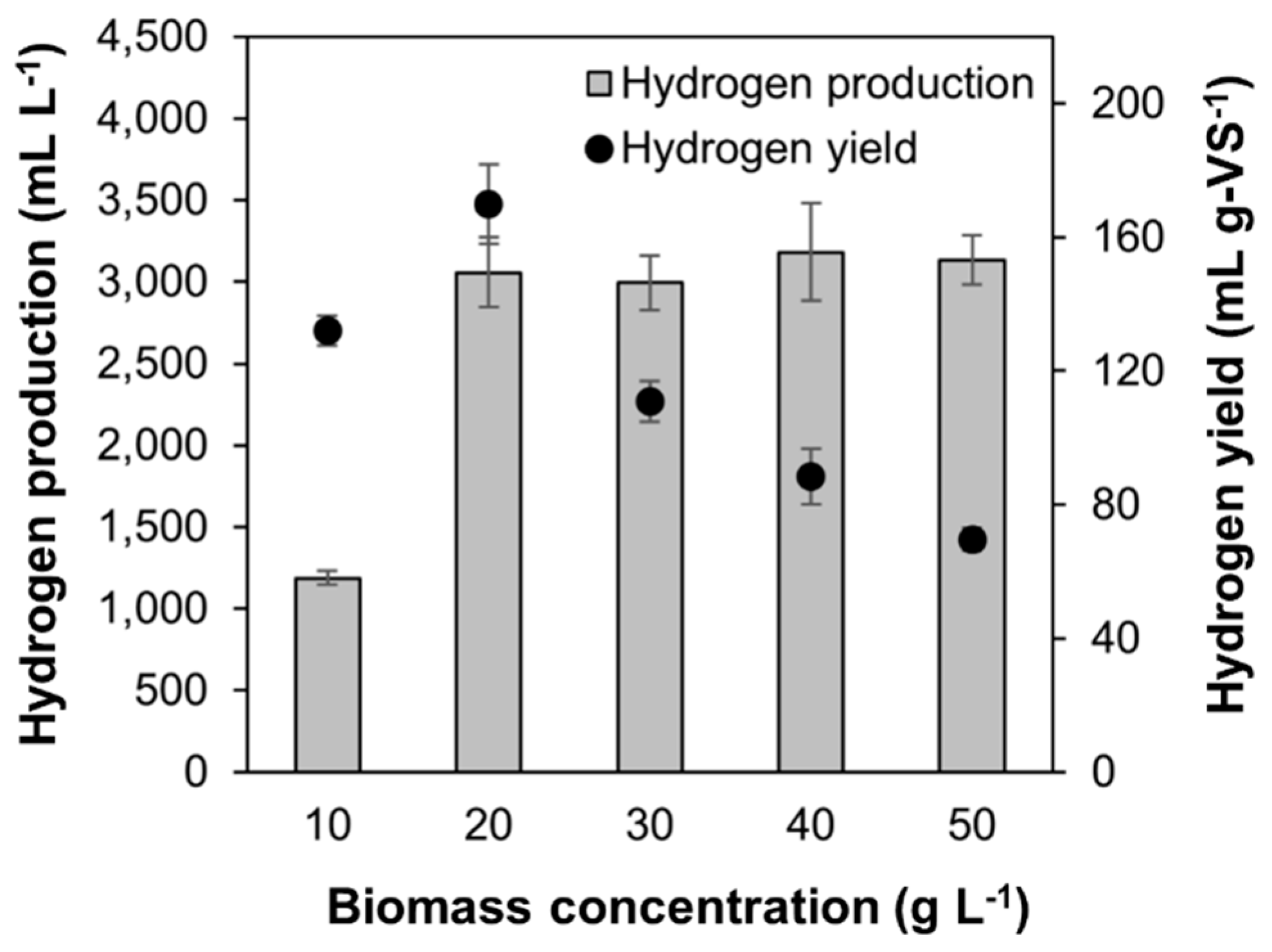
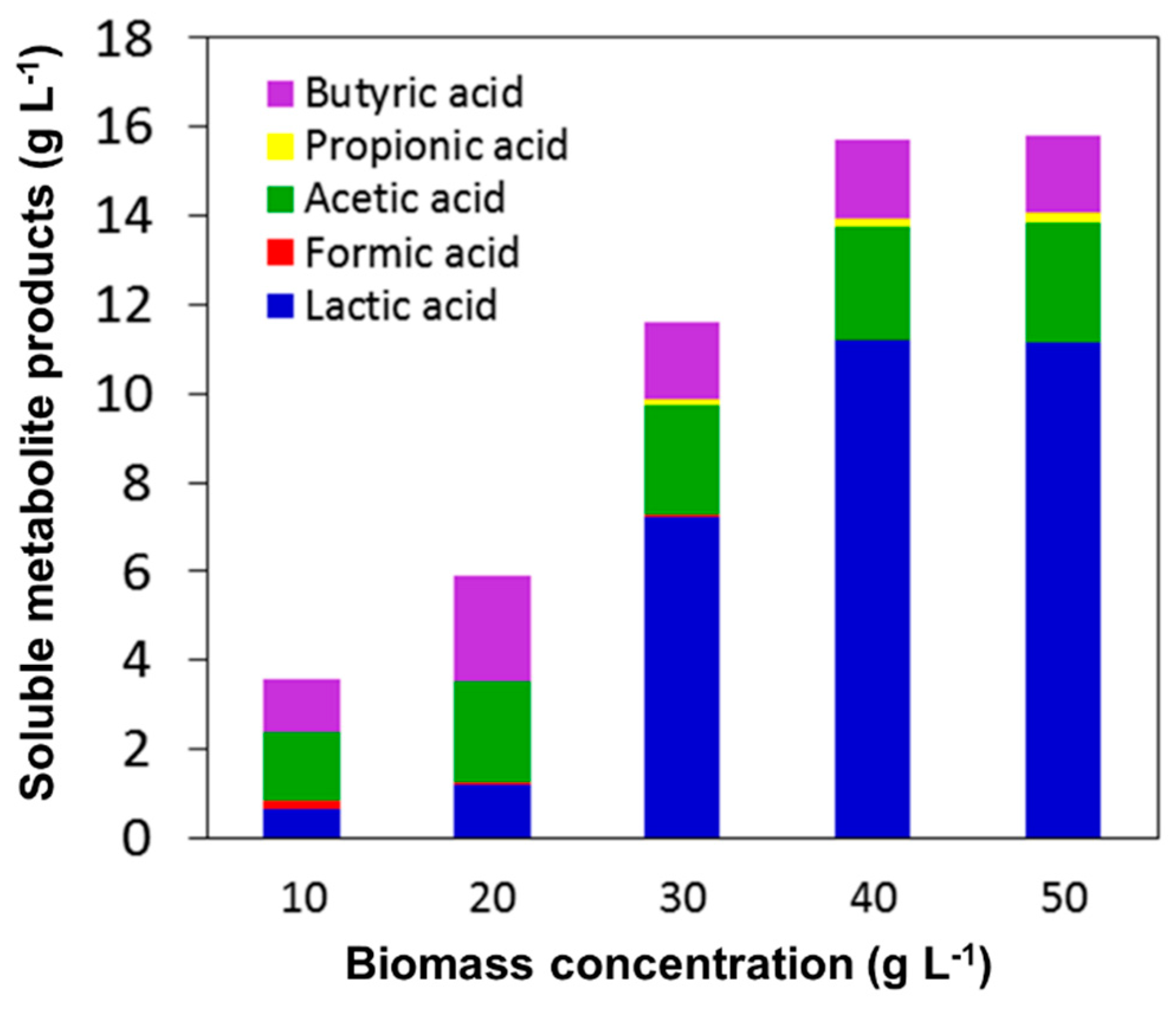
2.2. Effect of Initial Biomass Concentration on Hydrogen Production under SSF
2.3. Pre-Hydrolysis with Simultaneous Saccharification and Fermentation (PSSF) of Chlorella sp. Biomass
2.4. Analytical Methods
3. Results and Discussion
3.1. Effect of Initial Biomass Concentration on Hydrogen Production under SSF
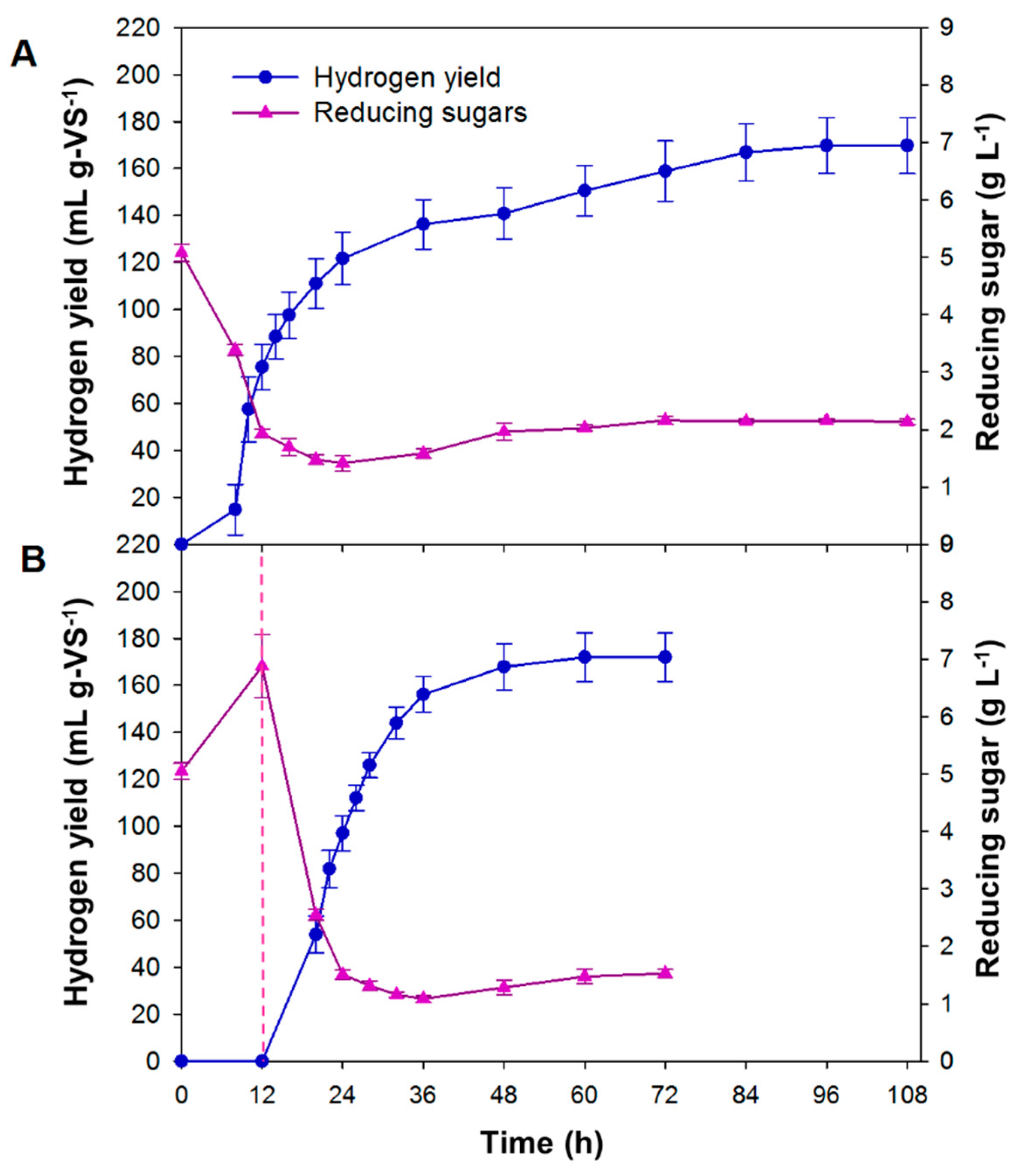
3.2. SSF and PSSF of Chlorella sp. Biomass for Hydrogen Production
3.3. Applicability of PSSF for Hydrogen Production from Chlorella sp. Biomass
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fayaz, H.; Saidur, R.; Razali, N.; Anuar, F.S.; Saleman, A.R.; Islam, M.R. An overview of hydrogen as a vehicle fuel. Renew. Sustain. Energy Rev. 2012, 16, 5511–5528. [Google Scholar] [CrossRef]
- Salvi, B.L.; Subramanian, K.A. Sustainable development of road transportation sector using hydrogen energy system. Renew. Sustain. Energy Rev. 2015, 51, 1132–1155. [Google Scholar] [CrossRef]
- Nagarajan, D.; Lee, D.J.; Kondo, A.; Chang, J.S. Recent insights into biohydrogen production by microalgae—From biophotolysis to dark fermentation. Bioresour. Technol. 2017, 227, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Zhen, G.; Sivagurunathan, P.; Bakonyi, P.; Nemestóthy, N.; Bélafi-Bakó, K.; Kobayashi, T.; Xu, K.-Q. Biogenic H2 production from mixed microalgae biomass: Impact of pH control and methanogenic inhibitor (BESA) addition. Biofuel Res. J. 2016, 3, 470–474. [Google Scholar] [CrossRef]
- Moreira, F.S.; Machado, R.G.; Romão, B.B.; Batista, F.R.X.; Ferreira, J.S.; Cardoso, V.L. Improvement of hydrogen production by biological route using repeated batch cycles. Process Biochem. 2017, 58, 60–68. [Google Scholar] [CrossRef]
