Electro-Microbiology as a Promising Approach Towards Renewable Energy and Environmental Sustainability
by
 , ,
, ,
Jafar Ali
1,2 ,
,
Aaqib Sohail
3, 
Lei Wang
1,*,
Muhammad Rizwan Haider
2,4,
Shahi Mulk
1,2 and
Gang Pan
1,5,* 1
Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
2
Research Center for Environmental Material and Pollution Control Technology, University of Chinese Academy of Sciences, Beijing 100049, China
3
Research Group Biomarkers for Infectious Diseases, TWINCORE Centre for Clinical and Experimental Infection Research, 30625 Hannover, Germany
4
Key Laboratory of Environmental Biotechnology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
5
School of Animal, Rural and Environmental Sciences, Nottingham Trent University, Brackenhurst Campus, Southwell NG25 0QF, UK
*
Authors to whom correspondence should be addressed.
Energies 2018, 11(7), 1822; https://doi.org/10.3390/en11071822
Submission received: 22 May 2018
/
Revised: 7 July 2018
/
Accepted: 9 July 2018
/
Published: 12 July 2018
Abstract
:Microbial electrochemical technologies provide sustainable wastewater treatment and energy production. Despite significant improvements in the power output of microbial fuel cells (MFCs), this technology is still far from practical applications. Extracting electrical energy and harvesting valuable products by electroactive bacteria (EAB) in bioelectrochemical systems (BESs) has emerged as an innovative approach to address energy and environmental challenges. Thus, maximizing power output and resource recovery is highly desirable for sustainable systems. Insights into the electrode-microbe interactions may help to optimize the performance of BESs for envisioned applications, and further validation by bioelectrochemical techniques is a prerequisite to completely understand the electro-microbiology. This review summarizes various extracellular electron transfer mechanisms involved in BESs. The significant role of characterization techniques in the advancement of the electro-microbiology field is discussed. Finally, diverse applications of BESs, such as resource recovery, and contributions to the pursuit of a more sustainable society are also highlighted.
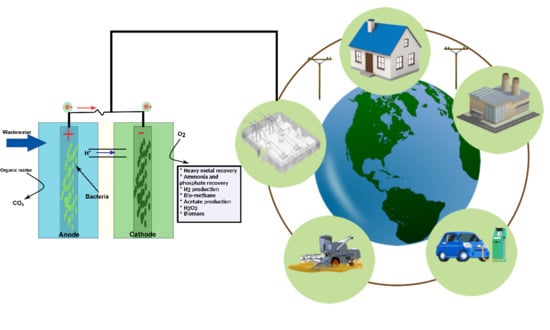
1. Introduction
Overexploitation of fossil fuels has resulted in scarcity of natural resources and environmental deterioration. Development of innovative technologies to explore sustainable and renewable energy resources is highly desirable. Recently, microorganisms have gained considerable attention for transforming organic waste into renewable energy and value-added products [1]. We live in a microbial world, where diverse and numerous species of microorganisms have adapted to almost all kinds of surroundings, ranging from moderate to extreme conditions and have developed diverse metabolisms to cope with their environment [2]. Electroactive (EA) and metal-reducing microbes are exciting examples with diverse metabolisms, which can transfer electrons to and from extracellular soluble/insoluble substances (electrodes) or use metal salts as terminal electron acceptors in the absence of molecular oxygen. Perhaps this unusual metabolism represents the primitive cellular nature during the process of evolution. Electro-microbiology is the study of the interactions between the electronic devices and novel electrical properties of microorganisms [3]. Electro-microbiology emerged as a multidisciplinary field during the early 1900s in experiments that revealed the potential of microbes to generate electricity by metabolizing organic carbon [4]. Later, the discovery of numerous dissimilatory metal reducing microbes revitalized research into electro-microbiology [5].
Key players in electro-microbiology are the so-called electroactive bacteria (EAB), which can interact with electrodes or some compounds in the vicinity via extracellular electron transfer (EET). EAB are used in various applications ranging from small power sources, pollution remediation, wastewater treatment, and metal transformation to electrosynthesis and energy recovery by bio-electrochemical systems (BESs) [1]. Interestingly, all BESs, such as microbial fuel cells (MFCs), microbial electrosynthesis (MES), microbial electrolysis cells (MECs), microbial desalination cells (MDCs), and microbial solar cells (MSCs), are governed by the electro-microbiology principles used for exploring the diverse potential of EAB. Generally, applications of BESs are determined by the reactor configuration, products required, and provided environmental conditions. Oxidation of organic substrates in the anode chamber and subsequent electron transfer to the anode are characteristics shared by all kinds of BESs [6]. These electrons are subsequently transferred via external circuit to the cathode where the ultimate consumption of electrons will determine the nature of the byproducts. In this regard, MFCs have emerged as promising alternative technologies for wastewater treatment (WWT) and concurrent generation of renewable energy [1,7]. Although considerable improvements in the power output of MFCs have been made during last decade, the process is not yet efficient enough for practical applications. There is a paradigm shift to explore the potential of electro-microbiological devices from electricity production alone to harvesting various fuels and valuable products in cathodic chambers.
Microbially-assisted chemical production in MECs offers a novel and sustainable way to cope with future challenges [8]. Therefore, maximizing power output, enhancing resource recovery, and creating useful products are highly desired for achieving sustainable systems. A better understanding of electro-active bacterial communities (EABc) and electron transfer strategies may help to improve the power density for envisioned application of BESs. The multidisciplinary nature of electro-microbiology will present additional challenges for reaching thorough understanding. Moreover, variations in results may appear because of inappropriate techniques and misinterpretations [9]. A combination of microbiological and electrochemical techniques can provide effective insights into complex interactions of microbe-electrode interfaces. Thus, collaborative interactions between various scientific disciplines will enable identification of the limitations of research methods to interpret complex data and determine its implications in electro-microbiology. Although several research articles and reviews have been published to evaluate the reactor configurations, electrode materials and operational parameters for BESs [1,3,10]. Unfortunately, understanding of electro-microbiological aspects of BESs, including detailed electron transfer pathways, important characterization methods, and recent trends in sustainability perspective, remain insufficient. Therefore, the current review aims to provide useful information for understanding the insights of EET mechanisms and how various electro-microbiological techniques have contributed to advancement of the field during the last decade.
Finally, recent applications of BESs for concurrent waste treatment and electricity generation along with various value-added bio-products are discussed. We anticipate that this study will expand knowledge about electro-microbiology and the critical role of electrochemical characterization techniques in sustainable remediation.
2. Microbe-Electrode Interaction
Electron movement is the essence of metabolic life for both eukaryotes and prokaryotes. Microbes extract energy through electron transfer from organic or inorganic substrates to the terminal electron acceptor, which can be oxygen, metal salts or electrodes [3]. It is assumed that EET at the microbe–electrode interface might have resulted from electron transfer strategies that originated with microbes respiring using insoluble minerals [Fe(III) oxides] under anoxic conditions over billions of years of evolution. Elucidation of electron transfer mechanisms at the microbe-electrode interface will reveal the evolutionary adaptations of EAB. However, microbe-electrode and microbe–minerals interactions are quite different because an electrode may provide a long-term surface for electron exchange. Microbe-electrode interaction is a basic feature of electro-microbiology and must be evaluated further as a function of current production. Recently, a study highlighted a strain-specific microbe-electrode interaction using pure cultures of Geobacter sulfurreducens and Shewanella oneidensis [11]. Microbial physiology can be altered in various conditions accordingly, such as in Fe(III) oxide-reducing Geobacter species, which may also develop flagella and become motile because they must find alternative sources of Fe(III). In contrast, Geobacter species that reside with the electrodes are not planktonic and provide a more stable thick electroactive biofilm. Thus, electrodes may serve as long-term electron acceptors for microbes involved in bioremediation of organic pollutants in anaerobic environments [12]. Additional studies of electrodes and analogous materials could enhance understanding of microbe-electrode interactions, which are anticipated to be useful for optimizing practical applications and coupling of biogeochemical reactions.
2.1. Mechanism of Electron Transfer
Electroactive biofilms are abundant in natural environments [2]. Various soluble electron shuttles can mediate the electron exchange at complex interfaces. Some organisms can also exchange electrons with electrodes or other microbes through artificial shuttles, such as natural red and methyl viologen.
These artificial shuttles accept the electrons from electroactive moieties in cells and subsequently transfer to the electrodes [13]. However, artificial electronic shuttles have drawbacks such as toxicity, lack of economic viability, and lack of applicability in an open environment. When soluble shuttles are not readily available for electron exchange, an electroactive biofilm will respire with minerals or electrodes [14]. Microorganisms can transfer electrons to the electrodes in two possible ways, which may follow direct or indirect pathways (Figure 1): (a) direct exchange can be short-range direct electron transfer between redox-active molecules on the outer cell surface and electrodes, or long-range direct electron transport through conductive pili, also referred as microbial nanowires. Direct electron exchange (DET) between an electroactive biofilm and the electrode appears to be most effective in producing the highest current density for a pure culture of Geobacter sulfurreducens, mediated by an extracellular c-type cytochrome [15]. (b) Indirect electron transfer occurs by microbially produced soluble redox-active molecules.
2.1.1. Direct Electron Transfer (DET)
Direct electron transfer (DET) is the most evident mechanism for EAB and can take place via conductive nanowires (pili) and membrane-bound cytochromes. DET has several advantages such as its ability to transfer electrons to the same or different bacterial species and independence of soluble redox mediators, and it may or may not involve cell-to-cell or cell-to-appendages contact with the electrodes or minerals. Typically, c-type OMCs (reductases) are involved in EET to and from electrodes for iron reducers such as G. sulferreducens. The role of c-type cytochromes (C-Cyts) in establishment of electrochemical communication with the electrodes has been documented mainly for G. sulfurreducens. However, C-Cyts may not assist EET for Fe(III) oxide reduction in Geobacter metallireducens. Cells in the vicinity or monolayers can easily transfer electrons and contribute to electricity generation, whereas a thick biofilm may face hindrance for EET. Thus, a highly conductive network of pili mediates the long-range transfer of electrons for electro-microbiological processes. Many bacterial strains are capable of producing pili, but electrically conductive pili are primarily associated with G. sulfurreducens [17]. Metal-like electrically conductive wires known as type IV pili have the ability to transfer electrons from cell to cell in a biofilm. Although the molecular mechanisms and structures of electrically active nanowires are not completely understood, it has been suggested that electron transfer is mediated by long aromatic–aromatic interaction chains, which result from stacking of conserved aromatic amino acids of GSu PilA105 [18].
The role of metallic conductive materials in biological EET compensations has been also evaluated for G. sulfurreducens. Another study provided the electrochemical and spectroscopic information on the bidirectional mechanism for microbe-electrode electron transfer in Geobacter soli biofilms [19]. Various researchers have contributed for the confirmation of the conductive role of nanowires, and some of these results are summarized in Table 1. Electrode-respiring Shewanella oneidensis MR-1 usually performs EET via the Mtr pathway, which further comprises five proteins: MtrA, CymA, MtrC, MtrB, and OmcA (Fig. 1A). Perhaps the earliest study to probe the DET mechanism in Shewanella oneidensis MR-1 showed that it was mediated by the OmcA–MtrCAB protein complex. In-vivo demonstration supported by cyclic voltammetry has ruled out the role of the CymA protein or secreted menaquinone in the DET process [20]. For short-range electron transfer across the outer membrane, various protein mediators (porin–protein complex) have been identified in G. sulfurreducens PCA. Generally, it is proposed that electron flow starts from the inner membrane protein (MacA) to periplasmic protein (PpcA) and then to the cell surface proteins (OMCs). Numerous genetic studies have revealed the involvement of other outer-membrane c-type cytochromes, OmcB, OmcS, and OmcZ, in electro-chemically active biofilms [21].
A prominent role of the OmcZ molecule in DET to the electrodes has been described previously. OmcZ consist of eight hemes and displays a wide range of redox potentials (−0.420 to −0.060 V). Therefore, OmcZ has been assigned as the principal c-type cytochrome contributing to the effective electrochemical communication between EAB and electrodes [22]. Remarkably, OMCs can store electrons (like a capacitor) when EET is not possible. Lovley [21] postulated that the conductivity of the electroactive biofilm of G. sulfurreducens is decisive for high current production. Electroactive biofilms perform as supercapacitors because of the presence of conductive cytochrome networks along with EET. Electrode modifications with graphene or reduced graphene for enhancing electrode-microbe interactions result in long-term EET and accelerating the oxygen reduction reaction (ORR) are still undebated. Direct interspecies electron transfer (DIET) is another newly discovered syntrophic metabolic pathway for cell-to-cell electron transfer between bacterial and methanogenic communities. DIET strategies have great biogeochemical significance and can stimulate the syntrophic conversion of various organic substances to methane in anaerobic digestion. Several conductive materials such as magnetite, graphite granules and carbon nanotubes have been tried to promote DIET kinetics between methanogens and volatile fatty acid (VFA) degraders in anaerobic digesters [23]. Generally, electrically conductive pili and outer surface cytochromes are considered essential electrical conduits for DIET. Primarily DIET-based mechanisms have been observed in defined co-cultures containing Geobacter species as one of the DIET partners.
However, DIET can take place under anoxic and aerobic conditions in the presence of nitrate, sulfate and oxygen and in the absence of CO2, thus permitting the deep cells of the biofilm to metabolize normally even when oxygen diffusion is limited [24]. Microbes with exceptional capability of DIET to serve as an electron source in anaerobic photosynthesis may further enhance their environmental applications in BESs. A recent investigation reported metatranscriptomic-based evidence for DIET between methanogenic rice paddy soils and Geobacter methanothrix species [25]. Further studies are needed to fully understand the electron flow, microbial physiology and biodiversity of microbes in DIET. Despite of the involvement of various electrochemical and microscopic techniques in EET research, thermodynamics and charge transfer resistance still need to be explored to optimize BES applications and minimize internal resistance.
2.1.2. Indirect Electron Transfer (IET)
Sometime electron transfer takes place without direct contact with solid-phase electron acceptors (Figure 1B). A number of microbes have been reported to produce electron shuttles such as flavins, pyocyanin, quinones, etc. to promote the electron transfer processes [26]. G. sulfurreducens secretes riboflavin in biofilm to interacts with C-type cytochromes (OM c-Cyts) of the outer membrane and mediate the electron transfer processes. Apparently, riboflavin production and binding in G. sulfurreducens biofilm is regulated with intracellular redox homeostasis [27]. Similarly, self-secreted electron shuttles have been linked with microbes that transfer the electrons from bacterial cells to Fe(III) oxides. An interesting example is reduction of Fe(III) enclosed in porous alginate beads by secreted electron shuttles from Geothrix fermentans, which can produce two different soluble redox-active shuttles with different redox potentials [28]. Shewanella sp. can also reduce Fe(III) oxides despite not being in direct contact through the release of flavin [27]. A P. aeruginosa strain KRP that produces pyocyanin and phenazine-1-carboxamide mediators has been isolated from MFCs. In another study, addition of sophorolipid was used to alter the cell membrane permeability for enhanced production of pyocyanin, eventually increasing the power output, whereas EET efficiency and power output were further improved when P. aeruginosa-phzM had an over-expressed methyltransferase encoding gene [29].
