
Two New Components from an Association of Marine Sponges Poecillastra sp. and Jaspis sp. and Their Inhibitory Effects on Biomarkers for Benign Prostatic Hyperplasia

Abstract
:1. Introduction
2. Results and Discussion
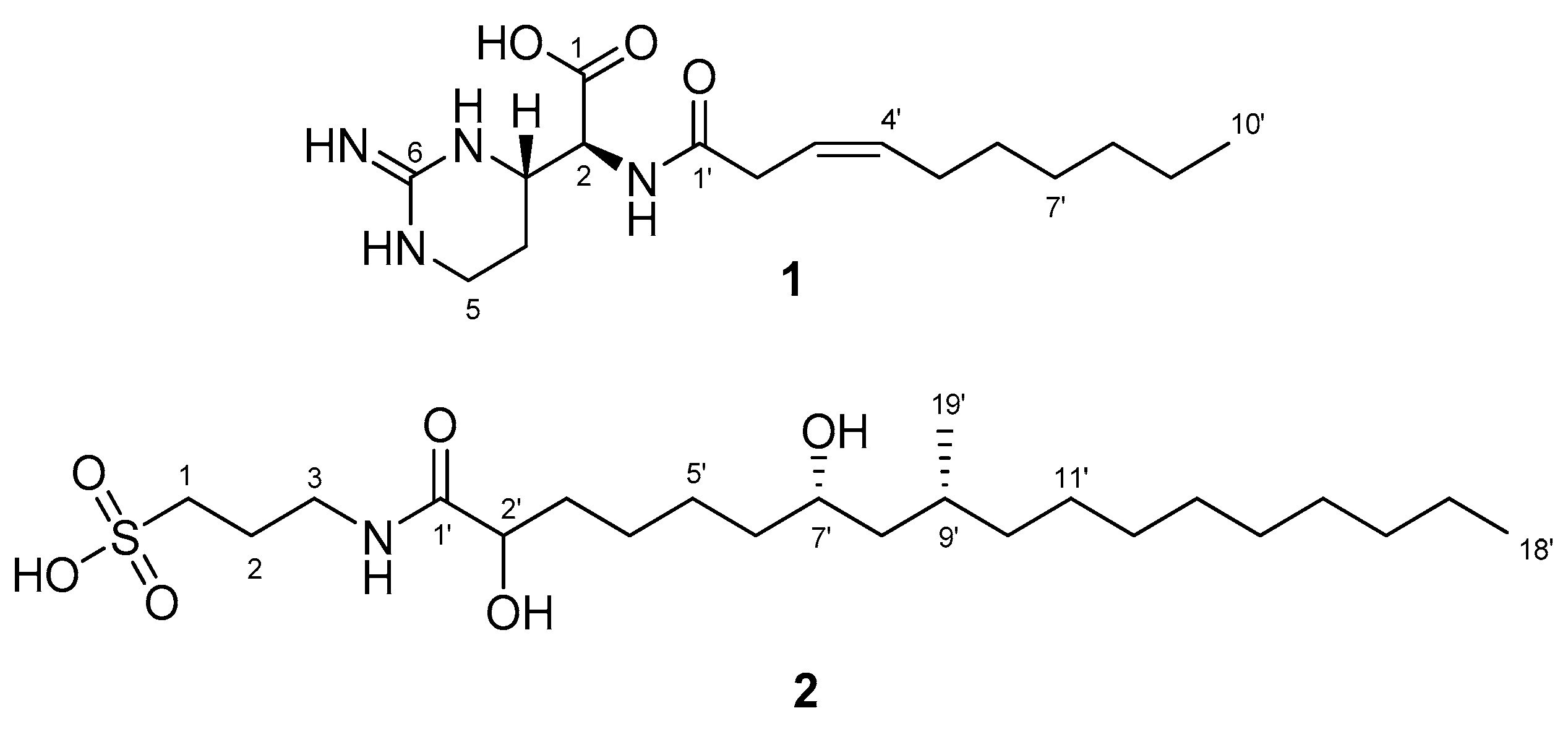
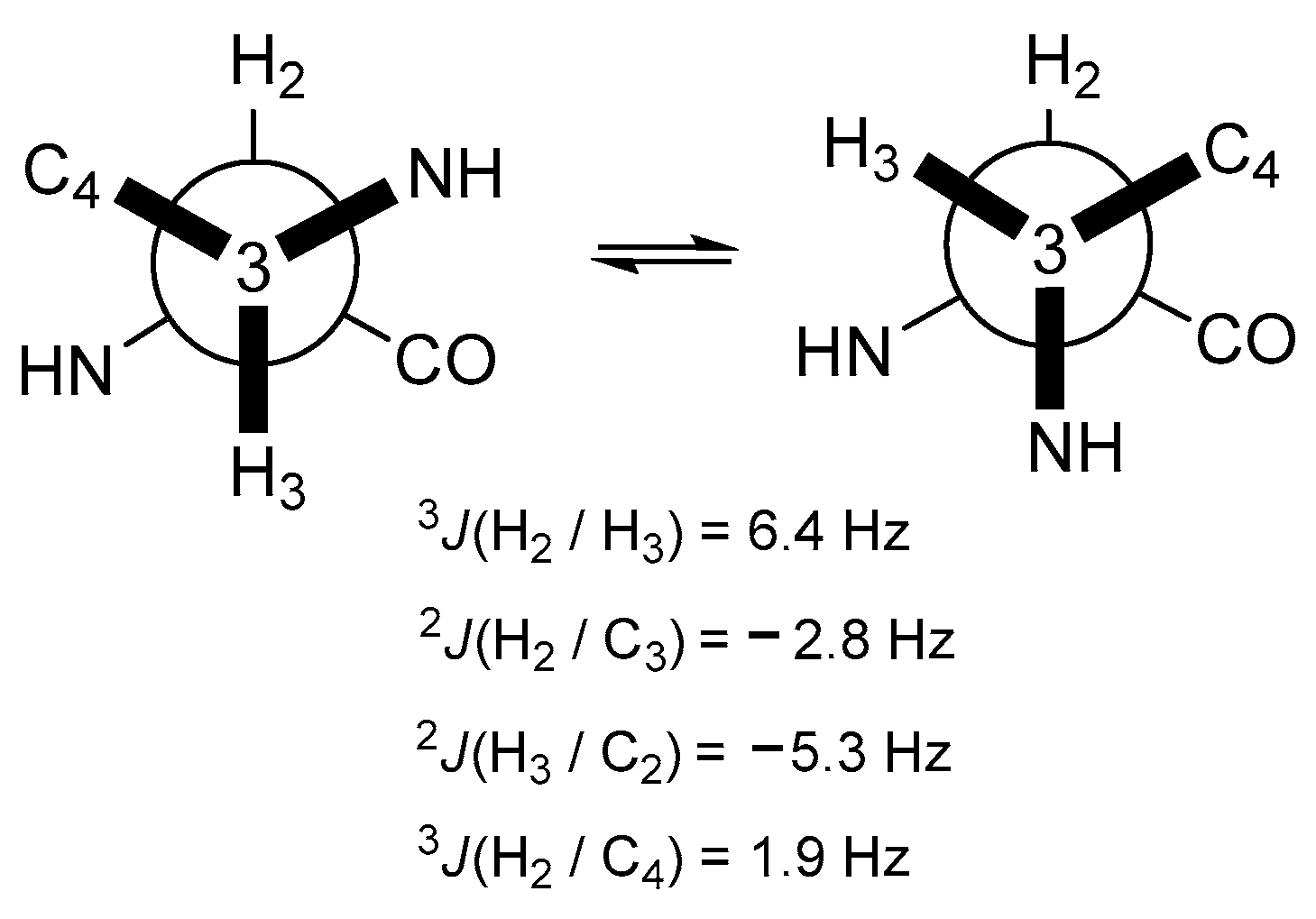
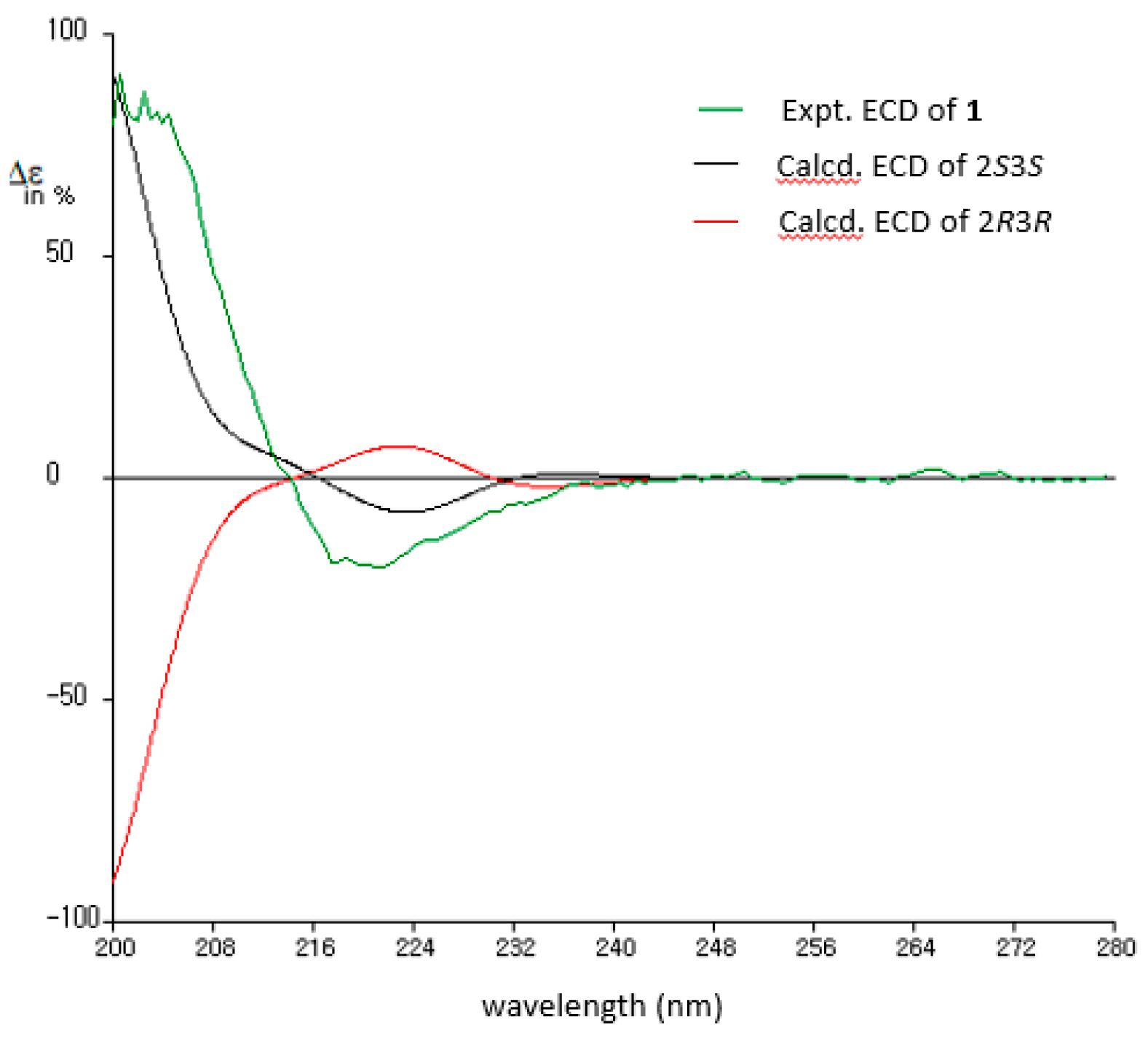
2.1. Structural Determination of Compounds 1 and 2
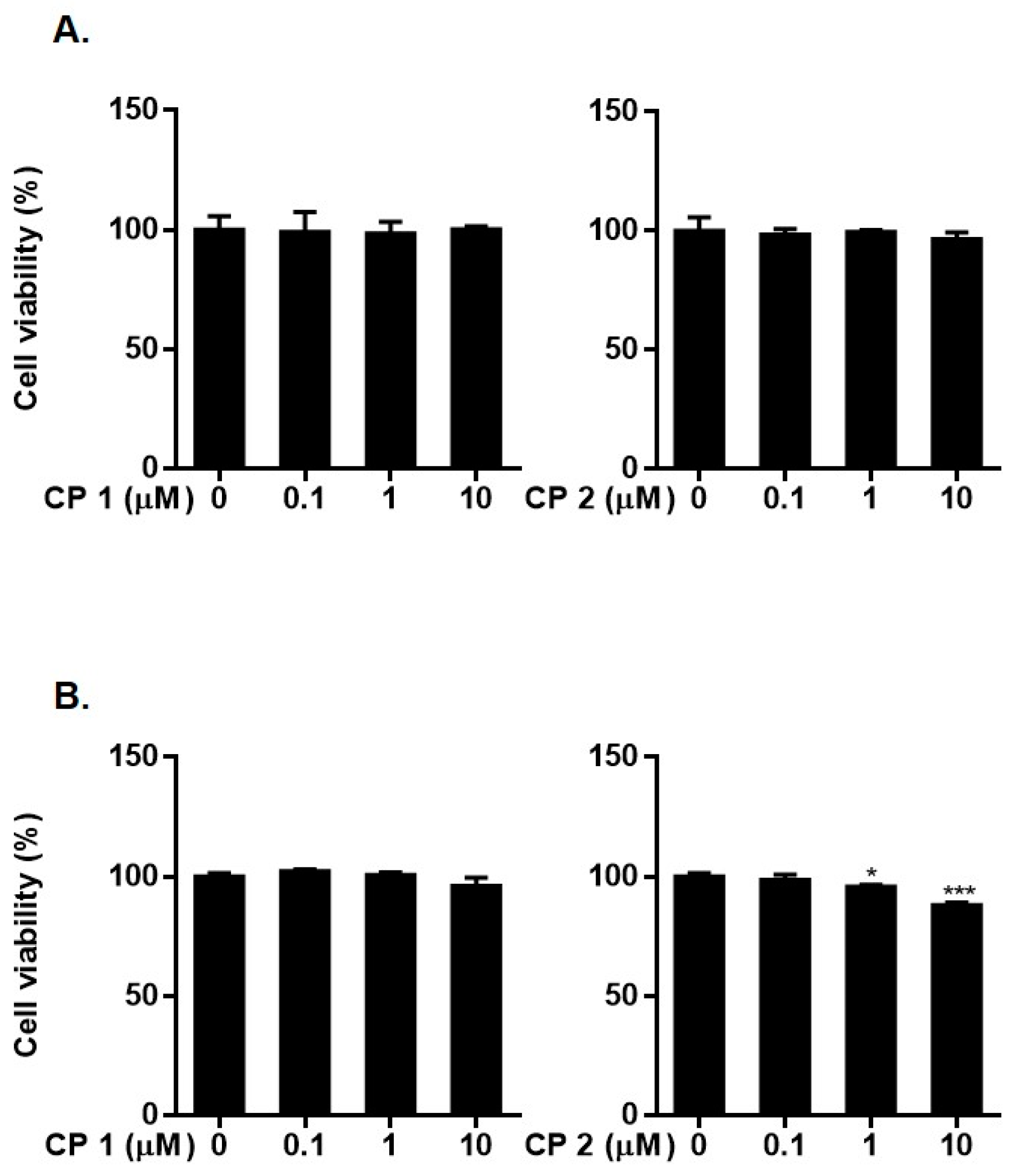
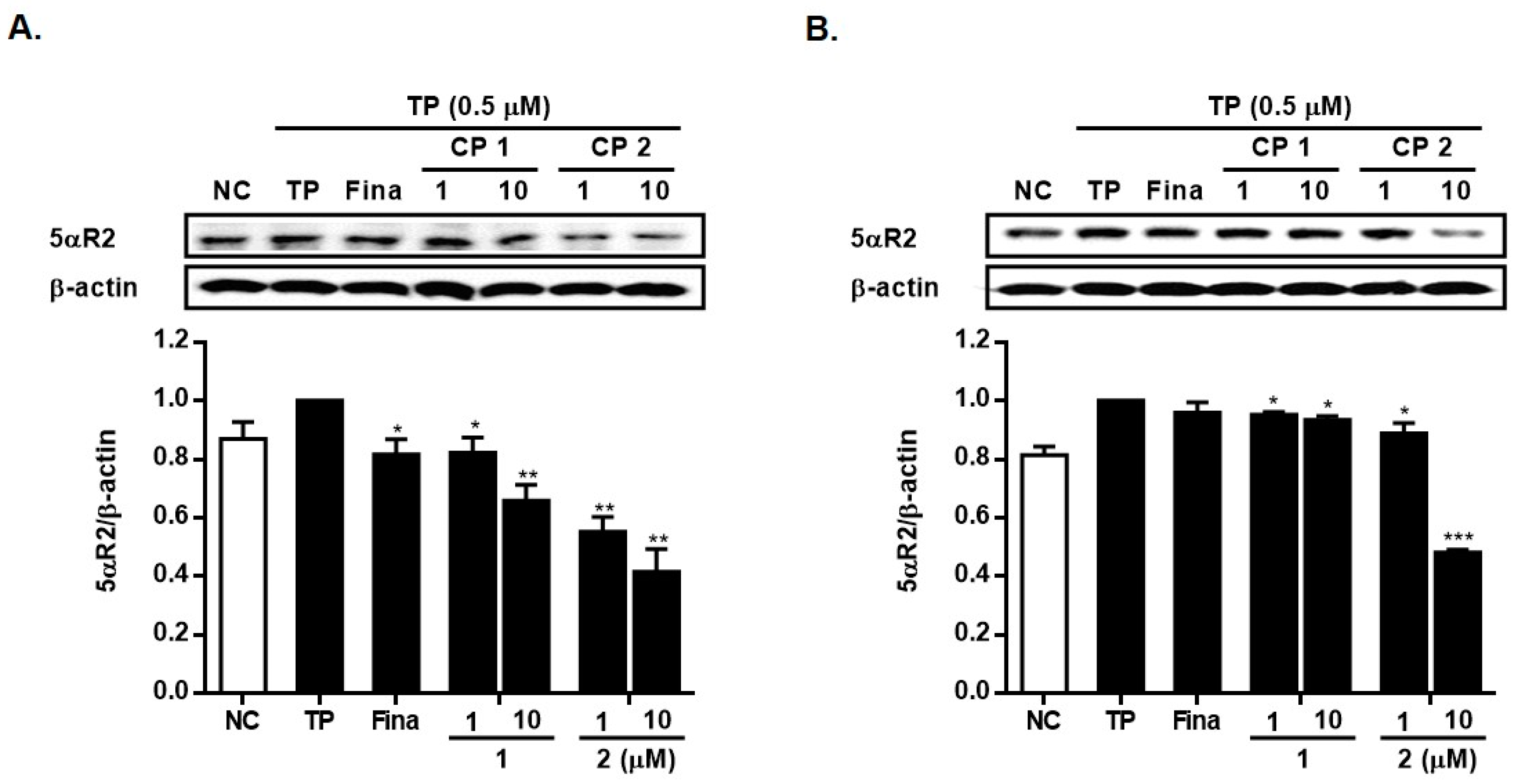
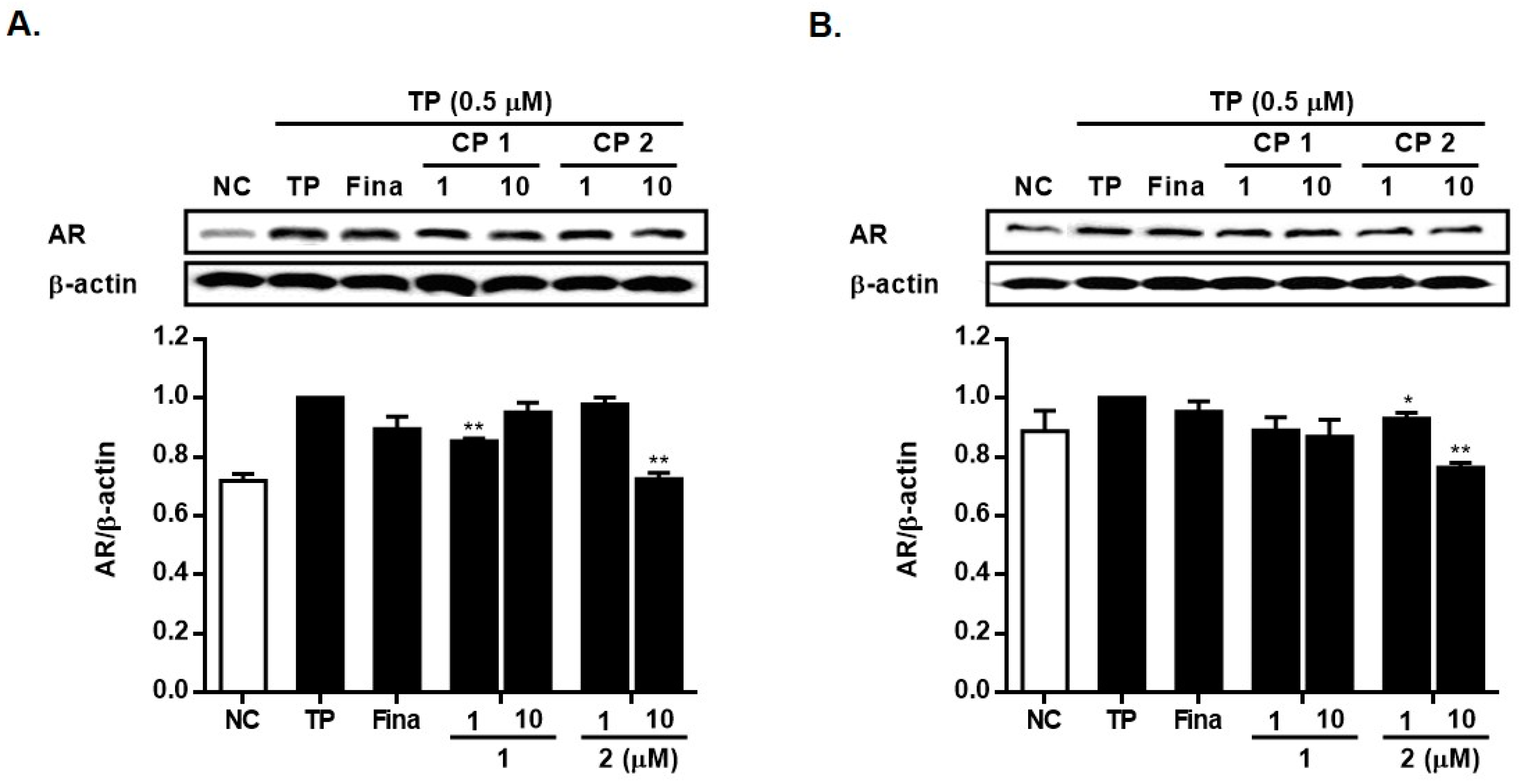
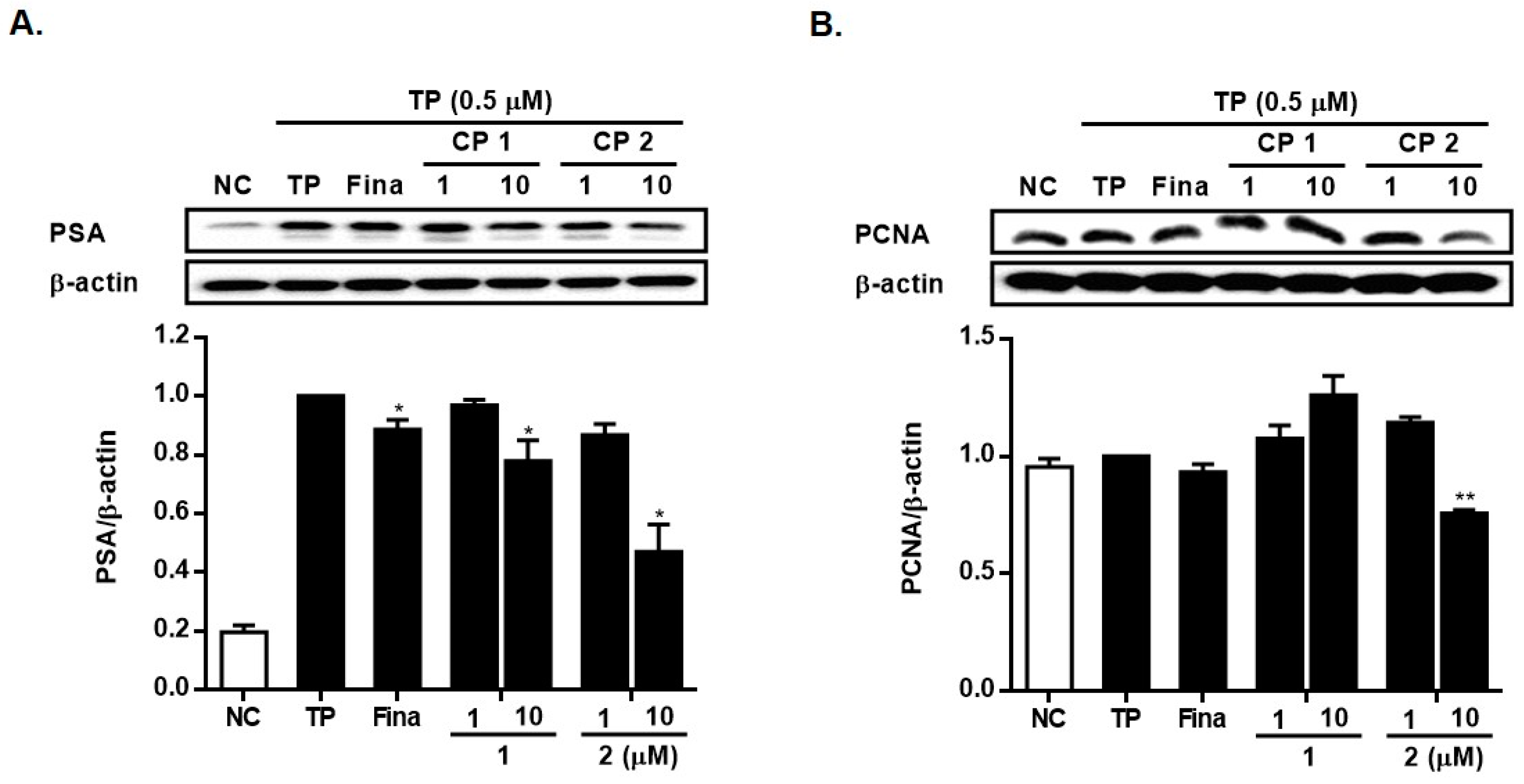
2.2. Biological Activity of Compounds 1 and 2
3. Materials and Methods
3.1. General Experiment Procedures
3.2. Animal Material
3.3. Isolation of Compounds 1 and 2
3.4. ECD Calculation of Compound 1
3.5. Cell Cultures
3.6. Cell Viability Assay
3.7. Western Blotting Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roehrborn, C.G. Benign prostatic hyperplasia: An overview. Rev. Urol. 2005, 7, S3–S14. [Google Scholar]
- Briganti, A.; Capitanio, U.; Suardi, N.; Gallina, A.; Salonia, A.; Bianchi, M.; Tutolo, M.; Di Girolamo, V.; Guazzoni, G.; Rigatti, P.; et al. Benign prostatic hyperplasia and its aetiologies. Eur. Urol. Suppl. 2009, 8, 865–871. [Google Scholar] [CrossRef]
