Report on the First Detection of Pectenotoxin-2, Spirolide-A and Their Derivatives in French Shellfish



Abstract
:Introduction
Materials and methods
Reference materials
Shellfish samples
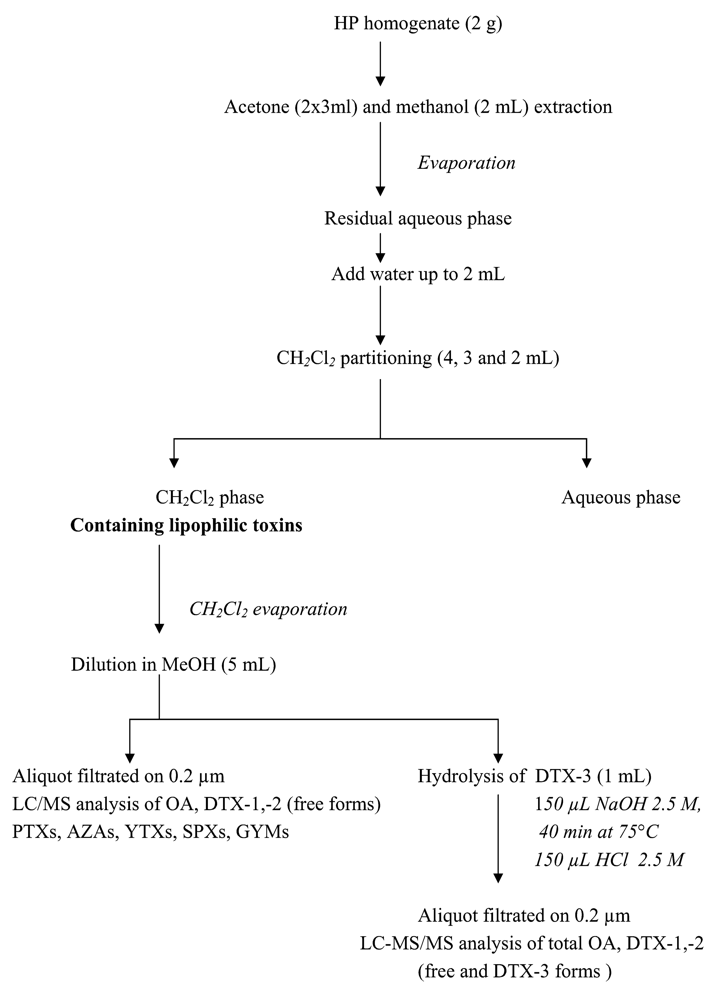
Extraction procedure for lipophilic toxins
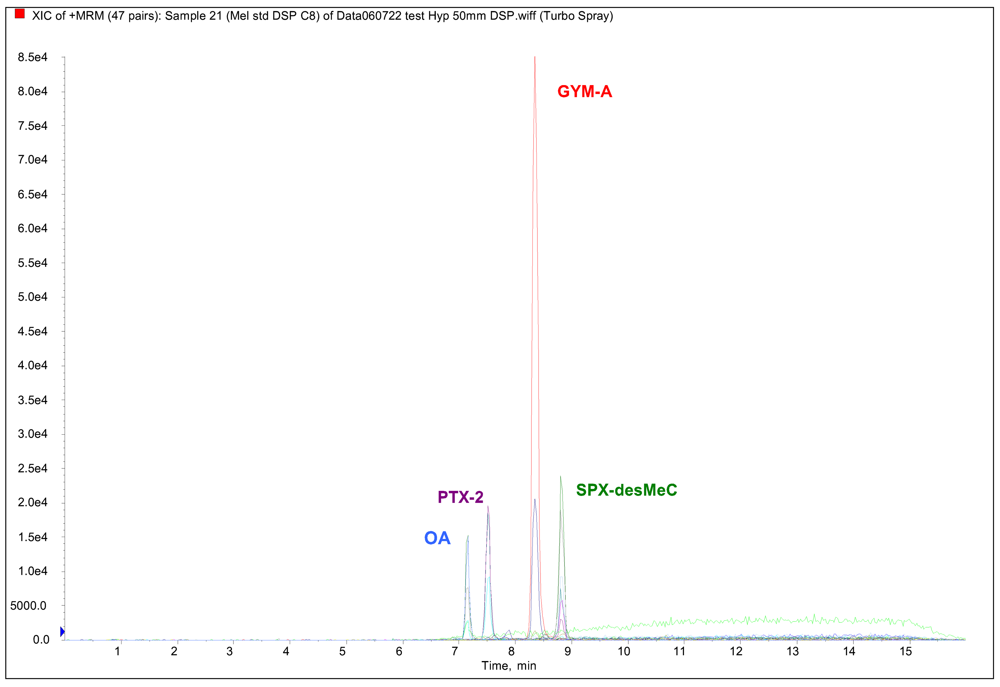
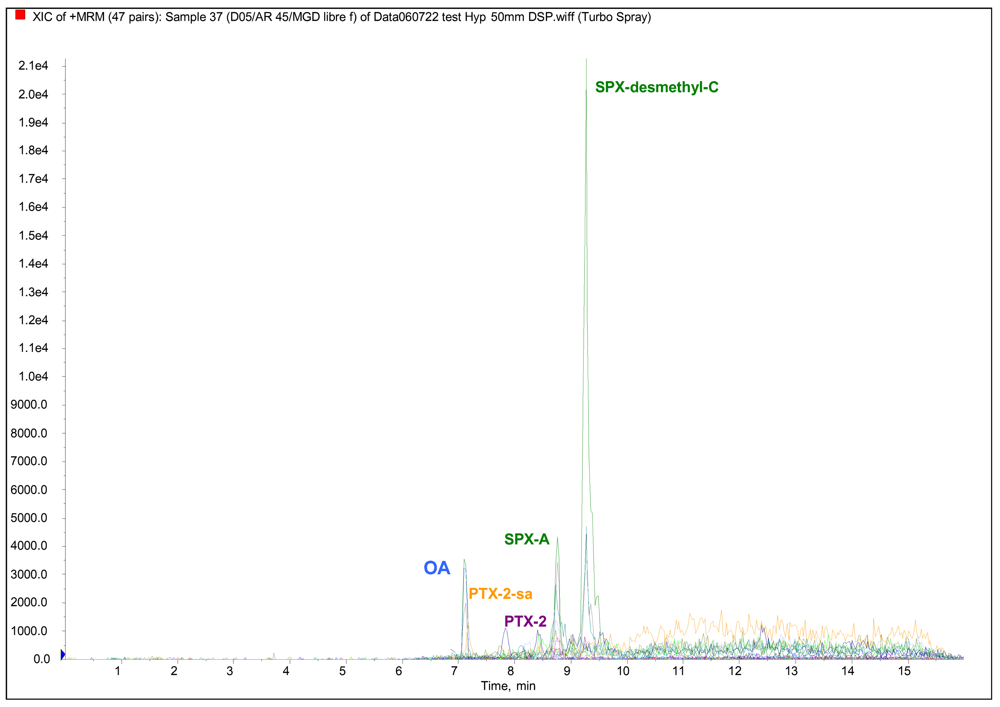
LC/MS-MS analyses of lipophilic toxins