The role of flavin molecules in promoting the EET in S. oneidensis has been well documented. Flavin molecules can interact and become reduced with c-type cytochrome MtrC at the outer membrane of S. oneidensis. In addition, the S. oneidensis MR-1 flavin mediates EET more toward insoluble electron acceptors. Flavin adenine dinucleotide (FAD) molecules have also been identified in S. oneidensis MR-1 for exportation of flavin. [30]. Another investigation on the role of the FAD coding gene has demonstrated that a sharp decrease in flavin concentration led to low reduction in insoluble Fe(III), whereas overexpression enhanced EET and the reduction of insoluble electron acceptors [31]. Moreover, elegant studies endorsed the indirect electron transfer mechanism in current production, whereas direct contact was prevented between the electrode and S. oneidensis by a non-conducting mask with nanoholes. Flavin binds with outer membrane cytochrome molecules to form complexes, which can mediate EET strategies. Table 1 presents various studies that have used MtrCAB to investigate EET by mediators. To demonstrate the aforementioned role of flavin in a synthetic porin–cytochrome complex (MtrCAB) encapsulated in liposomes, Shewanella was exposed to various iron salts. Interestingly, current findings corroborated the previous results demonstrating EET enhancement by flavin–cytochrome complexes [32,33].
Okamoto, et al. [34] observed that Geobacter sp. also utilizes self-secreted flavin as a redox cofactor in the outer membrane c-type cytochrome MtrC. The effects of various artificially induced mediators on the enhancement of current production and remediation of pollutants such as azo dyes in BESs have been reported previously (Table 1).
Recently, fumarate was evaluated for promoting EET by inhibiting the synthesis of riboflavin in MFCs [33]. Along with EET to electrodes, mediators have also been used to study internal resistance based on the electrochemical impedance spectroscopy (EIS) technique [9]. Riboflavin-mediated EET processes are not limited only to EAB; yeasts such as Pachysolen tannophilus, Pichia stipitis, and Saccharomyces cerevisiae are also promising candidates in this regard. Kaneko, et al. [48] designed cyto-compatible amphiphilic redox-active polymers for efficient and improved EET in Saccharomyces cerevisiae. These significant findings not only provide a systematic understanding of EET but also highlighted the role of secreted riboflavin in intracellular redox homeostasis. Although mediator-based fuel cells are unsustainable for long-term operations, further investigations of fundamental aspects will facilitate BESs applications like telemetry circuities, underwater monitoring devices, bio-sensing, bioremediation and biocathode studies. Conclusively, electron transfer via soluble mediators is an important phenomenon even in established biofilms.
3. Role of Characterization Techniques in the Advancement of Electro-Microbiology
Although the study of electro-microbiology originated about a century ago [4], it took several decades to establish mediator-less MFCs. Significant design developments and enhancement in power production have been achieved during the last few years. Yet, there is still great potential for further improvements to make MFC technology more sustainable and cost effective. A better understanding of each component is essential to identify the bottlenecks in MFCs for improving efficiency. Selection of the most appropriate techniques to evaluate MFCs performance could be crucial [49]. Interaction between various technological disciplines, such as electrochemistry, microbiology, materials science, and environmental engineering, is needed to understand the multidisciplinary challenges of MFCs. Of course, many detailed review articles have discussed the history, construction, operational conditions, and limitations of MFCs. However, few publications have focused on the most appropriate electro-microbiological techniques required for MFCs diagnosis and for continued innovation. Microbiological practices combine with electrochemical principles to participate in the development of electro-microbiology field.
3.1. Fundamental and Culturing Techniques
Isolation of microorganisms is a conventional way to purify and characterize microbes. Pure cultures can provide detailed information on electro-microbiological samples and the current generation strategies employed by different microbes. However, it is always essential to check the purity of a culture at the end of the experiment, which may be performed using several molecular or microscopic techniques in combination with bacterial culturing methods. It is well documented that only 1% of microbes have been cultured, and researchers are still trying to recover more “unculturable” microbes. Isolation also provides a comprehensive depiction of physiological and biochemical pathways for pure cultures. Therefore, isolation of EAB from BESs is necessary to fully understand the insights of electro-microbiology [50]. Dilution methods, physical enhancement, and the most probable number (MPN) method are major strategies employed to isolate microbes. Recently, Heidrich, et al. [51] proposed the details of the MPN method for correlating the number of EAB capable of electricity generation in MFCs.
The dilution to extinction strategy is used to isolate the most abundant strains by pour plate, streak plate and spread plate. Isolation of various EAB by dilution methods have proven effective. Nevertheless, pre-enrichment of electroactive microbes is needed, which underscores the limitation of this method in MFCs. Non-essential flora may cause difficulties in isolation of the targeted population. Physical enhancement of bacterial cultures under specific conditions may improve isolation processes, such as isolation of dissimilatory metal-reducing bacteria (DMRBs), which can also provide indirect selection of EAB and subsequent production of electricity in MFCs [52]. Thus, purified and isolated cultures of EAB are necessary to clearly understand the microbial physiology within the BESs. Furthermore, it is convenient to study the effects of various parameters, such as substrate concentration, temperature and pH, on purified culture as compared with mixed consortia. Therefore, isolation in combination with other molecular techniques can be useful for exploring the diversity of EAB. We have summarized different EAB isolated from various BESs using a combination of culturing methods (Table 2).
3.2. Molecular Techniques for Characterization of Electroactive Bacteria
Purified isolated cultures of EAB must be further validated by molecular techniques using 16S rRNA-gene sequencing, which are considered the “gold standard” in identification and comparative study of microbial communities. In this regard, the use of operational taxonomic units (OTUs) will compare the community structures without assigning sequences into substantial taxonomic ranks. To date, more than 60 EAB have been isolated from various sources. Phylogenetic studies have revealed that electroactive communities mainly belong to five major classes of Proteobacteria, as well as to the phyla Firmicutes, Acidobacteria, and Bacteroidetes [66]. Presumably, a number of EAB will be discovered in the future [3]. However, population dynamics could serve as an important indicator for understanding the microbial profiles with physiological and electrochemical activities. For example, 16S rRNA sequencing analysis has shown the dominant microbial community and species diversity in BESs. Denaturing gradient gel electrophoresis (DGGE) is another powerful technique for evaluating the successive communities after different treatments and experimental conditions in MFCs. Recently, Baranitharan, et al. [67] reported the dynamics of a microbial community during MFCs operation using palm oil mill effluent through DGGE analysis. Thus, genomic sequencing and DGGE can be employed to explore the ecophysiology and metabolic interactions in BESs within various microbial communities under various treatments. Samples analyzed before and after the specific treatments from BESs will suggest how EAB are interacting for the current production and substrate utilization.
Fluorescence in situ hybridization (FISH) has been successfully utilized for phylogenetic study based on the 16S rRNA sequences for understanding the spatial interactions in complex microbial communities of BESs [68]. Apart from its advantage of high sensitivity, FISH is independent of DNA extraction and PCR amplification for microbial identification, which shows the simplicity and usefulness of this method. However, drawbacks may involve time consumption and the laborious nature of this technique. Interestingly, Parameswaran, et al. [69] have suggested a novel way to explore microbial communities through detecting 16S rRNA gene sequences and formyltetrahydrofolate synthetase (FTHFS) gene sequences using qPCR. Thus, rapid detection of genes in structurally different EABc is also possible. FISH analysis can be useful for differentiating two different types of biofilm compositions, established even under slightly different conditions such as surface charge and hydrophobicity of electrodes. Terminal restriction fragment length polymorphism (T-RFLP) along with 16S rRNA clone libraries have been used to investigate EAB community profiles under varying conditions. Construction of clone libraries for the determination of dominant species and further quantification of community components under various conditions have been employed by many researchers in BESs [70]. However, a clone library also requires PCR amplification, like other fingerprinting techniques (DGGE and T-RFLP). Although several molecular techniques have been practiced to investigate the EABc, it is desirable to employ some more reliable methods that may avoid too many steps involved in genome extraction and clone libraries.
In recent times, Ishii, et al. [71] have studied the population dynamics of EABc by clone libraries in MFCs. They have reported that microbial communities were highly diverse and that physiologically important EABc exist in widely natural ecosystems; however, the nature and source of the inoculum can influence performance. DNA microarray is also a powerful and unbiased technology, which analyzes the expression of thousands of genes at once. PhyloChip is a manufactured DNA microarray for high-precision microbial community assessment. DNA Microarray (G3) identifies all known bacteria and archaea along with minor communities; thus, PhyloChip is cost effective and relatively efficient. Paitier, et al. [72] reported the population dynamics of anodic biofilm dynamics during MFC start-up based on DNA microarray analysis. Stable isotope probing (SIP) can characterize metabolically active microbes within specific involved complex environments [73]. Combination of SIP and molecular biological techniques is potentially favorable for widening the scope of the microbial ecology of BESs. Moreover, the SIP method does not need any supporting data about enzymatic and genomic profiles. Thus, SIP can be employed to characterize novel EAB. Meta-transcriptomics is a high-throughput technique that can simultaneously recover community composition, functional activity and functional adaptability based on phylogenetic and metabolic markers.
A novel meta-transcriptomic approach was applied to identify gene expression dynamics during extracellular electron transfer [74]. Meta-transcriptomics results are more reliable for comparative studies than microarray and RT-PCR, which may be influenced by probe design, composition, array construction and primer and hybridization conditions. Despite the highly precise measurements, several limitations are associated with the meta-transcriptomics approach; for instance, the sequence may not have close matches in existing databases. Hence, several genes from a microbial community may not be correctly annotated. Moreover, meta-omics approaches are entirely dependent on previous knowledge based on pure cultures; thus, genetic and physiological studies must be carried out in mixed consortia of BESs to completely understand the ecophysiology. This technique may be expedient in the selection of inoculum for large-scale reactors and addressing practical issues along with providing cost effectiveness. Fatty acid methyl ester (FAME) analysis is a rapid and cost-effective method to describe the microbial community structures without traditional cultivation techniques that may undermine community diversity. However, interpretation of FAME profiles from various environmental samples can be difficult because of the presence of common fatty acids in different microorganisms. Therefore, additional support from other microbiological and molecular methods must be provided to ensure the precise identification of EABs. The above-mentioned molecular studies based on metabolites also explored the unexpected features of EAB and actively involved the advancement of electro-microbiology.
Microscopic imaging of biofilm is another standard method used for investigating the different aspects of a biofilm. The correlation between the current and the biofilm was later studied with the help of confocal laser scanning microscopy (CLSM) by another research group [75]. This study created the possibility to analyze various parameters of biofilm study, such as surface coverage, biofilm roughness and biomass along with electricity generation. Nevin, et al. [76] monitored the variations in biofilm structure development by G. sulfurreducens biofilms grown on an electrode fed with or without fumarate. Likewise, single-cell electron transfer rates can be studied under high-resolution microscopes [75]. It should be taken into account that the above-mentioned single cell electron transfer rates are very important in the research associated with electron transfer mechanisms by mutants. Notably, different growth rates compared with wild type counterparts are the basic ideas for present studies, which implies that electricity generation can be quantified on a per-cell basis by comparing the mutant strains with the wild type. Thus, comparative study of current production by various biofilms would help to minimize the effect of growth rates on experimental steps. Hence, microscopic investigations have an important role in unveiling the complex biofilm structures in different BESs, which may allow microbiologists and environmentalists to critically correlate biofilm structures with various functions, such as electron transfer rates and substrate utilization.
3.3. Electrochemical Characterization of Microbial Communities
Electrochemical characterization is the next step when microbial communities have been successfully established on electrode surfaces. However, adequate understanding of complex electrochemical methods is highly desirable for dissecting the insights of population dynamics in BESs. Otherwise, incorrect interpretation of data or even incomplete description of information may result in misunderstandings and confusion. Therefore, we are presenting some literature and discussion about electrochemical technique utilization from an electro-microbiological perspective. This collective information is proposed to overcome the challenges of the multidisciplinary nature of this work by integrating electrochemical characterization with power production.
3.3.1. Cyclic Voltammetry (CV)
Cyclic voltammetry is a versatile electroanalytical technique for investigating EAB species. Because of its versatility and ease of measurements, CV is extensively used in various fields of bio-electrochemistry and electro-microbiology. CV is often the primary technique used to analyze electrochemical experiments [1]. The effectiveness of CV can be estimated based on rapid detection of redox molecules over a wide potential range. CV is also often described as “spectroscopy for the electrochemist”. A voltammogram conveys information as a function of an energy scan [14]. Despite wide range effectiveness and diverse applications, CV is not well understood like other instrumental methods such as spectroscopy and chromatography. In this review, we describe a basic understanding and emphasize the kind of information that can be obtained by this complex technique. Several parameters can influence the information obtained in CV, such as the potential window, scan rate, and growth stage of biofilms. Pre-scanning of the biofilm at a slow scan rate will help to select the suitable potential range. Growth stages may vary according to the tests required [14,31]. An electron transfer mechanism involving oxidation or reduction reactions can be explored using a three-electrode configuration. When an electroactive biofilm is successfully established and a steady-state current is achieved, CV analysis can identify the electric potential at which redox-active couples may be oxidized or reduced.
For CV analysis, electric potential is scanned from the initial to final potential at a specific scan rate, and thus current is measured. CV can determine a particular range of potentials in which redox-active molecules can exist and electron transfer is possible for EAB [77]. Moreover, for controlled conditions, the mechanistic pathway of electron transfer in EAB can be elucidated; that is, either biofilm itself is involved in electron transfer, or surface-adsorbed molecules and freely diffusing species are responsible for this transfer [78]. A distinct role of OmcB and OmcZ in the extracellular electron transfer for Geobacter biofilms was studied by CV analysis. However, CV does not reveal any information about long-term current production by EAB. Mostly, long-term electrode polarization studies are coupled with CV to investigate how active systematic changes are influential for active redox coupled under controlled conditions such as pH. Rotating disk electrodes have solved the reproducibility issue relating to mass transfer conditions. Mutant studies with particular gene deletions of the EAB have provided additional strength to CV analysis, although reaction kinetics and mass transfer for EAB are similarly important to study [78]. A basic tutorial on investigating the EABc through CV analysis was presented by Frick et al [14]. They have acknowledged that detailed information can be derived from CV, such as the thermodynamics, mass transfer, nature of the mechanism involved in electron transfer, reversibility of redox species and its dependency on the scan rate, formal potentials of redox species, identification of electrochemically active sites and kinetic parameters.
