
Alexandrium spp.: From Toxicity to Potential Biotechnological Benefits
by
 , , , and
, , , and
Eleonora Montuori
1,2 ,
,
Daniele De Luca
3 ,
,
Antonella Penna
4 ,
,
Darta Stalberga
5 and
Chiara Lauritano
2,*
1
Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, Viale F. Stagno d’Alcontres 31, 98166 Messina, Italy
2
Department of Ecosustainable Marine Biotechnology, Stazione Zoologica Anton Dohrn, Via Acton 55, 80133 Napoli, Italy
3
Research Infrastructure for Marine Biological Resources Department, Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy
4
Department of Biomolecular Sciences, University of Urbino, Campus E. Mattei, 61029 Urbino, Italy
5
Department of Biomedical and Clinical Sciences, Division of Clinical Chemistry and Pharmacology, Linköping University, SE-58183 Linköping, Sweden
*
Author to whom correspondence should be addressed.
Mar. Drugs 2024, 22(1), 31; https://doi.org/10.3390/md22010031
Submission received: 10 November 2023
/
Revised: 27 December 2023
/
Accepted: 28 December 2023
/
Published: 30 December 2023
Abstract
:Many dinoflagellates of the genus Alexandrium are well known for being responsible for harmful algal blooms (HABs), producing potent toxins that cause damages to other marine organisms, aquaculture, fishery, tourism, as well as induce human intoxications and even death after consumption of contaminated shellfish or fish. In this review, we summarize potential bioprospecting associated to the genus Alexandrium, including which Alexandrium spp. produce metabolites with anticancer, antimicrobial, antiviral, as well as anti-Alzheimer applications. When available, we report their mechanisms of action and targets. We also discuss recent progress on the identification of secondary metabolites with biological properties favorable to human health and aquaculture. Altogether, this information highlights the importance of studying which culturing conditions induce the activation of enzymatic pathways responsible for the synthesis of bioactive metabolites. It also suggests considering and comparing clones collected in different locations for toxin monitoring and marine bioprospecting. This review can be of interest not only for the scientific community, but also for the entire population and industries.
1. Introduction
Dinoflagellates represent one of the most important groups of marine phytoplankton, with more than 1500 species in the marine environment [1] and a total of around 2400 species also considering freshwater habitats [2,3]. This high species richness is also reflected when encompassing diverse biochemical (pigments and toxins produced), metabolic (autotrophs, mixotrophs, and grazers) and ecological (free-living, symbionts, and parasitic) strategies [4,5,6,7]. Some species have been the object of in-depth investigations due to their importance in ecosystem stability and resilience (e.g., zooxanthellae in coral reefs) or production of toxins with relevant impact on human health and activities (e.g., aquaculture). Regarding the latter aspect, some dinoflagellate species are responsible for harmful algal blooms (HABs), producing potent toxins that cause human intoxications and even death after consumption of contaminated shellfish or fish [8]. Moreso, these toxins can cause damage to aquaculture and the fishery industry [9,10], impairment of tourism and recreational activities [11,12], alterations in marine trophic structures [13,14], and mortality events of marine mammals, fish, and birds [15,16]. The globally distributed genus Alexandrium Halim contains 33 accepted species [17], of which some are involved in the formation of HABs (algal grow out of control which have toxic or harmful effects on humans or marine organisms and may differ in terms of severity, diversity, and distribution [18,19,20]). As a consequence, this genus is well studied among dinoflagellates [20]. The taxonomy of dinoflagellates, thanks to morphological and molecular analyses, is continuously improving with occasional controversies. For instance, John et al. 2014 [21] assigned the Alexandrium species names as Group I, A. fundyense; Group II, A. mediterraneum; Group III, A. tamarense; Group IV, A. pacificum; and Group V, A. australiense.
Previous reviews on Alexandrium are available: Long et al. in 2021 focused on Alexandrium bioactive extracellular compounds (BECs) [22], Lewis et al. in 2018 reviewed global distribution of Alexandrium minutum with a focus on Northern Europe [23], Casabianca et al. in 2012 investigated the population genetic structure and connectivity of A. minutum in the Mediterranean Sea [24], while Bi et al. in 2019 focused on studies in dinoflagellates, summarizing Alexandrium papers on expressed sequence tags, transcriptomes and other molecular approaches [25]. Our review is the first to discuss Alexandrium potential also for marine bioprospecting. Considering the harmfulness of many Alexandrium species, several studies have focused on the identification of toxins, enzymatic pathways responsible of their synthesis [26], transcriptomics [26,27,28], metabolomics [29], as well as various detection methods, ranging from microscopic examinations to real-time PCR-based assays [30,31]. However, species which do not produce toxins have been shown to exert bioactivities [32]. Our review highlights which Alexandrium produces metabolites with possible biotechnological applications, such as with anticancer, antimicrobial, antiviral, as well as anti-Alzheimer activity. This review explores their mechanisms of action and targets, where such information is available.
Alexandrium cells are roughly spherical. They occur as solitary or in short chains in rapidly growing populations, and possess two flagella that allow vertical migrations in the water column [33]. The genus includes both autotrophic and mixotrophic species (see Lim et al. [34] and references therein), with the latter nutritional strategy triggered by specific life phases and conditions [18]; bioluminescence is also reported for some species [24]. Like other dinoflagellates, some Alexandrium species possess a two-phase life strategy (Figure 1): a planktonic phase characterized by motile cells at different reproductive stages, and a cystic life phase where cells lie dormant in the sediments [35,36]. The cyst phase is an adaptation that allows survival during unfavorable growing conditions and is at the basis of the formation of blooms when favorable conditions for life return. Such blooms occur seasonally and are influenced by abiotic (temperature, salinity, nutrient availability) and biotic (predation, viral infection) factors [36].
As other microalgae, Alexandrium species produce primary and secondary metabolites, but, among these, they may produce toxic substances [20,37]. Toxic Alexandrium species are summarized in Table 1. Saxitoxin (STX), a tetrahydropurine alkaloid also known as paralytic shellfish toxin (PST), is one of the toxins produced by the genus Alexandrium and it is known to be one of the most toxic and harmful substances to humans. Saxitoxin was isolated and characterized in 1966 by Schantz et al. from Alexandrium catenella [20,38]. The lethal dose corresponds to 1–4 mg/person, depending on the sex and physiological predisposition of the individual [18,39,40].
Saxitoxin is also known for its use as a chemical agent in war [67]. It is also present in the Schedule I annex list of toxic chemicals of the Chemical Weapons Convention available on the Organization for the Prohibition of Chemical Weapons Chemicals website (https://www.opcw.org/chemical-weapons-convention/annexes/annex-chemicals/schedule-1; accessed on 21 October 2023) [67] and this is a further confirmation of its dangerousness and potential use as a weapon.
The first record of paralytic shellfish poisoning (PSP) events dated back to the end of 18th century in British Columbia [68], but regular data on such events were available from the end of 1930′s. Evidence showed that the phenomenon was reported along the coasts worldwide and mostly matched the known distribution of Alexandrium species (Figure 2). The genus Alexandrium produces other bioactive lipophilic compounds, called phycotoxins, such as spirolides, gymnodimines, goniodomins. The only known species that produces spirolides is A. ostenfeldii which also produces, gymnodimines and goniodomins [69,70]. The macrocyclic polyketide Goniodomin was produced also by A. hiranoi [71], A. monilatum [53], A. pseudogonyaulax [61], and A. taylorii [22,66].
2. Alexandrium Secondary Metabolites and Possible Biotechnological Applications
As mentioned in the previous paragraph, Alexandrium spp. can produce large quantities of varieties of molecules, including toxins such as saxitoxin, cyclic imine toxins, goniodomines and tetrodotoxin. Each toxin may have a similar structural variant, and each analogue may have different levels of toxicity and biological activity. Therefore, it is essential to continue the search and characterization of new analogues of these toxins for the assessment of their toxic effects and possible food safety measures to be taken into account for the prevention of intoxication by their ingestion. We report below the characteristics of the main toxins actually known and summarize the Alexandrium compound and extract bioactivities. Figure 3 summarizes the available chemical structures of Alexandrium compounds reported to exert possible bioactivities useful for human applications.
2.1. Saxitoxin
2.1.1. Chemistry
Saxitoxin (STX) is a compound belonging to the alkaloid class with the molecular formula C10H17N7O4 (Molecular weight = 299) and is composed of a 3,4-propinoperhydropurine tricyclic system (Figure 3A). Saxitoxin was the first PST whose purification dates back to 1957. Since then, 57 analogues of saxitoxin have been described [67,79]. The presence of two guanidinium groups in its structures makes this molecule highly polar [38,80]. Saxitoxin and its structural analogues differ in the fractions of the lateral groups (R1–R4) and, therefore, are commonly grouped according to these variable residues. They can be structurally classified as non-sulfated, such as STX and neosaxitoxin (neoSTX), mono-sulfated, such as gonyautoxins (GTXs 1–6), or di-sulfates (C1–4 toxins), decarbamoylated as decarbamoyl-saxitoxins (dcSTX, dcneoSTX), and deoxy-decarbamoylated like decarbamoyl-gonyautoxins (dcGTXs 1–4), and the 13-deoxy-decarbamoyl derivatives (doSTX, doGTX 2,3) [40,79,81].
2.1.2. Toxicity
Given the structural variation between saxitoxin and analogues, there is also a fluctuation in the toxicity range of these PSTs which varies by two orders of magnitude ranging from 15 MU·μmol−1 of C1 toxin to 2483 MU·μmol−1 of STX [81]. The proton group 7,8,9 guanidinium has been shown to be the key site to the bioactivity of the toxin as it allows efficient binding to Na [40]. It blocks the ion flux by binding the resting receptor site of the channels NaV1.1.–NaV1.4 and NaV1.6, which are the one of 10 subtypes of the alpha subunits of the NaV channels [82,83,84]. Saxitoxin acts by interacting with the extracellular regions of the Nav channels, without any dependence on tension [83]. However, it also binds to calcium [85] and potassium channels [86]. Saxitoxin is not directly involved in blocking both channels as in the case of NaVs. In K+ channels, 4 saxitoxins molecules bind to the extracellular site causing strong transmembrane depolarization that causes the canal to open and reduce potassium exchange [67,86]. In calcium channels, saxitoxin is associated with binding to the selectivity filter, but this binding does not cause complete blockage of the channel [85,87]. Llewellyn et al. in 2008 also observed that saxitoxin behaved as a weak inhibitor of nitric oxide neuronal synthase, which catabolizes the production of nitric oxide from the guanidine group of arginine [40]. To date, there is no clinically approved antidote on the market for poisoning with saxitoxin or its derivatives. The only common treatment is with activated carbon and application of artificial respiration in the early stages of poisoning [88].
2.1.3. Possible Applications
Toxins produced by the genus Alexandrium play an important role in research. For instance, saxitoxin and its analogues have been used to study and understand how sodium potassium channels work [22].
In 2000, saxitoxin, neosaxitoxin, decarbamoyl saxitoxin, and tetrodotoxin were tested for rat sciatic nerve block, exploring their potential as agents for extending the duration of local anesthesia [89]. Lethality, onset, and duration of action for thermal analgesia (hot-plate testing) and motor block (weight-bearing) were evaluated. Analgesia for 60 min was obtained with neosaxitoxin at 34 ± 2 micromol/L, saxitoxin at 58 ± 3 micromol/L, TTX at 92 ± 5 micromol/L, and decarbamoyl saxitoxin at 268 ± 8 micromol/L [89]. Wand et al. in 2006 detected the presence of goyautoxins in ATGX02 strain of Alexandrium tamarense isolated near Weizhou Island in the Beibu Gulf [81], while Xu et al. in 2021 found that Alexandrium tamiyavanichii isolated from Beibu Gulf waters in China produces low levels of saxitoxins and gonyautoxins [90]. The gonyautoxins are phycotoxins that act by reversibly blocking voltage-gated sodium channels at axonal levels. In Lettes et al. 2009, the therapeutic properties of gonyautoxin 2/3 epimers local infiltration in chronic tension-type headache patients were evaluated. The clinical trial was conducted from September 2004 to 2005 in Hospital Clínico Universidad de Chile. Twenty-seven patients with chronic tension-type headaches aged 18–80 years old were locally infiltrated with a dose of 50 µg of gonyautoxins. A total of 70% of patients responded to the treatment with long-lasting effects [91].
