Characterization of Intracellular and Extracellular Saxitoxin Levels in Both Field and Cultured Alexandrium spp. Samples from Sequim Bay, Washington


Abstract
:1. Introduction
2. Materials and Methods
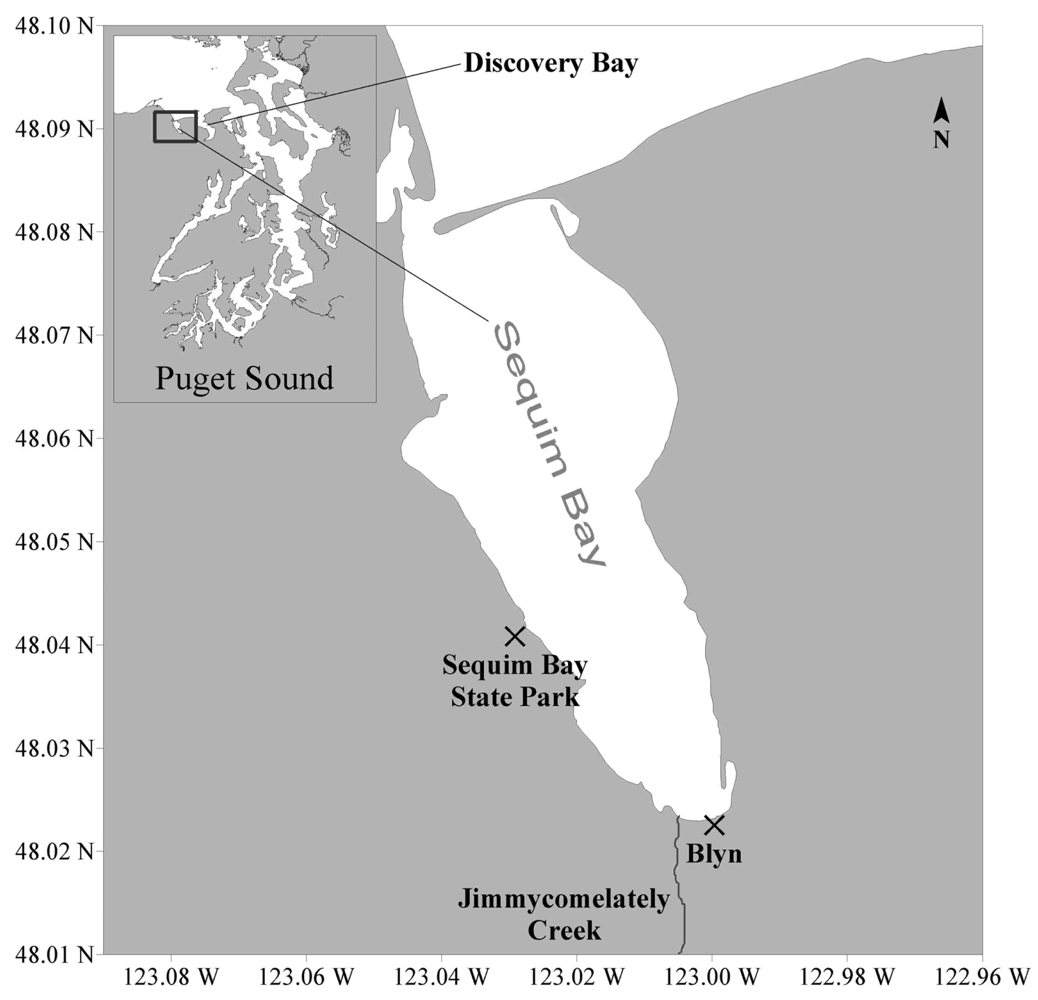
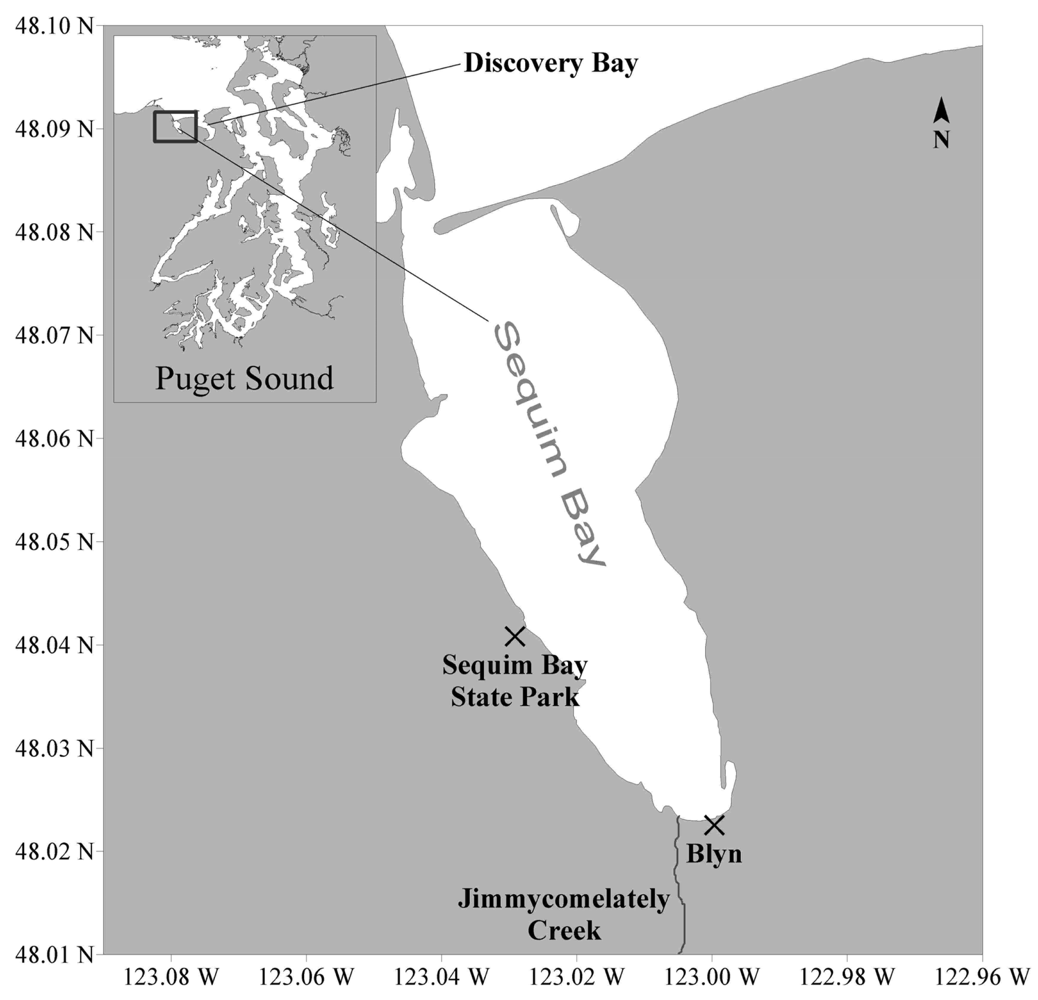
2.1 Collection of Sequim Bay seawater samples
2.2 Toxin and cell quantification
2.3 Isolation and analysis of laboratory culture samples for growth studies
2.4 Quality control experiments for extraction and filtering techniques
2.5 Characterization of toxin profiles in Sequim Bay Alexandrium spp. isolates
3. Results