Hence, CV can help to identify the mechanism of electron transfer and to select suitable electrode materials with less start-up time and high catalytic currents for enhanced performance of BESs. Moreover, the wide scan range of CV may reveal the presence of EAB in unknown samples for effective removal in MFCs. Integration of CV with bioinformatics tools and mathematical modeling is highly needed for collaborative interpretation and comprehensive understanding of microbe-electrode interactions in detail.
3.3.2. Electrochemical Impedance Spectroscopy (EIS)
EIS is a powerful and non-intrusive electrochemical technique that has been employed for corrosion studies during the past few years. Despite its extensive use in microbiologically influenced corrosion (MIC), it was not frequently exploited for evaluation of MFC performance until Manohar, et al. [79] published their work. Later, He and Mansfeld [80] provided comprehensive information on use of EIS for investigating the performance of MFCs based on internal resistances analysis of catalysts, biofilm development, and electrochemical reactions on electrodes. They postulated the EIS measurement procedure and basic considerations associated with data interpretations [80]. During the last five years, various studies involving the EIS in MFCs have been published. Interestingly, an improved MFC performance of microbes for reducing electron transfer resistance was demonstrated by the use of EIS [81]. In another study, the effect of anolyte and biofilm development was investigated on MFC performance by EIS. A significant reduction in electron transfer resistance and stable power generation in MFCs was attributed to electroactive biofilm by EIS after a long period of operation. Thus, EIS can work as an indicator system for biofilm establishment in a BES under various experimental conditions [82]. EIS can also provide quality-control-based catalyst fabrications by measuring the polarization and charge transfer resistance. In addition, the EIS technique can open the black box of the diffusion process and reveal the reaction kinetics. Because high internal resistance is a major drawback in the large-scale application of BESs, EIS has improved the depiction of research gaps in this area.
EIS has been shown to be more efficient and reliable than conventional current interrupt methods of observing internal resistance. Thus, EIS has gained a great respect for its use in characterization of new catalytic materials needed in electrode modification for achieving high power densities in MFCs. Furthermore, microbes and metabolites can act as biocatalysts for improving kinetically controlled electrochemical reactions and lowering internal resistance; thus, BESs combined with EIS techniques may also be used in biosensor applications.
3.3.3. Square Wave Voltammetry (SWV)
SWV is a reliable electrochemical technique used to detect low concentrations of electrochemically active species that are not easily detectable by CV [14]. Suppression of background currents and highly sensitive screening enable SWV in numerous electrochemical measurements. We have aimed to summarize how differently SWV can be used to infer bio-electrochemical behaviors. In one study, overpotential approximation of electroactive biofilm impedance was achieved for Geobacter sulfurreducens [83]. Moreover, SWV reflects the rapid analytical detection of reversibility of electrochemical reactions. Recently, an electroactive biofilm capable of simultaneously consuming substrate (electrotrophy) and producing electricity was studied by SWV. To promote electricity production by Pseudomonas sp. biofilm, phenazine-induced redox reaction was examined by SWV [84]. The ratio of the magnitudes of the peak currents in the forward and reverse directions can be utilized to estimate electrochemical reversibility. In addition to this, SWV has been coupled with other analytical techniques like high-pressure liquid chromatography and thin-layer chromatography (TLC) for electrochemical measurements of various pollutants. The high sensitivity of SWV enables the detection of very low (pico-molar) concentrations of electroactive analytes in solutions, which is a direct indication for studying EAB even after a few hours of incubation. However, to determine the electron transfer rate and electrode kinetics, SWV must be combined with simulation-based approaches.
3.3.4. Chronoamperometry (CA)
CA is an electrochemical technique where the potential of a working electrode is stepped and the resulting current is observed as a function of time. However, limited information can be extracted from the ratio between anodic and cathodic peaks. A better noise-to-peak ratio is obtained with a relatively high current integration time interval. Cercado-Quezada, et al. [85] studied various food-industry wastes for electricity production, and microbial biofilm was evaluated using the CA technique. Possibly the first demonstration of effective biofilm formation on stainless steel electrodes in marine MFCs was supported by the CA results. A continuous rise in catalytic current was revealed by cathode. These findings were incorporated with previous findings of biofilm-driven catalysis of oxygen reduction examined through CA experiments. Fricke, et al. [86] coupled CA with CV to explore the underlying mechanism involved in anodic electron transfer from biofilm. A high-performance rotating graphite fiber brush air-cathode was designed for enhanced ORR and power production by MFCs, and characterization was supported by CA [87].
A recent study has used carbon materials derived from waste tiers as high-performance anodes in microbial fuel cells. The anode was developed using tire waste material, and electrochemical characterizations were performed by CV and CA to optimize the design of carbonized waste. In this MFC, environmental problems caused by waste tires were alleviated [88]. In another study, a tungsten carbide-based anode was fed with glucose and inoculated with heat-treated soil, and microbial growth and oxidation of metabolites were then recorded as a function of time [89]. Thus, CA can be a reliable technique to understand the bottlenecks and limitations of electro-microbiology regarding the role of electrode material modifications for improving the overall performance of BESs. Moreover, detailed insights of biofilm formation on various electrode materials in different orientations can be obtained, which may help to fully optimize the growth parameters for specific current density.
3.3.5. Differential Pulse Voltammetry (DPV)
Compared with other voltammetry techniques, DPV is a highly precise electroanalytical technique used to measure the cell current as a function of time and potential between the working and reference electrodes. The high sensitivity of DPV can be attributed to extraction of faradic current from capacitive current. Consequently, much more reliable information can be obtained from redox-active molecules than can be using the CV and LSV techniques. However, in DPV analysis, potential is varied using pulses of rising amplitude, and the current is sampled before and after each voltage pulse. Interestingly, DPV is a preferred tool for exploring the electron transfer mechanism in EAB in MFCs and other electrochemical systems. In DPV measurement, a carbon electrode was colonized by Shewanella in a sterile medium. Marsili, Rollefson, Baron, Hozalski and Bond [77] observed that an adsorbed flavin layer on an electrode functioned as an electron acceptor in the presence of secondary metabolites in electrolyte solutions. Recent investigations on polyaniline-hybridized mesoporous carbon anodes using DPV have enabled fast electron transfer via inner endogenous electron mediators and outer membrane redox centers [90]. Combination of DPV and meta-omic approaches will open new horizons for studying electroactive species under different conditions. In general, DPV has significantly contributed to the field of electro-microbiology by elucidating the reaction kinetics of electron transfer processes.
4. Electro-Microbiology and Environmental Sustainability
4.1. Simultaneous Electricity Generation & Wastewater Treatment by MFCs
Electricity production is an exceptional feature of MFCs, wherein EAB convert chemical energy into electrical energy. Bioelectricity generation is not a new idea [4], but it is now believed that most of the organic substances can be converted into electricity [6]. MFC technology offers several advantages over other renewable and competitive technologies, such as (a) high conversion efficiency (90%) because of the direct conversion of substrate chemical energy to electricity, (b) efficient operation at ambient temperature, (c) safe and quiet performance, (d) electricity obtained from MFCs is sustainable and not limited by the Carnot cycle and heat loss as in chemical fuel cells. A highest coulombic efficiency of about 97% has been reported from formate oxidation in the presence of Pt-black [91]. Some very attractive power densities from MFCs are presented in Table 3. Although bioelectricity generation is the primary outcome of MFC technology, wastewater treatment is also considered an important application. All conventional biological treatment practices for low-concentration wastewater treatment are expensive and energy-consuming. Thus, cost-effective and sustainable treatment technologies are in high demanded. BESs can achieve wastewater treatment and electricity generation along with valuable resource recovery [92].
Activated sludge process consumes about 0.3 kWhm−3 of energy in aeration and additional pumping power during domestic wastewater treatment. Anaerobic conversion of organic waste to methane can recover only a small portion of the energy, while most of the organic fraction of waste is lost in aerobic oxidation processes. Therefore, anaerobic digestion can replace aerobic treatment processes, but still requires very complex working conditions such as specific temperature and a concentrated waste stream. Interestingly, wastewater treatment by BESs will result in no sludge, and no additional sludge management or treatment strategies are required. Despite these advantages, fuel cell technologies still face challenges that limit their practical applications. Low power densities and the high costs of materials are the major drawbacks in this regard. High overpotentials at electrodes include activation losses, ohmic losses, and mass transport loses. Overpotentials mainly contribute toward high internal resistance of reactors. The high cost of cathodic catalysts for oxygen reduction reaction (ORR) and fouling of proton exchange membranes are considered the most effective areas to improve the power density and lower costs [93]. The above-mentioned factors must be taken into account before the commercialization of MFCs technology. Despite low power densities, organic waste removal is much more sustainable in MFCs from an economic perspective. In this context, wastewaters can serve as a resource for energy and valuable chemicals via in-situ processes in MEC/MESs.
Hydrogen production along with H2O2 from wastewater in MECs is energetically more favorable than conventional electrolysis methods with further production of valuable products. The chemical oxygen demand (COD) removal efficiency is considered one of the critical parameter to assess the performance of MFCs. A comparative study based on COD removal and energy recovery revealed that MFCs have better performance than MECs [94]. Organic waste is considered as best nutrient media for EAB in MFCs. However, wastewater types range from paper industry wastewater, chocolate industry wastewater, human feces wastewater, fermented vegetable waste, brewery wastewater, manure sludge, domestic wastewater, landfill leachate, and other food processing industrial wastewaters [6]. Recently, BESs have been successfully applied for the treatment of emerging pollutants, anti-inflammatory drugs, estrogens and antibiotics with high mineralization rates. The removal of such pollutants is more problematic than that of organic wastes. Antibiotics have been frequently detected in the effluents of wastewater treatment plants and receiving water bodies, which shows the inability of conventional treatment systems to metabolize the trace levels of such contaminants. Long-term exposure to these contaminants could lead to the evolution of antibiotic-resistant bacteria and genes, which pose serious risks for human health [95]. Therefore, effective treatment is needed for the removal of recalcitrant compounds and antibiotics.
For example, in a study, about 85% of sulphamethoxazol (20 ppm) was degraded within 12 h with further mineralization of degradation products as well [96]. Antibiotics reportedly increased the power output of MFCs when used as electron donors along with glucose or acetate [97] and improved the stability of electricity production by inhibiting bacterial growth in the cathode chamber [98]. Another promising application of BES-driven advanced oxidation processes is in-situ production of H2O2 for Fenton degradation of emerging contaminants [99]. Therefore, BESs may be an interesting option for degradation of pollutants along with energy recovery; however, the generation and release of antibiotic-resistant bacteria and genes should not be overlooked. Numerous types of wastes have been tried as substrates in MFCs and have provided comprehensive and state-of-the-art information on various wastewater substrates [100]. An overview of COD removal efficiencies and power densities was provided in Table 3.
4.2. Resource Recovery and Sustainability
Nutrients recovery from wastewater is a more sustainable approach than nutrient removal alone that can minimize the environmental footprint of wastewater treatment. The recovered nutrients such as ammonia and phosphorus can be further utilized for fertilizer production, thereby BESs enables the enhanced energy recovery and reduce the serious environmental impacts of wastewater treatment processes. In addition, BESs have been developed to recover various useful nutrients and heavy metals along with generation of value-added compounds such as methane, acetate, hydrogen, hydrogen peroxide, and biomass. Herein, we provide an overview of how electro-microbiology is contributing in the pursuit of sustainable developments to address the energy and environmental challenges.
4.2.1. Metal Recovery
Wastewaters contain a wide range of metals such as copper, lead, iron, zinc and manganese. Their concentrations are usually low, for example, 1mg/L and depend upon the type of wastewater. Initially, the main focus was the waste treatment and electricity production, but now the emphasis has been shifted towards bioremediation and bioproduction processes in MECs. It is proposed that compounds with high redox potentials can serve as an effective electron acceptors in the cathode of MFCs. Electrons produced from anodic oxidation are further used to remove the persistent pollutants and heavy metals. When contaminants are electrons acceptors, the plating-out phenomena of metals will take place. However, metals with low redox potentials (e.g., Zn, Ni, Pb) can also be recovered using biocathode and external power supply. Numerous heavy metals including silver, arsenic, copper and cobalt have been treated and recovered in anodic and cathodic chambers of BESs. Earlier, BESs have been applied for the metal removal and recovery from the leachate/effluents of metallurgical processes which successfully resulted in metals reuse. Toxic metals such as chromium (VI) and vanadium can be reduced to their less toxic forms along with power production. A recent study reported the electrochemical reduction of copper at the cathode in the presence of bipolar membrane where acetate was oxidized. Reduced copper was plated on graphite electrode with the current density of 3.2 mA−2 in BESs [131].
Although metal recovery from landfill leachates presents a big challenge, the economic feasibility of the process still needs more investigation. Different factors such as the age of leachates, pH and organic content may influence the recovering metals. So, their complex interactions with methanogenic phases and acidogenic phases must be well understood for enhanced metal recovery. Another possible way to improve the metal recovery processes is the use of ion exchange membranes for anolyte before introduction to the cathode. Moreover, several other studies have described the precipitation and recovery of pure metals [132]. Metal recovery and current generation indeed reduce the energy needed for treatment processes. Thus, BESs have shown great potential for efficient removal of toxic heavy metals with simultaneous recovery of precious metals even in very low concentrations, which is not possible by using any other process. In addition to the cost-effectiveness, it also decreases the environmental and health risks associated with toxic heavy metals and thus increase sustainability.
4.2.2. Ammonia and Phosphorus Recovery
In addition to the metals, nutrient recovery has also been the focus of concerns over nutrient removal and waste treatment. Nitrogen in the form of ammonia and phosphorous as orthophosphate are the main species of wastewater or leachate. Agricultural wastes are also enriched with nitrogen and phosphorous due to the extensive use of synthetic fertilizers to boost the crop yield. Intensive mining to supplement the worldwide demand has led to a speculation that current phosphorous deposits may not last until the end of 21st century. Thus, alternate and renewable methods (BESs, MECs) are needed for sustainable supply of nutrients. The removal of high concentrated ammonia is the real issue for leachate treatment. Conventional strategies for ammonia removal include biological nitrogen removal (BNR) and ammonia stripping. Moreover, they need intensive biological systems and also energy/chemical consuming processes. Ionic solutions can be directed under the internal electric field of fuel cells. Therefore, BESs takes advantage of ion transport in the electrical field and enable the concurrent recovery of phosphorus and nitrogen, which is inconvenient in conventional water treatment processes. Recently, new practices were employed in BESs for ammonia oxidation at cathode along with denitrification at anode [133]. The ammonium (NH4+) recovery from urine has been achieved recently. Electrons required for NH4+ migration are provided by anodic oxidation, whereas at the cathode diffusion of ammonia (NH3) promotes the total ammonium. Localized pH facilitates the NH3 stripping, hence further improving the transport of ammonium.