2.2. Spiroimines (Gymnodimines and Spirolides)
2.2.1. Chemistry
Both gymnodimines and spirolides are members of the spiroimine group of lipophilic macrocyclic imine toxins [92]. Together with prorocentrolides, spiro-prorocentrimine, pinnatoxins, and pteriatoxins, they form a part of the cyclic imine (CI) family and are collectively linked to sporomine mollusc poisoning (SMP) [93,94]. The CI family toxins are characterized by macrocyclic compounds with imine, i.e., a double carbon–nitrogen bond, and spiro-linked ether moieties [95]. The spiroimine subunit has a substantial carbocyclic polyether structure, ranging from 14 members for portimines to 27 members for pinnatoxines. Spirolides and gymnodimines have a carbocycle of 23 and 16 members, respectively [96,97]. Currently, 17 individual members of the spirolides subgroup have been identified [98]. An important feature to consider regarding spirolides is their water stability, as it is parallel to the potency of these marine toxins. Spirolides A (Figure 3B) and B are known to be unstable, while a C32-monomethyl fraction is rapidly hydrolyzed to more stable and common spirolides such as E and F. Spirolides C (Figure 3C), D, G, H, and I instead have an imine ring replaced with 31,32-dimethyl and appear to be much more resistant to hydrolysis [96].
On the contrary of spirolides, gymnodimines have been detected in smaller amounts and variety in microalgae [98]. To date, eight members of gymnodimines have been described [98]. Gymnodimines are the smallest members of the cyclic imine family by molecular weight (with a molecular weight of 507.7 g/mol) [96]. Gymnodimines are characterized by a labile 6,6-spirocyclic imine fragment embedded in a 16-membered carbocycle incorporating a tetrahydrofuran ring and one (GYM (A))5 or two (GYMs B11 and C)12 trisubstituted E-double bonds [96].
2.2.2. Toxicity
Generally, CI toxins are antagonists of muscle and neural-type nicotinic acetylcholine receptors (nAChRs) [99]. Specifically, Gymnodimine A (Figure 3D) and 13-desmethyl spirolide C (Figure 3E) express neurotoxicity by broadly targeting and reversibly blocking with high-affinity nicotinic acetylcholine receptors (nAChR) [100,101]. The toxicity of CId as “fast acting” induces a set of neurological symptoms followed by death in mouse bioassays [102]. Other observed negative impacts of the CI toxins are expressed on neuromuscular, sensory, digestive, and respiratory systems [93,94]. Munday et al. in 2004 [102] observed that the gymnodimine expressed high toxicity in mouse by injection (LD50 = 96 µg/kg) However, when gymnodimine was ingested together with food, exceeding the LD50 approximately 10 times of oral LD50, it expressed low toxicity [102]. Spirolides express a similar toxicity profile of decreased toxicity when the toxin is ingested with food [103]. In 2001, three new spirolides derived from Alexandrium ostenfeldii called Spirolide A, Spirolide C, and 13-Desmethyl spirolide C were isolated [104].
2.2.3. Possible Applications
The compound 13-Desmethyl spirolide C is of strong pharmacological interest as it targets nicotinic receptors. In 2006 [105], in vivo studies were conducted in mice 3xTg-AD affected by Alzheimer’s, and 11.9 µg/kg of 13-desmethyl spirolide C by intraperitoneal injections was given. Authors noted that 2 min after injection, 13-desmethyl spirolide C was located in the brain and remains detectable even 24 h after administration. The 13-desmethyl spirolide C induced in mice 3xTg-AD positive effects on Alzheimer’s disease markers with an increase in N-acetyl aspartate (NAA) levels, an increase in synaptophytin, and a decrease in intracellular amyloid beta in the hypothalamus, further concluding that the 13-desmethyl spirolide C was able to cross the blood–brain barrier, making this a toxin of inspiration for the development of new drugs for neurodegenerative diseases [105].
2.3. Goniodomins
2.3.1. Chemistry
Goniodomins are characterized as polyether macrolide compounds consisting of 32- membered macrolactone with an array of five- and six-membered cyclic ethers [106].
2.3.2. Toxicity
These toxins have lipophilic characteristics and are stored intracellularly and expressing significantly higher toxicity when released in the environment upon cytolysis [53,106]. However, even slight changes in the chemical structure of Goniodomin A (Figure 3F) induces a large change in its cytotoxicity. Goniodomin A, however, expresses a toxicity 10 times higher than Goniodomin B. For the latter, the opening of the C-ring and the isomeric changes between C-24 and C-29 appear to be the modifications responsible for a decrease in the toxicity of the molecule [49].
2.3.3. Possible Applications
The Goniodomin A toxin has a wide range of biological activities, such as the inhibition of fungal growth, the termination of cell division in fertilized sea-urchin eggs, and even causes morphological changes in the liver [49,106]. Goniodomins also disturbs the actomyosin ATPase activity by binding stoichiometrically to actin monomers in filaments, causing conformational changes. This activity is highly affected by myosin and the toxin disturbs the ATPase activity if its concentration is >10−6 M, otherwise the ATPase activity is enhanced [49,107].
Some suppressive angiogenic activity in endothelial cells induced by Goniodomin A has also been found acting by inhibition of actin reorganization. The presence of Goniodomin confirmed this property by inhibiting angiogenesis in vivo. It also inhibited the development of the vascular system in a chorioallantoic membrane, but no effect was observed on the already established structures of the strain fiber formed with G-betagamma expression (Gβγ) [108].
Goniodomin A is also believed to express neurotoxicity. It induced an increase in cytosolic calcium and depolarization of the cell membrane when this ion is present in the extracellular medium. In addition, as in the study by Espiña et al. in 2016, neuroblastoma cells were more sensitive to the toxin than rat hepatocytes when they affected their metabolic rate, concluding that Goniodomin A may induce high neurotoxicity if it reaches the nervous system [49].
Mizuno et al. in 1998 reported that Goniodomin A induced morphological changes in human brain astrocytoma cells by increasing the filamentous actin content. The cells were becoming refractile from flat and it was accompanied by the outgrowth of a needle-like structures from the cell surface [109]. Successively, Matsunaga et al. confirmed that even if Goniodomin A was significantly inhibited by the troponin–tropomyosin complex, as it induced a change in the distribution in the same human astrocyte cell line (1321N1 cells) by increasing the intracellular content of F-actin [107]. The authors also demonstrated that Goniodomin A inhibited ATPase activities of atrial myofibrils, natural actomyosin, and reconstituted actomyosin [107]. Goniodomin A is known for its ability to suppress the angiogenic properties of endothelial cells at least in part through the inhibition of actin reorganization in cells [108].
2.4. Tetrodotoxin
2.4.1. Chemistry
Tetrodotoxin (TTX; Figure 3G), also called heterocyclic organic molecule hydroperquinozolinine (aminoperhydroquinazo-lone), is an orthoester with a group of guanidinium and a system of polyoxygenated rings, similar to saxitoxin, found in Alexandrium tamarense [110,111]. This toxin is heat stable, water soluble [112], and degrades at alkaline pH’s [110]. The TTX structure is similar to a prism with a tetrahedral core [110]. There have been 30 structural analogues of the tetrodotoxin reported, of which 26 are known to be naturally occurring [113,114].
2.4.2. Toxicity
The active moiety of tetrodotoxin, same as for saxitoxin, is known to be the only 1,2,3 guanidinium moiety. It is connected to a highly oxygenated carbon skeleton that possesses a 2,4-dioxaadamantane portion containing five hydroxyl groups and expresses its bioactivity when the guanidium group is protonated [110,115]. The toxicity degree of tetrodotoxin is also affected by the number and position of hydroxyl groups present in the structure. As demonstrated by Yang and Kao reported in 1998, hydroxyl groups at C-4, C-6, C-9, C-10 and C-11 are involved in the binding of the sodium channel [116]. Subsequently, Yotsu-Yamashita et al. in 1999 confirmed the involvement of hydroxyl groups to tetrodotoxin C-6 and C-11 in acting as hydrogen bond donors for sodium channel binding [113].
The guanidinium moieties on tetrodotoxin and on its analogs are the pharmacophores, and therefore are responsible for the blocking activity on the NaV channels [82]. The binding of tetrodotoxin to NaV results from the interaction between the positively charged guanidine group on the TTX with the negatively charged carboxylate groups on the side chains in the mouth of site 1 of the sodium channel, thereby partially or completely blocking the inward Na+ ion current [117,118].
The guanidinium toxins express toxicity to various ion channel types, but their primary target is voltage-gated ion channel blockage by binding to the NaV channels of excitable cells, mainly in nerve and muscle cell [82,110]. Tetrodotoxin binds and inhibits specifically six out of nine sodium channel isoforms expressed in mammalian cells: NaV1.1, NaV1.2, NaV1.3, and NaV1.7 [82,119,120]. It then terminates the nerve conduction and the muscle action potentials [121] leading to progressive paralysis and, in extreme cases, to death because of respiratory and heart failure [122,123]. Its toxicity is often emphasized by comparing it as being over a thousand times more toxic to humans than cyanide [124]. Even now, tetrodotoxin has no known antidote which is officially approved [125]. The only treatment for the toxin in case of intoxication is observation and the appropriate supportive care [124]. The supportive care includes practices such as gastric lavage with activated charcoal to remove unabsorbed toxin and machine-assisted ventilation when respiration is severely affected in conjunction with the use of dopamine and atropine to correct a problem with hypotension and arrhythmia, accordingly [126,127,128]. Typically, symptoms are expressed within 1 to 6 h if the toxin is ingested via fish [125].
2.4.3. Possible Applications
As summarized by Nieto et al. in 2012 [129], the administration of tetrodotoxin has been evaluated in both humans and experimental animals under different pain conditions [129]. In Hagen et al. 2017, the bioactivity of tetrodotoxin was investigated by daily subcutaneous injection for four consecutive days of 30 µg of tetrodotoxins in patients randomized to receive tetrodotoxins or placebo. All the patients were diagnosed with cancer. A clinically important analgesic signal was shown to moderate pain related to the tetrodotoxin cancer group [130]. An example of a drug based on tetrodotoxin is Halneuron®, a pain medication drug undergoing phase 3 clinical trials as a new class of non-opioid analgesics (https://wexpharma.com/; accessed on 27 December 2023). This drug was developed for the treatment of moderate-to-severe chemotherapy-induced neuropathic pain, cancer pain, and other pain indications. It has been already tested on 700 people in over 15 clinical trials and is going for a double-blinded, placebo-controlled Phase 3 trial (https://wexpharma.com/; accessed on 27 December 2023).
2.5. Other Molecules
Galasso et al. in 2018 [131] investigated the anti-cancer activity of Alexandrium minutum against a panel of cancer cell lines such as human pancreatic cancer cells PC-3, human colorectal adenocarcinoma cells HT29, and human lung adenocarcinoma cells A549. The authors found a specific anti-proliferative fraction from the total extract of A. minutum against A459, named fraction 3B. The activity was specific against A549 with an IC50 of 0.4 µg/mL. They evaluated the activation of the mitophagy pathway with a differential expression gene analysis. After a cell treatment with 0.4 µg/mL of fraction 3B of A. minutum for 2 h, it was observed that a strong up-regulation of the autophagy-related protein 12/gene (Atg12) with a consequent increase in ATPase H+ Transporting V1 Subunit G2 (ATP6V1G2) and BCL2/Adenovirus E1B19 kDa Interacting Protein 3 (BNIP3) occurred. In addition, there was up-regulation of two genes involved in mitophagy: PTEN induced putative kinase1 (PINK1) and the Parkin gene. The authors did not find the same cytotoxic and mitophagy activities against the normal lung fibroblast WI38, used as a control cell line. After a chemical analysis by colorimetric the phenol–sulphuric acid method and Breadford assay, Galasso et al. in 2018 revealed that this fraction was rich in carbohydrates (94%) and protein (4%) [131].