- Qiu, C.; Shi, P.; Xiao, S.; Sun, L. Effects of pH and substrate concentrations on dark fermentative biohydrogen production from xylose by extreme thermophilic mixed culture. World J. Microbiol. Biotechnol. 2017, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Voloshin, R.A.; Rodionova, M.V.; Zharmukhamedov, S.K.; Nejat Veziroglu, T.; Allakhverdiev, S.I. Review: Biofuel production from plant and algal biomass. Int. J. Hydrogen Energy 2016, 41, 17257–17273. [Google Scholar] [CrossRef]
- Holladay, J.D.; Hu, J.; King, D.L.; Wang, Y. An overview of hydrogen production technologies. Catal. Today 2009, 139, 244–260. [Google Scholar] [CrossRef]
- Rittmann, S.; Herwig, C. A comprehensive and quantitative review of dark fermentative biohydrogen production. Microb. Cell Fact. 2012, 11, 115. [Google Scholar] [CrossRef] [Green Version]
- Stephen, A.J.; Archer, S.A.; Orozco, R.L.; Macaskie, L.E. Advances and bottlenecks in microbial hydrogen production. Microb. Biotechnol. 2017, 10, 1120–1127. [Google Scholar] [CrossRef] [Green Version]
- Balachandar, G.; Khanna, N.; Das, D. Biohydrogen production from organic wastes by dark fermentation. In Biohydrogen; Pandey, A., Chang, J.-S., Hallenbeck, P., Larroche, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 103–144. ISBN 9780444595553. [Google Scholar]
- El-Dalatony, M.; Salama, E.-S.; Kurade, M.; Hassan, S.; Oh, S.-E.; Kim, S.; Jeon, B.-H. Utilization of microalgal biofractions for bioethanol, higher alcohols, and biodiesel production: A review. Energies 2017, 10, 2110. [Google Scholar] [CrossRef]
- Medipally, S.R.; Yusoff, F.M.; Banerjee, S.; Shariff, M. Microalgae as sustainable renewable energy feedstock for biofuel production. BioMed Res. Int. 2015, 2015, 519513. [Google Scholar] [CrossRef] [PubMed]
- Yeong, T.K.; Jiao, K.; Zeng, X.; Lin, L.; Pan, S.; Danquah, M.K. Microalgae for biobutanol production—Technology evaluation and value proposition. Algal Res. 2018, 31, 367–376. [Google Scholar] [CrossRef]