The stripped ammonia can be absorbed in dilute sulfuric acid to form ammonium sulphate, which has the potential applications in industry for synthesis of valuable nitrogen polymers, such as fertilizer in agriculture and food production. Pure ammonium sulphate crystals can also be obtained by pre-saturated ammonium sulphate for stripped ammonia absorption, which is a valuable salt and can be applied in forward osmosis (FO). Ammonia recovery in BESs can greatly affect the energy performance (production and consumption) of the system. However, phosphorous recovery has to rely on precipitation in the cathode under high pH conditions. Recently, some research groups have revealed phosphorus recovery in MFCs. The phosphorous removal could reach up to 80% in an air cathode MFC [134]. Simultaneous recovery of phosphorus and nitrogen with electricity generation will be an exciting choice. Magnesium ammonium phosphorus (MAP) or striate can also form under high pH conditions, which may also help to recover phosphorus from high strength wastewater [135]. But phosphorus recovery from leachates via BESs is not well explored and issues exist regarding the effect of precipitation on electrode performance for reuse, and stoichiometry of struvite formation under various conditions. The recovered nutrients in BESs particularly phosphorus and nitrogen can be applied in agricultural lands as an alternate to synthetic fertilizers. Hence, nutrient recovery may help to cope with the high demand of phosphorous fertilizers. Taken together, nutrient recovery by BESs is an extremely attractive option and gives additional benefits than just sustainable treatment of leachate and wastewater. However, optimization of the innovative reactor models will be required to commercialize this possibility.
4.2.3. Biosynthesis Prospects of Electro-Microbiology
To cope with the challenges of the twenty-first century, with researchers looking for alternative renewable energy sources, the electro-microbiology is significantly contributing to the production of biofuels and renewable resources. Development of innovative technologies for concurrent enhancement in power output and their transfer in biosynthetic pathways for production of valuable compounds is highly needed. Electro-microbiological systems such as BESs can considerably meet these challenges by producing various fuels and chemicals. Moreover, microbial assisted chemical production via MECs provides a highly attractive and novel route for producing valuable products from wastewater along with electricity generation.
Biomethane
Methane gas can be produced by BESs rather than fossil fuels such as gasoline/petrol and diesel. Methane contributes as a major component of liquefied natural gas (LNG) and liquid methane rocket fuel. Bio-methane production from organic wastes through MECs is preferred over other conventional processes because of the potential of electro-active bacteria to work under normal conditions, without aeration, with less sludge production, and the high yield of methane, which make the processes more sustainable and cost effective as compared to other processes [136]. Various experimental studies have revealed the electricity and methane production from glucose, acetate, beer wastewater, paddy soil and alkaline pre-treated sludge. Initially, it was considered that methane production will affect the EABc in anode section while, Satar, et al. [137] reported the immobilized mixed-culture reactor for simultaneous production of hydrogen and methane from glucose. Thereby, suggesting the high efficiency of BESs for biomethane production. Methane production from CO2 and hydrogen resolves the environmental issue and encourage the recently developed area of carbon capture and utilization for reducing greenhouse gas. Efforts for maximizing the biomethane production from CO2 and extracting all benefits of its use must be promoted against chemical methods. Effective utilization of electricity in BESs to produce H2 and a suitable source of CO2 will certainly make this process economically viable.
Another study investigated the integrated BESs-AD system for relatively high methane production than AD under poised cathode potential of 0.90 V (vs. Ag/AgCl) [138]. Bio-electrochemical devices may reduce the CO2 into methane under the specific poised potential. Numerous in-situ methods such as CO2 desorption, H2 addition, pressurized reactor, and electro-methanogenesis are employed to improve quality of biomethane in anaerobic digestion. Therefore, biocathode executing electro-methanogenesis are considered as a promising way to convert the effluents of dark fermentation into methane [139]. The integration of BESs with anaerobic digesters will substantially reduce the cost of biogas cleaning and upgrading. In contrast, a recent study reported that methane was used as a source for direct conversion into electricity by reversing the methanogenesis with the help of synthetic consortia in an MFCs [140]. Authors have claimed that this strategy will help to reduce leaks (from distribution, transportation and storage) and capital cost. Conclusively, in order to develop an efficient and sustainable system for high methane production, selection of optimum strains of EAB from MECs can play a crucial role. Furthermore, optimization of other factors such as reactor design type, electrode design, materials and microbial communities will also be influential in methane production. However, most of the results obtained so far are based on lab scale studies, more efforts projecting current knowledge about biomethane production at large scales are required. It is believed that efficient methane production strategies without transportation and substantial leaks will be achieved in future to improve the cost effectiveness and sustainability.
Biohydrogen
Hydrogen gas is extensively used as a starting fuel in different industrial processes. Hydrogen is also considered as an attractive source of renewable energy. There is high demand for hydrogen production due to enormous applications in industry and its absence in nature. Initially, bulk hydrogen was obtained from non-renewable sources such as coal, natural gas, naphthalene distillates and sometimes by electrical methods [32]. Therefore, alternative and eco-friendly sources are necessary for hydrogen production. In this regard, Kreysa and Håkansson [141] have made the first attempt by suggesting the combination of an MFC and water electrolysis. Principally, bacteria oxidize organic compounds and generate electrons, protons and CO2. The electrons are transferred to an anode, which further aids with external voltage and reacts with protons at the cathode to form hydrogen. Hydrogen (H2) production and WWT in MECs can offer new methods of resource recovery. Hydrogen generation from organic wastes through MFCs is termed as electro-hydrogenesis [142], and considered a favorable, eco-friendly and economical source. However, low yields and thermodynamic limitations in microbial metabolism are some bottlenecks in fermentation-based processes. Several strategies have been applied to increase the hydrogen yield from dark fermentation, such as pretreatment of substrates, metabolic engineering, reducing hydrogen partial pressure and optimization of parameters. Recently, coupling the dark fermentation with bioelectrochemical technologies (BESs, MECs) to improve the hydrogen production has attracted much attention [143]. The MESs offers a promising and alternative technology to enhance hydrogen yield by utilization of residual liquid from dark fermentation and devoided of high-energy inputs. The high conversion efficiency [26,144] and diverse substrates as compared to conventional electrolysis of water are the additional advantages of MESs.
Integration of MECs with dark fermenters and subsequent utilization of DF-effluents (VFA) in anode may disturb particular pH range, thereby demonstrating the governing role of pH for anode respiring biofilm. Adjustment of pH window by buffering solutions will be challenging on large-scale applications. Therefore, there is need to develop some acid tolerant EAB for continuous and enhanced one step production of hydrogen. Effect of various biotic and abiotic parameters such as nature of influent, type of microbial community, reactor design and electrode materials still needs further investigation for predicting the ultimate hydrogen generation capacity. Theoretically, hydrogen formation at the cathode must overcome the endothermic barrier of −0.414 V vs. SHE based on an external power supply (0.14 V) to the MEC, and the remaining voltage (0.279 V) is provided by EAB via anodic oxidation of waste. However, carbon cathodes give slow hydrogen evolution reactions, which can be improved by metal catalysts, such as titanium (Ti), platinum (Pt), and nickel (Ni) utilized to accomplish higher production of hydrogen. Nowadays biocathode has emerged as an alternative to abiotic cathodes, which is eco-friendly and cost effective catalyst and not only regenerative but also sustainable for efficient electro-hydrogenesis in MECs. Recently, Jafary, et al. [145] have presented a comparative study on the development of biocathode for hydrogen production in MECs. Authors have described the effective enrichment technique for biocathode development from sulfate reducing microbes to improve the hydrogen production rate. However, more studies are needed to explore the mixed and pure consortia to enhance hydrogen generation and restrict the methane production in MECs. Although biohydrogen production by BESs-related technology is still in infancy, it is considered as most eco-friendly and sustainable methods to generate biofuels. Researchers must focus on efficient materials and optimal catalysts for up scaling and improving the H2 production in MECs to sustainably meet the future energy challenges.
Acetate
Electro-microbiology-based reactions have taken advantage of converting CO2 to multi-carbon compounds like acetate and other liquid fuels. Alternatively, energy is recovered and stored from wastewater treatment via C–C bonds. Moreover, acetate can serves as an intermediate molecule for the production of other biochemical compounds. The production rate of acetate was not very high in the start, but it was improved in later studies. An experimental setup produced approximately 89.9% acetic acid, more than conventionally achieved by aerated fermentation (75.8%). Xiao, et al. [146] suggested the first microbial electro-synthesis of acetate and ox butyrate by the acetogen Sporomusa ovata, which can utilize electrons derived from a graphite cathode to reduce the CO2. Since then, wide ranges of microbes have been exploited for CO2 reduction to acetate in MECs. These studies depicted the high electron recovery in MEC-mediated CO2 reduction by numerous acetogenic bacteria including Clostridium ljungdahlii, Moorella thermoacetica, and Sporomusa silvacetica. Nowadays, biocathode driven acetate producing MECs are attracting attention of researchers. An earlier investigation showed the microbial electro-synthesis of acetate via biocathode driven CO2 conversion process, which is a sustainable and stable approach for liquid fuel generation [147].
Long-term operation of biocathode MECs in semi batch mood indicated the possibility of commercializing the microbial electro synthesis pathways. The high acetate production in MECs can be supported by biologically induced hydrogen at cathode. Stable colonization of microbes and suitable electron acceptors to allow optimal metabolic activity at the biocathode are key parameters in acetate production. An applied potential of −280 mV vs. SHE must be provided to overcome the thermodynamic barrier of a biological reaction for effective reduction in MEC [148]. However, practically much lower potential can overcome this overpotential due to microbial energy uptake and mass and charge transfer resistances involved in bioelectrochemical devices [149]. Although microbial electrosynthesis is a promising way to capture CO2 and produce multi-carbon compounds, which are intermittent energy source and precursor of complex compounds, there is still need to optimize the acetogenic communities for maximum acetate production and to prevent the side reactions (methanogenesis) in MECs. A selective enrichment was used to establish a stable community for continuous conversion of CO2 to acetate. This will provide an instant start-up for electrosynthesis processes and reduce the thermodynamic loses. Apart from the aforementioned parameters, several other underlying factors, such as mediators in electron transfer and the mechanisms of electron transfer in cathode and electrode materials with specific reactor designs, can influence the overall efficiency. Currently, MEC technology is in its infancy; future studies must be focused on exploring novel aspects in the electro-microbiology domain.
Hydrogen Peroxide
Hydrogen peroxide (H2O2) is a valuable industrial chemical widely used in the water and wastewater industries for advanced oxidation of emerging contaminants and complex organic chemicals. Currently, 95% of H2O2 is commercially produced by anthraquinone oxidation, which is a toxic and energy-consuming process. Electro-microbiology based technologies can provide a cheap and easy way for H2O2 production at a cathode [150]. Several studies have contributed to the sustainable production of H2O2 from various substrates in BESs. Likewise, sulfate reducing microbes can degrade chlorinated pollutant in MECs with simultaneous power generation and H2O2 production. It implies that MECs can provide a sustainable source for simultaneous waste treatment and H2O2 generation, which could be further utilized in advanced oxidation processes for degradation of pharmaceutical wastewater. Primarily, two electron pathway for oxygen reduction reaction (ORR) is directly linked with peroxide production. Slow kinetics of oxygen reduction reaction (ORR) at the cathode and high cost of catalytic materials are the major drawbacks in the large scale applications of MFCs. Moreover, inorganic ORR catalysts suffer from high overpotentials, which is mainly attributed to the complex nature of ORR mechanism [151]. Some enzymatic catalysts such as laccases and bilirubin oxidases have shown much lower overpotentials at neutral pH.
Unfortunately, high cost, low durability and ineffectiveness in real wastewater hinders their scalability. Therefore some cheap, eco-friendly and cost effective catalysts must be found to enhance the performance of MFCs and production of H2O2. Electrode modifications and pretreatments with the acidic solution can introduce some important functional groups such as nitrogen which will help to increase the production of H2O2 and power densities [152]. Possibly, functional groups will also decrease the startup time, thereby maximizing the efficiency of MECs and MFCs. It was speculated that two electron pathway for oxygen reduction can be controlled by using spectrographically pure graphite (SPG) electrode in wastewater treatment [153]. Asghar, et al. [154] proposed the similar setup for H2O2 production and power generation in MFCs using nitrogen doped graphite cathode. Undoubtedly, carbon based cathodes modified with various metal nanocomposites have been extensively adopted as cathode materials due to the high active surface area and low cost, but still there is a need for further improvements. To explore the underlying biotic processes involved in the H2O2 production, impact of various operational parameters on the performance of microbial peroxide producing cells must be investigated in future. Generally, more research is needed to maximize the production of H2O2 and power generation from wastewater treatment processes. Slow reaction kinetics at the cathode is limiting factors for enhanced production of H2O2. Therefore, it is suggested to develop efficient cathodic catalysts to maximize the H2O2 production by two electrons based pathways. The bottlenecks associated with sluggish reaction kinetics and other limiting factors must be identified and resolved in a sustainable way to enhance the peroxide production. Moreover, integration of in-situ electro Fenton based processes with MECs and photo electrochemical systems may provide a new gateway for enhanced remediation in different environments.
Biomass
Microalgae display high efficiency in nutrient removal relative to other microorganisms due to presence of numerous essential nutrients (ammonia, nitrate, phosphate, urea and trace elements) required for micro algal growth in various wastewaters [155]. Thus, inexhaustible availability of CO2 could be employed for algae biomass production with simultaneous energy recovery [156,157]. Microalgae have been suggested as a promising substitute to conventional fossil fuels that could provide an alternate eco-friendly sustainable energy source. Renewable energy researchers have since shifted their focus toward photosynthetic MFC technology, which is evident from published studies. Although plant based MFCs have been also practiced for concurrent biomass and bioelectricity production, but slow growth and low biomass limits their large-scale applications. Recently, Commault, et al. [158] have demonstrated the electricity and biomass production in a bacteria-Chlorella based MFC by treating wastewater. Microalgae can produce unlimited oxygen at cathode by utilizing the atmospheric CO2 and generating the biomass in photosynthetic MFCs. Along with providing a sustainable electron acceptor, microalgae can interact with biocathodic communities to enhance the catalytic reacations. Different growth parameters affecting the algal biomass growth such as light duration and aeration time in photosynthetic-MFCs has been thoroughly reviewed in the literature [159]. Algal cultivation can effectively reduce the antibiotic load in pharmaceutical wastewater, which may also help to cope with antibiotic resistance due to the intrinsic eukaryotic nature. Algal biomass can further be converted into various biofuel depending upon the type of contents. Algal species rich in fatty acid content can be utilized for biodiesel production, whereas carbohydrate rich algal biomass can be used for biohydrogen, bioethanol, and biogas via microbial conversions.