Yamasaki et al. 2008 [132] isolated a polysaccharide-based toxin from the dinoflagellate Alexandrium tamarense. They purified the AT-toxin by gel filtration on the fast protein liquid chromatography-FPLC system and thanks to sugar composition assay, noted that this toxin contains galactose, fucose, mannose, N-acetylglucosamine, xylose, and other minor saccharides. The polysaccharide-based AT-toxin showed cytotoxic effects against different cell lines such as the HeLa human immortalized cervix carcinoma cells, Vero human kidney epithelial cells, XC rat Rous Sarcoma cells, CHO Chinese hamster ovary epithelial cells, and U937 human monocyte Histiocytic Lymphoma cells. On U937, the treatment with polysaccharide-based TA-toxin induced a consistent cytotoxicity with an IC50 of 110 ng/mL. The authors demonstrated that AT-toxin was an apoptotic inducible toxin by measuring the increasing DNA fragmentation that is a hallmark of apoptotic cell death in U937 cells by diphenylamine assay after treatment with TA-toxin [132].
Sansone et al. 2018 [133] also found that the Alexandrium andersoni raw methanol total extract and the solid-phase extraction-SPE fractions induced high cytotoxicity on HT29 human colorectal cancer cells and A549 human lung adenocarcinoma cells, while they did not observe cytotoxicity for the two normal fibroblast cell lines WI38 and BEAS-2B. Fraction B was the most active on A549 cells, with the minimum concentration for inhibition cell viability ≤ 1 µg/mL, while fraction D was the most active fraction on HT29 cells, where the minimum concentration for inhibition of cell viability was >1 μg/mL. Nuclear magnetic resonance-NMR and liquid chromatography-mass spectrometry LC-MS spectra supported a different distribution of the metabolites in SPE fractions. The glycolipids, mostly digalactosyl diacyl glycerol (MGDG) and sulfoquinovosyl diacyl glycerol (SQDG), were the predominant metabolites in fraction B, whereas fraction D was mostly composed by sterols (ST) and triacylglycerols (TAG). Sansone et al. found that A. andersoni activated two different cell death pathways in the two tumor cell lines. Both signaling pathways observed were triggered by tumor necrosis factor receptor (TNFR), as observed by a gene expression analysis after 2 h of treatment with 1 μg/mL concentration of the two fractions in A549 and HT29. They noted that SPE fraction B was able to induce a down-regulation of anti-apoptotic molecules (BIRC3 and TRAF2) and an up-regulation of death receptors (CD27, DR3, TNF, and TNFS8) in A549 cells. In HT29 cells, it was observed that fraction D directly induced cell death through a particular TNF-related pathway also involving DNA cell-signaling damage. Two tumor necrosis factors, FASLG and TNF, were over-expressed as well as Gadd45 alpha and gamma, and Foxl1, confirming a cell-signaling pathway in response to the DNA damage. Gadd45 alpha and gamma proteins play important roles in suppressing cell proliferation, mediating cell cycle arrest, promoting apoptosis, inducing DNA repair, and stabilizing chromatin assessment [116].
De vera et al. in 2018 [134] conducted a bioprospecting study on the extracts of 33 strains of microalgae. Between these strains, they found that the extracts of the microalga Alexandrium tamarense had an anti-proliferative activity at concentration of 50 µg/mL against tumorigenic breast cancer cell MCF-7, non-tumorigenic breast cancer cell MCF-10A, and the two tumorigenic cell lines of prostate cancer cells LNCaP and PC-3. Authors also evaluated the apoptotic activity of A. tamarense extracts with an annexin V binding assay, showing that it induced apoptosis on the human hepatocellular carcinoma cell line HepG2 tested at 100 µg/mL. Raw extracts of Alexandrium andersonii also showed a weak cell death on human lung carcinoma A549 and on colorectal carcinoma HT29 cell lines [134]. Lauritano et al. in 2016 also evaluated the antiproliferative activity against human melanoma A2058 cells of 32 microalgal species, including three Alexandrium, i.e., A. andersonii (FE108), A. tamutum (FE107), and A. minutum (FE126) from the Mediterranean Sea [32]. In particular, they found that FE108 and FE107 reduced almost to 0% A2058 cell viability already at 10 µg/mL. FE107 extract (FE107/3) derived from a culture in phosphate starvation had weaker effect, showing antiproliferative activity at 25–100 µg/mL. Finally, FE126 reduced cell viability at 80% at 50 µg/mL, while at 100 µg/mL cell viability was reduced at 30%. All the three Alexandium species also showed cytotoxicity on normal human lung fibroblast (MRC-5). The authors showed how different culturing conditions as well as different clones of the same species may exert different bioactivities. The compounds responsible of those activities are still not characterized [32].
Some species of Alexandrium genus also produce a series of bioactive extracellular compounds (BECs) not yet completely characterized. One example of characterized BEC is alexandrolide, a microalgal growth inhibitor isolated from A. catenella which also showed cytotoxicity against mouse lymphoid P388. The production of BECs has an energy cost for the cell like the production of any organic molecule, but the mechanisms for their production are still not clear [22]. Extracellular particles produced by some species of Alexandrium may have anti-parasitic activities [22]. Long et al. [135] showed Alexandrium minutum activity against the dinoflagellate parasite Amoebophrya sp. The authors noted that exposure of free-life infectious spores of Amoebophrya sp. to the filtered exudates with allelochemical properties of A. minutum affected the mortality rate of Amoebophrya sp. In addition to the infectious capacity of the latter, exposure to A. minutum exudates quickly resulted in permeation of the membranes of Amoebophrya sp. dinospores, leading to the loss of dinofluorescence associated in turn with the loss in virulence of the parasite in cultures. After 20 min of exposure with A. minutum extracts, a reduction of 40% of viability was observed while after 2 h of exposure the reduction was almost 98% [135]. Lassudrie et al. in 2015 [136] observed the anti-viral properties of the species Alexandrium catenella against herpesvirus OsHV-1 μvar affecting oysters. When the herpes virus OsHV-1 μvar was exposed to A. catenella, the prevalence of viral infection in oysters decreased. The authors concluded that the reduction in viral infection is due to an antagonistic relationship that is established between the virus and toxic microalgae assuming various mechanisms of action. Firstly, there may be a direct interaction between OsHV-1 mvar and A. catenella that could reduce viral transmission and algae availability for oyster consumption, or the antagonistic action could be the consequence of immune and physiological responses to the virus and/or microalgae [136]. The potential direct deleterious effects of Alexandrium BECs on marine viruses can then mitigate the infection by decreasing the viral load. To our knowledge, the effects of BECs on pathogenic bacteria have not yet been studied. Bioactivities reported for Alexandrium spp. compounds and extracts are summarized in Table 2.
3. Conclusions
This review describes Alexandrium toxins and their possible useful applications. In particular, anti-cancer, anti-microbial, anti-parasite, and anti-Alzheimer activities have been reported for some Alexandrium compounds or extracts. These data highlight that even if some Alexandrium spp. are very toxic and dangerous for other marine organisms and humans, some strains may produce metabolites with possible beneficial effects for human health. This was also found for other toxic species such as cyanobacteria [138,139]. This testifies how important it is for biological inter-species diversity, and how it can protect and maintain this diversity for the health status of the marine environment and humans. The diverse range of compounds produced, both toxic and beneficial, can vary under different growth conditions, representing an opportunity for further research. By delving deeper into this aspect, it will be possible to identify growth conditions to enhance the production of specific compounds. By improving production efficiency and conducting in-depth molecular studies, we can gain a detailed understanding of the molecular and metabolic pathways involved in microalgae bioactive compound production. This knowledge can then be used to develop genome editing techniques that transform microalgae into a versatile biotechnological tool. One approach to achieve this is to increase transcriptomic studies on the same algae cultivated under various conditions (e.g., variation of temperature, pH, and salinity, as well as the composition of culture media) in order to identify differentially expressed genes involved in compound metabolism and potentially target them for silencing (e.g., by using RNA interference molecular techniques). This research approach holds great potential for enhancing the production of specific compounds and optimizing the beneficial properties of microalgae. Exciting research directions in light of bioactivity discovery useful for human health can be represented by the search for new Alexandrium strains/clones, new metabolites, and through evaluation of the growth conditions in which these compounds are mostly produced, as well as through considering the possibility of synthetic modification of some molecule structures to reduce side effects and increase specificity and efficacy. Results will be of interest not only for the scientific community, but also for human health and the pharmaceutical industries.
Author Contributions
Conceptualization, C.L.; writing—original draft preparation, E.M., D.D.L., D.S. and C.L.; writing—review and editing, E.M., D.D.L., A.P., D.S. and C.L.; supervision, C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gómez, F. A List of Dinoflagellates in the World’s Oceans. Acta Bot. Croat. 2005, 64, 129–212. [Google Scholar]
- Gómez, F. A Checklist and Classification of Living Dinoflagellates (Dinoflagellata, Alveolata). Cicimar Oceánides 2012, 27, 65–140. [Google Scholar] [CrossRef]
- Gómez, F.; Artigas, L.F. Redefinition of the Dinoflagellate Genus Alexandrium Based on Centrodinium: Reinstatement of Gessnerium and Protogonyaulax, and Episemicolon Gen. Nov. (Gonyaulacales, Dinophyceae). J. Mar. Biol. 2019, 2019, 1284104. [Google Scholar] [CrossRef]
- Taylor, F.J.R.; Hoppenrath, M.; Saldarriaga, J.F. Dinoflagellate Diversity and Distribution. Biodivers. Conserv. 2008, 17, 407–418. [Google Scholar] [CrossRef]
- Cohen, N.R.; McIlvin, M.R.; Moran, D.M.; Held, N.A.; Saunders, J.K.; Hawco, N.J.; Brosnahan, M.; DiTullio, G.R.; Lamborg, C.; McCrow, J.P.; et al. Dinoflagellates Alter Their Carbon and Nutrient Metabolic Strategies across Environmental Gradients in the Central Pacific Ocean. Nat. Microbiol. 2021, 6, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Smayda, T.J. Adaptive Ecology, Growth Strategies and the Global Bloom Expansion of Dinoflagellates. J. Oceanogr. 2002, 58, 281–294. [Google Scholar] [CrossRef]
- Schnepf, E.; Elbrächter, M. Nutritional Strategies in Dinoflagellates. Eur. J. Protistol. 1992, 28, 3–24. [Google Scholar] [CrossRef]
- Camacho, F.G.; Rodríguez, J.G.; Mirón, A.S.; García, M.C.C.; Belarbi, E.H.; Chisti, Y.; Grima, E.M. Biotechnological Significance of Toxic Marine Dinoflagellates. Biotechnol. Adv. 2007, 25, 176–194. [Google Scholar] [CrossRef]
- Matsuyama, Y. Impacts of the Harmful Dinoflagellate Heterocapsa circularisquama Bloom on Shellfish Aquaculture in Japan and Some Experimental Studies on Invertebrates. Harmful Algae 2012, 14, 144–155. [Google Scholar] [CrossRef]
- Sakamoto, S.; Lim, W.A.; Lu, D.; Dai, X.; Orlova, T.; Iwataki, M. Harmful Algal Blooms and Associated Fisheries Damage in East Asia: Current Status and Trends in China, Japan, Korea and Russia. Harmful Algae 2021, 102, 101787. [Google Scholar] [CrossRef]
- Bechard, A. Red Tide at Morning, Tourists Take Warning? County-Level Economic Effects of HABS on Tourism Dependent Sectors. Harmful Algae 2019, 85, 101689. [Google Scholar] [CrossRef] [PubMed]
- Berge, E.; Whiteley, W.; Audebert, H.; De Marchis, G.; Fonseca, A.C.; Padiglioni, C.; Pérez De La Ossa, N.; Strbian, D.; Tsivgoulis, G.; Turc, G. European Stroke Organisation (ESO) Guidelines on Intravenous Thrombolysis for Acute Ischaemic Stroke. Eur. Stroke J. 2021, 6, I–LXII. [Google Scholar] [CrossRef] [PubMed]
- Bricelj, V.M.; Haubois, A.-G.; Sengco, M.R.; Pierce, R.H.; Culter, J.K.; Anderson, D.M. Trophic Transfer of Brevetoxins to the Benthic Macrofaunal Community during a Bloom of the Harmful Dinoflagellate Karenia Brevis in Sarasota Bay, Florida. Harmful Algae 2012, 16, 27–34. [Google Scholar] [CrossRef]
- Alves, T.; Mafra, L. Diel Variations in Cell Abundance and Trophic Transfer of Diarrheic Toxins during a Massive Dinophysis Bloom in Southern Brazil. Toxins 2018, 10, 232. [Google Scholar] [CrossRef] [PubMed]
- Gibble, C.M.; Hoover, B.A. Interactions between Seabirds and Harmful Algal Blooms. In Harmful Algal Blooms; Shumway, S.E., Burkholder, J.M., Morton, S.L., Eds.; Wiley: Hoboken, NJ, USA, 2018; pp. 223–242. ISBN 978-1-118-99467-2. [Google Scholar]
- Reis Costa, P. Impact and Effects of Paralytic Shellfish Poisoning Toxins Derived from Harmful Algal Blooms to Marine Fish. Fish Fish. 2016, 17, 226–248. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. 30 March 2023. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 11 August 2023).