- Sambusiti, C.; Bellucci, M.; Zabaniotou, A.; Beneduce, L.; Monlau, F. Algae as promising feedstocks for fermentative biohydrogen production according to a biorefinery approach: A comprehensive review. Renew. Sustain. Energy Rev. 2015, 44, 20–36. [Google Scholar] [CrossRef]
- Argun, H.; Gokfi, P.; Karapinar, I. Biohydrogen production potential of different biomass sources. In Biohydrogen Production: Sustainability of Current Technology and Future Perspective; Singh, A., Rathore, D., Eds.; Springer: New Delhi, India, 2016; pp. 11–48. ISBN 978-81-322-3575-0. [Google Scholar]
- Wieczorek, N.; Kucuker, M.A.; Kuchta, K. Fermentative hydrogen and methane production from microalgal biomass (Chlorella vulgaris) in a two-stage combined process. Appl. Energy 2014, 132, 108–117. [Google Scholar] [CrossRef]
- Cheng, J.; Lin, R.; Song, W.; Xia, A.; Zhou, J.; Cen, K. Enhancement of fermentative hydrogen production from hydrolyzed water hyacinth with activated carbon detoxification and bacteria domestication. Int. J. Hydrogen Energy 2015, 40, 2545–2551. [Google Scholar] [CrossRef]
- Dickinson, S.; Mientus, M.; Frey, D.; Amini-Hajibashi, A.; Ozturk, S.; Shaikh, F.; Sengupta, D.; El-Halwagi, M.M. A review of biodiesel production from microalgae. Clean Technol. Environ. Policy 2017, 19, 637–668. [Google Scholar] [CrossRef]
- Heaven, S.; Milledge, J.; Zhang, Y. Comments on “Anaerobic digestion of microalgae as a necessary step to make microalgal biodiesel sustainable”. Biotechnol. Adv. 2011, 29, 164–167. [Google Scholar] [CrossRef]
- Phanduang, O.; Lunprom, S.; Salakkam, A.; Reungsang, A. Anaerobic solid-state fermentation of bio-hydrogen from microalgal Chlorella sp. biomass. Int. J. Hydrogen Energy 2017, 42, 9650–9659. [Google Scholar] [CrossRef]
- Lakaniemi, A.-M.; Hulatt, C.J.; Thomas, D.N.; Tuovinen, O.H.; Puhakka, J.A. Biogenic hydrogen and methane production from Chlorella vulgaris and Dunaliella tertiolecta biomass. Biotechnol. Biofuels 2011, 4, 34. [Google Scholar] [CrossRef]
