
Low-Molecular-Weight Fish Collagen Peptide (Valine-Glycine-Proline-Hydroxyproline-Glycine-Proline-Alanine-Glycine) Prevents Osteoarthritis Symptoms in Chondrocytes and Monoiodoacetate-Injected Rats

 , and
, and
Abstract
:1. Introduction
2. Results
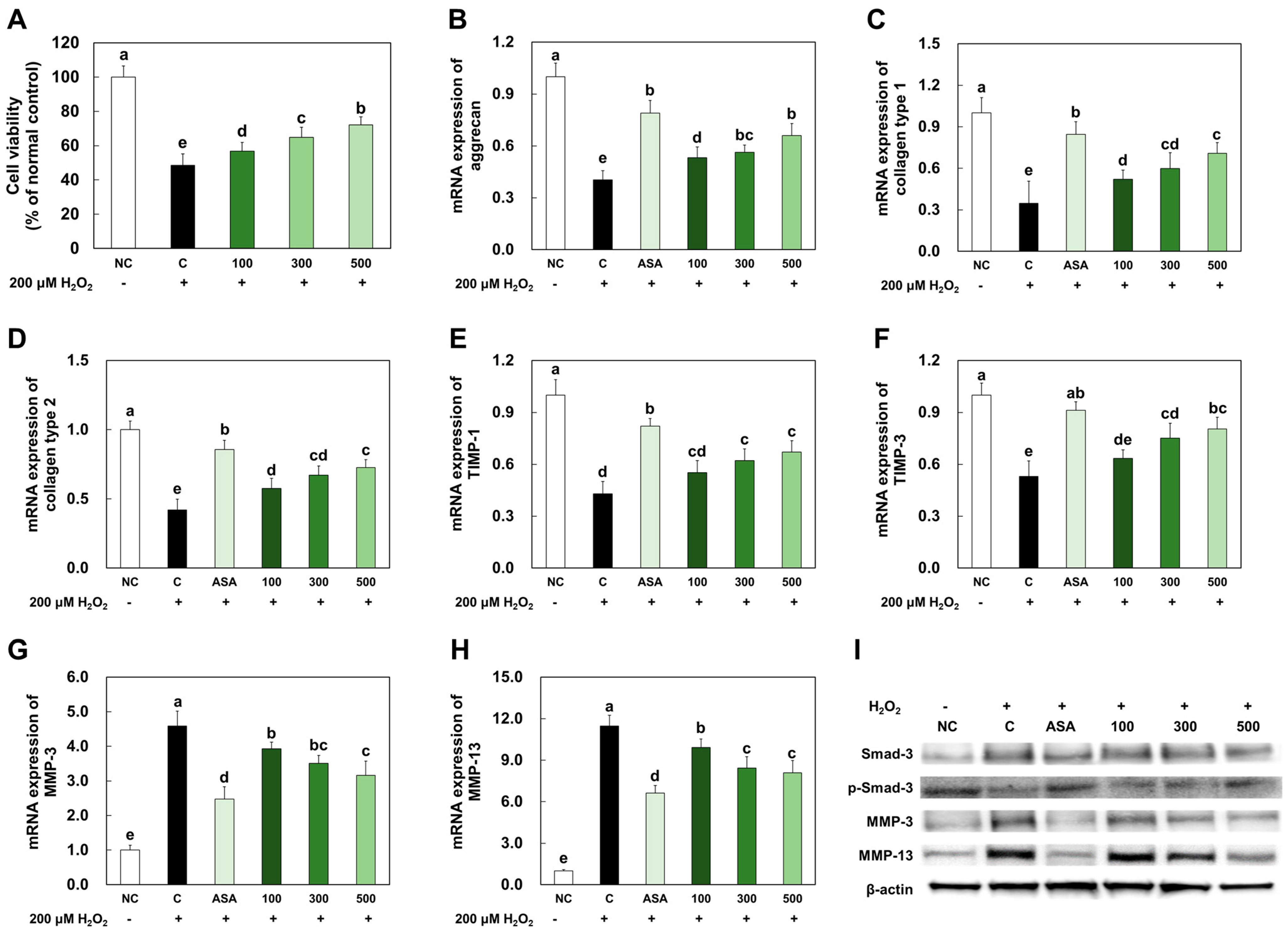
2.1. LMWCP Ameliorated Cell Damage and the Expression of Catabolic Factors in H2O2-Treated Chondrocytes
2.2. LMWCP Ameliorated Inflammation in LPS-Treated Chondrocytes
2.3. LMWCP Ameliorated Apoptosis in LPS-Treated Chondrocytes
2.4. LMWCP Ameliorated Morphological Change of Cartilage Tissue in MIA-Injected Rats
2.5. LMWCP Ameliorated Catabolic Factors Expression in Cartilage Tissue from MIA-Injected Rats
2.6. LMWCP Ameliorated Inflammation in Cartilage Tissue from MIA-Injected Rats
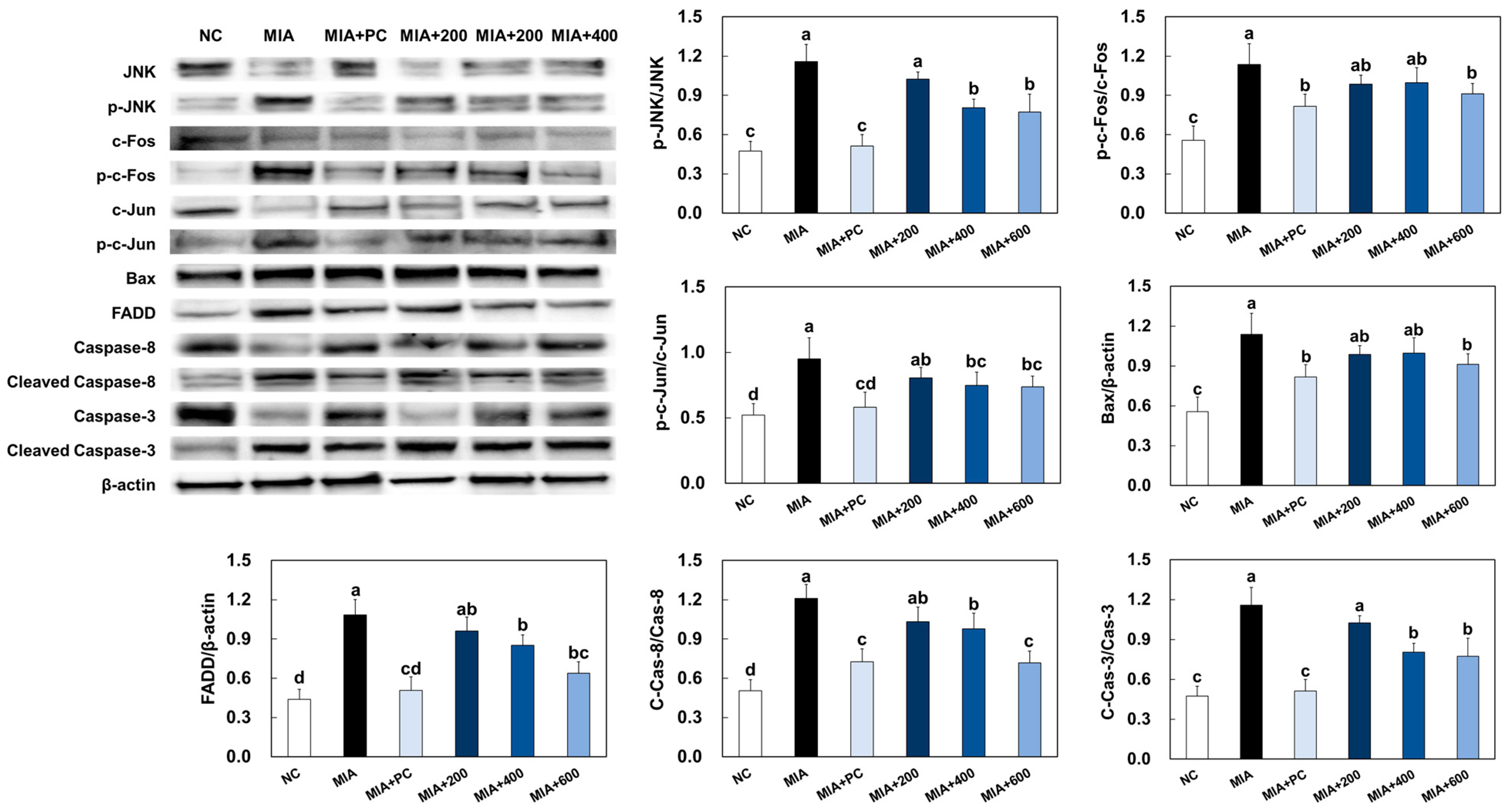
2.7. LMWCP Ameliorated Apoptosis in Cartilage Tissue from MIA-Injected Rats
3. Discussion
4. Materials and Methods
4.1. Preparation of LMWCP
4.2. Primary Culture of Chondrocytes
4.3. Animals and Induction of Osteoarthritis
4.4. Micro-CT and ELISA Analysis
4.5. Total RNA Extraction and Real-Time PCR
4.6. Western Blot
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Arendt-Nielsen, L.; Eskehave, T.N.; Egsgaard, L.L.; Petersen, K.K.; Graven-Nielsen, T.; Hoeck, H.C.; Simonsen, O.; Siebuhr, A.S.; Karsdal, M.; Bay-Jensen, A.C. Association between experimental pain biomarkers and serologic markers in patients with different degrees of painful knee osteoarthritis. Arthritis Rheumatol. 2014, 66, 3317–3326. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, M.; Fragassi, A.; Pannuzzo, M.; Ferreira, M.; Brahmachari, S.; Decuzzi, P. Management of osteoarthritis: From drug molecules to nano/micromedicines. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnology 2022, 14, 1780. [Google Scholar] [CrossRef]
- Martin, J.A.; Buckwalter, J.A. Roles of articular cartilage aging and chondrocyte senescence in the pathogenesis of osteoarthritis. Iowa Orthop. J. 2001, 21, 1–7. [Google Scholar] [PubMed]