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The Globally Distributed Genus Alexandrium: Multifaceted Roles in Marine Ecosystems and Impacts on Human Health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef]
- Van Dolah, F.M. Marine Algal Toxins: Origins, Health Effects, and Their Increased Occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful Algal Blooms and Eutrophication: Nutrient Sources, Composition, and Consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal Revision of the Alexandrium tamarense Species Complex (Dinophyceae) Taxonomy: The Introduction of Five Species with Emphasis on Molecular-Based (rDNA) Classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef]
- Long, M.; Krock, B.; Castrec, J.; Tillmann, U. Unknown Extracellular and Bioactive Metabolites of the Genus Alexandrium: A Review of Overlooked Toxins. Toxins 2021, 13, 905. [Google Scholar] [CrossRef]
- Lewis, A.M.; Coates, L.N.; Turner, A.D.; Percy, L.; Lewis, J. A Review of the Global Distribution of Alexandrium minutum (Dinophyceae) and Comments on Ecology and Associated Paralytic Shellfish Toxin Profiles, with a Focus on Northern Europe. J. Phycol. 2018, 54, 581–598. [Google Scholar] [CrossRef] [PubMed]
- Casabianca, S.; Penna, A.; Pecchioli, E.; Jordi, A.; Basterretxea, G.; Vernesi, C. Population Genetic Structure and Connectivity of the Harmful Dinoflagellate Alexandrium minutum in the Mediterranean Sea. Proc. R. Soc. B 2012, 279, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Wang, F.; Zhang, W. Omics Analysis for Dinoflagellates Biology Research. Microorganisms 2019, 7, 288. [Google Scholar] [CrossRef] [PubMed]
- Vingiani, G.M.; Štālberga, D.; De Luca, P.; Ianora, A.; De Luca, D.; Lauritano, C. De Novo Transcriptome of the Non-Saxitoxin Producing Alexandrium tamutum Reveals New Insights on Harmful Dinoflagellates. Mar. Drugs 2020, 18, 386. [Google Scholar] [CrossRef]
- Meng, F.-Q.; Song, J.-T.; Zhou, J.; Cai, Z.-H. Transcriptomic Profile and Sexual Reproduction-Relevant Genes of Alexandrium minutum in Response to Nutritional Deficiency. Front. Microbiol. 2019, 10, 2629. [Google Scholar] [CrossRef]
- Jaeckisch, N.; Yang, I.; Wohlrab, S.; Glöckner, G.; Kroymann, J.; Vogel, H.; Cembella, A.; John, U. Comparative Genomic and Transcriptomic Characterization of the Toxigenic Marine Dinoflagellate Alexandrium ostenfeldii. PLoS ONE 2011, 6, e28012. [Google Scholar] [CrossRef]
- Brown, E.R.; Moore, S.G.; Gaul, D.A.; Kubanek, J. Differentiating Toxic and Nontoxic Congeneric Harmful Algae Using the Non-Polar Metabolome. Harmful Algae 2021, 110, 102129. [Google Scholar] [CrossRef]
- Galluzzi, L.; Penna, A.; Bertozzini, E.; Vila, M.; Garcés, E.; Magnani, M. Development of a Real-Time PCR Assay for Rapid Detection and Quantification of Alexandrium Minutum (a Dinoflagellate). Appl. Environ. Microbiol. 2004, 70, 1199–1206. [Google Scholar] [CrossRef]
- Galluzzi, L.; Bertozzini, E.; Penna, A.; Perini, F.; Garcés, E.; Magnani, M. Analysis of rRNA Gene Content in the Mediterranean Dinoflagellate Alexandrium catenella and Alexandrium taylori: Implications for the Quantitative Real-Time PCR-Based Monitoring Methods. J. Appl. Phycol. 2010, 22, 1–9. [Google Scholar] [CrossRef]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef]
- Balech, E. The Genus Alexandrium halim (Dinoflagellata); Sherkin Island Marine Station: Sherkin Island, Ireland, 1995. [Google Scholar]
- Lim, A.S.; Jeong, H.J.; Ok, J.H. Five Alexandrium Species Lacking Mixotrophic Ability. ALGAE 2019, 34, 289–301. [Google Scholar] [CrossRef]
- Wyatt, T.; Jenkinson, I.R. Notes on Alexandrium Population Dynamics. J. Plankton Res. 1997, 19, 551–575. [Google Scholar] [CrossRef]
- Anderson, D.M. Physiology and Bloom Dynamics of Toxic Alexandrium Species, with Emphasis on Life Cycle Transitions. Nato Asi. Ser. G Ecol. Sci. 1998, 41, 29–48. [Google Scholar]
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the Health Potential of Microalgae as Sustainable Sources of Bioactive Compounds. IJMS 2021, 22, 4383. [Google Scholar] [CrossRef] [PubMed]
- Schantz, E.J.; Ghazarossian, V.E.; Schnoes, H.K.; Strong, F.M.; Springer, J.P.; Pezzanite, J.O.; Clardy, J. Structure of Saxitoxin. J. Am. Chem. Soc. 1975, 97, 1238–1239. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-Z. Neurotoxins from Marine Dinoflagellates: A Brief Review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, L.E. Saxitoxin, a Toxic Marine Natural Product That Targets a Multitude of Receptors. Nat. Prod. Rep. 2006, 23, 200. [Google Scholar] [CrossRef]
- Kim, H.-S.; Park, H.; Wang, H.; Kim, T.; Ki, J.-S. Saxitoxins-Producing Potential of the Marine Dinoflagellate Alexandrium affine and Its Environmental Implications Revealed by Toxins and Transcriptome Profiling. Mar. Environ. Res. 2023, 185, 105874. [Google Scholar] [CrossRef]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Montresor, M. Saxitoxin and Neosaxitoxin as Toxic Principles of Alexandrium andersoni (Dinophyceae) from the Gulf of Naples, Italy. Toxicon 2000, 38, 1871–1877. [Google Scholar] [CrossRef]
- Sampedro, N.; Franco, J.M.; Zapata, M.; Riobó, P.; Garcés, E.; Penna, A.; Caillaud, A.; Diogène, J.; Cacho, E.; Camp, J. The Toxicity and Intraspecific Variability of Alexandrium andersonii Balech. Harmful Algae 2013, 25, 26–38. [Google Scholar] [CrossRef]
- Navarro, J.M.; Contreras, A.M. An Integrative Response by Mytilus Chilensis to the Toxic Dinoflagellate Alexandrium catenella. Mar. Biol. 2010, 157, 1967–1974. [Google Scholar] [CrossRef]
- Tatters, A.O.; Flewelling, L.J.; Fu, F.; Granholm, A.A.; Hutchins, D.A. High CO2 Promotes the Production of Paralytic Shellfish Poisoning Toxins by Alexandrium catenella from Southern California Waters. Harmful Algae 2013, 30, 37–43. [Google Scholar] [CrossRef]
- Anderson, D.M.; Fachon, E.; Pickart, R.S.; Lin, P.; Fischer, A.D.; Richlen, M.L.; Uva, V.; Brosnahan, M.L.; McRaven, L.; Bahr, F.; et al. Evidence for Massive and Recurrent Toxic Blooms of Alexandrium catenella in the Alaskan Arctic. Proc. Natl. Acad. Sci. USA 2021, 118, e2107387118. [Google Scholar] [CrossRef] [PubMed]
- Eckford-Soper, L.K.; Bresnan, E.; Lacaze, J.-P.; Green, D.H.; Davidson, K. The Competitive Dynamics of Toxic Alexandrium Fundyense and Non-Toxic Alexandrium tamarense: The Role of Temperature. Harmful Algae 2016, 53, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Okita, Y.; Matsuda, H.; Okino, T.; Yamaguchi, K. From the Dinoflagellate Alexandrium hiranoi. Phytochemistry 1998, 48, 85–88. [Google Scholar] [CrossRef]
- Espiña, B.; Cagide, E.; Louzao, M.C.; Vilariño, N.; Vieytes, M.R.; Takeda, Y.; Sasaki, M.; Botana, L.M. Cytotoxicity of Goniodomin A and B in Non Contractile Cells. Toxicol. Lett. 2016, 250–251, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhai, X.; Hao, S.; Shang, L.; Deng, Y.; Chai, Z.; Chen, J.; Hu, Z.; Tang, Y.Z. Novel Non-Paralytic Shellfish Toxin and Non-Spirolide Toxicity to Finfish, Brine Shrimp, and Rotifer Observed in a Culture of the Dinoflagellate Alexandrium insuetum Isolated From the Coastal Water of China. Front. Mar. Sci. 2021, 8, 735752. [Google Scholar] [CrossRef]
- Shang, L.; Xu, Y.; Leaw, C.P.; Lim, P.T.; Wang, J.; Chen, J.; Deng, Y.; Hu, Z.; Tang, Y.Z. Potent Allelopathy and Non-PSTs, Non-Spirolides Toxicity of the Dinoflagellate Alexandrium leei to Phytoplankton, Finfish and Zooplankton Observed from Laboratory Bioassays. Sci. Total Environ. 2021, 780, 146484. [Google Scholar] [CrossRef]
- Penna, A.; Perini, F.; Dell’Aversano, C.; Capellacci, S.; Tartaglione, L.; Giacobbe, M.G.; Casabianca, S.; Fraga, S.; Ciminiello, P.; Scardi, M. The Sxt Gene and Paralytic Shellfish Poisoning Toxins as Markers for the Monitoring of Toxic Alexandrium Species Blooms. Environ. Sci. Technol. 2015, 49, 14230–14238. [Google Scholar] [CrossRef]
- Hsia, M.H.; Morton, S.L.; Smith, L.L.; Beauchesne, K.R.; Huncik, K.M.; Moeller, P.D.R. Production of Goniodomin A by the Planktonic, Chain-Forming Dinoflagellate Alexandrium monilatum (Howell) Balech Isolated from the Gulf Coast of the United States. Harmful Algae 2006, 5, 290–299. [Google Scholar] [CrossRef]
- May, S.P.; Burkholder, J.M.; Shumway, S.E.; Hégaret, H.; Wikfors, G.H.; Frank, D. Effects of the Toxic Dinoflagellate Alexandrium monilatum on Survival, Grazing and Behavioral Response of Three Ecologically Important Bivalve Molluscs. Harmful Algae 2010, 9, 281–293. [Google Scholar] [CrossRef]
- Mackenzie, L.; White, D.; Oshima, Y.; Kapa, J. The Resting Cyst and Toxicity of Alexandrium ostenfeldii (Dinophyceae) in New Zealand. Phycologia 1996, 35, 148–155. [Google Scholar] [CrossRef]
- Cembella, A.D.; Lewis, N.I.; Quilliam, M.A. The Marine Dinoflagellate Alexandrium ostenfeldii (Dinophyceae) as the Causative Organism of Spirolide Shellfish Toxins. Phycologia 2000, 39, 67–74. [Google Scholar] [CrossRef]
- Gribble, K.E.; Keafer, B.A.; Quilliam, M.A.; Cembella, A.D.; Kulis, D.M.; Manahan, A.; Anderson, D.M. Distribution and Toxicity of Alexandrium ostenfeldii (Dinophyceae) in the Gulf of Maine, USA. Deep. Sea Res. Part. II Top. Stud. Oceanogr. 2005, 52, 2745–2763. [Google Scholar] [CrossRef]
- Borkman, D.G.; Smayda, T.J.; Tomas, C.R.; York, R.; Strangman, W.; Wright, J.L.C. Toxic Alexandrium peruvianum (Balech and de Mendiola) Balech and Tangen in Narragansett Bay, Rhode Island (USA). Harmful Algae 2012, 19, 92–100. [Google Scholar] [CrossRef]