- Pagliaccia, P.; Gallipoli, A.; Gianico, A.; Montecchio, D.; Braguglia, C.M. Single stage anaerobic bioconversion of food waste in mono and co-digestion with olive husks: Impact of thermal pretreatment on hydrogen and methane production. Int. J. Hydrogen Energy 2016, 41, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.M.; Han, S.K.; Kim, J.T.; Lee, C.Y. Optimization of combined (acid + thermal) pretreatment for enhanced dark fermentative H2 production from Chlorella vulgaris using response surface methodology. Int. Biodeterior. Biodegrad. 2016, 108, 191–197. [Google Scholar] [CrossRef]
- Sivagurunathan, P.; Kumar, G.; Kobayashi, T.; Xu, K.; Kim, S.H. Effects of various dilute acid pretreatments on the biochemical hydrogen production potential of marine macroalgal biomass. Int. J. Hydrogen Energy 2017, 42, 27600–27606. [Google Scholar] [CrossRef]
- Jegannathan, K.R.; Nielsen, P.H. Environmental assessment of enzyme use in industrial production-a literature review. J. Clean. Prod. 2013, 42, 228–240. [Google Scholar] [CrossRef]
- Quéméneur, M.; Bittel, M.; Trably, E.; Dumas, C.; Fourage, L.; Ravot, G.; Steyer, J.-P.; Carrère, H. Effect of enzyme addition on fermentative hydrogen production from wheat straw. Int. J. Hydrogen Energy 2012, 37, 10639–10647. [Google Scholar] [CrossRef]
- de Barros, E.M.; Carvalho, V.M.; Rodrigues, T.H.S.; Rocha, M.V.P.; Gonçalves, L.R.B. Comparison of strategies for the simultaneous saccharification and fermentation of cashew apple bagasse using a thermotolerant Kluyveromyces marxianus to enhance cellulosic ethanol production. Chem. Eng. J. 2017, 307, 939–947. [Google Scholar] [CrossRef]
- Tsai, M.H.; Lee, W.C.; Kuan, W.C.; Sirisansaneeyakul, S.; Savarajara (Akaracharanya), A. Evaluation of different pretreatments of Napier grass for enzymatic saccharification and ethanol production. Energy Sci. Eng. 2018, 6, 683–692. [Google Scholar] [CrossRef]
- Chen, K.; Liu, D.; Chen, X.; Fan, Z. Kinetic analysis of Fe(II)-promoted ethanol preparation from cornstalks. BioResources 2018, 13, 4973–4985. [Google Scholar] [CrossRef]
