1. Introduction
Acute kidney injury (AKI), a syndrome engendered by sepsis, cardiorenal syndrome, urinary tract obstruction, and nephrotoxins, is known to increase the level of serum creatinine and/or decrease urine output. It is an important public health issue because of being a serious complication for 10–15% of hospitalized patients and ~50% of those in intensive care [
1].
Animal models of AKI are induced by administering a drug or toxicant (e.g., HgCl
2) [
2,
3]. Mercury targets the kidney by binding to thiol-containing proteins in the tubular and glomerular nephron portion, disrupting the tubular transport mechanism related to Na
+/K
+-ATPase [
4]. It also alters the intracellular calcium current and consequently the redox environment. The increase in oxidants is not counteracted by the antioxidant system and therefore leads to oxidative stress [
5,
6] and endoplasmic reticulum (ER) stress [
3].
ER stress disrupts proteostasis in this organelle, causing the accumulation of unfolded and misfolded proteins. To maintain ER function, the unfolded protein response is activated through the protein kinase RNA-like ER kinase (PERK), activating transcription factor 6α (ATF6α), and inositol-requiring enzyme 1α (IRE1α) pathways.
The PERK pathway, crucial in regulating the unfolded protein response, reduces transcription through phosphorylation of the eukaryotic translation initiation factor-2α (eIF2α). If ER stress is controlled, protein folding can resume, and the phosphorylated eIF2α dephosphorylates. In the event that ER stress is sustained, the activating transcription factor 4 (ATF4)/growth arrest and DNA damage-inducible gene 153 (GADD153, also called CHOP) pathway activates the expression of genes that participate in redox homeostasis, autophagy, and/or apoptosis. The particular genes involved depend on the level of ER stress [
7,
8].
In parallel, the ATF6α pathway diminishes ER stress by regulating genes that encode ER chaperones and enzymes responsible for promoting folding, maturation, secretion, or degradation of misfolded proteins. When ER stress is sustained, the cell activates autophagy and apoptosis by upregulating the generation of reactive oxygen species (ROS) and activating ER membrane-associated caspase 12 through the ATF4/GADD153 pathway [
7,
8].
Additionally, IRE1α contributes to adaptation or apoptosis under chronic ER stress. The adaptation response of IRE1α is activated by selective cleavage of X-box binding protein 1 (XBP1) mRNA to produce spliced isoforms of XBP1, which enhance the transcription of chaperones, foldases, and components of the ER-associated protein degradation (ERAD) response to restore proteostasis. In case ER stress is still uncontrolled, IRE1α activates c-Jun N-terminal kinases 1 (JNK1) to promote the translocation of B-cell lymphoma 2 (Bcl-2)-associated X protein (Bax) into the mitochondrial membrane, which triggers the release of cytochrome c and the second mitochondrial-activated factor (Smac), leading to the activation of caspases 3 and 9 [
9].
Some research groups have been developing eco-friendly therapeutic strategies for AKI from microalgae pigments such as phycobiliproteins, toroidal light-harvesting proteins in cyanobacteria, and the photosynthetic apparatus in algae. The most studied phycobiliprotein with nephroprotective activity is C-phycocyanin (C-PC). It impedes kidney failure by decreasing oxidative stress and ER stress in mice intoxicated with mercury [
6,
10,
11]. Moreover, other phycobiliproteins, including C-phycoerythrin (C-PE), have nutraceutical activity against metabolic and toxic injury that affects certain organs (e.g., the liver) in animal models [
12,
13].
C-PE, an oligomeric chromoprotein of cyanobacteria, is composed of monomers αβ and prosthetic covalently linked open-chain tetrapyrrole moieties denominated C-phycoerythrobilin. In
Phormidium sp., the monomer units oligomerize to form trimers (αβ)
3 and then stack as hexamers [(αβ)
3]
2 [
14]. C-PE is widely used in the food and cosmetic industries, as well as in diagnosis and research. There are reports on its nutraceutical properties, which stem from scavenging and antioxidant activity [
15]. Our research group demonstrated that a C-PE-rich protein extract from
Pseudanabaena tenuis has nephroprotective activity [
16], although its mechanism is still not completely understood. The aim of the current contribution was to determine whether the nephroprotective activity of C-PE (purified from
Phormidium persicinum) is related to a reduction in oxidative stress and ER stress, and consequently an attenuation of the alterations in the levels of nephrin and podocin normally caused by HgCl
2-induced AKI.