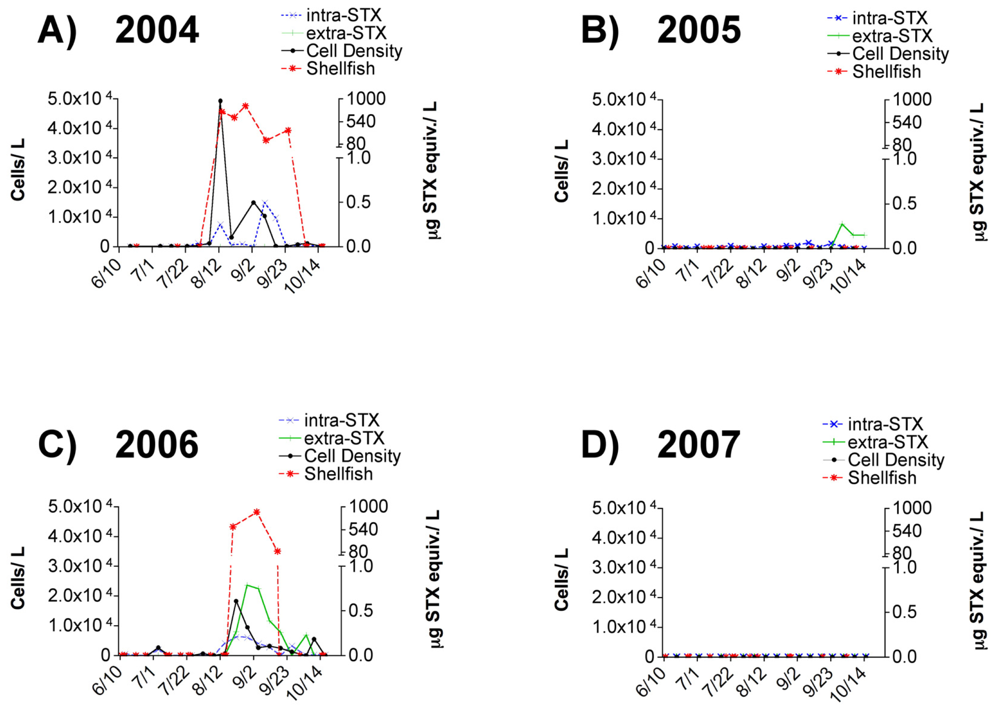
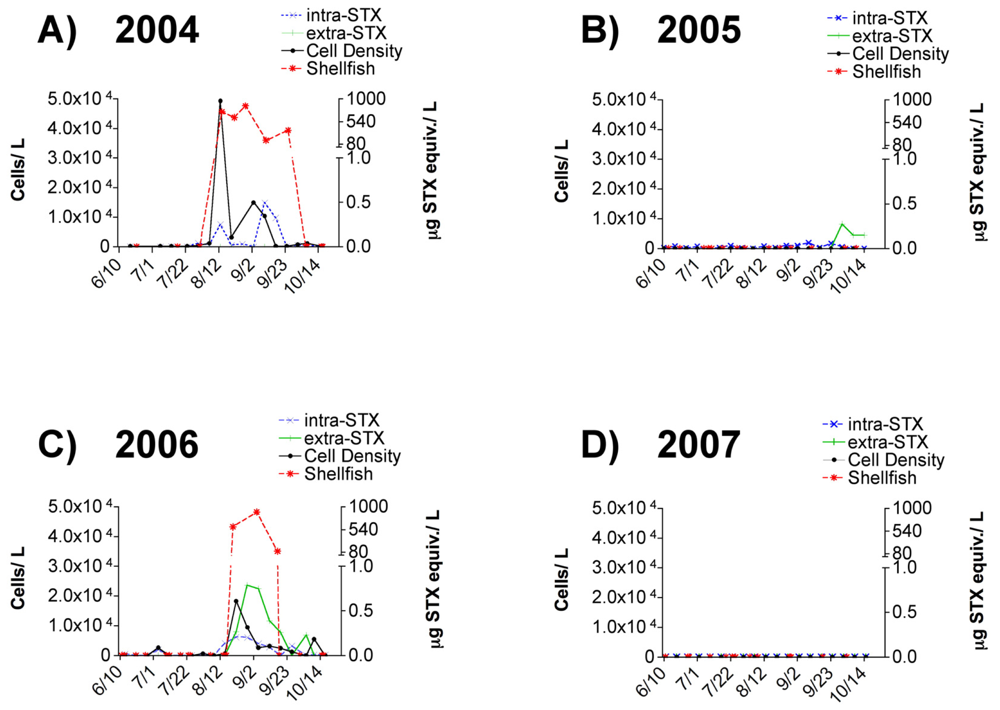
3.1 Alexandrium spp. cell densities and toxin presence in field-collected seawater
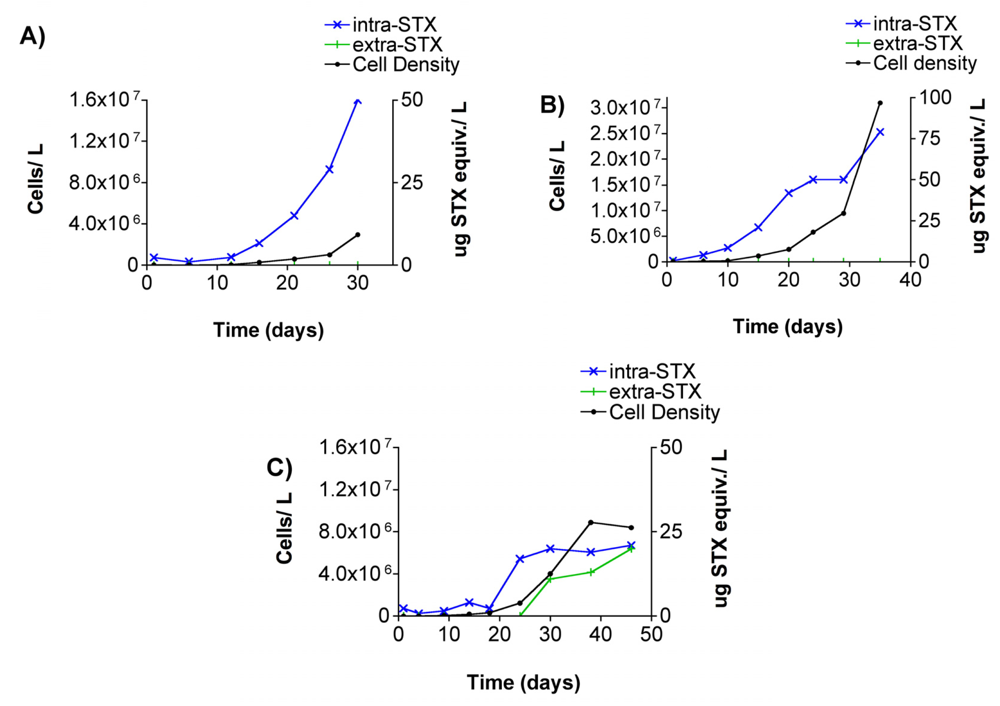
3.2 Alexandrium spp., cell densities and toxin presence in culture growth studies
3.3 Results of quality control experiments for extraction and filtration procedures
3.4 Toxin profiles in Sequim Bay Alexandrium spp. isolates
4. Discussion and Conclusions
4.1 Fisheries and PSTs in Sequim Bay
4.2 Dissolved PST levels in field-collected seawater and culture media samples