- Lin, Y.S.; Lee, W.C. Simultaneous saccharification and fermentation of alkali-pretreated cogongrass for bioethanol production. BioResources 2011, 6, 2744–2756. [Google Scholar] [CrossRef]
- Fulke, A.B.; Mudliar, S.N.; Yadav, R.; Shekh, A.; Srinivasan, N.; Ramanan, R.; Krishnamurthi, K.; Devi, S.S.; Chakrabarti, T. Bio-mitigation of CO2, calcite formation and simultaneous biodiesel precursors production using Chlorella sp. Bioresour. Technol. 2010, 101, 8473–8476. [Google Scholar] [CrossRef] [PubMed]
- Chinnasamy, S.; Ramakrishnan, B.; Bhatnagar, A.; Das, K.; Chinnasamy, S.; Ramakrishnan, B.; Bhatnagar, A.; Das, K.C. Biomass production potential of a wastewater alga Chlorella vulgaris ARC 1 under elevated levels of CO2 and temperature. Int. J. Mol. Sci. 2009, 10, 518–532. [Google Scholar] [CrossRef] [PubMed]
- Fangkum, A.; Reungsang, A. Biohydrogen production from mixed xylose/arabinose at thermophilic temperature by anaerobic mixed cultures in elephant dung. Int. J. Hydrogen Energy 2011, 36, 13928–13938. [Google Scholar] [CrossRef]
- Zhang, H.; Liao, Q.; Fu, Q.; Chen, H.; Huang, Y.; Xia, A.; Zhu, X.; Reungsang, A.; Liu, Z.; Li, J. Rheological properties of microalgae slurry under subcritical conditions for hydrothermal hydrolysis systems. Algal Res. 2018, 33, 78–83. [Google Scholar] [CrossRef]
- Owen, W.F.; Stuckey, D.C.; Healy, J.B.; Young, L.Y.; McCarty, P.L. Bioassay for monitoring biochemical methane potential and anaerobic toxicity. Water Res. 1979, 13, 485–492. [Google Scholar] [CrossRef]
- Association of Official Analysis Chemists. Standard Methods for the Examination of Water and Wastewater, 19th ed.; American Public Health Association: Washington, DC, USA, 2012; ISBN 0935584544. [Google Scholar]
- Adney, B.; Baker, J. Measurement of Cellulase Activities Laboratory; National Renewable Energy Laboratory: Golden, CO, USA, 2008.