3. Discussion
C-PE is reported to have nutraceutical activity against the damage resulting from cell insult [
12,
13]. Our group has demonstrated that treatment with a protein extract rich in C-PE prevented oxidative stress and cellular damage in an animal model of HgCl
2-induced AKI [
16]. This model was chosen because mercury produces ER stress, which leads to renal damage. However, the aforementioned study only associated the nutraceutical properties of C-PE with scavenging and antioxidant activity. Thus, the aim of the current contribution was to explore the molecular mechanism of action of C-PE (purified from
P. persicinum) by examining its nephroprotective activity against HgCl
2-induced ER stress, oxidative stress, and alterations in the redox environment in the same animal model.
HgCl
2 produces oxidative stress and alterations in the redox environment by three mechanisms: Fenton and Haber-Weiss reactions that generate free radicals and ROS [
17], the activation of ER stress [
3], and the binding of Hg
2+ with intracellular sulfhydryl-containing proteins and low-molecular-weight compounds (e.g., GSH) capable of affecting the redox environment and protein function [
18]. As a consequence of these reactions, nephrin and podocin are downregulated, and the slit diaphragm is injured, which is observed as HgCl
2-induced AKI. The resulting inflammatory process participates in the progression of AKI [
19].
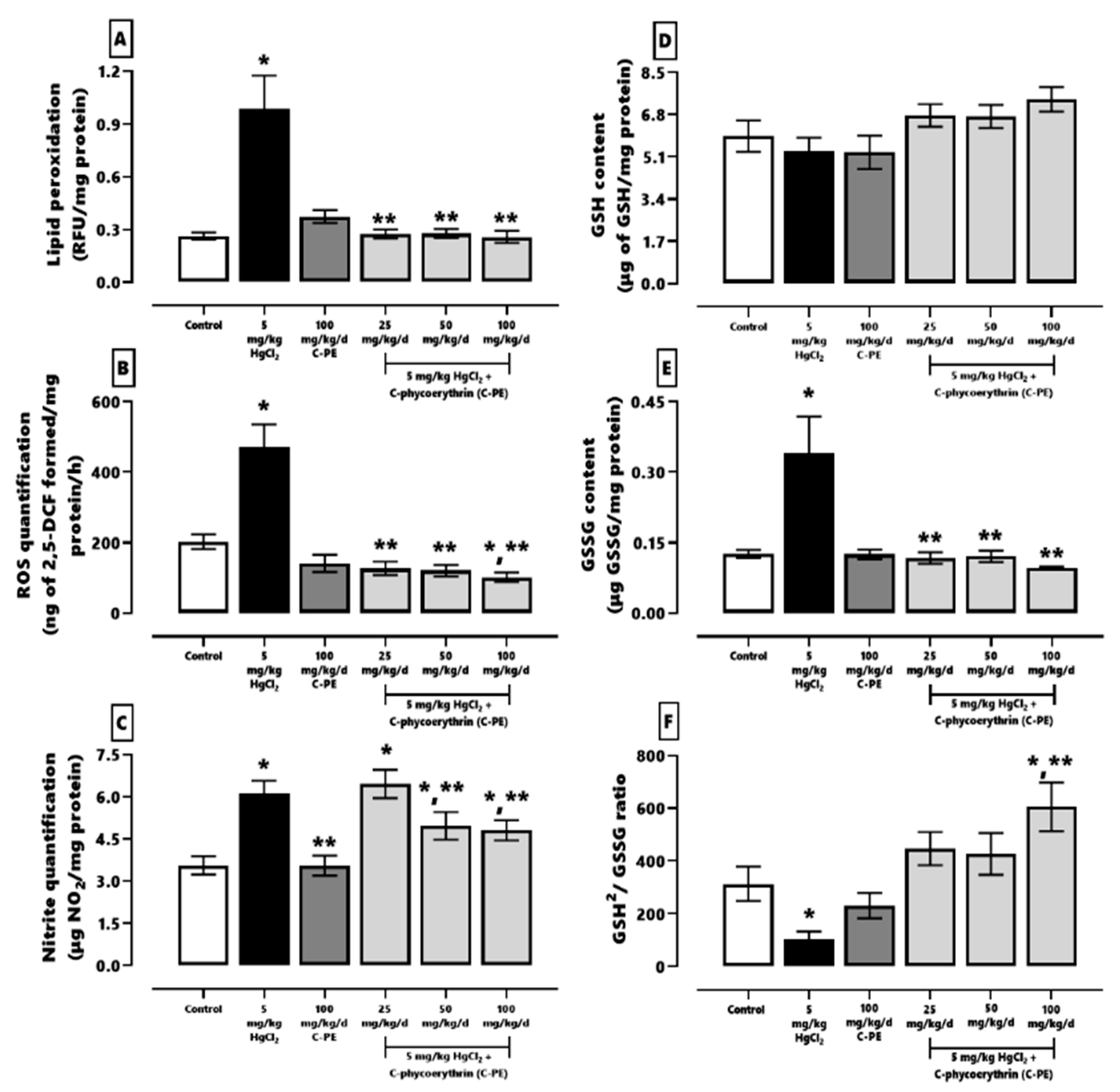
In recent years, the use of nutraceuticals from cyanobacteria and their metabolites has proven effective against renal damage (e.g., AKI) stemming from toxicants or chronic kidney disease [
16,
20,
21,
22]. Purified C-PE presently demonstrated nephroprotective activity when tested against HgCl
2-induced AKI, as evidenced by the reduction found in oxidative stress and ER stress.
C-PE, a protein with a molecular weight of ~240 KDa, has nutraceutical properties in vitro as an ROS scavenger [