- Loeser, R.F.; Collins, J.A.; Diekman, B.O. Ageing and the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Loeser, R.F. Aging and osteoarthritis: The role of chondrocyte senescence and aging changes in the cartilage matrix. Osteoarthr. Cartil. 2009, 17, 971–979. [Google Scholar] [CrossRef]
- Giannoni, P.; Cancedda, R. Articular chondrocyte culturing for cell-based cartilage repair: Needs and perspectives. Cells Tissues Organs 2006, 184, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Camarero-Espinosa, S.; Rothen-Rutishauser, B.; Foster, E.J.; Weder, C. Articular cartilage: From formation to tissue engineering. Biomater. Sci. 2016, 4, 734–767. [Google Scholar] [CrossRef]
- Sandell, L.J.; Aigner, T. Articular cartilage and changes in arthritis. An introduction: Cell biology of osteoarthritis. Arthritis Res. 2001, 3, 107–113. [Google Scholar] [CrossRef]
- Aida, Y.; Maeno, M.; Suzuki, N.; Shiratsuchi, H.; Motohashi, M.; Matsumura, H. The effect of IL-1beta on the expression of matrix metalloproteinases and tissue inhibitors of matrix metalloproteinases in human chondrocytes. Life Sci. 2005, 77, 3210–3221. [Google Scholar] [CrossRef]
- Bondeson, J.; Wainwright, S.D.; Lauder, S.; Amos, N.; Hughes, C.E. The role of synovial macrophages and macrophage-produced cytokines in driving aggrecanases, matrix metalloproteinases, and other destructive and inflammatory responses in osteoarthritis. Arthritis Res. Ther. 2006, 8, 187. [Google Scholar] [CrossRef]
- Rovati, L.C.; Girolami, F.; D’Amato, M.; Giacovelli, G. Effects of glucosamine sulfate on the use of rescue non-steroidal anti-inflammatory drugs in knee osteoarthritis: Results from the Pharmaco-Epidemiology of GonArthroSis (PEGASus) study. Semin. Arthritis Rheum. 2016, 45, 34–41. [Google Scholar] [CrossRef]
- Bannuru, R.R.; Vaysbrot, E.E.; Sullivan, M.C.; McAlindon, T.E. Relative efficacy of hyaluronic acid in comparison with NSAIDs for knee osteoarthritis: A systematic review and meta-analysis. Semin. Arthritis Rheum. 2014, 43, 593–599. [Google Scholar] [CrossRef]