Furthermore, after lipid extraction the biomass feedstock can be converted into algae biochar by conventional pyrolysis [160], torrefaction or hydrothermal carbonization. Algae biochar represent a new class of efficient fertilizers and absorbent materials for toxicants [161]. A combination with other sorbent material i.e., chitosan can enhance the application of bio char for treating the sewage wastewater prior to irrigate the agricultural fields. Thus providing a sustainable solution to environmental pollution. Though enhancement of biomass is considered as an unexplored area. However, photosynthetic MFCs are utilizing microalgae at cathode to provide oxygen, which is a free electron acceptor and biomass. Some groups are targeting the anolyte to grow algae in the cathode chamber, therefore providing the maximum sequestration of waste carbon and converting into algae biomass for further applications. It is apparent from the above discussion that integration of wastewater treatment with microalgae farming for biomass generation is an effective and efficient approach to biofuel generation. Despite the potential benefits, algae-bacterial MFCs have some limitations and barriers to successful commercialization in terms of biomass and wastewater treatment. However, cost-effective cultivation and harvesting constitute a major challenge. In this regard, appropriate selection of micro-algae species for specific wastewaters and pre-treatment of wastewater can improve and enhance biomass cultivation to create a more sustainable society.
5. Future Perspectives
This review summarizes various electron transfer strategies at the electrode-microbe interface, and electrochemical characterization techniques for understanding the insights of electro-microbiology and BES applications in the pursuit of a more sustainable society. Energy shortage and environmental deterioration have become severe global issues. Fossil fuels is a non-renewable source of energy because of limited reserves and have harmful environmental impacts. Therefore, development of new technologies for achieving alternative and renewable energy resources is in high demand. Electro-microbiology holds promise for converting waste into green and renewable resources. Despite low power densities, MFCs remain a promising method of organic waste removal and are much more sustainable than conventional processes from an economic perspective. Thus, MFCs can power the underwater monitoring devices and remote sensors, which are usually powered by batteries. Limited lifetime and regular replacement for recharging are the major disadvantages of battery-based devices. To ensure continuous power and to avoid operational inconveniences, MFCs can serve as sustainable power sources for wireless data transmitters and telemetry systems using the organic content of the environment. Several other applications in medical devices, robotics, and biosensors have been demonstrated successfully. The interaction between researchers with different backgrounds is needed to fully explore the new gateways in waste to energy challenges. We have described the insights of various electron transfer strategies in BESs. Combinations of electrochemical and microbiological techniques have elucidated the complex physiology within BESs. Although the molecular techniques and electrochemical methodologies needed for electro-microbiology study are still in their infancy, it is anticipated that reliable and rapid techniques will be discovered in future to advance this innovative field. Selecting an appropriate method and thorough understanding of electro-microbiology may help to optimize the efficiency of MFCs and other BESs for enhanced waste treatment along with energy production. Moreover, this review has also focused on how electro-microbiological systems (BESs) are contributing to sustainability and resource recovery. The paradigm shift from wastewater treatment alone to electricity generation and now toward resource recovery in BESs has completely changed the approach for sustainable WWT.
6. Conclusions
BESs are promising technologies for sustainable wastewater treatment (WWT) along with electricity generation. Complex interactions at the electrode-microbe interface can enhance the power output for practical applications in various monitoring devices and telemetry circuitries. Electrochemical characterization techniques in combination with microbiological methods are significantly contributing to the advancement of electro-microbiology. However, more collaborative interaction is highly recommended within various scientific disciplines to completely explore this area. Resource recovery along with WWT thorough BESs has become a progressing area of electro-microbiology, but more research is needed to enhance the efficiency of BESs for integration in future practices for progress toward a more sustainable society.
Author Contributions
J.A. conceived study, data analysis and wrote the manuscript, A.S. revised the tabulated data and helped in graphical work. L.W. critically revised article and M.R.H. and S.M. helped in data collection. G.P. critically revised and final approval of the manuscript to be published.
Acknowledgments
This work was supported by a University of Chinese Academy of Sciences (UCAS) Scholarship for International Students, the National Key R&D Program of China (2017YFA0207204, 2017YFA0207203), National Natural Science Foundation of China (21407160), Strategic Priority Research Program of the Chinese Academy of Sciences (XDA09030203) and Program of Research and Demonstration of the Large Lake Control Technology for Algal Blooms and Endogenous Pollution Control (Y7A0011001).
Conflicts of Interest
The authors declare no conflict of interests.
References
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial fuel cells: Methodology and technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef] [PubMed]
- Chabert, N.; Ali, O.A.; Achouak, W. All ecosystems potentially host electrogenic bacteria. Bioelectrochem. 2015, 106, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Nealson, K.H. Bioelectricity (electromicrobiology) and sustainability. Microb. Biotechnol. 2017, 10, 1114–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, M.C. Electrical effects accompanying the decomposition of organic compounds. Proc. R. Soc. Lond. Ser. B Contain. Papers Biol. Character 1911, 84, 260–276. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J. Novel mode of microbial energy metabolism: Organic carbon oxidation coupled to dissimilatory reduction of iron or manganese. Appl. Environ. Microbiol. 1988, 54, 1472–1480. [Google Scholar] [PubMed]
- Pant, D.; Van Bogaert, G.; Diels, L.; Vanbroekhoven, K. A review of the substrates used in microbial fuel cells (mfcs) for sustainable energy production. Bioresour. Technol. 2010, 101, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Rittmann, B.E. Opportunities for renewable bioenergy using microorganisms. Biotechnol. Bioeng. 2008, 100, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabaey, K.; Rozendal, R.A. Microbial electrosynthesis—revisiting the electrical route for microbial production. Nat. Rev. Microbiol. 2010, 8, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Sekar, N.; Ramasamy, R.P. Electrochemical impedance spectroscopy for microbial fuel cell characterization. J. Microb. Biochem. Technol. 2013. [Google Scholar] [CrossRef]
- Zhi, W.; Ge, Z.; He, Z.; Zhang, H. Methods for understanding microbial community structures and functions in microbial fuel cells: A review. Bioresour. Technol. 2014, 171, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Dolch, K.; Danzer, J.; Kabbeck, T.; Bierer, B.; Erben, J.; Förster, A.H.; Maisch, J.; Nick, P.; Kerzenmacher, S.; Gescher, J. Characterization of microbial current production as a function of microbe-electrode-interaction. Bioresour. Technol. 2014, 157, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Nevin, K.P. A shift in the current: New applications and concepts for microbe-electrode electron exchange. Curr. Opin. Biotechnol. 2011, 22, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Logan, B.E. Electricity generation using an air-cathode single chamber microbial fuel cell in the presence and absence of a proton exchange membrane. Environ. Sci. Technol. 2004, 38, 4040–4046. [Google Scholar] [CrossRef] [PubMed]
- Harnisch, F.; Freguia, S. A basic tutorial on cyclic voltammetry for the investigation of electroactive microbial biofilms. Chemistry–An Asian Journal 2012, 7, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Leang, C.; Franks, A.E.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Specific localization of the c-type cytochrome omcz at the anode surface in current-producing biofilms of geobacter sulfurreducens. Environ. Microbiol. Rep. 2011, 3, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Hsu, L.H.-H.; Kavanagh, P.; Barrière, F.; Lens, P.N.; Lapinsonnière, L.; Schröder, U.; Jiang, X.; Leech, D. The ins and outs of microorganism-electrode electron transfer reactions. Nat. Rev. Chem. 2017, 1. [Google Scholar] [CrossRef]
- Reguera, G.; McCarthy, K.D.; Mehta, T.; Nicoll, J.S.; Tuominen, M.T.; Lovley, D.R. Extracellular electron transfer via microbial nanowires. Nature 2005, 435, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Dong, H.; Reguera, G.; Beyenal, H.; Lu, A.; Liu, J.; Yu, H.-Q.; Fredrickson, J.K. Extracellular electron transfer mechanisms between microorganisms and minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Huang, L.; You, L.; Zhuang, L.; Zhou, S. Electrochemical and spectroscopic insights into the mechanisms of bidirectional microbe-electrode electron transfer in geobacter soli biofilms. Electrochem. Commun. 2017, 77, 93–97. [Google Scholar] [CrossRef]
- Okamoto, A.; Nakamura, R.; Hashimoto, K. In-vivo identification of direct electron transfer from shewanella oneidensis mr-1 to electrodes via outer-membrane omca–mtrcab protein complexes. Electrochim. Acta. 2011, 56, 5526–5531. [Google Scholar] [CrossRef]
- Lovley, D.R. Electromicrobiology. Annu. Rev. Microbiol. 2012, 66, 391–409. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Qian, X.; Morgado, L.; Kim, B.-C.; Mester, T.; Izallalen, M.; Salgueiro, C.A.; Lovley, D.R. Purification and characterization of omcz, an outer-surface, octaheme c-type cytochrome essential for optimal current production by geobacter sulfurreducens. Appl. Environ. Microbiol. 2010, 76, 3999–4007. [Google Scholar] [CrossRef] [PubMed]
- Baek, G.; Kim, J.; Kim, J.; Lee, C. Role and potential of direct interspecies electron transfer in anaerobic digestion. Energies 2018, 11. [Google Scholar] [CrossRef]
- Lovley, D.R. Syntrophy goes electric: Direct interspecies electron transfer. Annu. Rev. Microbiol. 2017, 71, 643–664. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D.E.; Shrestha, P.M.; Walker, D.J.; Dang, Y.; Nevin, K.P.; Woodard, T.L.; Lovley, D.R. Metatranscriptomic evidence for direct interspecies electron transfer between geobacter and methanothrix species in methanogenic rice paddy soils. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qiao, Y.; Guo, C.X.; Lim, S.; Song, H.; Li, C.M. Graphene/carbon cloth anode for high-performance mediatorless microbial fuel cells. Bioresour. Technol. 2012, 114, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Squier, T.C.; Zachara, J.M.; Fredrickson, J.K. Respiration of metal (hydr) oxides by shewanella and geobacter: A key role for multihaem c-type cytochromes. Mol. Microbiol. 2007, 65, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Bond, D.R.; Lovley, D.R. Evidence for involvement of an electron shuttle in electricity generation by geothrix fermentans. Appl. Environ. Microbiol. 2005, 71, 2186–2189. [Google Scholar] [CrossRef] [PubMed]
- Yong, X.-Y.; Shi, D.-Y.; Chen, Y.-L.; Jiao, F.; Lin, X.; Zhou, J.; Wang, S.-Y.; Yong, Y.-C.; Sun, Y.-M.; OuYang, P.-K. Enhancement of bioelectricity generation by manipulation of the electron shuttles synthesis pathway in microbial fuel cells. Bioresour. Technol. 2014, 152, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ding, Y.; Hu, Y.; Cao, B.; Rice, S.A.; Kjelleberg, S.; Song, H. Enhancing bidirectional electron transfer of shewanella oneidensis by a synthetic flavin pathway. ACS Synth. Biol. 2015, 4, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Marsili, E.; Baron, D.B.; Shikhare, I.D.; Coursolle, D.; Gralnick, J.A.; Bond, D.R. Shewanella secretes flavins that mediate extracellular electron transfer. Proc. Natl. Acad. Sci. USA 2008, 105, 3968–3973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, G.; Bakonyi, P.; Zhen, G.; Sivagurunathan, P.; Koók, L.; Kim, S.-H.; Tóth, G.; Nemestóthy, N.; Bélafi-Bakó, K. Microbial electrochemical systems for sustainable biohydrogen production: Surveying the experiences from a start-up viewpoint. Renew. Sustain. Energy Rev. 2017, 70, 589–597. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, J.; Qu, Y.; Feng, Y. Enhanced shewanella oneidensis mr-1 anode performance by adding fumarate in microbial fuel cell. Chem. Eng. J. 2017, 328, 697–702. [Google Scholar] [CrossRef]
- Okamoto, A.; Saito, K.; Inoue, K.; Nealson, K.H.; Hashimoto, K.; Nakamura, R. Uptake of self-secreted flavins as bound cofactors for extracellular electron transfer in geobacter species. Energy Environ. Sci. 2014, 7, 1357–1361. [Google Scholar] [CrossRef]
- Lovley, D.R. Long-range electron transport to fe (iii) oxide via pili with metallic-like conductivity. Biochem. Soc. Trans. 2012, 40, 1186–1190. [Google Scholar] [CrossRef] [PubMed]
- Kracke, F.; Vassilev, I.; Krömer, J.O. Microbial electron transport and energy conservation–the foundation for optimizing bioelectrochemical systems. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Nevin, K.P.; Hensley, S.A.; Franks, A.E.; Summers, Z.M.; Ou, J.; Woodard, T.L.; Snoeyenbos-West, O.L.; Lovley, D.R. Electrosynthesis of organic compounds from carbon dioxide catalyzed by a diversity of acetogenic microorganisms. Appl. Environ. Microbiol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E. Exoelectrogenic bacteria that power microbial fuel cells. Nat. Rev. Microbiol. 2009, 7, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Bosire, E.M.; Blank, L.M.; Rosenbaum, M.A. Strain and substrate dependent redox mediator and electricity production by pseudomonas aeruginosa. Appl. Environ. Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, P.L.; Aklujkar, M.; Leang, C.; Nevin, K.P.; Lovley, D. A genetic system for geobacter metallireducens: Role of the flagellin and pilin in the reduction of fe (iii) oxide. Environ. Microbiol. Rep. 2012, 4, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.M.; Myers, C.R. Role for outer membrane cytochromes omca and omcb of shewanella putrefaciens mr-1 in reduction of manganese dioxide. Appl. Environ. Microbiol. 2001, 67, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Venkidusamy, K.; Megharaj, M. A novel electrophototrophic bacterium rhodopseudomonas palustris strain rp2, exhibits hydrocarbonoclastic potential in anaerobic environments. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, X.; Zhang, W.; Hu, M.; Li, F. Fe (iii) oxides accelerate microbial nitrate reduction and electricity generation by klebsiella pneumoniae l17. J. Colloid. Interface Sci. 2014, 423, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.W.; Ross, D.E.; Fichot, E.B.; Norman, R.S.; May, H.D. Electrosynthesis of commodity chemicals by an autotrophic microbial community. Appl. Environ. Microbiol. 2012, 78, 8412–8420. [Google Scholar] [CrossRef] [PubMed]