- Gandaglia, G.; Briganti, A.; Gontero, P.; Mondaini, N.; Novara, G.; Salonia, A.; Sciarra, A.; Montorsi, F. The role of chronic prostatic inflammation in the pathogenesis and progression of benign prostatic hyperplasia (bph). BJU Int. 2013, 112, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Governa, P.; Giachetti, D.; Biagi, M.; Manetti, F.; De Cico, L. Hypothesis on Serenoa repens (Bartram) small extract inhibition of prostatic 5α-reductase through an in silico approach on 5α-reductase x-ray structure. Peer J. 2016, 4, e2698. [Google Scholar] [CrossRef] [PubMed]
- Jena, A.K.; Vasisht, K.; Sharma, N.; Kaur, R.; Dhingra, M.S.; Karan, M. Amelioration of testosterone induced benign prostatic hyperplasia by Prunus species. J. Ethnopharmacol. 2016, 190, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Mansoori, B.; Aghapour, M.; Baradaran, B. Urtica dioica dichloromethane extract induce apoptosis from intrinsic pathway on human prostate cancer cells (PC3). Cell. Mol. Biol. 2016, 62, 78–83. [Google Scholar] [PubMed]
- Sugawara, K.; Kanki, D.; Watanabe, R.; Matsushima, R.; Lse, Y.; Yokose, H.; Morii, Y.; Yamawaki, N.; Ninomiya, A.; Okada, S.; et al. Aciculitin D, a cytotoxic heterodetic cyclic peptide from a Poecillastra sp. marine sponge. Tetrahedron 2022, 119, 132859. [Google Scholar] [CrossRef]
- Tang, S.; Deng, Z.; Proksch, P.; Lin, W. Jaspolides G and H, unique bisisomalaricanes from the Chinese marine sponge Jaspis sp. Tetrahedron Lett. 2007, 48, 5443–5447. [Google Scholar] [CrossRef]
- Shin, J.; Rho, J.-R.; Seo, Y.; Lee, H.-S.; Cho, K.W.; Kwon, H.J.; Sim, C.J. Wondonins A and B, new bis (dihydroxystyryl) imidazoles from a two-sponge association. Tetrahedron Lett. 2001, 42, 1965–1968. [Google Scholar] [CrossRef]
- Chang, Y.H.; Shin, D.; Na, Z.; Lee, H.-S.; Kim, D.-D.; Oh, K.-B.; Shin, J. Dihydroxystyrene metabolites from an association of the sponges Poecillastra wondoensis and Jaspis sp. J. Nat. Prod. 2008, 71, 779–783. [Google Scholar] [CrossRef]
- Shinde, P.B.; Lee, Y.M.; Dang, H.T.; Hong, J.; Lee, C.-O.; Jung, J.H. Cytotoxic bromotyrosine derivatives from a two-sponge association of Jaspis sp. and Poecillastra sp. Bioorganic Med. Chem. Lett. 2008, 18, 6414–6418. [Google Scholar] [CrossRef] [PubMed]