23]. Moreover, it prevents oxidative stress and cellular damage in vivo [
12,
13]. All reports on C-PE suggest that it is a potent antioxidant. By scavenging ROS, it avoids alterations in the redox environment and therefore impedes cellular damage [
12,
13,
24]. However, animal studies have not yet completely defined the nutraceutical protection mechanism.
C-PE may act as a prodrug that leads to the release of the phycoerythrobilin moiety into the gastrointestinal tract, as previously demonstrated by our group for C-PC and phycocyanobilin [
22]. C-PC is known to break down into chromo-peptides that contain phycocyanobilin, followed by the apparent absorption of linear tetrapyrrole compounds facilitated by the action of intestinal peptidases [
24,
25]. Once in serum, phycoerythrobilin could bind to albumin due to its low water solubility, which would extend its therapeutic activity into the entire organism [
26].
The protective effect of C-PE against HgCl
2-induced AKI is associated with antioxidant, anti-inflammatory, and chelation mechanisms. C-PE acts as an antioxidant because it contains PEB. In addition, the chemical structure of phycoerythrobilin acts as a nucleophilic compound, neutralizing free radicals and ROS [
24]. According to an in vitro model, the chelation of Hg
2+ by PEB suppresses the degranulation of RBL-2H3 mast cells and decreases the intracellular concentration of Ca
2+ [
27], giving rise to anti-inflammatory and nephroprotective effects. Hg
2+ binds to PEB thioether bridges in C-PE, which assume a cyclic helical form capable of chelation [
28]. The antioxidant and chelating activity of C-PE can avoid Fenton and Haber-Weiss reactions and consequently ameliorate the production of free radicals, the generation of oxidative stress, and the alteration of the redox environment in kidney cells. All the aforementioned mechanisms of C-PE are related to the maintenance of the redox environment and therefore prevent the dysfunction of organelles such as the ER.