- Freguia, S.; Masuda, M.; Tsujimura, S.; Kano, K. Lactococcus lactis catalyses electricity generation at microbial fuel cell anodes via excretion of a soluble quinone. Bioelectrochemistry 2009, 76, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Coursolle, D.; Gralnick, J.A. Modularity of the mtr respiratory pathway of shewanella oneidensis strain mr-1. Mol. Microbiol. 2010, 77, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Le Laz, S.; Kpebe, A.; Lorquin, J.; Brugna, M.; Rousset, M. H 2-dependent azoreduction by shewanella oneidensis mr-1: Involvement of secreted flavins and both [ni–fe] and [fe–fe] hydrogenases. Appl. Microbiol. Biotechnol. 2014, 98, 2699–2707. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Ishikawa, M.; Hashimoto, K.; Nakanishi, S. Molecular design of cytocompatible amphiphilic redox-active polymers for efficient extracellular electron transfer. Bioelectrochemistry 2017, 114, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Slade, R.C.; Varcoe, J.R. Techniques for the study and development of microbial fuel cells: An electrochemical perspective. Chem. Soc. Rev. 2009, 38, 1926–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, C.; Li, J.; Qin, D.; Chen, L.; Zhao, F.; Chen, S.; Hu, H.; Yu, C.-P. Characterization of exoelectrogenic bacteria enterobacter strains isolated from a microbial fuel cell exposed to copper shock load. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Heidrich, E.S.; Curtis, T.P.; Woodcock, S.; Dolfing, J. Quantification of effective exoelectrogens by most probable number (mpn) in a microbial fuel cell. Bioresour. Technol. 2016, 218, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Wrighton, K.C.; Agbo, P.; Warnecke, F.; Weber, K.A.; Brodie, E.L.; DeSantis, T.Z.; Hugenholtz, P.; Andersen, G.L.; Coates, J.D. A novel ecological role of the firmicutes identified in thermophilic microbial fuel cells. ISME J. 2008, 2, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Sacco, N.J.; Bonetto, M.C.; Cortón, E. Isolation and characterization of a novel electrogenic bacterium, dietzia sp. Rnv-4. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Deng, D.; Zhang, D.; Chen, Q.; Kang, J.; Fan, N.; Liu, Y. Microbial electricity generation and isolation of exoelectrogenic bacteria based on petroleum hydrocarbon-contaminated soil. Electroanalysis 2016, 28, 1510–1516. [Google Scholar] [CrossRef]
- Zuo, Y.; Xing, D.; Regan, J.M.; Logan, B.E. Isolation of the exoelectrogenic bacterium ochrobactrum anthropi yz-1 by using a u-tube microbial fuel cell. Appl. Environ. Microbiol. 2008, 74, 3130–3137. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhang, Y.; Quan, X.; Chen, S. Anaerobic biodecolorization of ao7 by a newly isolated fe (iii)-reducing bacterium sphingomonas strain dj. J. Chem. Technol. Biotechnol. 2015, 90, 158–165. [Google Scholar] [CrossRef]
- Xing, D.; Zuo, Y.; Cheng, S.; Regan, J.M.; Logan, B.E. Electricity generation by rhodopseudomonas palustris dx-1. Environ. Sci. Technol. 2008, 42, 4146–4151. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Call, D.; Wang, A.; Cheng, S.; Logan, B.E. Geobacter sp. Sd-1 with enhanced electrochemical activity in high-salt concentration solutions. Environ. Microbiol. Rep. 2014, 6, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Lee, D.-J.; Wang, A.; Ren, N.; Su, A.; Lai, J.-Y. Isolation of fe (iii)-reducing bacterium, citrobacter sp. Lar-1, for startup of microbial fuel cell. Int. J. Hydrogen Energy 2016, 41, 4498–4503. [Google Scholar] [CrossRef]
- Nercessian, O.; Parot, S.; Délia, M.-L.; Bergel, A.; Achouak, W. Harvesting electricity with geobacter bremensis isolated from compost. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Wang, Y.; Qin, D.; Yang, G.; Zhou, S. Bacillus sediminis sp. Nov., isolated from an electroactive biofilm. Antonie Van Leeuwenhoek 2013, 104, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Kim, B.H.; Kim, H.S.; Kim, H.J.; Kim, G.T.; Kim, M.; Chang, I.S.; Park, Y.K.; Chang, H.I. A novel electrochemically active and fe (iii)-reducing bacterium phylogenetically related to clostridium butyricum isolated from a microbial fuel cell. Anaerobe 2001, 7, 297–306. [Google Scholar] [CrossRef]
- Finneran, K.T.; Johnsen, C.V.; Lovley, D.R. Rhodoferax ferrireducens sp. Nov., a psychrotolerant, facultatively anaerobic bacterium that oxidizes acetate with the reduction of fe (iii). Int. J. Syst. Evol. Microbiol. 2003, 53, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Hyun, M.; Chang, I.; Kim, H.; Park, H.; Kim, B.; Kim, S.; Wimpenny, J.; Weightman, A.J. Dissimilatory fe (iii) reduction by an electrochemically active lactic acid bacterium phylogenetically related to enterococcus gallinarum isolated from submerged soil. J. Appl. Microbiol. 2005, 99, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Pham, C.A.; Jung, S.J.; Phung, N.T.; Lee, J.; Chang, I.S.; Kim, B.H.; Yi, H.; Chun, J. A novel electrochemically active and fe (iii)-reducing bacterium phylogenetically related to aeromonas hydrophila, isolated from a microbial fuel cell. FEMS Microbiol. Lett. 2003, 223, 129–134. [Google Scholar] [CrossRef]
- Wei, Z.; Hu, X.; Li, X.; Zhang, Y.; Jiang, L.; Li, J.; Guan, Z.; Cai, Y.; Liao, X. The rhizospheric microbial community structure and diversity of deciduous and evergreen forests in taihu lake area, china. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Baranitharan, E.; Khan, M.R.; Prasad, D.; Teo, W.F.A.; Tan, G.Y.A.; Jose, R. Effect of biofilm formation on the performance of microbial fuel cell for the treatment of palm oil mill effluent. Bioprocess Biosyst. Eng. 2015, 38, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Roy, S.; Das, D. Genomic and proteomic approaches for dark fermentative biohydrogen production. Renew. Sustain. Energy Rev. 2016, 56, 1308–1321. [Google Scholar] [CrossRef]
- Parameswaran, P.; Torres, C.I.; Lee, H.-S.; Rittmann, B.E.; Krajmalnik-Brown, R. Hydrogen consumption in microbial electrochemical systems (mxcs): The role of homo-acetogenic bacteria. Bioresour. Technol. 2011, 102, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Kiely, P.D.; Cusick, R.; Call, D.F.; Selembo, P.A.; Regan, J.M.; Logan, B.E. Anode microbial communities produced by changing from microbial fuel cell to microbial electrolysis cell operation using two different wastewaters. Bioresour. Technol. 2011, 102, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.i.; Suzuki, S.; Yamanaka, Y.; Wu, A.; Nealson, K.H.; Bretschger, O. Population dynamics of electrogenic microbial communities in microbial fuel cells started with three different inoculum sources. Bioelectrochemistry 2017. [Google Scholar] [CrossRef] [PubMed]
- Paitier, A.; Godain, A.; Lyon, D.; Haddour, N.; Vogel, T.M.; Monier, J.-M. Microbial fuel cell anodic microbial population dynamics during mfc start-up. Biosens. Bioelectron. 2017, 92, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Hungate, B.A.; Mau, R.L.; Schwartz, E.; Caporaso, J.G.; Dijkstra, P.; van Gestel, N.; Koch, B.J.; Liu, C.M.; McHugh, T.A.; Marks, J.C. Quantitative microbial ecology through stable isotope probing. Appl. Environ. Microbiol. 2015, 81, 7570–7581. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.i.; Suzuki, S.; Norden-Krichmar, T.M.; Tenney, A.; Chain, P.S.; Scholz, M.B.; Nealson, K.H.; Bretschger, O. A novel metatranscriptomic approach to identify gene expression dynamics during extracellular electron transfer. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef]
- McLean, J.S.; Wanger, G.; Gorby, Y.A.; Wainstein, M.; McQuaid, J.; Ishii, S.i.; Bretschger, O.; Beyenal, H.; Nealson, K.H. Quantification of electron transfer rates to a solid phase electron acceptor through the stages of biofilm formation from single cells to multicellular communities. Environ. Sci. Technol. 2010, 44, 2721–2727. [Google Scholar] [CrossRef] [PubMed]
- Nevin, K.P.; Kim, B.-C.; Glaven, R.H.; Johnson, J.P.; Woodard, T.L.; Methé, B.A.; DiDonato Jr, R.J.; Covalla, S.F.; Franks, A.E.; Liu, A. Anode biofilm transcriptomics reveals outer surface components essential for high density current production in geobacter sulfurreducens fuel cells. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Marsili, E.; Rollefson, J.B.; Baron, D.B.; Hozalski, R.M.; Bond, D.R. Microbial biofilm voltammetry: Direct electrochemical characterization of catalytic electrode-attached biofilms. Appl. Environ. Microbiol. 2008, 74, 7329–7337. [Google Scholar] [CrossRef] [PubMed]
- Strycharz-Glaven, S.M.; Snider, R.M.; Guiseppi-Elie, A.; Tender, L.M. On the electrical conductivity of microbial nanowires and biofilms. Energy Environ. Sci. 2011, 4, 4366–4379. [Google Scholar] [CrossRef]
- Manohar, A.K.; Bretschger, O.; Nealson, K.H.; Mansfeld, F. The use of electrochemical impedance spectroscopy (eis) in the evaluation of the electrochemical properties of a microbial fuel cell. Bioelectrochemistry 2008, 72, 149–154. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Mansfeld, F. Exploring the use of electrochemical impedance spectroscopy (eis) in microbial fuel cell studies. Energy Environ. Sci. 2009, 2, 215–219. [Google Scholar] [CrossRef]
- Borole, A.P.; Aaron, D.; Hamilton, C.Y.; Tsouris, C. Understanding long-term changes in microbial fuel cell performance using electrochemical impedance spectroscopy. Environ. Sci. Technol. 2010, 44, 2740–2745. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Herrera, D.; Pacheco-Catalan, D.; Valdez-Ojeda, R.; Canto-Canche, B.; Dominguez-Benetton, X.; Domínguez-Maldonado, J.; Alzate-Gaviria, L. Characterization of anode and anolyte community growth and the impact of impedance in a microbial fuel cell. BMC Biotechnol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Babauta, J.T.; Beyenal, H. Use of a small overpotential approximation to analyze geobacter sulfurreducens biofilm impedance. J. Power Sources 2017, 356, 549–555. [Google Scholar] [CrossRef]
- Qiao, Y.-J.; Qiao, Y.; Zou, L.; Wu, X.-S.; Liu, J.-H. Biofilm promoted current generation of pseudomonas aeruginosa microbial fuel cell via improving the interfacial redox reaction of phenazines. Bioelectrochemistry 2017. [Google Scholar] [CrossRef] [PubMed]
- Cercado-Quezada, B.; Delia, M.-L.; Bergel, A. Testing various food-industry wastes for electricity production in microbial fuel cell. Bioresour. Technol. 2010, 101, 2748–2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fricke, K.; Harnisch, F.; Schröder, U. On the use of cyclic voltammetry for the study of anodic electron transfer in microbial fuel cells. Energy Environ. Sci. 2008, 1, 144–147. [Google Scholar] [CrossRef]
- Chen, S.; Patil, S.A.; Schröder, U. A high-performance rotating graphite fiber brush air-cathode for microbial fuel cells. Appl. Energy 2018, 211, 1089–1094. [Google Scholar] [CrossRef]
- Tsai, W.-T.; Chen, C.-C.; Lin, Y.-Q.; Hsiao, C.-F.; Tsai, C.-H.; Hsieh, M.-H. Status of waste tires’ recycling for material and energy resources in taiwan. J. Mater. Cycles. Waste Manag. 2017, 19, 1288–1294. [Google Scholar] [CrossRef]
- Rosenbaum, M.; Zhao, F.; Quaas, M.; Wulff, H.; Schröder, U.; Scholz, F. Evaluation of catalytic properties of tungsten carbide for the anode of microbial fuel cells. Appl. Catal. B Environ. 2007, 74, 261–269. [Google Scholar] [CrossRef]
- Zou, L.; Qiao, Y.; Zhong, C.; Li, C.M. Enabling fast electron transfer through both bacterial outer-membrane redox centers and endogenous electron mediators by polyaniline hybridized large-mesoporous carbon anode for high-performance microbial fuel cells. Electrochim. Acta. 2017, 229, 31–38. [Google Scholar] [CrossRef]
- Rush, A.J.; Trivedi, M.H.; Wisniewski, S.R.; Nierenberg, A.A.; Stewart, J.W.; Warden, D.; Niederehe, G.; Thase, M.E.; Lavori, P.W.; Lebowitz, B.D. Acute and longer-term outcomes in depressed outpatients requiring one or several treatment steps: A star* d report. Am. J. Psychiatry 2006, 163, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E.; Rabaey, K. Conversion of wastes into bioelectricity and chemicals by using microbial electrochemical technologies. Science 2012, 337, 686–690. [Google Scholar] [CrossRef] [PubMed]
- Jafar Ali, S.Z.; Ali, N. Green synthesis of metal nanoparticles by microorganisms; a current prospective. J. Nanoanal 2015, 2, 32–38. [Google Scholar] [CrossRef]
- Cusick, R.D.; Kiely, P.D.; Logan, B.E. A monetary comparison of energy recovered from microbial fuel cells and microbial electrolysis cells fed winery or domestic wastewaters. Int. J. Hydrogen Energy 2010, 35, 8855–8861. [Google Scholar] [CrossRef]
- Liang, B.; Kong, D.; Ma, J.; Wen, C.; Yuan, T.; Lee, D.-J.; Zhou, J.; Wang, A. Low temperature acclimation with electrical stimulation enhance the biocathode functioning stability for antibiotics detoxification. Water Res. 2016, 100, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Liu, Y.; Ma, J.; Zhao, F. Rapid degradation of sulphamethoxazole and the further transformation of 3-amino-5-methylisoxazole in a microbial fuel cell. Water Res. 2016, 88, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.; Kong, F.; Zheng, H.; Yin, J.; Cao, D.; Ren, Y.; Wang, G. Simultaneous processes of electricity generation and ceftriaxone sodium degradation in an air-cathode single chamber microbial fuel cell. J. Power Sources 2011, 196, 2567–2572. [Google Scholar] [CrossRef]
- Liu, W.; Cheng, S.; Sun, D.; Huang, H.; Chen, J.; Cen, K. Inhibition of microbial growth on air cathodes of single chamber microbial fuel cells by incorporating enrofloxacin into the catalyst layer. Biosens. Bioelectron. 2015, 72, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feng, C.; Li, Y.; Gao, J.; Yu, C.-P. Enhancement of emerging contaminants removal using fenton reaction driven by h2o2-producing microbial fuel cells. Chem. Eng. J. 2017, 307, 679–686. [Google Scholar] [CrossRef]