- Bryjak, J. Glucoamylase, α-amylase and β-amylase immobilisation on acrylic carriers. Biochem. Eng. J. 2003, 16, 347–355. [Google Scholar] [CrossRef]
- Sitthikitpanya, S.; Reungsang, A.; Prasertsan, P.; Khanal, S.K. Two-stage thermophilic bio-hydrogen and methane production from oil palm trunk hydrolysate using Thermoanaerobacterium thermosaccharolyticum KKU19. Int. J. Hydrogen Energy 2017, 42, 28222–28232. [Google Scholar] [CrossRef]
- Zheng, X.J.; Yu, H.Q. Inhibitory effects of butyrate on biological hydrogen production with mixed anaerobic cultures. J. Environ. Manag. 2005, 74, 65–70. [Google Scholar] [CrossRef]
- Khanal, S.K.; Chen, W.H.; Li, L.; Sung, S. Biological hydrogen production: Effects of pH and intermediate products. Int. J. Hydrogen Energy 2004, 29, 1123–1131. [Google Scholar] [CrossRef]
- Nualsri, C.; Kongjan, P.; Reungsang, A. Direct integration of CSTR-UASB reactors for two-stage hydrogen and methane production from sugarcane syrup. Int. J. Hydrogen Energy 2016, 41, 17884–17895. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Wu, Z.; Shi, X. Rheological properties of Chlorella pyrenoidosa culture grown heterotrophically in a fermentor. J. Appl. Phycol. 2008, 20, 279–282. [Google Scholar] [CrossRef]
- Wileman, A.; Ozkan, A.; Berberoglu, H. Rheological properties of algae slurries for minimizing harvesting energy requirements in biofuel production. Bioresour. Technol. 2012, 104, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Schneider, N.; Gerber, M. Correlation between viscosity, temperature and total solid content of algal biomass. Bioresour. Technol. 2014, 170, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Adesanya, V.O.; Vadillo, D.C.; Mackley, M.R. The rheological characterization of algae suspensions for the production of biofuels. J. Rheol. 2012, 56, 925–939. [Google Scholar] [CrossRef]
- Srinorakutara, T.; Subkaree, Y.; Boonvitthya, N.; Kunhanon, T.; Bamrungchue, N. Effect of commercial cellulase enzymes on ethanol production from pretreated rice straw at high solid loading. J. Food Sci. Eng. 2015, 5, 67–75. [Google Scholar] [CrossRef]
- Park, C.; Lee, J.H.; Yang, X.; Yoo, H.Y.; Lee, J.H.; Lee, S.K.; Kim, S.W. Enhancement of hydrolysis of Chlorella vulgaris by hydrochloric acid. Bioprocess Biosyst. Eng. 2016, 39, 1015–1021. [Google Scholar] [CrossRef]
- Wang, J.; Wan, W. The effect of substrate concentration on biohydrogen production by using kinetic models. Sci. China Ser. B Chem. 2008, 51, 1110–1117. [Google Scholar] [CrossRef]
- Reungsang, A.; Sreela-or, C. Bio-hydrogen production from pineapple waste extract by anaerobic mixed cultures. Energies 2013, 6, 2175–2190. [Google Scholar] [CrossRef]
- Sangyoka, S.; Reungsang, A.; Lin, C.-Y. Optimization of biohydrogen production from sugarcane bagasse by mixed cultures using a statistical method. Sustain. Environ. Res. 2016, 26, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Kumar, K.; Ghosh, S.; Das, D. Thermophilic biohydrogen production using pre-treated algal biomass as substrate. Biomass Bioenergy 2014, 61, 157–166. [Google Scholar] [CrossRef]
- Yun, Y.M.; Jung, K.W.; Kim, D.H.; Oh, Y.K.; Shin, H.S. Microalgal biomass as a feedstock for bio-hydrogen production. Int. J. Hydrogen Energy 2012, 37, 15533–15539. [Google Scholar] [CrossRef]