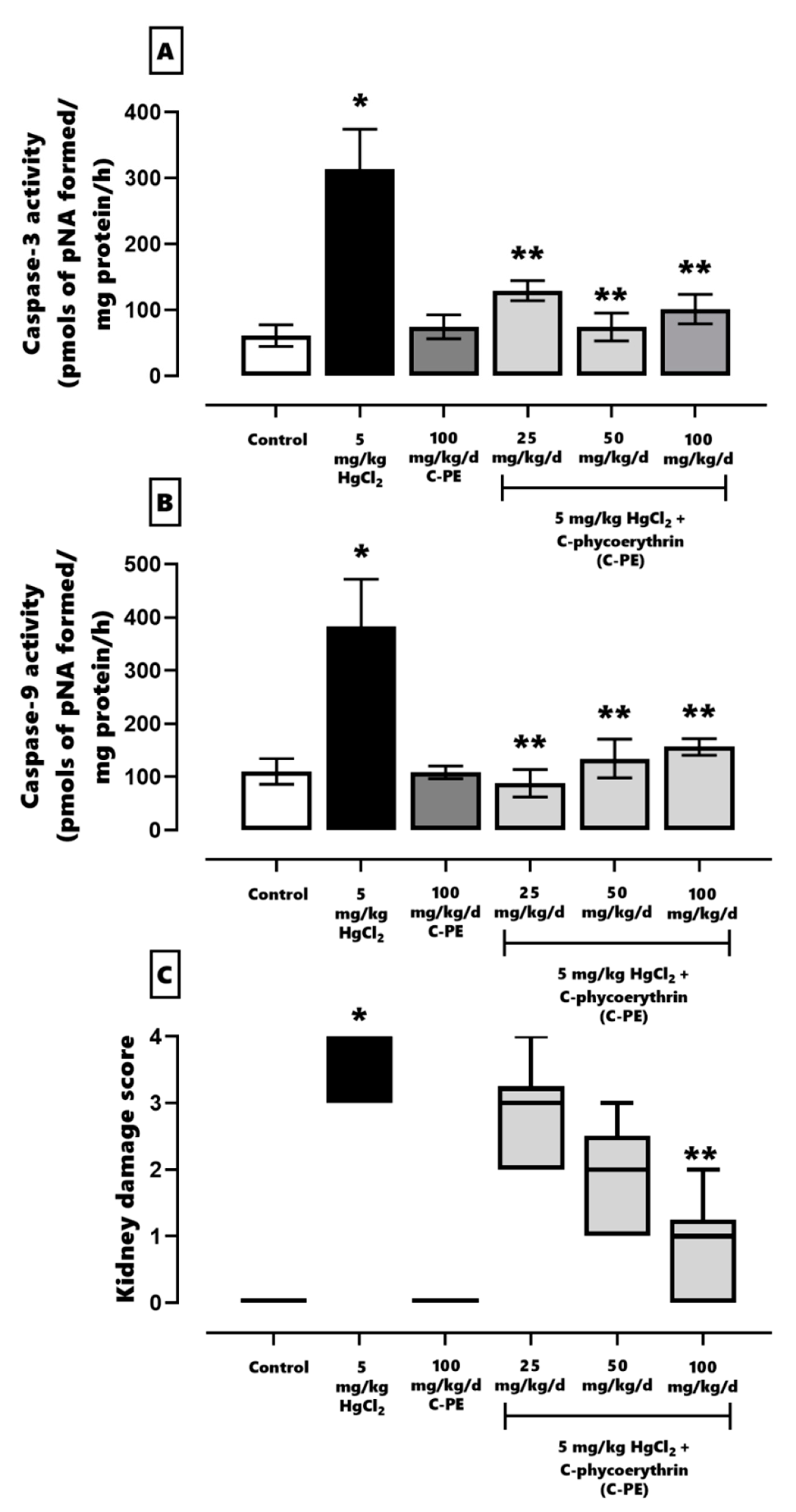
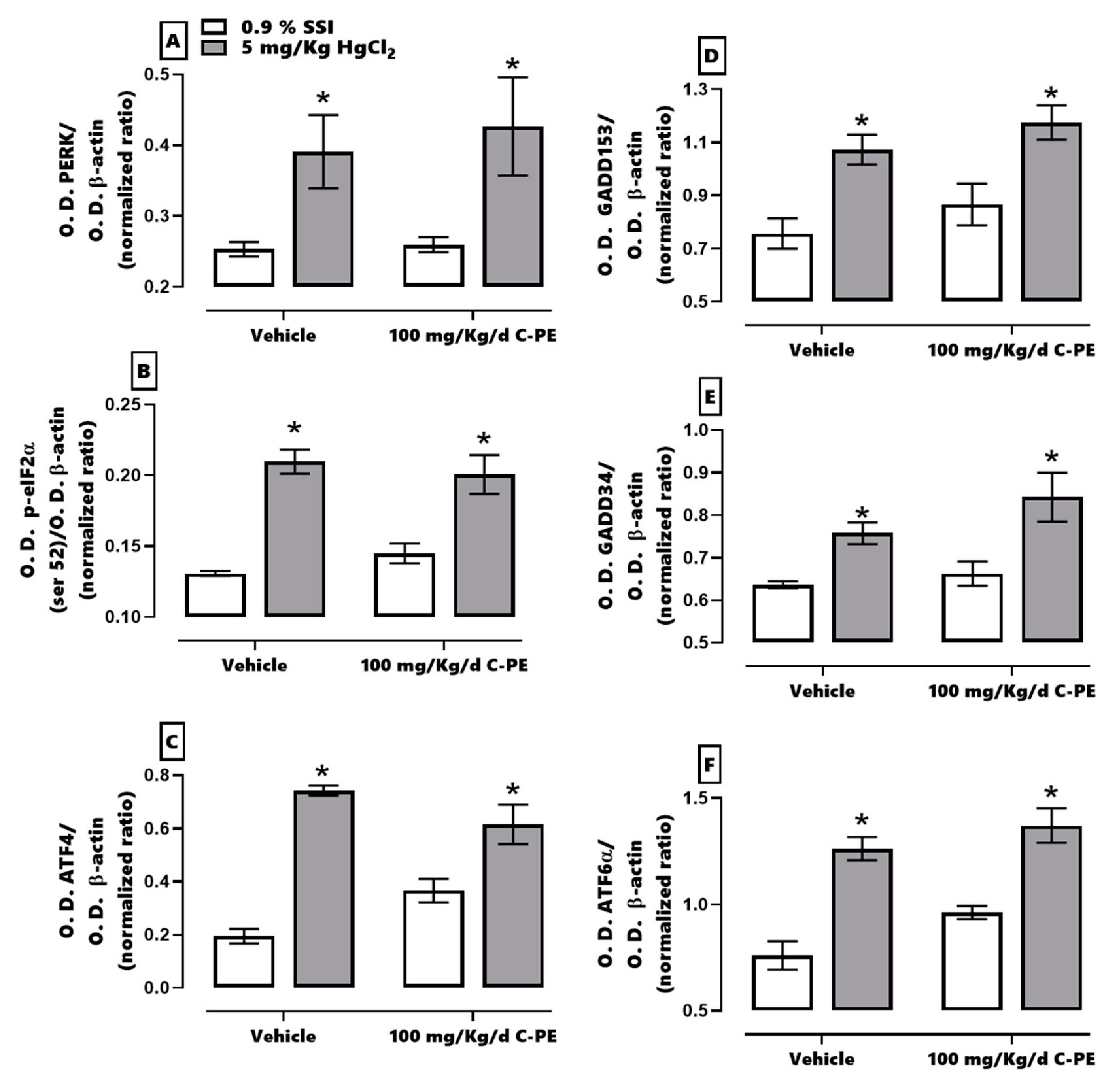
In the current evaluation of proteostasis, HgCl
2-induced ER stress was found to activate the IRE1α pathway and promote cell death. At the same time, mercury activated the PERK pathway, which restored proteostasis through PERK/eIF2α/ATF-4/GADD153. When the cell was incapable of compensating for imbalances in proteostasis, the activation of ATF4 and GADD153 in the same pathway led to the expression of proapoptotic proteins and the triggering of cell death. As can be appreciated, PERK and IRE1α have a synergic effect in prompting kidney cell death by increasing the Bax/Bcl-2 ratio and the level of caspases 3, 8, 9, and 12 [
3,
10]. Hence, HgCl
2 was capable of generating AKI in the present study by fomenting oxidative stress, an alteration in the redox environment, and ER stress. The resulting histological damage was considerable (grade 4), affecting over 75% of tubular and glomerular cells.
C-PE treatment enhanced the canonical ER response through the PERK/p-eIF2α (ser 52)/ATF-4/GADD153 pathway, involving ER-associated degradation (ERAD), known to process misfolded and unfolded proteins. The phosphorylation of eIF2α (ser 52) is able to suppress the overall translation of mRNA, thus reducing protein stress in the ER. Furthermore, the moderate increment in ATF6α upregulates several genes that participate in the adaptative phase of the unfolded protein response [
29]. C-PE treatment is herein proposed to have activated the PERK and ATF6 signaling pathways, maintaining proteostasis by avoiding oxidative stress and alterations in the redox environment and by activating the unfolded protein response [
30,
31].