- Büschleb, M.; Granitzka, M.; Stalke, D.; Cucho, C. A biomimetic domino reaction for the concise synthesis of capreomycidine and epicapreomycidine. Amino Acids 2012, 43, 2313–2328. [Google Scholar] [CrossRef]
- Caltagirone, C.; Ferrannini, L.; Marchionni, N.; Nappi, G.; Scapagnini, G.; Trabucchi, M. The potential protective effect of tramiprosate(homotaurine) against Alzheimer’s disease: A review. Aging Clin. Exp. Res. 2012, 24, 580–587. [Google Scholar] [PubMed]
- Rittmaster, R.S. 5α-reductase inhibitors in benign prostatic hyperplasia and prostate cancer risk reduction. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 389–402. [Google Scholar] [CrossRef]
- Bartsch, G.; Rittmaster, R.S.; Klocker, H. Dihydrotestosterone and the concept of 5α-reductase inhibition in human benign prostatic hyperplasia. Eur. Urol. 2000, 37, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Boyle, P.; Robertson, C.; Lowe, F.; Roehrborn, C. Updated meta-analysis of clinical trials of Serenoa repens extract in the treatment of symptomatic benign prostatic hyperplasia. BJU Int. 2004, 93, 751–756. [Google Scholar] [CrossRef]