- Guan, V.X.; Mobasheri, A.; Probst, Y.C. A systematic review of osteoarthritis prevention and management with dietary phytochemicals from foods. Maturitas 2019, 122, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.; Gagnier, J.J.; Little, C.V.; Parsons, T.J.; Blümle, A.; Chrubasik, S. Evidence of effectiveness of herbal medicinal products in the treatment of arthritis. Osteoarthritis. Phytother. Res. 2009, 23, 1497–1515. [Google Scholar] [CrossRef] [PubMed]
- Bello, A.E.; Oesser, S. Collagen hydrolysate for the treatment of osteoarthritis and other joint disorders: A review of the literature. Curr. Med. Res. Opin. 2006, 22, 2221–2232. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Puig, D.; Costa-Larrión, E.; Rubio-Rodríguez, N.; Gálvez-Martín, P. Collagen Supplementation for Joint Health: The Link between Composition and Scientific Knowledge. Nutrients 2023, 15, 1332. [Google Scholar] [CrossRef] [PubMed]
- Cúneo, F.; Costa-Paiva, L.; Pinto-Neto, A.M.; Morais, S.S.; Amaya-Farfan, J. Effect of dietary supplementation with collagen hydrolysates on bone metabolism of postmenopausal women with low mineral density. Maturitas 2010, 65, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K.; Hasegawa, T.; Taguchi, Y.; Morimatsu, F.; Sato, K.; Nakamura, Y.; Higashi, A.; Kido, Y.; Nakabo, Y.; Ohtsuki, K. Identification of food-derived collagen peptides in human blood after oral ingestion of gelatin hydrolysates. J. Agric. Food Chem. 2005, 53, 6531–6536. [Google Scholar] [CrossRef]
- Yazaki, M.; Ito, Y.; Yamada, M.; Goulas, S.; Teramoto, S.; Nakaya, M.A.; Ohno, S.; Yamaguchi, K. Oral Ingestion of Collagen Hydrolysate Leads to the Transportation of Highly Concentrated Gly-Pro-Hyp and Its Hydrolyzed Form of Pro-Hyp into the Bloodstream and Skin. J. Agric. Food Chem. 2017, 65, 2315–2322. [Google Scholar] [CrossRef]
- Maldonado, M.; Nam, J. The role of changes in extracellular matrix of cartilage in the presence of inflammation on the pathology of osteoarthritis. Biomed. Res. Int. 2013, 2013, 284873. [Google Scholar] [CrossRef]
- Murphy, G.; Nagase, H. Progress in matrix metalloproteinase research. Mol. Aspects Med. 2008, 29, 290–308. [Google Scholar] [CrossRef] [PubMed]