The response elicited by C-PE is distinct from that of other phycobiliproteins. For instance, C-PC averts the overexpression of GADD34 by activating GADD153, which is related to the inhibition of apoptosis [
11,
32]. On the other hand, both C-PC and C-PE maintain proteostasis. The differences between these two responses should be explored in depth in future research.
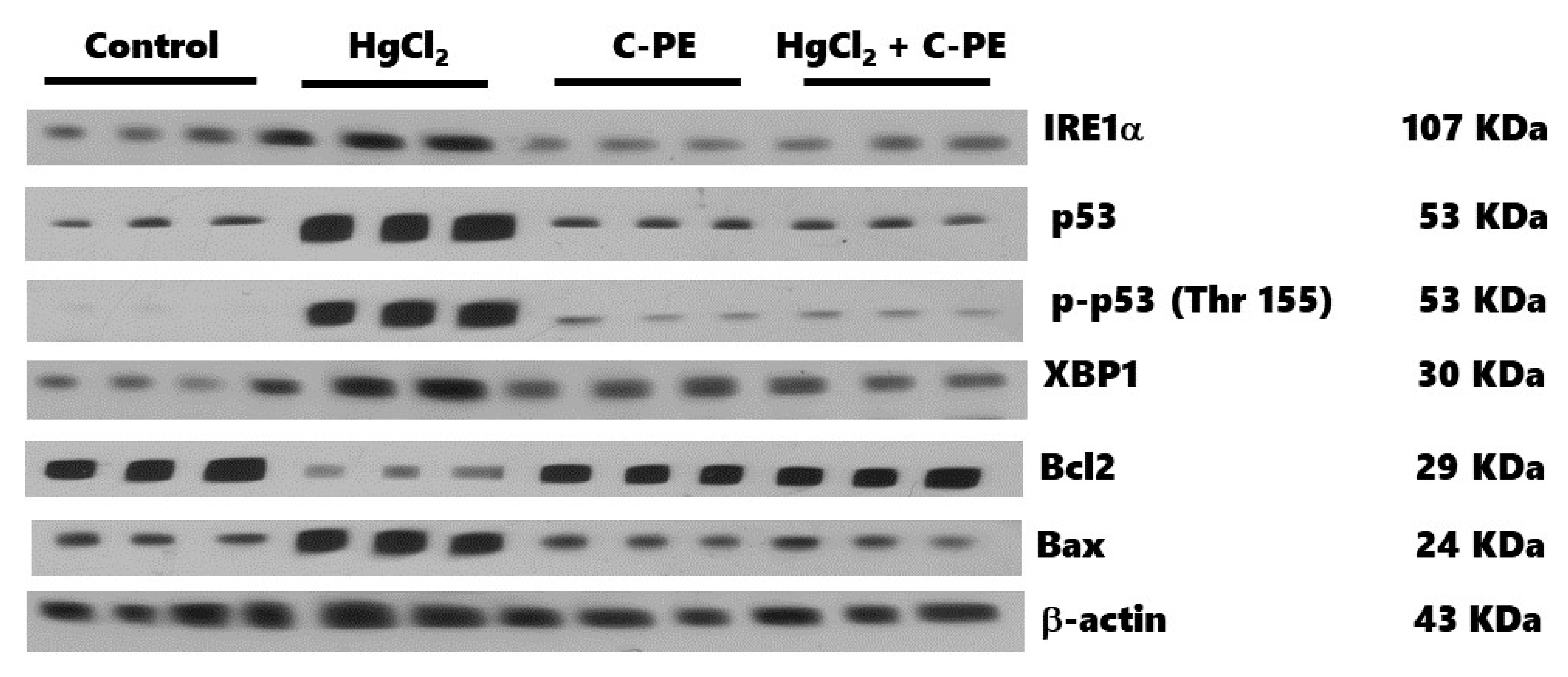
C-PE and C-PC have a similar effect on the IREα pathway, decreasing cell death mediated by caspases 3, 9, and 12 as well as reducing the disruption in p53 activation and the alteration of the Bax/Bcl2 ratio [
10,
11]. This idea is supported by neurotoxicological models, where C-PE prevents ER stress linked to calcium deregulation and mitochondrial dysfunction [
33].