- Izumi, K.; Mizokami, A.; Lin, W.-J.; Lai, K.-P.; Chang, C. Androgen receptor roles in the development of benign prostate hyperplasia. Am. J. Pathol. 2013, 182, 1942–1949. [Google Scholar] [CrossRef]
- Bello, D.; Webber, M.M.; Kleinman, H.K.; Wartinger, D.D.; Rhim, J.S. Androgen responsive adult human prostatic epithelial cell lines immortalized by human papillomavirus 18. Carcinogenesis 1997, 18, 1215–1223. [Google Scholar] [CrossRef]
- Silva, I.S.; Morsch, D.M.; Urnauer, L.; Spritzer, P.M. Androgen-induced cell growth and c-myc expression in human non-transformed epithelial prostatic cells in primary culture. Endocr. Res. 2001, 27, 153–169. [Google Scholar] [CrossRef]
- Schroeder, F.H.; Westerhof, M.; Bosch, R.J.; Kurth, K.H. Benign prostatic hyperplasia treated by castration or the LH-RH analogue buserelin: A report on 6 cases. Eur. Urol. 1986, 12, 318–321. [Google Scholar] [CrossRef]
- Bianchi, S.; Gravina, G.; Podestà, A.; Barletta, D.; Franchi, F.; Kicovic, P.; Luisi, M. Treatment of complicated benign prostatic hyperplasia with LHRH-analogues in aged patients. Int. J. Androl. 