- Pandey, P.; Shinde, V.N.; Deopurkar, R.L.; Kale, S.P.; Patil, S.A.; Pant, D. Recent advances in the use of different substrates in microbial fuel cells toward wastewater treatment and simultaneous energy recovery. Appl. Energy 2016, 168, 706–723. [Google Scholar] [CrossRef]
- Oh, S.; Logan, B.E. Hydrogen and electricity production from a food processing wastewater using fermentation and microbial fuel cell technologies. Water Res. 2005, 39, 4673–4682. [Google Scholar] [CrossRef] [PubMed]
- Mohanakrishna, G.; Mohan, S.V.; Sarma, P. Bio-electrochemical treatment of distillery wastewater in microbial fuel cell facilitating decolorization and desalination along with power generation. J. Hazard Mater. 2010, 177, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ni, J. Treatment of wastewater from dioscorea zingiberensis tubers used for producing steroid hormones in a microbial fuel cell. Bioresour. Technol. 2011, 102, 2731–2735. [Google Scholar] [CrossRef] [PubMed]
- Sharma, Y.; Parnas, R.; Li, B. Bioenergy production from glycerol in hydrogen producing bioreactors (hpbs) and microbial fuel cells (mfcs). Int. J. Hydrogen Energy 2011, 36, 3853–3861. [Google Scholar] [CrossRef]
- Sun, J.; Hu, Y.-Y.; Bi, Z.; Cao, Y.-Q. Simultaneous decolorization of azo dye and bioelectricity generation using a microfiltration membrane air-cathode single-chamber microbial fuel cell. Bioresour. Technol. 2009, 100, 3185–3192. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhang, A.; Li, J.; He, B. A continuous process for biodiesel production in a fixed bed reactor packed with cation-exchange resin as heterogeneous catalyst. Bioresour. Technol. 2011, 102, 3607–3609. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Zhou, S.-G.; Zhuang, L.; Zhang, J.-T.; Ni, J.-R. Electricity generation from starch processing wastewater using microbial fuel cell technology. Biochem. Eng. J. 2009, 43, 246–251. [Google Scholar] [CrossRef]
- Huang, L.; Logan, B.E. Electricity generation and treatment of paper recycling wastewater using a microbial fuel cell. Appl. Microbiol. Biotechnol. 2008, 80, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Behera, M.; Jana, P.S.; More, T.T.; Ghangrekar, M. Rice mill wastewater treatment in microbial fuel cells fabricated using proton exchange membrane and earthen pot at different ph. Bioelectrochemistry 2010, 79, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zhu, X.; Ni, J.; Borthwick, A. Palm oil mill effluent treatment using a two-stage microbial fuel cells system integrated with immobilized biological aerated filters. Bioresour. Technol. 2010, 101, 2729–2734. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.; Kong, F.; Zheng, H.; Cao, D.; Ren, Y.; Yin, J. Electricity generation from synthetic penicillin wastewater in an air-cathode single chamber microbial fuel cell. Chem. Eng. J. 2011, 168, 572–576. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, R.; Liu, G.; Li, J.; Li, M.; Zhang, C. Electricity generation from indole and microbial community analysis in the microbial fuel cell. J. Hazard. Mater. 2010, 176, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhao, H.; Zhou, S.; Shi, C.; Wang, C.; Ni, J. A novel uasb–mfc–baf integrated system for high strength molasses wastewater treatment and bioelectricity generation. Bioresour. Technol. 2009, 100, 5687–5693. [Google Scholar] [CrossRef] [PubMed]
- Catal, T.; Bermek, H.; Liu, H. Removal of selenite from wastewater using microbial fuel cells. Biotechnol. Lett. 2009, 31, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Ni, J. Simultaneous processes of electricity generation and p-nitrophenol degradation in a microbial fuel cell. Electrochem. Commun. 2009, 11, 274–277. [Google Scholar] [CrossRef]
- Luo, H.; Xu, P.; Roane, T.M.; Jenkins, P.E.; Ren, Z. Microbial desalination cells for improved performance in wastewater treatment, electricity production, and desalination. Bioresour. Technol. 2012, 105, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Mathuriya, A.S.; Sharma, V. Bioelectricity production from various wastewaters through microbial fuel cell technology. J. Biochem. Technol. 2010, 2, 133–137. [Google Scholar]
- Yu, C.-P.; Liang, Z.; Das, A.; Hu, Z. Nitrogen removal from wastewater using membrane aerated microbial fuel cell techniques. Water Res. 2011, 45, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Neburchilov, V.; Mehta, P.; Hussain, A.; Wang, H.; Guiot, S.; Tartakovsky, B. Microbial fuel cell operation on carbon monoxide: Cathode catalyst selection. Int. J. Hydrogen Energy 2011, 36, 11929–11935. [Google Scholar] [CrossRef] [Green Version]
- Pham, H.; Boon, N.; Marzorati, M.; Verstraete, W. Enhanced removal of 1, 2-dichloroethane by anodophilic microbial consortia. Water Res. 2009, 43, 2936–2946. [Google Scholar] [CrossRef] [PubMed]
- Raghavulu, S.V.; Mohan, S.V.; Reddy, M.V.; Mohanakrishna, G.; Sarma, P. Behavior of single chambered mediatorless microbial fuel cell (mfc) at acidophilic, neutral and alkaline microenvironments during chemical wastewater treatment. Int. J. Hydrogen Energy 2009, 34, 7547–7554. [Google Scholar] [CrossRef]
- You, S.J.; Zhao, Q.L.; Jiang, J.Q.; Zhang, J.N.; Zhao, S.Q. Sustainable approach for leachate treatment: Electricity generation in microbial fuel cell. J. Environ. Sci. Health Part A 2006, 41, 2721–2734. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Zhang, X.; Li, Z.; Lei, L. Studies on treatment of chlorophenol-containing wastewater by microbial fuel cell. Chin. Sci. Bull. 2007, 52, 3448–3451. [Google Scholar] [CrossRef]
- Fu, L.; You, S.-J.; Zhang, G.-q.; Yang, F.-L.; Fang, X.-h. Degradation of azo dyes using in-situ fenton reaction incorporated into h 2 o 2-producing microbial fuel cell. Chem. Eng. J. 2010, 160, 164–169. [Google Scholar] [CrossRef]
- Zhou, Y.-F.; Haynes, R.J. Removal of pb (ii), cr (iii) and cr (vi) from aqueous solutions using alum-derived water treatment sludge. Water Air Soil Pollut. 2011, 215, 631–643. [Google Scholar] [CrossRef]
- Frontera-Suau, R.; Bost, F.D.; McDonald, T.J.; Morris, P.J. Aerobic biodegradation of hopanes and other biomarkers by crude oil-degrading enrichment cultures. Environ. Sci. Technol. 2002, 36, 4585–4592. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Xu, Y.; Ye, Y.; Chen, Y.; Shen, S. Electricity generation from terephthalic acid using a microbial fuel cell. J. Chem. Technol. Biotechnol. 2009, 84, 356–360. [Google Scholar] [CrossRef]
- Morris, J.M.; Jin, S. Feasibility of using microbial fuel cell technology for bioremediation of hydrocarbons in groundwater. J. Environ. Sci. Health Part A 2007, 43, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.; Wu, Y.; Zhao, L.; Sun, Q. Production of electricity from the treatment of continuous brewery wastewater using a microbial fuel cell. Fuel 2010, 89, 1381–1385. [Google Scholar] [CrossRef]
- Kim, B.H.; Park, H.; Kim, H.; Kim, G.; Chang, I.; Lee, J.; Phung, N. Enrichment of microbial community generating electricity using a fuel-cell-type electrochemical cell. Appl. Microbiol. Biotechnol. 2004, 63, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Ter Heijne, A.; Hamelers, H.V.; De Wilde, V.; Rozendal, R.A.; Buisman, C.J. A bipolar membrane combined with ferric iron reduction as an efficient cathode system in microbial fuel cells. Environ. Sci. Technol. 2006, 40, 5200–5205. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ren, Z.J. Bioelectrochemical metal recovery from wastewater: A review. Water Res. 2014, 66, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P.T.; He, Z. Nutrients removal and recovery in bioelectrochemical systems: A review. Bioresour. Technol. 2014, 153, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Damiano, L.; Jambeck, J.R.; Ringelberg, D.B. Municipal solid waste landfill leachate treatment and electricity production using microbial fuel cells. Appl. Biochem. Biotechnol. 2014, 173, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Tanaka, Y.; Kuroda, K.; Hanajima, D.; Fukumoto, Y.; Yasuda, T.; Waki, M. Removal and recovery of phosphorous from swine wastewater by demonstration crystallization reactor and struvite accumulation device. Bioresour. Technol. 2007, 98, 1573–1578. [Google Scholar] [CrossRef] [PubMed]
- Van Eerten-Jansen, M.C.; Jansen, N.C.; Plugge, C.M.; de Wilde, V.; Buisman, C.J.; ter Heijne, A. Analysis of the mechanisms of bioelectrochemical methane production by mixed cultures. J. Chem. Technol. Biotechnol. 2015, 90, 963–970. [Google Scholar] [CrossRef]
- Satar, I.; Daud, W.R.W.; Kim, B.H.; Somalu, M.R.; Ghasemi, M. Immobilized mixed-culture reactor (imcr) for hydrogen and methane production from glucose. Energy 2017, 139, 1188–1196. [Google Scholar] [CrossRef]
- Kumar, G.; Saratale, R.G.; Kadier, A.; Sivagurunathan, P.; Zhen, G.; Kim, S.-H.; Saratale, G.D. A review on bio-electrochemical systems (bess) for the syngas and value added biochemicals production. Chemosphere 2017, 177, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Rader, G.K.; Logan, B.E. Multi-electrode continuous flow microbial electrolysis cell for biogas production from acetate. Int. J. Hydrogen Energy 2010, 35, 8848–8854. [Google Scholar] [CrossRef]
- McAnulty, M.J.; Poosarla, V.G.; Kim, K.-Y.; Jasso-Chávez, R.; Logan, B.E.; Wood, T.K. Electricity from methane by reversing methanogenesis. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Kreysa, G.; Håkansson, B. Electrocatalysis by amorphous metals of hydrogen and oxygen evolution in alkaline solution. J. Electroanal. Chem. Interfacial Electrochem. 1986, 201, 61–83. [Google Scholar] [CrossRef]
- Logan, B.; Cheng, S.; Watson, V.; Estadt, G. Graphite fiber brush anodes for increased power production in air-cathode microbial fuel cells. Environ. Sci. Technol. 2007, 41, 3341–3346. [Google Scholar] [CrossRef] [PubMed]
- Guwy, A.; Dinsdale, R.; Kim, J.; Massanet-Nicolau, J.; Premier, G. Fermentative biohydrogen production systems integration. Bioresour. Technol. 2011, 102, 8534–8542. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E.; Call, D.; Cheng, S.; Hamelers, H.V.; Sleutels, T.H.; Jeremiasse, A.W.; Rozendal, R.A. Microbial electrolysis cells for high yield hydrogen gas production from organic matter. Environ. Sci. Technol. 2008, 42, 8630–8640. [Google Scholar] [CrossRef] [PubMed]
- Jafary, T.; Daud, W.R.W.; Ghasemi, M.; Kim, B.H.; Carmona-Martínez, A.A.; Bakar, M.H.A.; Jahim, J.M.; Ismail, M. A comprehensive study on development of a biocathode for cleaner production of hydrogen in a microbial electrolysis cell. J. Clean. Prod. 2017, 164, 1135–1144. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, E.; Zhang, J.; Dai, Y.; Yang, Z.; Christensen, H.E.; Ulstrup, J.; Zhao, F. Extracellular polymeric substances are transient media for microbial extracellular electron transfer. Sci. Adv. 2017, 3. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.W.; Ross, D.E.; Fichot, E.B.; Norman, R.S.; May, H.D. Long-term operation of microbial electrosynthesis systems improves acetate production by autotrophic microbiomes. Environ. Sci. Technol. 2013, 47, 6023–6029. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekhar, K.; Lee, Y.-J.; Lee, D.-W. Biohydrogen production: Strategies to improve process efficiency through microbial routes. Int. J. Mol. Sci. 2015, 16, 8266–8293. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.A.; Arends, J.B.; Vanwonterghem, I.; Van Meerbergen, J.; Guo, K.; Tyson, G.W.; Rabaey, K. Selective enrichment establishes a stable performing community for microbial electrosynthesis of acetate from CO2. Environ. Sci. Technol. 2015, 49, 8833–8843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escapa, A.; Mateos, R.; Martínez, E.; Blanes, J. Microbial electrolysis cells: An emerging technology for wastewater treatment and energy recovery. From laboratory to pilot plant and beyond. Renew. Sustain. Energy Rev. 2016, 55, 942–956. [Google Scholar] [CrossRef]
- Tang, H.; Zeng, Y.; Zeng, Y.; Wang, R.; Cai, S.; Liao, C.; Cai, H.; Lu, X.; Tsiakaras, P. Iron-embedded nitrogen doped carbon frameworks as robust catalyst for oxygen reduction reaction in microbial fuel cells. Appl. Catal. B Environ. 2017, 202, 550–556. [Google Scholar] [CrossRef]
- Thostenson, J.O.; Ngaboyamahina, E.; Sellgren, K.L.; Hawkins, B.T.; Piascik, J.R.; Klem, E.J.; Parker, C.B.; Deshusses, M.A.; Stoner, B.R.; Glass, J.T. Enhanced H2O2 production at reductive potentials from oxidized boron-doped ultrananocrystalline diamond electrodes. ACS Appl. Mater. Interfaces 2017, 9, 16610–16619. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; You, S.J.; Yang, F.l.; Gao, M.m.; Fang, X.h.; Zhang, G.Q. Synthesis of hydrogen peroxide in microbial fuel cell. J. Chem. Technol. Biotechnol. 2010, 85, 715–719. [Google Scholar] [CrossRef]
- Asghar, A.; Raman, A.A.A.; Daud, W.M.A.W. Advanced oxidation processes for in-situ production of hydrogen peroxide/hydroxyl radical for textile wastewater treatment: A review. J. Clean. Prod. 2015, 87, 826–838. [Google Scholar] [CrossRef]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.-J.; Chang, J.-S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Ali, J.; Ali, N.; Jamil, S.U.U.; Waseem, H.; Khan, K.; Pan, G. Insight into eco-friendly fabrication of silver nanoparticles by pseudomonas aeruginosa and its potential impacts. J. Environ. Chem. Eng. 2017, 5, 3266–3272. [Google Scholar] [CrossRef]
- Ali, J.; Hameed, A.; Ahmed, S.; Ali, M.I.; Zainab, S.; Ali, N. Role of catalytic protein and stabilising agents in the transformation of ag ions to nanoparticles by pseudomonas aeruginosa. IET Nanobiotechnol. 2016, 10, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Commault, A.S.; Laczka, O.; Siboni, N.; Tamburic, B.; Crosswell, J.R.; Seymour, J.R.; Ralph, P.J. Electricity and biomass production in a bacteria-chlorella based microbial fuel cell treating wastewater. J. Power Sources 2017, 356, 299–309. [Google Scholar] [CrossRef]
- Saba, B.; Christy, A.D.; Yu, Z.; Co, A.C. Sustainable power generation from bacterio-algal microbial fuel cells (mfcs): An overview. Renew. Sustain. Energy Rev. 2017, 73, 75–84. [Google Scholar] [CrossRef]
- Xie, T.; Reddy, K.R.; Wang, C.; Yargicoglu, E.; Spokas, K. Characteristics and applications of biochar for environmental remediation: A review. Crit. Rev. Environ. Sci. Technol. 2015, 45, 939–969. [Google Scholar] [CrossRef]
- Lawton, R.J.; Cole, A.J.; Roberts, D.A.; Paul, N.A.; de Nys, R. The industrial ecology of freshwater macroalgae for biomass applications. Algal. Res. 2017, 24, 486–491. [Google Scholar] [CrossRef]
Figure 1.