- Klatt, A.R.; Paul-Klausch, B.; Klinger, G.; Kühn, G.; Renno, J.H.; Banerjee, M.; Malchau, G.; Wielckens, K. A critical role for collagen II in cartilage matrix degradation: Collagen II induces pro-inflammatory cytokines and MMPs in primary human chondrocytes. J. Orthop. Res. 2009, 27, 65–70. [Google Scholar] [CrossRef]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [PubMed]
- Choi, M.C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-κB Signaling Pathways in Osteoarthritic Cartilage Destruction. Cells 2019, 8, 734. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.S.; Kim, H.A. Chondrocyte Apoptosis in the Pathogenesis of Osteoarthritis. Int. J. Mol. Sci. 2015, 16, 26035–26054. [Google Scholar] [CrossRef]
- Takács-Buia, L.; Iordachel, C.; Efimov, N.; Caloianu, M.; Montreuil, J.; Bratosin, D. Pathogenesis of osteoarthritis: Chondrocyte replicative senescence or apoptosis? Cytometry B Clin. Cytom. 2008, 74, 356–362. [Google Scholar] [CrossRef]
- Zimmermann, K.C.; Green, D.R. How cells die: Apoptosis pathways. J. Allergy Clin. Immunol. 2001, 108, 99–103. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Diet | Supplement | MIA |
|---|---|---|---|
| Normal control | AIN93G | − | − |
| MIA | AIN93G | − | + |
| MIA + PC | AIN93G | Ibuprofen 20 mg/kg BW | + |
| MIA + 200 | AIN93G | LMWCP 200 mg/kg BW | + |
| MIA + 400 | AIN93G | LMWCP 400 mg/kg BW | + |
| MIA + 600 | AIN93G | LMWCP 600 mg/kg BW | + |
| Gene | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|
| GAPDH (NM_017008) | TGG CCT CCA AGG AGT AAG AAA C | CAG CAA CTG AGG GCC TCT CT |
| Aggrecan (NM_022190) | GAA GTG GCG TCC AAA CCA A | CGT TCC ATT CAC CCC TCT CA |
| Collagen Type I (NM_000088) | GAG CGG AGA GTA CTG GAT CGA | CTG ACC TGT CTC CAT GTT GCA |
| Collagen Type II (RATCOLLII) | GCA ACA GCA GGT TCA CGT ACA | TCG GTA CTC GAT GAT GGT CTT G |
| TIMP-1 (NM_053819) | ACA GCT TTC TGC AAC TCG GA | CGT CGA ATC CTT TGA GCA TC |
| TIMP-3 (RNU27201) | CTT CTG CAA CTC CGA CAT CGT | GGG GCA TCT TAC TGA ATC CTC |
| MMP-3 (NM_133523) | GAG TGT GGA TTC TGC CAT TGA G | TTA TGT CAG CCT CTC CTT CAG AGA |