4.3 Effects of extracellular PST exposure on fish larvae
4.4 Frequency of PSP blooms and importance of monitoring in Sequim Bay
4.5 Summary
Acknowledgments
References
- Horner, RA; Garrison, DL; Plumley, FG. Harmful algal blooms and red tide problems on the U.S. west coast. Limnol and Oceanogr 1997, 42, 1076–1088. [Google Scholar]
- Waldichuk, M. Shellfish toxicity and the weather in the Strait of Georgia during 1957. J Fish Res Board Can, Prog Rep Pacific Coast Stations 1958, 112, 10–14. [Google Scholar]
- Trainer, VL; Eberhart, B-TL; Wekell, JC; Adams, NG; Hanson, L; Cox, F; Dowell, J. Paralytic shellfish toxins in Puget Sound, Washington State. J Shellfish Res 2003, 22, 213–222. [Google Scholar]
- Determan, T. Puget Sound Research ’98 Proceedings; Puget Sound Water Quality Action Team: Seattle, WA, 1998; pp. 627–633. [Google Scholar]
- Moore, SK; Mantua, NJ; Trainer, VL; Hickey, BM. Long-term measurements of paralytic shellfish toxins in Puget Sound shellfish: The influence of climate and capacity for prediction. Mar Ecol Prog Ser. submitted.
- Rudeen, SK. In the “undevelopment” of Jimmycomelately creek. Wetland Venturer. 2003, 5. http://pcjv.org/newsletter_Wetland_Ventures/wetland_ventures_v5no3.pdf.