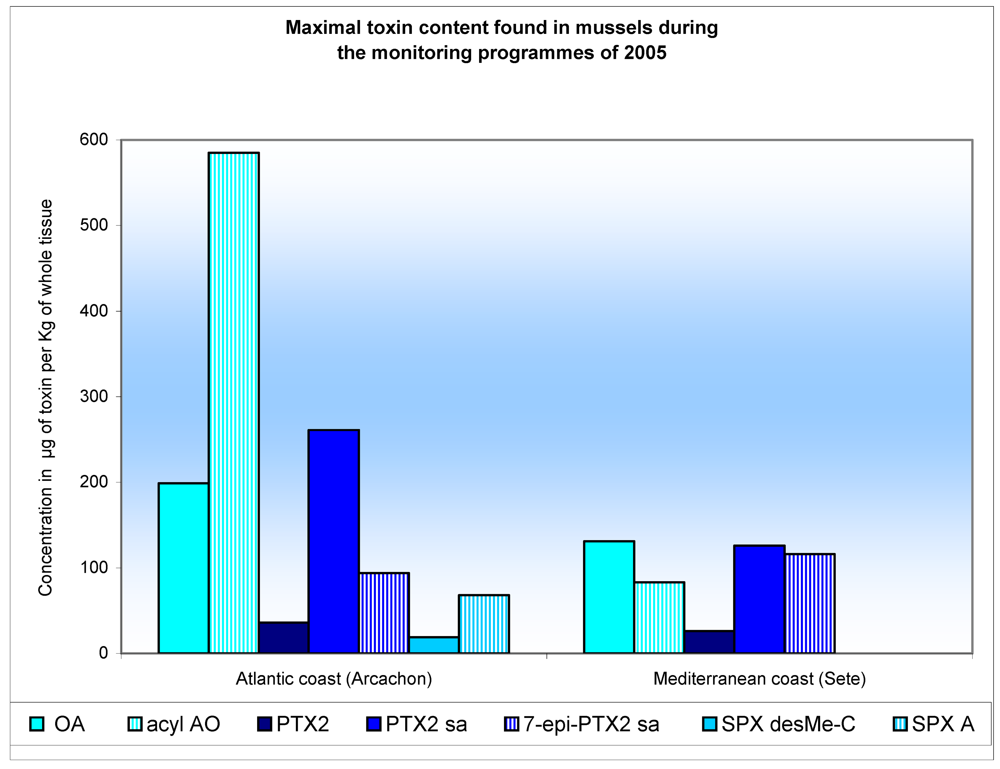
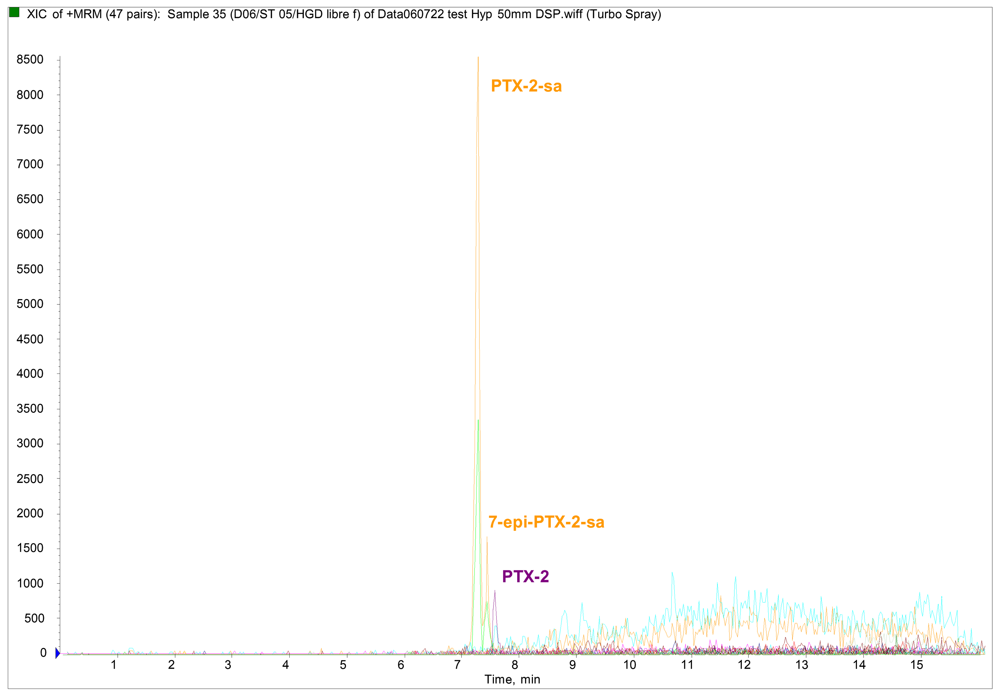
Results and Discussion
Conclusions
Acknowledgements
- Sample Availability: Contact the authors.