- Tomas, C.R.; Van Wagoner, R.; Tatters, A.O.; White, K.D.; Hall, S.; Wright, J.L.C. Alexandrium peruvianum (Balech and Mendiola) Balech and Tangen a New Toxic Species for Coastal North Carolina. Harmful Algae 2012, 17, 54–63. [Google Scholar] [CrossRef]
- Blossom, H.E.; Daugbjerg, N.; Hansen, P.J. Toxic Mucus Traps: A Novel Mechanism That Mediates Prey Uptake in the Mixotrophic Dinoflagellate Alexandrium pseudogonyaulax. Harmful Algae 2012, 17, 40–53. [Google Scholar] [CrossRef]
- Zmerli Triki, H.; Laabir, M.; Moeller, P.; Chomérat, N.; Kéfi Daly-Yahia, O. First Report of Goniodomin a Production by the Dinoflagellate Alexandrium pseudogonyaulax Developing in Southern Mediterranean (Bizerte Lagoon, Tunisia). Toxicon 2016, 111, 91–99. [Google Scholar] [CrossRef]
- Collins, C.; Graham, J.; Brown, L.; Bresnan, E.; Lacaze, J.-P.; Turrell, E.A. Identification and toxicity of Alexandrium tamarense (dinophyceae) in scottish waters. J. Phycol. 2009, 45, 692–703. [Google Scholar] [CrossRef]
- Ichimi, K.; Suzuki, T.; Ito, A. Variety of PSP Toxin Profiles in Various Culture Strains of Alexandrium tamarense and Change of Toxin Profile in Natural A. tamarense Population. J. Exp. Mar. Biol. Ecol. 2002, 273, 51–60. [Google Scholar] [CrossRef]
- Hashimoto, T.; Matsuoka, S.; Yoshimatsu, S.; Miki, K.; Nishibori, N.; Nishio, S.; Noguchi, T. First Paralytic Shellfish Poison (PSP) Infestation of Bivalves Due to Toxic Dinoflagellate Alexandrium tamiyavanichii, in the Southeast Coasts of the Seto Inland Sea, Japan. J. Food Hyg. Soc. Jpn. 2002, 43, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Emura, A.; Matsuyama, Y.; Oda, T. Evidence for the Production of a Novel Proteinaceous Hemolytic Exotoxin by Dinoflagellate Alexandrium taylorii. Harmful Algae 2004, 3, 29–37. [Google Scholar] [CrossRef]
- Tillmann, U.; Krock, B.; Wietkamp, S.; Beran, A. A Mediterranean Alexandrium taylorii (Dinophyceae) Strain Produces Goniodomin A and Lytic Compounds but Not Paralytic Shellfish Toxins. Toxins 2020, 12, 564. [Google Scholar] [CrossRef] [PubMed]
- Cusick, K.; Sayler, G. An Overview on the Marine Neurotoxin, Saxitoxin: Genetics, Molecular Targets, Methods of Detection and Ecological Functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, L.; Miller, A.; Kosatsky, T. Changing Trends in Paralytic Shellfish Poisonings Reflect Increasing Sea Surface Temperatures and Practices of Indigenous and Recreational Harvesters in British Columbia, Canada. Mar. Drugs 2021, 19, 568. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, R.M.; Misner, I.; Tomas, C.R.; Wright, J.L.C. Occurrence of 12-Methylgymnodimine in a Spirolide-Producing Dinoflagellate Alexandrium peruvianum and the Biogenetic Implications. Tetrahedron Lett. 2011, 52, 4243–4246. [Google Scholar] [CrossRef]
- Hansen, P.J.; Cembella, A.D.; Moestrup, Ø. The marine dinoflagellate Alexandrium ostenfeldii: Paralytic shellfish toxin concentration, composition, and toxicity to a tintinnid ciliate. J. Phycol. 1992, 28, 597–603. [Google Scholar] [CrossRef]
- Harris, C.M.; Reece, K.S.; Stec, D.F.; Scott, G.P.; Jones, W.M.; Hobbs, P.L.M.; Harris, T.M. The Toxin Goniodomin, Produced by Alexandrium Spp., Is Identical to Goniodomin A. Harmful Algae 2020, 92, 101707. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID:56947150, Saxitoxin. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/56947150#section=2D-Structure (accessed on 19 August 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 21590535, Spirolide A. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/21590535#section=2D-Structure (accessed on 12 December 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 101136678, Spirolide C. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/101136678#section=2D-Structure (accessed on 12 December 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 11649137, Gymnodimine A. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/11649137#section=2D-Structure (accessed on 13 December 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 101136679, 13-Desmethyl Spirolide C. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/101136679#section=2D-Structure (accessed on 29 August 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 6506119, Goniodomin A. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/6506119#section=2D-Structure (accessed on 30 August 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 11174599, Tetrodoxin. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/tetrodotoxin#section=2D-Structure (accessed on 11 December 2023).
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic Alkaloids: Saxitoxin and Its Analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef]
- Bordner, J.; Thiessen, W.E.; Bates, H.A.; Rapoport, H. Structure of a Crystalline Derivative of Saxitoxin. Structure of Saxitoxin. J. Am. Chem. Soc. 1975, 97, 6008–6012. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Zhang, S.-F.; Zhang, Y.; Lin, L. Paralytic Shellfish Toxin Biosynthesis in Cyanobacteria and Dinoflagellates: A Molecular Overview. J. Proteom. 2016, 135, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Durán-Riveroll, L.; Cembella, A. Guanidinium Toxins and Their Interactions with Voltage-Gated Sodium Ion Channels. Mar. Drugs 2017, 15, 303. [Google Scholar] [CrossRef] [PubMed]
- Mattei, C.; Legros, C. The Voltage-Gated Sodium Channel: A Major Target of Marine Neurotoxins. Toxicon 2014, 91, 84–95. [Google Scholar] [CrossRef] [PubMed]
- England, S.; De Groot, M.J. Subtype-selective Targeting of Voltage-gated Sodium Channels. Br. J. Pharmacol. 2009, 158, 1413–1425. [Google Scholar] [CrossRef]
- Su, Z.; Sheets, M.; Ishida, H.; Li, F.; Barry, W.H. Saxitoxin Blocks L-Type I Ca. J. Pharmacol. Exp. Ther. 2004, 308, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Salata, J.J.; Bennett, P.B. Saxitoxin Is a Gating Modifier of hERG K+ Channels. J. Gen. Physiol. 2003, 121, 583–598. [Google Scholar] [CrossRef]
- Zhang, F.; Xu, X.; Li, T.; Liu, Z. Shellfish Toxins Targeting Voltage-Gated Sodium Channels. Mar. Drugs 2013, 11, 4698–4723. [Google Scholar] [CrossRef]
- Kao, C.Y. Paralytic Shellfish Poisoning. In Algal Toxins in Seafood and Drinking Water; Elsevier: Amsterdam, The Netherlands, 1993; pp. 75–86. ISBN 978-0-12-247990-8. [Google Scholar]
- Kohane, D.; Lu, N.; Gokgolkline, A.; Shubina, M.; Kuang, Y.; Hall, S.; Strichartz, G.; Berde, C. The Local Anesthetic Properties and Toxicity of Saxitonin Homologues for Rat Sciatic Nerve Block in Vivo. Reg. Anesth. Pain. Med. 2000, 25, 52–59. [Google Scholar] [CrossRef]
- Xu, Y.; He, X.; Li, H.; Zhang, T.; Lei, F.; Gu, H.; Anderson, D.M. Molecular Identification and Toxin Analysis of Alexandrium Spp. in the Beibu Gulf: First Report of Toxic A. Tamiyavanichii in Chinese Coastal Waters. Toxins 2021, 13, 161. [Google Scholar] [CrossRef]
- Lattes, K.; Venegas, P.; Lagos, N.; Lagos, M.; Pedraza, L.; Rodriguez-Navarro, A.J.; García, C. Local Infiltration of Gonyautoxin Is Safe and Effective in Treatment of Chronic Tension-Type Headache. Neurol. Res. 2009, 31, 228–233. [Google Scholar] [CrossRef]
- Molgó, J.; Marchot, P.; Aráoz, R.; Benoit, E.; Iorga, B.I.; Zakarian, A.; Taylor, P.; Bourne, Y.; Servent, D. Cyclic Imine Toxins from Dinoflagellates: A Growing Family of Potent Antagonists of the Nicotinic Acetylcholine Receptors. J. Neurochem. 2017, 142, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.; Murphy, M.; Clausen, J.; Richard, D.; Quilliam, M.; MacKinnon, S.; LaBlanc, P.; Mueller, R.; Pulido, O. Neural Injury Biomarkers of Novel Shellfish Toxins, Spirolides: A Pilot Study Using Immunochemical and Transcriptional Analysis. NeuroToxicology 2003, 24, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Guéret, S.M.; Brimble, M.A. Spiroimine Shellfish Poisoning (SSP) and the Spirolide Family of Shellfish Toxins: Isolation, Structure, Biological Activity and Synthesis. Nat. Prod. Rep. 2010, 27, 1350. [Google Scholar] [CrossRef] [PubMed]
- Otero, A.; Chapela, M.-J.; Atanassova, M.; Vieites, J.M.; Cabado, A.G. Cyclic Imines: Chemistry and Mechanism of Action: A Review. Chem. Res. Toxicol. 2011, 24, 1817–1829. [Google Scholar] [CrossRef] [PubMed]
- Stivala, C.E.; Benoit, E.; Aráoz, R.; Servent, D.; Novikov, A.; Molgó, J.; Zakarian, A. Synthesis and Biology of Cyclic Imine Toxins, an Emerging Class of Potent, Globally Distributed Marine Toxins. Nat. Prod. Rep. 2015, 32, 411–435. [Google Scholar] [CrossRef] [PubMed]
- Stivala, C.E.; Zakarian, A. Studies Toward the Synthesis of Spirolides: Assembly of the Elaborated E-Ring Fragment. Org. Lett. 2009, 11, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Zurhelle, C.; Nieva, J.; Tillmann, U.; Harder, T.; Krock, B.; Tebben, J. Identification of Novel Gymnodimines and Spirolides from the Marine Dinoflagellate Alexandrium ostenfeldii. Mar. Drugs 2018, 16, 446. [Google Scholar] [CrossRef]
- Albuquerque, E.X.; Pereira, E.F.R.; Alkondon, M.; Rogers, S.W. Mammalian Nicotinic Acetylcholine Receptors: From Structure to Function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef]