- Lefebvre, KA; Trainer, VL; Scholz, NL. Morphological abnormalities and sensorimotor deficits in larval fish exposed to dissolved saxitoxin. Aquat Toxicol 2004, 66, 159–170. [Google Scholar]
- Lefebvre, KA; Elder, NE; Hershberger, PK; Trainer, VL; Stehr, CM; Scholz, NL. Dissolved saxitoxin causes transient inhibition of sensorimotor function in larval Pacific herring (Clupea harengus pallasi). Mar Biol 2005, 147, 1393–1402. [Google Scholar]
- Velzeboer, RMA; Bater, PD; Rositano, J. Saxitoxins associated with the growth of the cyanobacterium Anabaena circinalis (Nostocales, Cyanophyta) under varying sources and concentrations of nitrogen. Phycologia 2001, 40, 305–312. [Google Scholar]
- Robertson, A; Negri, AP; Burnell, JN; Llewellyn, LE. Development and assessment of radioreceptor binding assays for the detection of saxitoxin binding proteins in biological extracts. Anal Biochem 2006, 356, 66–75. [Google Scholar]
- Trainer, V; Poli, M. Rochat, H, Marein-Eauclaire, MF, Eds.; Assays for Dinoflagellate Toxins, Specifically Brevetoxin, Ciguatoxin, and Saxitoxin. In Animal Toxins; Birkhäuser Verlag: Basel, 2000; pp. 1–19. [Google Scholar]
- Guillard, RRL. Culture of Marine Invertebrate Animals; Plenum Press: New York, NY, 1975; pp. 26–60. [Google Scholar]
- Ravn, H; Anthoni, U; Christophersen, C; Nielsen, PH; Oshima, Y. Standardized extraction method for paralytic shellfish toxins in phytoplankton. J Appl Phycol 1995, 7, 589–594. [Google Scholar]
- Lawrence, JF; Niedzwiadek, B; Menard, C. Quantitative determination of paralytic shellfish poisoning toxins in shellfish using prechromatographic oxidation and liquid chromatography with fluorescence detection: collaborative study. J AOAC Int 2005, 88, 1714–1732. [Google Scholar]
- Hsieh, D; Huxtable, S; Chang, G. Hallegraeff, G, Ed.; Total production of C1/C2 by Alexandrium tamarense. In Proceedings from the Ninth International Conference on Harmful Algal Blooms; University of Tasmania: Hobart, Tasmania, 2000. [Google Scholar]
- Llewellyn, LE; Doyle, J; Jellett, J; Barrett, R; Alison, C; Bentz, C; Quilliam, MA. Measurement of paralytic shellfish toxins in molluscan extracts: Comparison of the microtitre plate saxiphilin and sodium channel radioreceptor assays with mouse bioassay, HPLC analysis and a commercially available cell culture assay. Food Addit Contam 2001, 18, 970–980. [Google Scholar]
- Mortensen, AM. Anderson, DM, White, AW, Baden, DG, Eds.; Massive fish mortalities in the Faroe Islands caused by Gonyaulax excavata red tide. In Toxic Dinoflagellates; Elsevier: New York, 1985; pp. 165–170. [Google Scholar]
- Yazdandoust, MH. Anderson, DM, White, AW, Baden, DG, Eds.; Cancer crab larvae and goby fish: vector and victim of paralytic shellfish poisons (PSP). In Toxic Dinoflagellates; Elsevier: New York, 1985; pp. 419–424. [Google Scholar]
- Gosselin, S; Fortier, L; Gagne, JA. Vulnerability of marine fish larvae to the toxic dinoflagellate Protogonyaulax tamarensis. Mar Ecol Pro Ser 1989, 57, 1–10. [Google Scholar]