References
- Murata, M; Shimatani, M; Sugitani, H; Oshima, Y; Yasumoto, T. Isolation and structural elucidation of the causative toxin of diarrhetic shellfish poisoning. Bull Jap Soc Fish 1982, 48, 549–552. [Google Scholar]
- Kumagai, M; Yanagi, T; Murata, M; Yasumoto, T; Kat, M; Lassus, P; Rodriguez-Vasquez, JA. Okadaic acid as causative toxin of diarrhetic shellfish poisoning in Europe. Agric Biol Chem 1986, 50, 2853–2857. [Google Scholar]
- Hu, T; Doyle, J; Jackson, D; Marr, J; Nixon, E; Pleasance, S; Quilliam, MA; Walter, JA; Writh, JLC. Isolation of a new diarrhetic shellfish poison from Irish mussels. Chem Commun 1992, 39–41. [Google Scholar]
- Marr, JC; Hu, T; Pleasance, S; Quilliam, MA; Wright, JLC. Detection of new 7-O-acyl derivatives of diarrhetic shellfish poisoning toxins by liquid chromatography-mass spectrometry. Toxicon 1992, 30, 1621–1630. [Google Scholar]
- Vale, P; Antonia, M; Sampayo, M. Esters of okadaic acid related to human poisonings. Toxicon 1999, 37, 1109–1121. [Google Scholar]
- Suzuki, T; Ota, H; Yamasaki, M. Direct evidence of transformation of dinophysistoxin-1 to 7-O-acyl-dinophysistoxin-1 (dinophysistoxin-3) in the scallop Patinopecten yessoensis. Toxicon 1999, 37, 187–198. [Google Scholar]
- Yasumoto, T; Murata, M; Oshima, Y; Sano, M; Matsumoto, GK; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar]
- Murata, M; Kumagai, M; Lee, JS; Yasumoto, T. Isolation and structure of yessotoxin, a polyether compound implicated in diarrhetic shellfish poisoning. Tetrahedron Lett 1987, 28, 5869–5872. [Google Scholar]
- Satake, M; Mackenzie, L; Yasumoto, T. Identification of Protoceratium reticulatum as the biogenetic origin of yessotoxin. Natural Toxins 1997, 5, 164–167. [Google Scholar]
- Ofuji, K; Satake, M; McMahon, T; James, KJ; Naoki, H; Oshima, Y; Yasumoto, T. Two analogs of azaspiracids isolated from mussels, Mytilus edulis, involved in human intoxication in Ireland. Natural Toxins 1999, 7, 99–102. [Google Scholar]
- James, KJ; Furey, A; Lehane, M; Ramstad, H; Aune, T; Hovgaard, P; Morris, S; Higman, W; Satake, M; Yasumoto, T. First evidence of an extensive northern European distribution of azaspiracid poisoning (AZP) toxins in shellfish. Toxicon 2002, 40, 909–915. [Google Scholar]
- Yasumoto, T; Murata, M; Oshima, Y; Matsumoto, GK; Clardy, J. Ragelis, EP, Ed.; Diarrhetic Shellfish Poisoning. In Sea Food Toxins; ACS Symposium Series: A.C.S: Washington DC, 1984; Volume 262, pp. 208–214. [Google Scholar]
- Hu, T; Curtis, JM; Oshima, Y; Quilliam, MA; Walter, JA; Watson-Wright, WM; Wright, JLC. Spirolides B and D, two novel macrocycles isolated from the digestive glands of shellfish. J Chem Soc, Chem Commun 1995, 2159–2161. [Google Scholar]
- Cembella, AD; Quilliam, MA; Lewis, NI; Bauder, AG; Wright, JLC. Reguera, B, Blanco, J, Fernandez, ML, Wyatt, T, Eds.; Identifying the plankton origin and distribution of spirolides in coastal Nova Scotian waters. In Harmful Algae; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO, 1998; pp. 481–484. [Google Scholar]
- Cembella, AD; Lewis, NI; Quilliam, MA. The marine dinoflagellate Alexandrium ostenfeldii (Dinophyceae) as the causative organism of spirolides shellfish toxins. Phycologia 2000, 39, 67–74. [Google Scholar]