- Kharrat, R.; Servent, D.; Girard, E.; Ouanounou, G.; Amar, M.; Marrouchi, R.; Benoit, E.; Molgó, J. The Marine Phycotoxin Gymnodimine Targets Muscular and Neuronal Nicotinic Acetylcholine Receptor Subtypes with High Affinity. J. Neurochem. 2008, 107, 952–963. [Google Scholar] [CrossRef]
- Hauser, T.A.; Hepler, C.D.; Kombo, D.C.; Grinevich, V.P.; Kiser, M.N.; Hooker, D.N.; Zhang, J.; Mountfort, D.; Selwood, A.; Akireddy, S.R.; et al. Comparison of Acetylcholine Receptor Interactions of the Marine Toxins, 13-Desmethylspirolide C and Gymnodimine. Neuropharmacology 2012, 62, 2239–2250. [Google Scholar] [CrossRef]
- Munday, R.; Towers, N.R.; Mackenzie, L.; Beuzenberg, V.; Holland, P.T.; Miles, C.O. Acute Toxicity of Gymnodimine to Mice. Toxicon 2004, 44, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Quilliam, M.A.; LeBlanc, P.; Lewis, N.; Gallant, P.; Sperker, S.A.; Ewart, H.S.; MacKinnon, S.L. Investigations into the Toxicology of Spirolides, a Group of Marine Phycotoxins. Toxins 2011, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Burton, I.W.; Cembella, A.D.; Curtis, J.M.; Quilliam, M.A.; Walter, J.A.; Wright, J.L.C. Characterization of Spirolides A, C, and 13-Desmethyl C, New Marine Toxins Isolated from Toxic Plankton and Contaminated Shellfish. J. Nat. Prod. 2001, 64, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Alonso, E.; Otero, P.; Vale, C.; Alfonso, A.; Antelo, A.; Gimenez-Llort, L.; Chabaud, L.; Guillou, C.; Botana, M.L. Benefit of 13-Desmethyl Spirolide C Treatment in Triple Transgenic Mouse Model of Alzheimer Disease: Beta-Amyloid and Neuronal Markers Improvement. CAR 2013, 10, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Makabe, K.; Yamaguchi, K.; Konosu, S.; Wälchli, M.R. Goniodomin a, a Novel Polyether Macrolide from the Dinoflagellate Goniodoma pseudogoniaulax. Tetrahedron Lett. 1988, 29, 1149–1152. [Google Scholar] [CrossRef]
- Furukawa, K.; Sakai, K.; Watanabe, S.; Maruyama, K.; Murakami, M.; Yamaguchi, K.; Ohizumi, Y. Goniodomin A Induces Modulation of Actomyosin ATPase Activity Mediated through Conformational Change of Actin. J. Biol. Chem. 1993, 268, 26026–26031. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Inoue, D.; Matsunaga, K.; Ohizumi, Y.; Ueda, H.; Asano, T.; Murakami, M.; Sato, Y. Goniodomin A, an Antifungal Polyether Macrolide, Exhibits Antiangiogenic Activities via Inhibition of Actin Reorganization in Endothelial Cells. J. Cell. Physiol. 2002, 190, 109–116. [Google Scholar] [CrossRef]
- Mizuno, K.; Nakahata, N.; Ito, E.; Murakami, M.; Yamaguchi, K.; Ohizumi, Y. Goniodomin A, an Antifungal Polyether Macrolide, Increases the Filamentous Actin Content of 1321N1 Human Astrocytoma Cells. J. Pharm. Pharmacol. 1998, 50, 645–648. [Google Scholar] [CrossRef]
- Llewellyn, L.E. Sodium Channel Inhibiting Marine Toxins. In Marine Toxins as Research Tools; Fusetani, N., Kem, W., Eds.; Progress in Molecular and Subcellular Biology; Springer: Berlin/Heidelberg, Germany, 2009; Volume 46, pp. 67–97. ISBN 978-3-540-87892-6. [Google Scholar]
- Kodama, M.; Sato, S.; Sakamoto, S.; Ogata, T. Occurrence of Tetrodotoxin in Alexandrium tamarense, a Causative Dinoflagellate of Paralytic Shellfish Poisoning. Toxicon 1996, 34, 1101–1105. [Google Scholar] [CrossRef]
- Tsuda, K.; Ikuma, S.; Kawamura, M.; Tachikawa, R.; Sakai, K.; Tamura, C.; Amakasu, O. Tetrodotoxin. VII. On the Structures of Tetrodotoxin and Its Derivatives. Chem. Pharm. Bull. 1964, 12, 1357–1374. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Sugimoto, A.; Takai, A.; Yasumoto, T. Effects of Specific Modifications of Several Hydroxyls of Tetrodotoxin on Its Affinity to Rat Brain Membrane. J. Pharmacol. Exp. Ther. 1999, 289, 1688–1696. [Google Scholar] [PubMed]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, Toxicity, Source, Distribution and Detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the Origins and Biosynthesis of Tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Kao, C.Y. Actions of Chiriquitoxin on Frog Skeletal Muscle Fibers and Implications for the Tetrodotoxin/Saxitoxin Receptor. J. Gen. Physiol. 1992, 100, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Denac, H.; Mevissen, M.; Scholtysik, G. Structure, Function and Pharmacology of Voltage-Gated Sodium Channels. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2000, 362, 453–479. [Google Scholar] [CrossRef] [PubMed]
- Hille, B. The Permeability of the Sodium Channel to Organic Cations in Myelinated Nerve. J. Gen. Physiol. 1971, 58, 599–619. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Cellular and Molecular Biology of Voltage-Gated Sodium Channels. Physiol. Rev. 1992, 72, S15–S48. [Google Scholar] [CrossRef]
- Walker, J.R.; Novick, P.A.; Parsons, W.H.; McGregor, M.; Zablocki, J.; Pande, V.S.; Du Bois, J. Marked Difference in Saxitoxin and Tetrodotoxin Affinity for the Human Nociceptive Voltage-Gated Sodium Channel (Na v 1.7). Proc. Natl. Acad. Sci. USA 2012, 109, 18102–18107. [Google Scholar] [CrossRef]
- Lee, C.H.; Ruben, P.C. Interaction between Voltage-Gated Sodium Channels and the Neurotoxin, Tetrodotoxin. Channels 2008, 2, 407–412. [Google Scholar] [CrossRef]
- Hwang, D.; Noguchi, T. Tetrodotoxin Poisoning. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2007; Volume 52, pp. 141–236. ISBN 978-0-12-373711-3. [Google Scholar]
- Lago, J.; Rodríguez, L.; Blanco, L.; Vieites, J.; Cabado, A. Tetrodotoxin, an Extremely Potent Marine Neurotoxin: Distribution, Toxicity, Origin and Therapeutical Uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef]
- Noguchi, T.; Ebesu, J.S.M. Puffer Poisoing: Epidemiologi and Tretment. J. Toxicol. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Saoudi, M.; Abdelmouleh, A.; El Feki, A. Tetrodotoxin: A Potent Marine Toxin. Toxin Rev. 2010, 29, 60–70. [Google Scholar] [CrossRef]
- How, C.-K.; Chern, C.-H.; Huang, Y.-C.; Wang, L.-M.; Lee, C.-H. Tetrodotoxin Poisoning. Am. J. Emerg. Med. 2003, 21, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.K.; Ostman, D.C. Pufferfish Poisoning: Emergency Diagnosis and Management of Mild Human Tetrodotoxication. Ann. Emerg. Med. 1986, 15, 1094–1098. [Google Scholar] [CrossRef]
- Lange, W.R. Puffer Fish Poisoning. Am. Fam. Physician 1990, 42, 1029–1033. [Google Scholar] [PubMed]
- Nieto, F.R.; Cobos, E.J.; Tejada, M.Á.; Sánchez-Fernández, C.; González-Cano, R.; Cendán, C.M. Tetrodotoxin (TTX) as a Therapeutic Agent for Pain. Mar. Drugs 2012, 10, 281–305. [Google Scholar] [CrossRef] [PubMed]
- Hagen, N.A.; Cantin, L.; Constant, J.; Haller, T.; Blaise, G.; Ong-Lam, M.; Du Souich, P.; Korz, W.; Lapointe, B. Tetrodotoxin for Moderate to Severe Cancer-Related Pain: A Multicentre, Randomized, Double-Blind, Placebo-Controlled, Parallel-Design Trial. Pain Res. Manag. 2017, 2017, 7212713. [Google Scholar] [CrossRef]
- Galasso, C.; Nuzzo, G.; Brunet, C.; Ianora, A.; Sardo, A.; Fontana, A.; Sansone, C. The Marine Dinoflagellate Alexandrium minutum Activates a Mitophagic Pathway in Human Lung Cancer Cells. Mar. Drugs 2018, 16, 502. [Google Scholar] [CrossRef]
- Yamasaki, Y.; Katsuo, D.; Nakayasu, S.; Salati, C.; Duan, J.; Zou, Y.; Matsuyama, Y.; Yamaguchi, K.; Oda, T. Purification and Characterization of a Novel High Molecular Weight Exotoxin Produced by Red Tide Phytoplankton, Alexandrium tamarense. J. Biochem. Mol. Toxicol. 2008, 22, 405–415. [Google Scholar] [CrossRef]
- Sansone, C.; Nuzzo, G.; Galasso, C.; Casotti, R.; Fontana, A.; Romano, G.; Ianora, A. The Marine Dinoflagellate Alexandrium andersoni Induces Cell Death in Lung and Colorectal Tumor Cell Lines. Mar. Biotechnol. 2018, 20, 343–352. [Google Scholar] [CrossRef]
- De Vera, C.; Díaz Crespín, G.; Hernández Daranas, A.; Montalvão Looga, S.; Lillsunde, K.-E.; Tammela, P.; Perälä, M.; Hongisto, V.; Virtanen, J.; Rischer, H.; et al. Marine Microalgae: Promising Source for New Bioactive Compounds. Mar. Drugs 2018, 16, 317. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Marie, D.; Szymczak, J.; Toullec, J.; Bigeard, E.; Sourisseau, M.; Le Gac, M.; Guillou, L.; Jauzein, C. Dinophyceae Can Use Exudates as Weapons against the Parasite Amoebophrya sp. (Syndiniales). ISME Commun. 2021, 1, 34. [Google Scholar] [CrossRef] [PubMed]
- Lassudrie, M.; Soudant, P.; Nicolas, J.-L.; Fabioux, C.; Lambert, C.; Miner, P.; Le Grand, J.; Petton, B.; Hégaret, H. Interaction between Toxic Dinoflagellate Alexandrium catenella Exposure and Disease Associated with Herpesvirus OsHV-1 μVar in Pacific Oyster Spat Crassostrea Gigas. Harmful Algae 2015, 45, 53–61. [Google Scholar] [CrossRef]
- Lauritano, C.; Martín, J.; De La Cruz, M.; Reyes, F.; Romano, G.; Ianora, A. First Identification of Marine Diatoms with Anti-Tuberculosis Activity. Sci. Rep. 2018, 8, 2284. [Google Scholar] [CrossRef]
- Chen, L.; Giesy, J.P.; Adamovsky, O.; Svirčev, Z.; Meriluoto, J.; Codd, G.A.; Mijovic, B.; Shi, T.; Tuo, X.; Li, S.-C.; et al. Challenges of Using Blooms of Microcystis Spp. in Animal Feeds: A Comprehensive Review of Nutritional, Toxicological and Microbial Health Evaluation. Sci. Total Environ. 2021, 764, 142319. [Google Scholar] [CrossRef]
- Bashir, F.; Bashir, A.; Bouaïcha, N.; Chen, L.; Codd, G.A.; Neilan, B.; Xu, W.-L.; Ziko, L.; Rajput, V.D.; Minkina, T.; et al. Cyanotoxins, Biosynthetic Gene Clusters, and Factors Modulating Cyanotoxin Biosynthesis. World J. Microbiol. Biotechnol. 2023, 39, 241. [Google Scholar] [CrossRef]
Figure 1.