- White, AW; Fukuhara, O; Anraku, M. Okaichi, T, Anderson, DM, Nemoto, T, Eds.; Mortality of fish larvae from eating toxic dinoflagellates or zooplankton containing dinoflagellate toxins. In Red Tides: Biology, Environmental Science and Toxicology; Elsevier: New York, 1989; pp. 395–398. [Google Scholar]
- Mills, LJ; Klein-MacPhee, GK. Taylor, KL, Seliger, HH, Eds.; Toxicity of the New England red tide dinoflagellate to winter flounder larvae. In Toxic Dinoflagellate Blooms; Elsevier: New York, 1979; pp. 389–394. [Google Scholar]
- White, AW. Ragelis, EP, Ed.; Paralytic shellfish toxins and finfish. In Seafood Toxins; American Chemical Society: Washington, DC, 1984. [Google Scholar]
- White, AW. The scope of impact of toxic dinoflagellate blooms on finfish in Canada. Can Tech Rep Fish Aquat Sci 1982, 1063, 1–5. [Google Scholar]
- Oberemm, A; Becker, J; Codd, GA; Steinberg, C. Effects of cyanobacterial toxins and aqueous crude extracts of cyanobacteria on the development of fish and amphibians. Environ Toxicology 1999, 14, 77–88. [Google Scholar]
- Nishitani, L; Chew, KK. Recent developments in paralytic shellfish poisoning research. Aquaculture 1984, 39, 317–329. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | A. dH2O | B. 0.1 M acetic acid | C. dH2O/HCl | D. 0.1 M acetic acid/HCl |
|---|---|---|---|---|
| 1 | 154 | 131 | 121 | 145 |
| 2 | 64 | 89 | 75 | 95 |
| 3 | 81 | 121 | 84 | 105 |
| 4 | 66 | 66 | 49 | 69 |
| 5 | 95 | 100 | 87 | 111 |
| 6 | 107 | 98 | 108 | 116 |
| 7 | 31 | 36 | 35 | 49 |
| 8 | 69 | 63 | 48 | 65 |
| sample | Alexandrium cells/L | normal vacuum extracellular STX | light vacuum extracellular STX |
|---|---|---|---|
| 1 | 3.5 x 106 | 30 | 28 |
| 2 | 2.3 x 106 | 24 | 27 |
| 3 | 2.2 x 106 | 26 | 30 |
| 4 | 1.8 x 106 | 23 | 31 |
| 5 | 2.9 x 106 | 24 | 24 |
| Sample | Cells/L | Intracellular STX (RBA) | Intracellular STX (ELISA) | Extracellular STX (RBA) | Extracellular STX (ELISA) |
|---|---|---|---|---|---|
| SQ-1 | 6.2 X 106 | 61 | 1.0 | 14 | 2.9 |
| SQ-2 | 6.8 X 105 | 17 | 3.0 | 15 | 2.0 |
| SQ-3 | 2.2 X 106 | 127 | 6.8 | 29 | 3.4 |
| SQ-4 | 3.3 X 106 | 96 | 2.1 | 12 | 1.4 |
Share and Cite
Lefebvre, K.A.; Bill, B.D.; Erickson, A.; Baugh, K.A.; O’Rourke, L.; Costa, P.R.; Nance, S.; Trainer, V.L. Characterization of Intracellular and Extracellular Saxitoxin Levels in Both Field and Cultured Alexandrium spp. Samples from Sequim Bay, Washington. Mar. Drugs 2008, 6, 103-116. https://doi.org/10.3390/md6020103
Lefebvre KA, Bill BD, Erickson A, Baugh KA, O’Rourke L, Costa PR, Nance S, Trainer VL. Characterization of Intracellular and Extracellular Saxitoxin Levels in Both Field and Cultured Alexandrium spp. Samples from Sequim Bay, Washington. Marine Drugs. 2008; 6(2):103-116. https://doi.org/10.3390/md6020103
Chicago/Turabian StyleLefebvre, Kathi A., Brian D. Bill, Aleta Erickson, Keri A. Baugh, Lohna O’Rourke, Pedro R. Costa, Shelly Nance, and Vera L. Trainer. 2008. "Characterization of Intracellular and Extracellular Saxitoxin Levels in Both Field and Cultured Alexandrium spp. Samples from Sequim Bay, Washington" Marine Drugs 6, no. 2: 103-116. https://doi.org/10.3390/md6020103