- Aasen, J; Mackinnon, SL; Le Blanc, P; Walter, JA; Hovgaard, P; Aune, T; Quilliam, MA. Detection and identification of spirolides in Norwegian shellfish and plankton. Chem Res Toxicol 2005, 18, 509–515. [Google Scholar]
- Villar Gonzalez, A; Rodriguez-Valasco, ML; Ben-Gigirey, B; Botana, LM. First evidence of spirolides in Spanish shellfish. Toxicon 2006, 48, 1068–1074. [Google Scholar]
- Ciminiello, P; Dell’Aversano, C; Fattorusso, E; Magno, S; Tartaglione, L; Cangini, M; Pompei, M; Guerrini, F; Boni, L; Pistochi, R. Toxin profile of Alexandrium ostenfeldii (Dinophyceae) from the Northern Adriatic sea revealed by liquid chromatography-mass spectrometry. Toxicon 2006, 47, 597–604. [Google Scholar]
- MacKenzie, L; Haywood, A; Adamson, J; Truman, P; Till, D; Deki, T; Satake, M; Yasumoto, T. Yasumoto, T, Oshima, Y, Fukuyo, Y, Eds.; Gymnodimine contamination of shellfish in New Zealand. In Harmful and toxic algal blooms; IOC of UNESCO Publish: Paris, 1996; pp. 97–100. [Google Scholar]
- McKenzie, L; Beuzenberg, V; McNabb, P. Steidinger, KA, Landsberg, CR, Vargo, GA, Eds.; Production of gymnodimine by Karenia selliformis. (Haywood et al). In Harmful Algae 2002; Florida Fish and Wildlife Conservation Commission, Florida Institute of Aceanography, and IOC of UNESCO Publish: Paris, 2004; pp. 160–162. [Google Scholar]
- Biré, R; Krys, S; Frémy, JM; Dragacci, S; Stirling, D; Kharrat, R. First evidence on occurrence of gymnodimine in clams from Tunisia. J Nat Toxins 2002, 11, 269–275. [Google Scholar]
- Lassus, P; Bardouil, M; Berthome, JP; Maggi, P; Truquet, P; Le Dean, L. Seasonal occurrence of Dinophysis sp. along the French coast between 1983 and 1987. Aquat Living Ressour 1988, 1, 155–164. [Google Scholar]
- Lassus, P; LeDoux, M; Bardouil, M; Bohec, M; Erard-LeDenn, E. Kinetics of Alexandrium minitum Halim toxin accumulation in mussels and clams. Natural Toxins 1994, 2, 329–333. [Google Scholar]
- Masselin, P; Lassus, P; Bardouil, M. High performance liquid chromatography analysis of diarrhoeic toxins in Dinophysis spp. from the French coast. J Appl Phycol 1992, 4, 385–389. [Google Scholar]
- Masselin, P; Amzil, Z; Abadie, E; Nézan, E; Le Bec, C; Chiantella, C; Truquet, P. Hallegraeff, GM, Blackburn, SI, Bolch, CJ, Lewis, RJ, Eds.; Paralytic shellfish poisoning on the French Mediterranean coast in the autumn 1998: Alexandrium tamarense complex (Dinophyceae) as causative agent. In Harmful Algal Blooms; JOC of UNESCO Publish: Paris, 2001; pp. 407–410. [Google Scholar]
- Amzil, Z; Fresnel, J; Le Gal, D; Billard, C. Domoic acid accumulation in french shellfish in relation to toxic species of Pseudonitzschia multiseries and P. pseudodelicatissima. Toxicon 2001, 39, 1245–1251. [Google Scholar]
- Amzil, Z; Mathias, A. Henshilwood, K, Deegan, B, McMahon, T, Cusak, C, Keaveney, S, Silke, J, O’ Cinneide, M, Lyons, D, Hess, P, Eds.; First report on detection of okadaic acid 7-O-acyl-ester derivatives (DTX-3) in French shellfish. In Molluscan Shellfish Safety; The Marine Institute: Rinville, Oranmore, Galway, Ireland, 2006; pp. 150–161. [Google Scholar]
- Quilliam, MA; Hess, P; Dell’Aversano, C. De Koe, WJ, Sampson, RA, Van Egmond, HP, Gilbert, J, Sabino, M, Eds.; Recent developments in the analysis of phycotoxins by liquid chromatography-mass spectrometry. In Mycotoxins and Phycotoxins in Perspective at the Turn of the Millenium; DeKoe W.J., Wageningen: The Netherlands, 2001; pp. 383–391. [Google Scholar]