Schematic illustration of extracellular electron transfer mechanisms at microbe-electrode interface in bioelectrochemical systems, (A) Direct electron transfer; (B) Indirect electron transfer, adapted and modified from [16] with the permission from nature publishing group.
Figure 1.
Schematic illustration of extracellular electron transfer mechanisms at microbe-electrode interface in bioelectrochemical systems, (A) Direct electron transfer; (B) Indirect electron transfer, adapted and modified from [16] with the permission from nature publishing group.
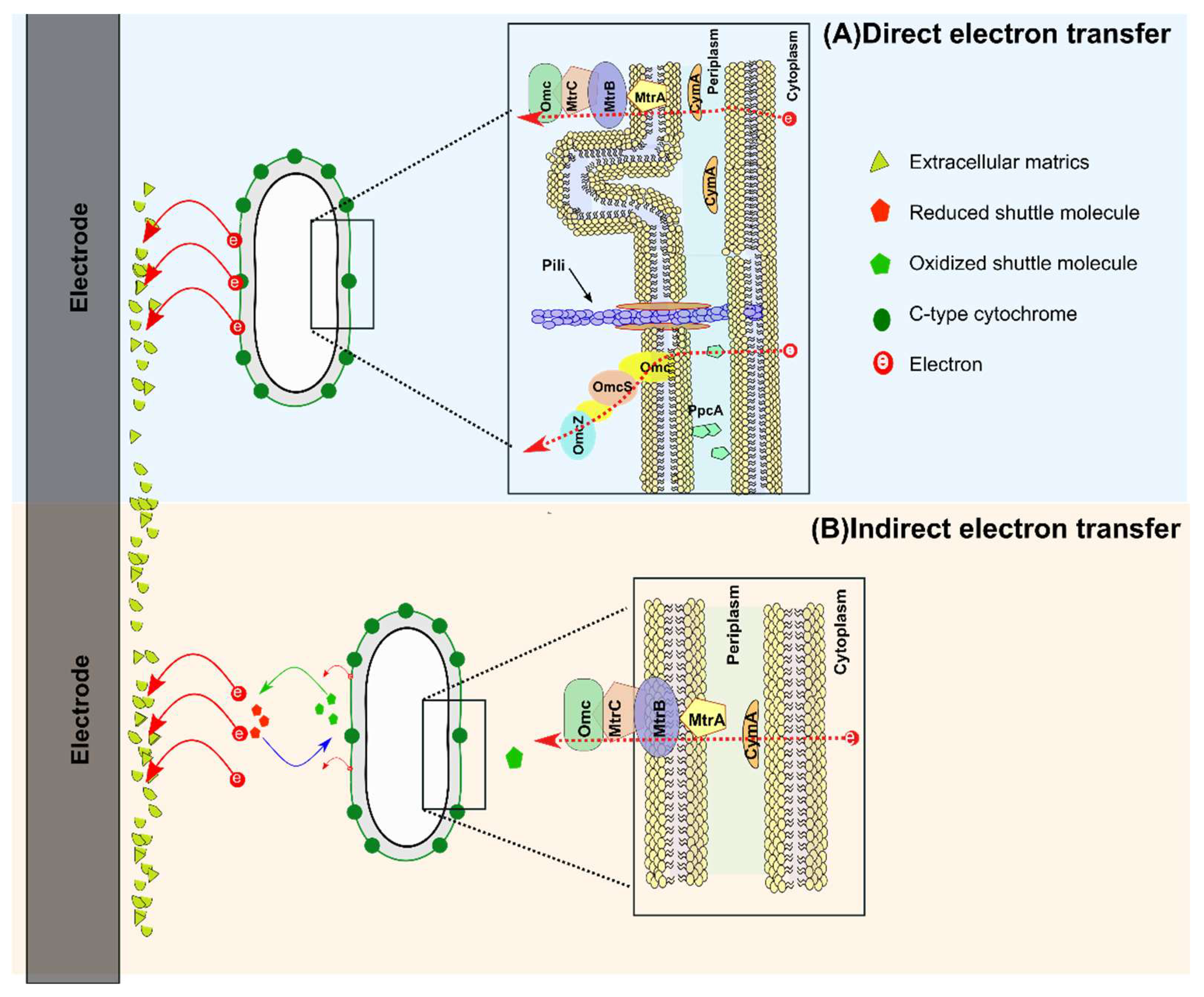

{kind=link}
{kind=link}
Table 1.
Electroactive bacteria and molecules involved in extracellular electron transfer (EET) within the electrode-microbe interfaces in bioelectrochemical systems (BESs).
Table 1.
Electroactive bacteria and molecules involved in extracellular electron transfer (EET) within the electrode-microbe interfaces in bioelectrochemical systems (BESs).
| Name of Bacteria | Electron Transfer Mechanism | Molecules Involved in Electron Transfer | Remarks | References |
|---|---|---|---|---|
| Geobacter sulfurreducens | DET (nanowires) | Branched OMCs system: cytochromes (c-, d-types) and Type IV pili | Oxidation of organic compounds to iron reduction leads to electron release | [35] |
| Thermincola ferriacetica | DET | c-type cytochromes | Electrons are transported to electrode through multiheme c-type cytochromes | [36] |
| Clostridium ljungdahlii | DET | Rnf complex (Ferredoxin:NAD+ -oxidoreductase) (No cytochromes, no quinones) | Electron bifurcating ferredoxin reduction | [37,38] |
| Pseudomonas aeruginosa | DET | cytochromes (a-, b-, c-, o-type), phenazines, flavines (soluble and bound), quinones and dehydrogenases | Electron transfer through the production of versatile phenazine redox mediators. | [39] |
| Geobacter metallireducens | DET | c-Type cytochromes, that is, OmcB and OmcE | Fe(III) oxide reduction | [40] |
| Shewanella putrefaciens | DET | c-Type cytochromes, that is, MtrC and OmcA, FAD transporter | NIF | [41] |
| Rhodopseudomonas Palustris | DET | c-Type cytochromes | NIF | [42] |
| Klebsiella pneumoniae | DET | 2,6-di-tert-butyl-p-benzoquinone | Reduction of iron oxide and generate electricity | [43] |
| Moorella thermoacetica | IET (methylviologen) | Cytochromes (b, d-type), quinones and/or Ech-complex | Reduction of CO2 to other organic compounds | [37] |
| Acetobacterium woodii | IET (biotic H2) | Rnf complex (Ferredoxin: NAD+ -fuctase), membrane bound corrinoids (No cytochromes, no quinones) | Electron bifurcating ferredoxin hydrogenase catalysis H2 formation | [37,44] |
| Lactococcus lactis | IET | 2-Amino-3-dicarboxy-1,4-naphthoquinone | NIF | [45] |
| Shewanella oneidensis | IET (self-produced shuttles),DET (nanowires) | Metal-reducing pathway components CymA, MtrA, MtrB, MtrC, and OmcA | CymA in inner-membrane oxidizes the quinol and transfers the released electrons to MtrA directly or indirectly | [46] |
| Sporomusa ovata | DET and IET | Membrane-bound cytochromes (b-, c-types) and quinones | CO2 is used as an electron acceptor and reduced to organic compounds via the Wood-Ljungdahl pathway | [37] |
| Shewanella oneidensis | IET (self-produced shuttles), DET (nanowires) | Flavins, riboflavin | NIF | [47] |
| Thermincola ferriacetica | NIF | Anthraquinone-2,6-disulfonate | Fe(III) reduction | [36] |
NIF: no information found.
Table 2.
Isolated electroactive bacteria from various bioelectrochemical systems.
| Isolated Electroactive Bacteria | Source of Isolation | Technique Used before Culturing | Reference |
|---|---|---|---|
| Dietzia sp. RNV-4 | Single Chamber MFC | Dilution | [53] |
| Geobacter OS1 | Petroleum hydrocarbon-contaminated soil | Dilution | [54] |
| Ochrobactrum OS2 | Petroleum hydrocarbon-contaminated soil | Dilution | [54] |
| Ochrobactrum anthropi YZ-1 | Single Chamber MFC | Dilution & Enrichment | [55] |
| Sphingomonas strain DJ | Microbial electrolysis cell (MECs) | Dilution | [56] |
| Rhodopseudomonas palustris DX-1 | Air cathode MFC | Dilution | [57] |
| Geobacter SD-1 | Microbial electrolysis cell (MECs) | Dilution & Enrichment | [58] |
| Citrobacter LAR-1 | Sediments | Dilution & Enrichment | [59] |
| Geobacter bremensis | Compost | Dilution & Enrichment | [60] |
| Bacillus sediminis DX-5T | Microbial electrolysis cell (MECs) | Dilution | [61] |
| Clostridium butyricum | Mediatorless MFC | Enrichment | [62] |
| Rhodoferax ferrireducens | Sediments | Enrichment | [63] |
| Enterococcus gallinarum MG25 | Submerged soil | Dilution & Enrichment | [64] |
| Aeromonas hydrophila | Mediatorless MFC | Enrichment | [65] |
Table 3.
Simultaneous wastewater treatment of various substrates, COD removal efficiency and power densities reported in the literature.
Table 3.
Simultaneous wastewater treatment of various substrates, COD removal efficiency and power densities reported in the literature.
| Type of Wastewater Treated | Inoculum Source | Hydraulic Retention Time | Initial COD | COD Removal Efficiency | Power Density | Reference |
|---|---|---|---|---|---|---|
| Cereal wastewater | Sludge | 120 h | 595 mg/L | 95% | 371 ± 10 mW/m2 | [101] |
| Dairy wastewater | Anaerobic mixed consortia as | NIF | 4.44 kgCOD/m3 | 95.49% | 1.10 W/m3 | [102] |
| Dioscorea zingiberensis wastewater | Anaerobic sludge | 72 h | 91 000 mg/L | 93.50% | 118.1 mW/m2 | [103] |
| Effluent from hydrogen producing biofermentor | Domestic wastewater | 23 h | 6.3 g/L | 97% | 4200 mW/m3 | [104] |
| Confectionary wastewater | Aerobic sludge, anaerobic sludge, | NIF | 22 000 mg/L | 92% | NIF | [105] |
| Biodiesel waste | Domestic wastewater | NIF | 1400 mg/L | 90% | 1310 ± 15 mW/m2 | [106] |
| Starch processing wastewater | Wastewater itself | four cycles of 140 days | 4852 mg/L | 98% | 239.4 mW/m2 | [107] |
| Paper wastewater | Wastewater itself | 500 h | 0.48 g/L | 76 ± 4% TCOD; | 501 ± 20 mW/m2 | [108] |
| Rice mill wastewater | Anaerobic sludge | 288 h | 1100–1125 mg/L | 96.5% COD | 2.3 W/m3 | [109] |
| Palm oil effluent | 3.5 g/L sludge | 48 h | 10000 mg/L | COD; 93.6% | NIF | [110] |
| Synthetic penicillin wastewater | Bacteria from another MFC | 24 h | 50 mg/L penicillin: 1000 mg/L glucose mix | 98% | 101.2 mW/m3 | [111] |
| Indole | Mixed aerobic and anaerobic activated | 12 h | 500 mg/L | Complete removal | 1410.2 mW/m2 | [112] |
| Quinoline | – | 6 h | 500 mg/L | Complete removal | 16.4 mW/m2 | [113] |
| Selenite wastewater | Mixed bacterial culture | 48 and 72 h | 50 and 200 mg/L | 99% | 2900 mW/m2 | [114] |
| Pyridine | Mixed aerobic and anaerobic activated | 12 h | 500 mg/L | Complete removal | 1410.2 mW/m2 | [114] |
| Ceftriaxone sodium (Cs). | Bacteria from another glucose-fed mFC | 24 h | 50 mg/L (Cs): 1000 mg/L glucose | 96% | 11 mW/m2 | [111] |
| p-Nitrophenol wastewater | Anaerobic sludge | 12 h | NIF | Complete degradation | 143 mW/m2 | [115] |
| Refractory contaminants (Furfural) | Anaerobic and aerobic sludge | 60 h | 300 mg/L | 96% COD; 100% furfural | 15.9 W/m3 | [116] |
| Dairy wastewater | self-microbial population of | 10 days | 1487 mg/L | 81.29% | 10.89 mA | [117] |
| Carbonaceous and nitrogenous pollutants | digested sludge | 210 days | 545 ± 43 mg/L | 99% COD; <20% | by 50% 0.25 ± 0.07 V | [118] |
| CO | anaerobic sludge | 14 days | NIF | NIF | 260.3 mV | [119] |
| 1,2-Dichloro-ethane waste water | mixed natural consortium from a 1,2-DCA contaminated site | 1 month | 99 mg/L | 85% | 0.03 mA | [120] |
| Chemical wastewater | anaerobic mixed consortia | 96 h | 5900 mg/L | 58.98% | 186.34 mA/m2 | [121] |
| Landfill | anaerobic | NIF | 9800 mg/L | NIF | 6817.4 mW/m3 | [122] |
| 4-Chlorophenol | anaerobic sludge | 45 h | 60 mg/L | complete | 12.4 mW/m2 | [123] |
| Amaranth Dye | NIF | 1 h | 75 mg/L | 82.59% | 28.3W/m3 | [124] |
| Cr(VI) | NIF | 10 h | 177 mg/L | 92.80% | 108 mW/m2 | [125] |
| Refinery waste | NIF | 6 days | NIF | NIF | 120 mW/m2 | [126] |
| Terephthalic acid | anaerobic sludge | 210 h | 4000 mg/L | 80.30% | 96.3 mW/m2 | [127] |
| Diesel range organics | diesel contaminated groundwater | 21 days | 300 mg/L | 82% | 31 mW/m2 | [128] |
| Brewery wastewater | anaerobic mixed consortia | 2.13 h | 1501 mg/L | 47.60% | 669 mW/m2 | [129] |
| Starch processing wastewater | electrochemically active bacteria | 6 weeks | 1700 mg/L | 97% | 0.44 mA/cm2 | [130] |
NIF: no information found.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ali, J.; Sohail, A.; Wang, L.; Rizwan Haider, M.; Mulk, S.; Pan, G. Electro-Microbiology as a Promising Approach Towards Renewable Energy and Environmental Sustainability. Energies 2018, 11, 1822. https://doi.org/10.3390/en11071822
AMA Style
Ali J, Sohail A, Wang L, Rizwan Haider M, Mulk S, Pan G. Electro-Microbiology as a Promising Approach Towards Renewable Energy and Environmental Sustainability. Energies. 2018; 11(7):1822. https://doi.org/10.3390/en11071822
Chicago/Turabian StyleAli, Jafar, Aaqib Sohail, Lei Wang, Muhammad Rizwan Haider, Shahi Mulk, and Gang Pan. 2018. "Electro-Microbiology as a Promising Approach Towards Renewable Energy and Environmental Sustainability" Energies 11, no. 7: 1822. https://doi.org/10.3390/en11071822
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.