- Ghimire, A.; Frunzo, L.; Pirozzi, F.; Trably, E.; Escudie, R.; Lens, P.N.L.; Esposito, G. A review on dark fermentative biohydrogen production from organic biomass: Process parameters and use of by-products. Appl. Energy 2015, 144, 73–95. [Google Scholar] [CrossRef]
- Cappai, G.; Gioannis, G.D.; Muntoni, A.; Polettini, A.; Pomi, R.; Spiga, D. Effect of inoculum to substrate ratio (ISR) on hydrogen production through dark fermentation of food waste. In Proceedings of the Fifteenth International Waste Management and Landfill Symposium, Cagliari, Italy, 5–9 October 2015. [Google Scholar]
- Hallenbeck, P.C. Fermentative hydrogen production: Principles, progress, and prognosis. Int. J. Hydrogen Energy 2009, 34, 7379–7389. [Google Scholar] [CrossRef]
- Sikora, A.; Błaszczyk, M.; Jurkowski, M.; Zielenkiewicz, U. Lactic acid bacteria in hydrogen-producing consortia: On purpose or by coincidence. In Lactic Acid Bacteria—R&D for Food, Health and Livestock Purposes; Kongo, M., Ed.; InTech: Rijeka, Croatia, 2013; ISBN 9789533070940. [Google Scholar]
- Nualsri, C.; Reungsang, A.; Plangklang, P. Biochemical hydrogen and methane potential of sugarcane syrup using a two-stage anaerobic fermentation process. Ind. Crops Prod. 2016, 82, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Reungsang, A.; Sreela-Or, C.; Plangklang, P. Non-sterile bio-hydrogen fermentation from food waste in a continuous stirred tank reactor (CSTR): Performance and population analysis. Int. J. Hydrogen Energy 2013, 38, 15630–15637. [Google Scholar] [CrossRef]
- Cabrol, L.; Marone, A.; Tapia-Venegas, E.; Steyer, J.P.; Ruiz-Filippi, G.; Trably, E. Microbial ecology of fermentative hydrogen producing bioprocesses: Useful insights for driving the ecosystem function. FEMS Microbiol. Rev. 2017, 41, 158–181. [Google Scholar] [CrossRef] [PubMed]
- Cotana, F.; Cavalaglio, G.; Gelosia, M.; Coccia, V.; Petrozzi, A.; Ingles, D.; Pompili, E. A comparison between SHF and SSSF processes from cardoon for ethanol production. Ind. Crops Prod. 2015, 69, 424–432. [Google Scholar] [CrossRef]
- Hoyer, K.; Galbe, M.; Zacchi, G. The effect of prehydrolysis and improved mixing on high-solids batch simultaneous saccharification and fermentation of spruce to ethanol. Process Biochem. 2013, 48, 289–293. [Google Scholar] [CrossRef]
- de Souza, C.J.A.; Costa, D.A.; Rodrigues, M.Q.R.B.; dos Santos, A.F.; Lopes, M.R.; Abrantes, A.B.P.; dos Santos Costa, P.; Silveira, W.B.; Passos, F.M.L.; Fietto, L.G. The influence of presaccharification, fermentation temperature and yeast strain on ethanol production from sugarcane bagasse. Bioresour. Technol. 2012, 109, 63–69. [Google Scholar] [CrossRef]
- Yun, Y.-M.; Jung, K.-W.; Kim, D.-H.; Oh, Y.-K.; Cho, S.-K.; Shin, H.-S. Optimization of dark fermentative H2 production from microalgal biomass by combined (acid + ultrasonic) pretreatment. Bioresour. Technol. 2013, 141, 220–226. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chang, H.Y.; Chang, J.S. Producing carbohydrate-rich microalgal biomass grown under mixotrophic conditions as feedstock for biohydrogen production. Int. J. Hydrogen Energy 2016, 41, 4413–4420. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Arya, S.K. Hydrogen from algal biomass: A review of production process. Biotechnol. Rep. 2017, 15, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Müller, K.; Fleige, M.; Rachow, F.; Schmeißer, D. Sabatier based CO2-methanation of flue gas emitted by conventional power plants. Energy Procedia 2013, 40, 240–248. [Google Scholar] [CrossRef]