- Mondeguer, F; Genin, E; Mestre, G; Nsengiyumva, C. Henshilwood, K, Deegan, B, McMahon, T, Cusak, C, Keaveney, S, Silke, J, O’, Cinneide M, Lyons, D, Hess, P, Eds.; Validation of an automated procedure of solid phase extraction of mussels digestive glands for identification and quantification of Dinophysistoxins in LC/ESI/MS2 by quadruple ion trap. In Molluscan Shellfish Safety; The Marine Institute: Rinville, Oranmore, Galway, Ireland, 2006; pp. 94–99. [Google Scholar]
- Miles, CO; Wilkins, AL; Munday, R; Dines, MH; Hawkes, AD; Briggs, LR; Sandvik, M; Jensen, DJ; Cooney, JM; Holland, P; Quilliam, MA; MacKenzie, AL; Beuzenberg, V; Towers, NR. Isolation of pectenotoxin-2 from Dinophysis acuta and its conversion to pectenotoxin-2 seco acid, and preliminary assessment of their acute toxicities. Toxicon 2004, 43, 1–9. [Google Scholar]
- Suzuki, T; MacKenzie, L; Stirling, D; Adamson, J. Pectenotoxin-2 seco acid: a toxin converted from pectenotoxin-2 by the New Zealand Greenshell mussel, Pena canaliculus. Toxicon 2001, 39, 207–514. [Google Scholar]
- McKenzie, L; Beuzenberg, V; Holland, P; McNabb, P; Suzuki, T; Selwood, A. Pectenotoxin and okadaic acid-based toxin profiles in Dinophysis acuta and Dinophysis acuminata from New Zealand. Harmful Algae 2005, 4, 75–85. [Google Scholar]
- Vrancic, MP; Mestrovic, V; Marasovic, I; Gillman, M; Furey, A; James, KK. The occurrence of 7-epipectenotoxin-2 seco acid in the coastal waters of the central Adriatic (Kastela Bay). Toxicon 2001, 39, 771–779. [Google Scholar]
- Vale, P; De Sampayo, MA. Pectenotoxin-2 seco acid, 7-epipectenotoxin-2 seco acid and pectenotoxin-2 in shellfish and plankton from Portugal. Toxicon 2002, 40, 972–987. [Google Scholar]
- Blanco, J; Alvarez, G; Uribe, E. Identification of pectenotoxins in plankton, filter feeders, and isolated cells of a Dinophysis acuminata with an atypical toxin profile, from Chile. Toxicon 2007, 49 , 710–716. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| South Brittany | Atlantic coast (Arcachon bay) | Mediterranean coast (Thau Lagoon) | ||||
|---|---|---|---|---|---|---|
| Lipophilic toxins | Clams | Mussels | Oysters | Mussels | Oysters | Mussels |
| OA | 37 | 86 | DLa | 199 | DLa | 131 |
| Acyl-AO (DTX3) | 554 | 231 | 180 | 585 | 46 | 83 |
| DTX2 | 26 | DLa | DLa | 5 | DLa | DLa |
| Acyl-DTX-2 (DTX3) | 17 | DLa | DLa | 2 | DLa | DLa |
| PTX2 | 2 | DLb | 5 | 36 | 22 | 26 |
| PTX2-sa | 22 | DLb | 4 | 261 | 170 | 126 |
| 7-epi-PTX2-sa | 16 | DLb | 2 | 94 | 33 | 116 |
| SPX-desMeC | 8 | 14 | 47 | 19 | DLc | DLc |
| SPX-A | DLc | 7 | DLc | 68 | DLc | DLc |
| SPX-desMeD | 2 | 2 | DLc | DLc | DLc | DLc |
Share and Cite
Amzil, Z.; Sibat, M.; Royer, F.; Masson, N.; Abadie, E. Report on the First Detection of Pectenotoxin-2, Spirolide-A and Their Derivatives in French Shellfish. Mar. Drugs 2007, 5, 168-179. https://doi.org/10.3390/md504168
Amzil Z, Sibat M, Royer F, Masson N, Abadie E. Report on the First Detection of Pectenotoxin-2, Spirolide-A and Their Derivatives in French Shellfish. Marine Drugs. 2007; 5(4):168-179. https://doi.org/10.3390/md504168
Chicago/Turabian StyleAmzil, Zouher, Manoella Sibat, Florence Royer, Nadine Masson, and Eric Abadie. 2007. "Report on the First Detection of Pectenotoxin-2, Spirolide-A and Their Derivatives in French Shellfish" Marine Drugs 5, no. 4: 168-179. https://doi.org/10.3390/md504168