Life cycle of Alexandrium spp. The life cycle of Alexandrium spp. is divided into two phases, the planktonic vegetative phase with asexual reproduction and the dormancy phase in the sediment in the form of cysts resulting in sexual reproduction. The duration of each phase is influenced by the conditions in the surrounding environment (temperature, pH, nutrients, etc.). The cycle starts from the germination of the cyst in planomeiocyte (1.) that under favorable conditions for growth undertakes asexual reproduction by demoschisis giving rise to the vegetative phase of the cells (2.1; 3.1). Under not optimal conditions of growth, it gives rise to the sexual reproduction forming two haploid gametes (2.) that subsequently merge (3.), forming the diploid and biflagellated planozygote (4.) which forms a new cyst called hypnozygote (5.).
Figure 1.
Life cycle of Alexandrium spp. The life cycle of Alexandrium spp. is divided into two phases, the planktonic vegetative phase with asexual reproduction and the dormancy phase in the sediment in the form of cysts resulting in sexual reproduction. The duration of each phase is influenced by the conditions in the surrounding environment (temperature, pH, nutrients, etc.). The cycle starts from the germination of the cyst in planomeiocyte (1.) that under favorable conditions for growth undertakes asexual reproduction by demoschisis giving rise to the vegetative phase of the cells (2.1; 3.1). Under not optimal conditions of growth, it gives rise to the sexual reproduction forming two haploid gametes (2.) that subsequently merge (3.), forming the diploid and biflagellated planozygote (4.) which forms a new cyst called hypnozygote (5.).
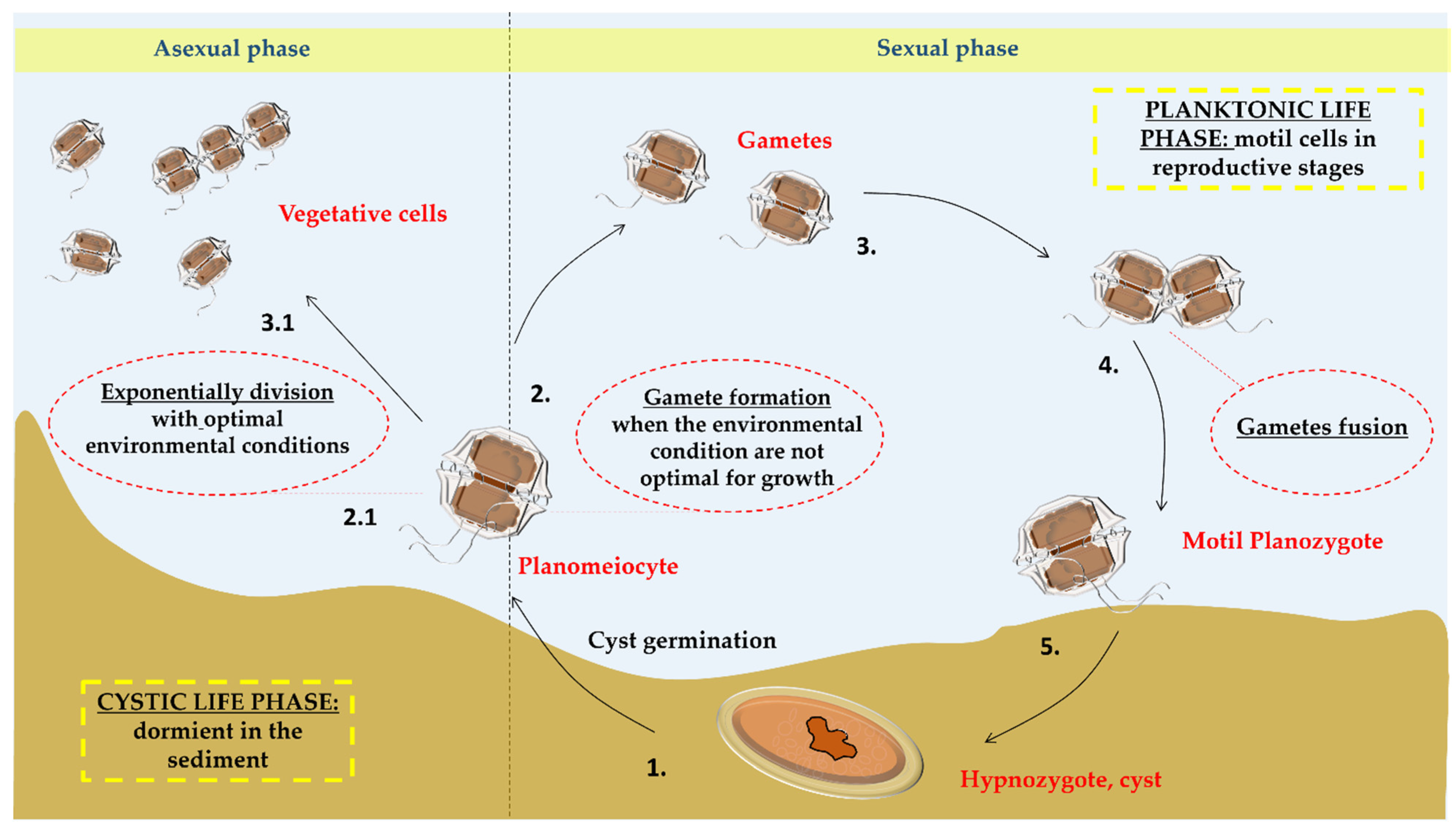
Figure 2.
Occurrence of Alexandrium spp. (light blue squares) and paralytic shellfish poisoning (PSP) events (orange). Species occurrences are from 1904, while PSP records from 1938. The map was built using the Harmful Algal Information System (HAIS) available at https://data.hais.ioc-unesco.org/ (accessed on 11 August 2023).
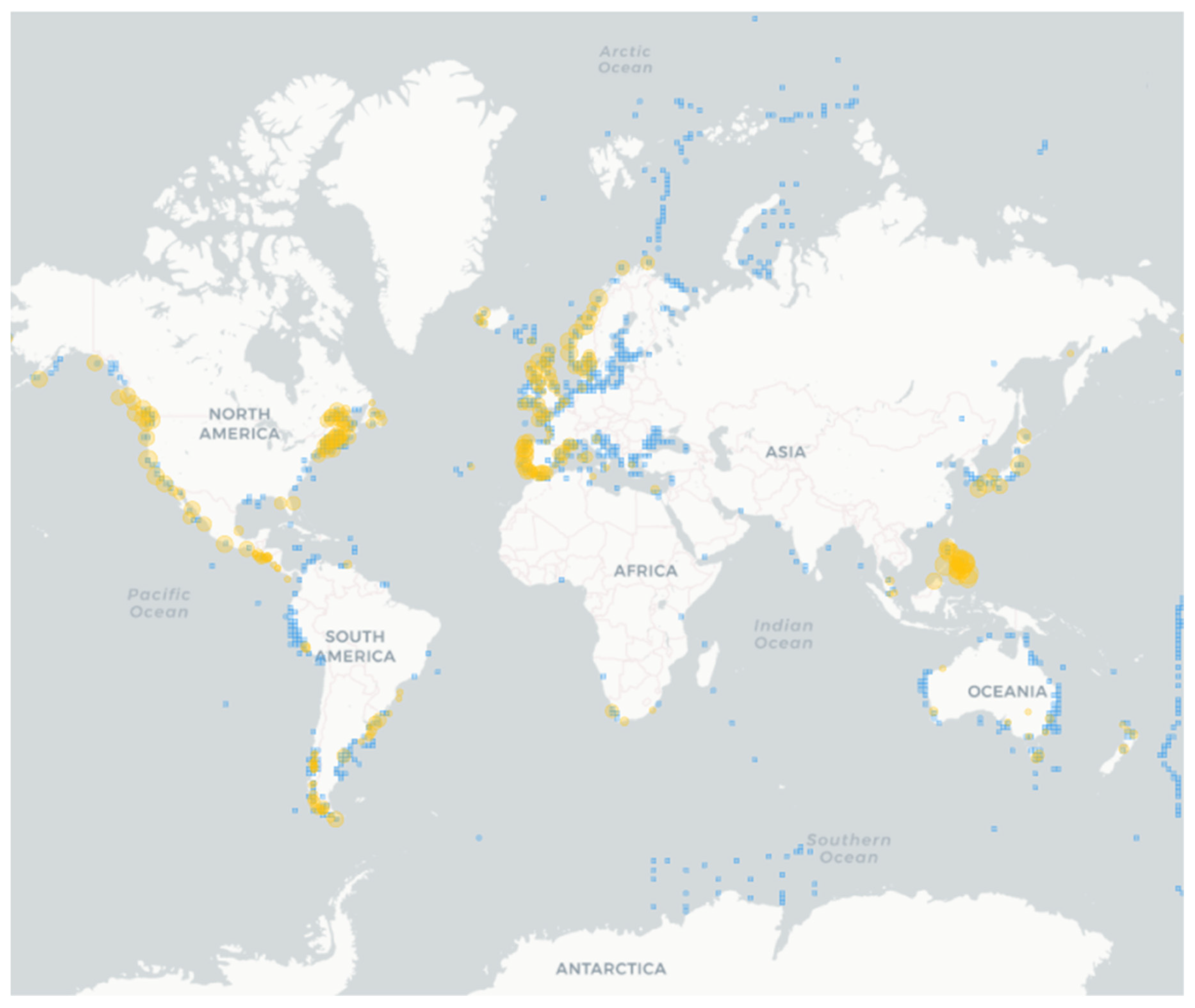
Figure 2.
Occurrence of Alexandrium spp. (light blue squares) and paralytic shellfish poisoning (PSP) events (orange). Species occurrences are from 1904, while PSP records from 1938. The map was built using the Harmful Algal Information System (HAIS) available at https://data.hais.ioc-unesco.org/ (accessed on 11 August 2023).
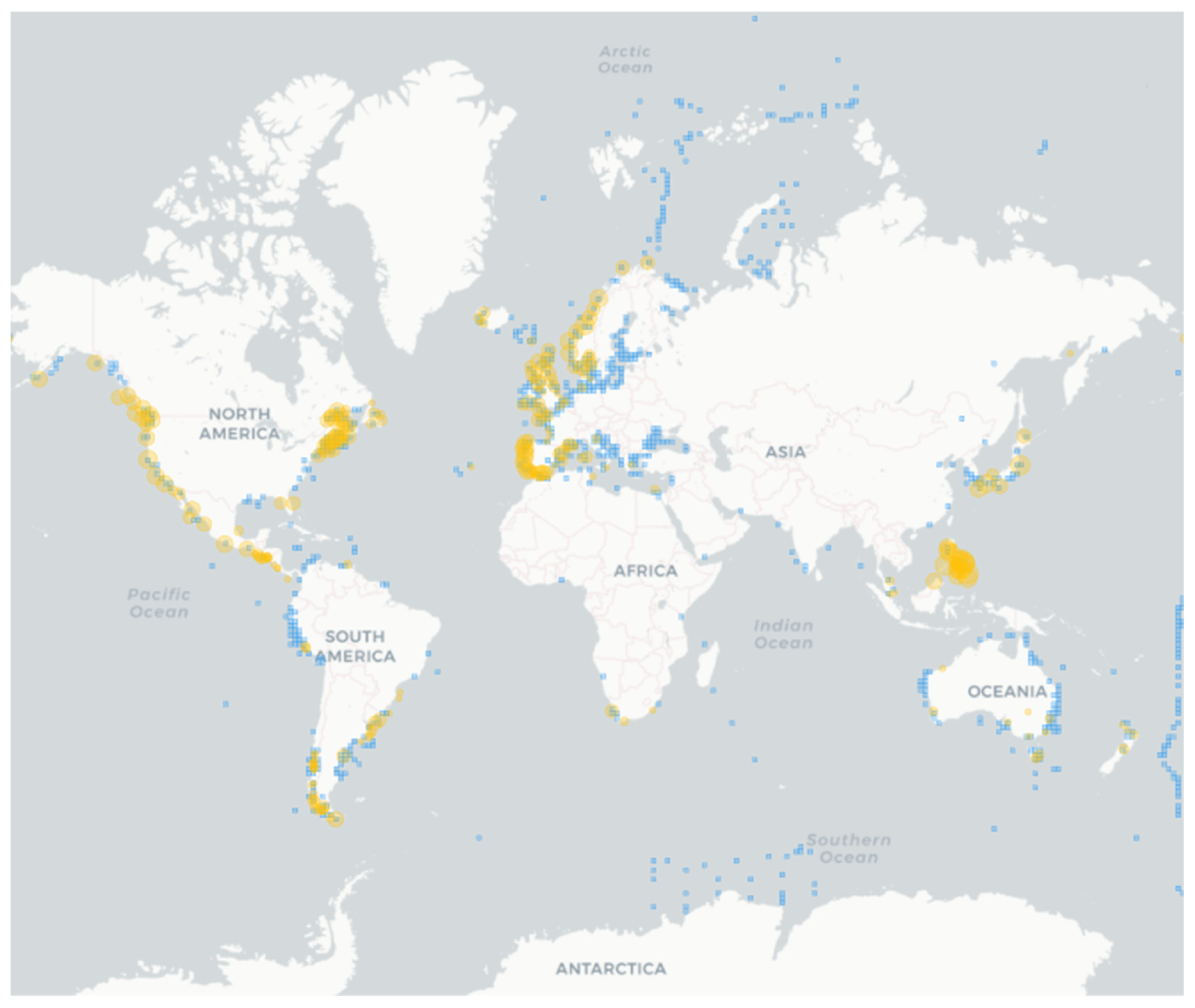
Figure 3.