1989, 12, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P. The nuclear receptor superfamily: The second decade. Cell 1995, 8, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Tsui, K.H.; Wu, L.L.; Chang, P.L.; Hsieh, M.L.; Juang, H.H. Identifying the combination of the transcriptional regulatory sequences on prostate specific antigen and human glandular kallikrein genes. J. Urol. 2004, 172, 2029–2034. [Google Scholar] [CrossRef] [PubMed]
- Hehre, W.; Klunzinger, P.; Deppmeier, B.; Driessen, A.; Uchida, N.; Hashimoto, M.; Fukushi, E.; Takara, Y. Efficient protocol for accurately calculating 13C chemical shifts of conformationally flexible natural products: Scope, assessment, and limitations. J. Nat. Prod. 2019, 82, 2299–2306. [Google Scholar] [CrossRef] [PubMed]

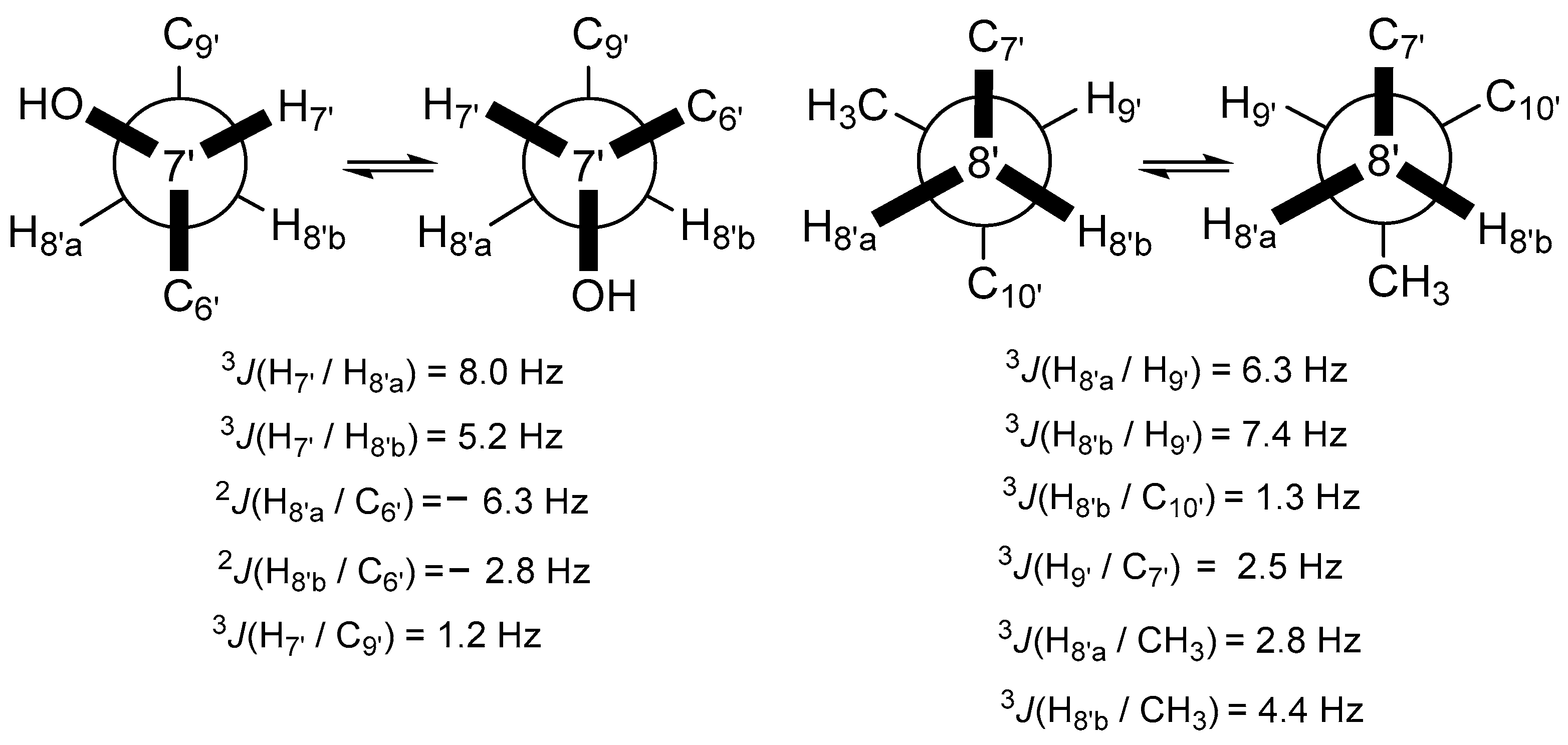

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | δC, Mult | δH, (J Hz) | COSY | HMBC |
|---|---|---|---|---|
| 1 2 3 4 5 6 1′ 2′ 3′ 4′ 5′ 6′ 7′ 8′ 9′ 10′ | 174.5, C 57.6, CH 53.6, CH 23.2, CH2 38.4, CH2 155.5, C 174.7, C 35.6, CH2 122.8, CH 135.1, CH 28.4, CH2 30.5, CH2 30.1, CH2 32.9, CH2 23.7, CH2 