| MMP-13 (NM_133530.1) | ACG TTC AAG GAA TCC AGT CT | GGA TAG GGC TGG GTC ACA CTT |
| Antibodies | Distributor |
|---|---|
| Smad3 | Cell signaling (#9523, Beverly, MA, USA) |
| p-Smad3 | Cell signaling (#9520) |
| MMP-3 | Abcam (ab53015) |
| MMP-13 | Abcam (ab39012) |
| IκBα | Cell signaling (#9242) |
| p-IκBα | Cell signaling (#2859) |
| p65 | Abcam (ab16502) |
| p-p65 | Cell signaling (#3031) |
| COX-2 | Cell signaling (#12282) |
| JNK | Cell signaling (#9252) |
| p-JNK | Cell signaling (#9251) |
| c-Fos | Cell signaling (#4384 |
| p-c-Fos | Cell signaling (#5348) |
| c-Jun | Cell signaling (#9165) |
| p-c-Jun | Cell signaling (#2361) |
| Bax | Cell signaling (#2772) |
| FADD | LSbio (LS-C766496, Settle, WA, USA) |
| Caspase-8 | Cell signaling (#4790) |
| Cleaved caspase-8 | Cell signaling (#8592) |
| Caspase-3 | Cell signaling (#9662) |
| Cleaved caspase-3 | Cell signaling (#9661) |
| β-actin | BETHYL (A300-491A, Waltham, MA, USA) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, W.; Park, J.; Kim, J.; Lee, M.; Park, S.J.; Kim, K.S.; Jun, W.; Kim, O.-K.; Lee, J. Low-Molecular-Weight Fish Collagen Peptide (Valine-Glycine-Proline-Hydroxyproline-Glycine-Proline-Alanine-Glycine) Prevents Osteoarthritis Symptoms in Chondrocytes and Monoiodoacetate-Injected Rats. Mar. Drugs 2023, 21, 608. https://doi.org/10.3390/md21120608
Cho W, Park J, Kim J, Lee M, Park SJ, Kim KS, Jun W, Kim O-K, Lee J. Low-Molecular-Weight Fish Collagen Peptide (Valine-Glycine-Proline-Hydroxyproline-Glycine-Proline-Alanine-Glycine) Prevents Osteoarthritis Symptoms in Chondrocytes and Monoiodoacetate-Injected Rats. Marine Drugs. 2023; 21(12):608. https://doi.org/10.3390/md21120608
Chicago/Turabian StyleCho, Wonhee, Jeongjin Park, Jinhee Kim, Minhee Lee, So Jung Park, Kyung Seok Kim, Woojin Jun, Ok-Kyung Kim, and Jeongmin Lee. 2023. "Low-Molecular-Weight Fish Collagen Peptide (Valine-Glycine-Proline-Hydroxyproline-Glycine-Proline-Alanine-Glycine) Prevents Osteoarthritis Symptoms in Chondrocytes and Monoiodoacetate-Injected Rats" Marine Drugs 21, no. 12: 608. https://doi.org/10.3390/md21120608