Chemical structures of: (A) Saxitoxin [72]; (B) Spirolide A [73]; (C) Spirolide C [74]; (D) Gymnodimine A [75]; (E) 13-desmeethyl spirolide C [76]; (F) Goniodomin A [77]; (G) tetrodoxin [78].


{kind=link}
{kind=link}
{kind=link}
Table 1.
List of known toxic Alexandrium species. Species names are as in AlgaeBase [17] and only accepted names are shown.
Table 1.
List of known toxic Alexandrium species. Species names are as in AlgaeBase [17] and only accepted names are shown.
| Species | Toxins | Effects | References |
|---|---|---|---|
| A. affine (H. Inoue and Y. Fukuyo) Balech | Saxitoxins (only genes found) | Not evaluated | [41] |
| A. andersonii Balech * | Saxitoxins (GTX2, NEO, STX) | Inhibition of voltage-gated sodium channels, hemolytic activity | [42,43] |
| A. catenella (Whedon and Kofoid) Balech | Saxitoxins (GTX1, GTX4, NEO, STX) | PSP | [44,45,46] |
| A. fundyense Balech | Saxitoxins (GTX3, Neo, STX) | PSP | [47] |
| A. hiranoi Kita and Fukuyo | Goniodomin A | Cytotoxic, disturbs the actomyosin ATPase activity in diverse cell types | [48,49] |
| A. insuetum Balech | Not characterized | PSP | [50] |
| A. leei Balech | Not characterized, but neither PSTs nor spirolides | Mortality of finfish and rotifers | [51] |
| A. minutum Halim * | Saxitoxins (C1, C2, NEO, GTX1, GTX2, GTX3, GTX4) | PSP | [23,52] |
| A. monilatum (Howell) Balech | Goniodomin A | Highly toxic to fish and shellfish | [53,54] |
| A. ostenfeldii (Paulsen) Balech and Tangen | Saxitoxins (GTX3, GTX5); spirolides (A, B, C2, D2) | PSP (saxitoxins); toxicity to shellfish (spirolides) | [55,56,57] |
| A. peruvianum (Balech and Mendiola) | Hemolytic substances, Saxitoxins (B1, C1, C2, GTX2, GTX3, B1) Spirolides (C and D) | PSP | [58,59] |
| A. pseudogonyaulax (Biecheler) Horiguchi ex K. Yuki and Y. Fukuyo | Goniodomin A | Fish mortality | [60,61] |
| A. tamarense (Lebour) Balech * | Saxitoxins (C2, NEO, GTX3, GTX4) | PSP | [62,63] |
| A. tamiyavanichii Balech | Saxitoxins (C1, C2, GTX1, GTX2, GTX3, GTX5, STX) | PSP | [64] |
| A. tamutum | Saxitoxins (only genes found) | Antiproliferative activity on melanoma (A2058) and normal human lung fibroblast (MRC-5) | [26,32] |
| A. taylori Balech | Goniodomin A, Hemolytic substances | Effects on oyster larvae, sea-monkey mortality | [65,66] |
PSP = paralytic shellfish poisoning; PSTs = paralytic shellfish toxins. * only some strains were found to be toxic.
Table 2.
Bioactivities reported for Alexandrium species, mechanisms of action, cell lines or model organisms used. BIRC3 stand for Baculoviral IAP Repeat Containing 3; TRAF2 for TNF receptor-associated factor 2; DR3 for Death receptor 3; TNF for tumor necrosis factor; TNFS8 for TNF superfamily member 8; FASLG for Fas ligand; Gadd45 for Growth Arrest and DNA Damage; Foxl1 for forkhead box L1; Atg12 for autophagy-related protein 12/gene; ATP6V1G2 for ATPase H+ Transporting V1 Subunit G2; BNIP3 for BCL2/Adenovirus E1B19 kDa Interacting Protein 3; PINK1 for PTEN induced putative kinase1; NAA for N-acetyl aspartate.
Table 2.
Bioactivities reported for Alexandrium species, mechanisms of action, cell lines or model organisms used. BIRC3 stand for Baculoviral IAP Repeat Containing 3; TRAF2 for TNF receptor-associated factor 2; DR3 for Death receptor 3; TNF for tumor necrosis factor; TNFS8 for TNF superfamily member 8; FASLG for Fas ligand; Gadd45 for Growth Arrest and DNA Damage; Foxl1 for forkhead box L1; Atg12 for autophagy-related protein 12/gene; ATP6V1G2 for ATPase H+ Transporting V1 Subunit G2; BNIP3 for BCL2/Adenovirus E1B19 kDa Interacting Protein 3; PINK1 for PTEN induced putative kinase1; NAA for N-acetyl aspartate.
| Species | Sampling Location and Culture Conditions | Active Compounds/Extracts/Fractions | Bioactivities | In Vivo/In Vitro | Mechanism | References |
|---|---|---|---|---|---|---|
| Alexandrium andersonii | Mediterranean Sea [137]. - | Fraction B (digalactosyl diacyl glycerol and sulfoquinovosyl diacyl glycerol, were the predominant metabolites) | Cytotoxic (Anti-cancer) | In vitro investigation on: A549 cells | Down-regulation of anti-apoptotic molecules (BIRC3 and TRAF2) and up-regulation of death receptors (CD27, DR3, TNF, and TNFS8) | [133] |
| Alexandrium andersonii | Mediterranean Sea. - | Fraction D (rich in sterols and triacylglycerols) | Cytotoxic (Anti-cancer) | In vitro investigation on: HT29 cells | Induction of cell death with over expression of FASLG and TNF genes Up-regulation of Gadd45 alpha and gamma together with Foxl1 genes | [133] |
| Alexandrium catenella | Thau lagoon, France. Cultivated in L1 medium | - | Anti-viral | Herpes virus OsHV-1 μvar Oyster affected | Antagonist relationship that is established between the virus and A. catenella | [136] |
| Alexandrium minutum | Gulf of Naples. Cultivated in Keller medium | Fraction 3B (rich in carbohydrates (94%) and protein (4%)) | Cytotoxic (Anti-cancer) | In vitro investigation on: PC-3, HT29, A549 cells | Up-regulation of autophagy-related protein Atg12 with a consequent increase in ATP6V1G2 and BNIP3 Up-regulation of mitophagy genes PINK1 and Parkin gene | [131] |
| Alexandrium minutum | Coastal marine waters of the NE Atlantic Ocean. Cultivated in F/2 medium, 27 of salinity (Guillard’s Marine Water Enrichment Solution) and Keller medium, 35 of salinity | A. minutum exudates | Anti-parasite | Anti-parasite activity against Amoebophrya sp. | Permeation of the membranes of Amoebophrya sp. dinospores, leading to the loss of dinofluorescence associated in turn with the loss of virulence of the parasite in culture | [135] |
| Alexandrium ostenfeldii | Coastal marine waters of the NE Atlantic Ocean. Cultivated in F/2 medium, 27 of salinity (Guillard’s Marine Water Enrichment Solution) and Keller medium, 35 of salinity | 13-Desmethyl Spirolide C | Anti-Alzheimer | In vivo investigation in mice 3xTg-AD | Increase in NAA levels; Increase in synaptophytin and decrease in intracellular amyloid beta in the hypothalamus | [105] |
| Alexandrium sp. | - | Goniodomin A | Anti-angiogenic | - | Induction of morphological changes in human brain astrocytoma cells. Inhibition of ATPase activities of atrial myofibrils, natural actomyosin, and reconstitution of actomyosin. | [107,109] |
| Alexandrium sp. | - | Saxitoxin, neosaxitoxin, decarbamoyl saxitoxin, and tetrodotoxin | Rat sciatic nerve block (local anesthesia) | In vivo; evaluation of lethality, onset and duration of action for thermal analgesia (hot-plate testing), and motor block (weight-bearing) | Analgesia for 60 min with neosaxitoxin at 34 ± 2 micromol/L, saxitoxin at 58 ± 3 micromol/L, TTX at 92 ± 5 micromol/L, and decarbamoyl saxitoxin at 268 ± 8 micromol/L | [89] |
| Alexandrium sp. | - | Gonyautoxins 2/3 epimers | Treatment of Chronic headache | In vivo: investigation on 27 patients affected of chronic headache | Reversible block of the voltage-gated sodium channels at axonal level. | [91] |
| Alexandrium sp. | - | Tetrodoxins | Analgesic effects. | In vivo: subcutaneous injection of 30 µg. | Blockage of one subclass of sodium channels, NaV 1.7. | [130] |
| Alexandrium tamarense | Hiroshima Bay, Japan. Cultivated in ESM (Erd-Schreiber modified) medium pH8.2 | Polysaccharide-based AT-toxin | Cytotoxic (Anti-cancer) | In vitro investigation on: HeLa, Vero, XC, CHO, and U937 cells | Induction of cell death in U937 by DNA fragmentation | [132] |
| Alexandrium tamarense | Cultivated in Keller medium | Total Extract | Anti-proliferative (Anti-cancer) | In vitro investigation on: MCF-7, MCF-10A, PC-3, and LNCaP cells | - | [134] |
| Alexandrium tamarense | Cultivated in Keller medium | Total Extract | Apoptotic (Anti-cancer) | In vitro investigation on: HepG2 cells. | - | [134] |
| Alexandrium tamutum, A. andersonii, A. minutum | Mediterranean Sea. Cultivated in Keller medium | Total Extract | Anti-proliferative (Anti-cancer) | In vitro on melanoma A2058 and MRC-5 cells | - | [32] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Montuori, E.; De Luca, D.; Penna, A.; Stalberga, D.; Lauritano, C. Alexandrium spp.: From Toxicity to Potential Biotechnological Benefits. Mar. Drugs 2024, 22, 31. https://doi.org/10.3390/md22010031
AMA Style
Montuori E, De Luca D, Penna A, Stalberga D, Lauritano C. Alexandrium spp.: From Toxicity to Potential Biotechnological Benefits. Marine Drugs. 2024; 22(1):31. https://doi.org/10.3390/md22010031
Chicago/Turabian StyleMontuori, Eleonora, Daniele De Luca, Antonella Penna, Darta Stalberga, and Chiara Lauritano. 2024. "Alexandrium spp.: From Toxicity to Potential Biotechnological Benefits" Marine Drugs 22, no. 1: 31. https://doi.org/10.3390/md22010031
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.