- Xia, A.; Cheng, J.; Song, W.; Su, H.; Ding, L.; Lin, R.; Lu, H.; Liu, J.; Zhou, J.; Cen, K. Fermentative hydrogen production using algal biomass as feedstock. Renew. Sustain. Energy Rev. 2015, 51, 209–230. [Google Scholar] [CrossRef]
- Alibardi, L.; Cossu, R. Effects of carbohydrate, protein and lipid content of organic waste on hydrogen production and fermentation products. Waste Manag. 2016, 47, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, J.B.; Felby, C.; Jørgensen, H. Yield-determining factors in high-solids enzymatic hydrolysis of lignocellulose. Biotechnol. Biofuels 2009, 2, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Biomass Concentration (g L−1) | Hmax (mL g-VS−1) | HPR (mL g-VS−1 h−1) | λ (h) |
|---|---|---|---|
| 10 | 132 ± 4 b | 11.1 ± 0.2 a | 2.7 ± 0.3 e |
| 20 | 153 ± 11 a | 11.2 ± 0.2 a | 5.8 ± 0.5 d |
| 30 | 108 ± 6 c | 11.8 ± 0.0 a | 7.5 ± 0.3 c |
| 40 | 87 ± 8 d | 9.3 ± 1.4 b | 9.5 ± 0.6 b |
| 50 | 68 ± 3 e | 9.2 ± 0.3 b | 11.0 ± 0.2 a |
| Process | HBu (g L−1) | HPr (g L−1) | HAc (g L−1) | HFo (g L−1) | HLa (g L−1) | Total VFAs (g L−1) |
|---|---|---|---|---|---|---|
| SSF | 2.4 ± 0.4 | 0.03 ± 0.00 | 2.3 ± 0.4 | 0.03 ± 0.02 | 1.2 ± 0.2 | 5.96 ± 1.02 |
| PSSF | 1.9 ± 0.3 | 0.07 ± 0.01 | 2.4 ± 0.3 | 0.11 ± 0.07 | 1.1 ± 0.1 | 5.58 ± 0.78 |
| Algae Strain | Fermentation Process | Pretreatment | Hydrogen Yield (mL g-biomass−1) | Energy Yield (kJ g-biomass−1) | Hydrogen Productivity (mL g-VS−1 h−1) | Reference | |
|---|---|---|---|---|---|---|---|
| Method | Conditions | ||||||
| C. vulgaris | SHF | Acid-ultrasonic | 0.79% (v/w) HCl, 49,600 kJ kg-DW−1, 36 min | 42.1 | 0.54 | n/a | [66] |
| C. vulgaris | SHF | Acid-thermal | 1% HCl, 92 °C, 47 min | 47.1 | 0.60 | n/a | [24] |
| C. vulgaris FSP-E | SHF | Enzymatic | Accellerase 1500, 45 °C, 3 days | 57.3 | 0.73 | n/a | [67] |
| C. vulgaris | SHF | Enzymatic | Onozuka R-10, Macerozyme R-10 | 124.9 | 1.59 | 1.1 | [17] |
| Chlorella sp. | SSF | No pretreatment | Cellic CTec2, Termamyl SC, and Dextrozyme GA, 35 °C | 153 | 1.94 | 1.6 | This study |
| PSSF | Pre-hydrolysis | Cellic CTec2, Termamyl SC, and Dextrozyme GA, 50 °C, 12 h | 154.8 | 1.97 | 2.4 | This study | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giang, T.T.; Lunprom, S.; Liao, Q.; Reungsang, A.; Salakkam, A. Enhancing Hydrogen Production from Chlorella sp. Biomass by Pre-Hydrolysis with Simultaneous Saccharification and Fermentation (PSSF). Energies 2019, 12, 908. https://doi.org/10.3390/en12050908
Giang TT, Lunprom S, Liao Q, Reungsang A, Salakkam A. Enhancing Hydrogen Production from Chlorella sp. Biomass by Pre-Hydrolysis with Simultaneous Saccharification and Fermentation (PSSF). Energies. 2019; 12(5):908. https://doi.org/10.3390/en12050908
Chicago/Turabian StyleGiang, Tran Thi, Siriporn Lunprom, Qiang Liao, Alissara Reungsang, and Apilak Salakkam. 2019. "Enhancing Hydrogen Production from Chlorella sp. Biomass by Pre-Hydrolysis with Simultaneous Saccharification and Fermentation (PSSF)" Energies 12, no. 5: 908. https://doi.org/10.3390/en12050908
APA StyleGiang, T. T., Lunprom, S., Liao, Q., Reungsang, A., & Salakkam, A. (2019). Enhancing Hydrogen Production from Chlorella sp. Biomass by Pre-Hydrolysis with Simultaneous Saccharification and Fermentation (PSSF). Energies, 12(5), 908. https://doi.org/10.3390/en12050908