14.4, CH3 | 4.39, d(6.4) 3.75, ddd(9.3, 6.4, 4.2) 1.75, m; 1.90, m 3.23, ddd(12.7, 10.3, 4.2) 3.40, dt(12.7, 4.65) 3.08, d(7.1) 5.55, m 5.61, m 2.10, dt(7.1, 6.9) 1.36, m 1.26~1.34 1.26~1.34 1.26~1.34 0.89, t(7.1) | H-3 H-2, H-4 H-3, H-5 H-4 H-3′ H-2′, H-4′ H-3′, H-5′ H-4′, H-6′ H-10′ H-9′ | C-1, C-3, C-4, C-1′ C-1, C-2, C-4, C-5, C-6 C-2, C-3, C-4 C-3, C-4, C-6 C-1′, C-3′, C-4′ C-2′ C-2′, C-5′, C-6′ C-3′, C-4′, C-6′ C-4′, C-7′ C-8′, C-9′ |
| No | δC, Mult | δH, (J Hz) | COSY | HMBC |
|---|---|---|---|---|
| 1 2 3 1′ 2′ 3′ 4′ 5′ 6′ 7′ 8′ 9′ 10′ 11′ 12′ 13′ 14′ 15′ 16′ 17′ 18′ 19′ | 38.8, CH2 26.3, CH2 50.0, CH2 177.6, C 72.8, CH 35.6, CH2 26.2, CH2 30.6, CH2 38.7, CH2 70.3, CH 46.2, CH2 30.6, CH 37.7, CH2 28.0, CH2 26.6, CH2 31.1, CH2 * 30.8, CH2 * 30.5, CH2 * 33.1, CH2 23.8, CH2 14.5, CH3 20.8, CH3 | 3.33, m 1.97, quint(7.8) 2.82, t(7.8) 3.98, dd(7.8, 3.9) a 1.57, m; b 1.74, m 1.43, m 1.33, m a 1.36, dt(7.1, 6.9); b 1.43, m 3.60, m a 1.27, m; b 1.34, m 1.60, m a 1.08, m; b 1.37, m a 1.27, m; b 1.35, m a 1.35, m; b 1.45, m 1.27~1.34, m 1.27~1.34, m 1.27~1.34, m 1.28, m 1.31, m 0.90, t(6.9) 0.89, d(6.9) | H-2 H-1, H-3 H-2 H-3′ H-2′, H-4′ H-3′ H-7′ H-8′, H-6′ H-7′, H-9′ H-8′, H-10′, H-19′ H-9′ H-18′ H-17′ H-9′ | C-1′, C-2, C-3 C-1, C-3 C-1, C-2 C-1′, C-3′, C-4′ C-1′, C-2′ C-4′, C-5′, C-7′ C-6′, C-7′, C-9′, C-19′ C-10′ C-11′ C-16′, C-17′ C-8′, C-9′, C-10′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, B.S.; Lee, S.; Jeong, E.J.; Rho, J.-R. Two New Components from an Association of Marine Sponges Poecillastra sp. and Jaspis sp. and Their Inhibitory Effects on Biomarkers for Benign Prostatic Hyperplasia. Mar. Drugs 2023, 21, 491. https://doi.org/10.3390/md21090491
Hwang BS, Lee S, Jeong EJ, Rho J-R. Two New Components from an Association of Marine Sponges Poecillastra sp. and Jaspis sp. and Their Inhibitory Effects on Biomarkers for Benign Prostatic Hyperplasia. Marine Drugs. 2023; 21(9):491. https://doi.org/10.3390/md21090491
Chicago/Turabian StyleHwang, Buyng Su, Sangbum Lee, Eun Ju Jeong, and Jung-Rae Rho. 2023. "Two New Components from an Association of Marine Sponges Poecillastra sp. and Jaspis sp. and Their Inhibitory Effects on Biomarkers for Benign Prostatic Hyperplasia" Marine Drugs 21, no. 9: 491. https://doi.org/10.3390/md21090491