In the control group, interestingly, C-PE per se increased the phosphorylation of p53 (Thr 155), which is a genome gatekeeper because it is a master transcriptional factor that induces cellular senescence and suppresses cell growth and tumor formation. Exposure to various cellular stressors, however, causes p53 to be overexpressed and phosphorylated in several regions, leading to cell cycle arrest or apoptosis. Accordingly, p53 is phosphorylated by the C-Jun activation domain-binding protein-1 (Jab1) in Thr 155, promoting its translocation into the cytoplasm to favor interaction with the COP9 signalosome complex. These nuclear export mechanisms of p53 provide a practical future approach to a possible C-PE-induced activation of anti-cancer therapy by p53 [
34], as evidenced by the lack of histological irregularities in the C-PE control group as well as the capacity of C-PE treatment of AKI mice to prevent oxidative stress, ER stress, and alterations in the redox environment and cell death markers.
4. Materials and Methods
4.1. Animals
Forty-eight male albino NIH Swiss mice (25–30 g) were kept in a cool room (21 ± 2 °C) with 40–60% relative humidity under a 12/12 h light/dark cycle (lights on at 8 AM). Food and water were provided ad libitum. The experimental procedures were in accordance with the Official Mexican Norm (NOM-062-ZOO-1999, technical specifications for the production, care, and use of laboratory animals) [
35]. The protocol was approved by the institutional Internal Bioethics Committee (ZOO-013-2021).
The animals were divided into two lots to carry out distinct protocols, one to assess oxidative stress and kidney damage and another to analyze ER stress. For the evaluation of oxidative stress and kidney damage, 36 mice were randomly allocated to 6 groups (n = 6). Three were control groups: (1) the vehicle (negative control), with 100 mM of phosphate buffer (PB, at pH 7.4) administered by oral gavage (og) + 0.9% of saline solution (SS) applied intraperitoneally (ip), (2) AKI induced by a single application of 5 mg/kg HgCl2 ip + the vehicle (PB) og, and (3) C-PE treatment, consisting of 100 mg/kg/day C-PE og + 0.9% SS ip. The other three groups received a single application of HgCl2 ip as well as 25, 50, or 100 mg/kg/day C-PE og. For the analysis of ER stress, twelve mice were randomly allocated to four groups with the following treatments (n = 3): (1) the control (vehicle), (2) mercury-induced AKI, (3) the C-PE treatment, and (4) the AKI + C-PE treatment (a single application of HgCl2 ip and 100 mg/kg/day C-PE og).
C-PE or the vehicle was administered 30 min before the injection of HgCl2 or 0.9% of SS. C-PE was administered once daily for five days (the first protocol) or for three days (the second protocol) at the same time (12:00 AM) each day. Whereas the mice assigned to the evaluation of oxidative stress and renal damage were euthanized 5 days after mercury intoxication, those employed for assessing ER stress were euthanized 3 days after the same event. The right kidneys were frozen at −70 °C to await examination of the markers of oxidative stress and the redox environment by Western blot, while the left kidneys were put into paraformaldehyde in PBS (4% v/v) to appraise cell damage.
4.2. Cultivation, Purification, and Characterization of C-PE from Phormidium persicinum
P. persicinum was obtained from the culture collection of the Centro de Investigaciones Biológicas del Noroeste, S. C. (CIB 84). It was grown in a synthetic medium (denominated NM), created and optimized by our group (composition: 29 g/L of commercial sea salt, 0.8 g/L NaHCO3, 0.05 g/L K2HPO4, 2.16 g/L NaNO3, 5 mg/L MgSO4, 1 mg/L FeSO4, and 1 mL of a micronutrient solution containing 0.2 mM EDTA, 46.2 mM H3BO3, 9.3 mM MnCl2, 0.95 mM ZnSO4, 2.03 mM Na2MoO4, 0.49 mM Ca(NO3)2, and 0.77 mM CuSO4). Incubation was carried out at 21 ± 2 °C with constant aeration provided by an air pump, under green LED illumination (24 W, 3000 Lx) and a 12/12 h light/dark cycle (lights on at 8:00 AM).
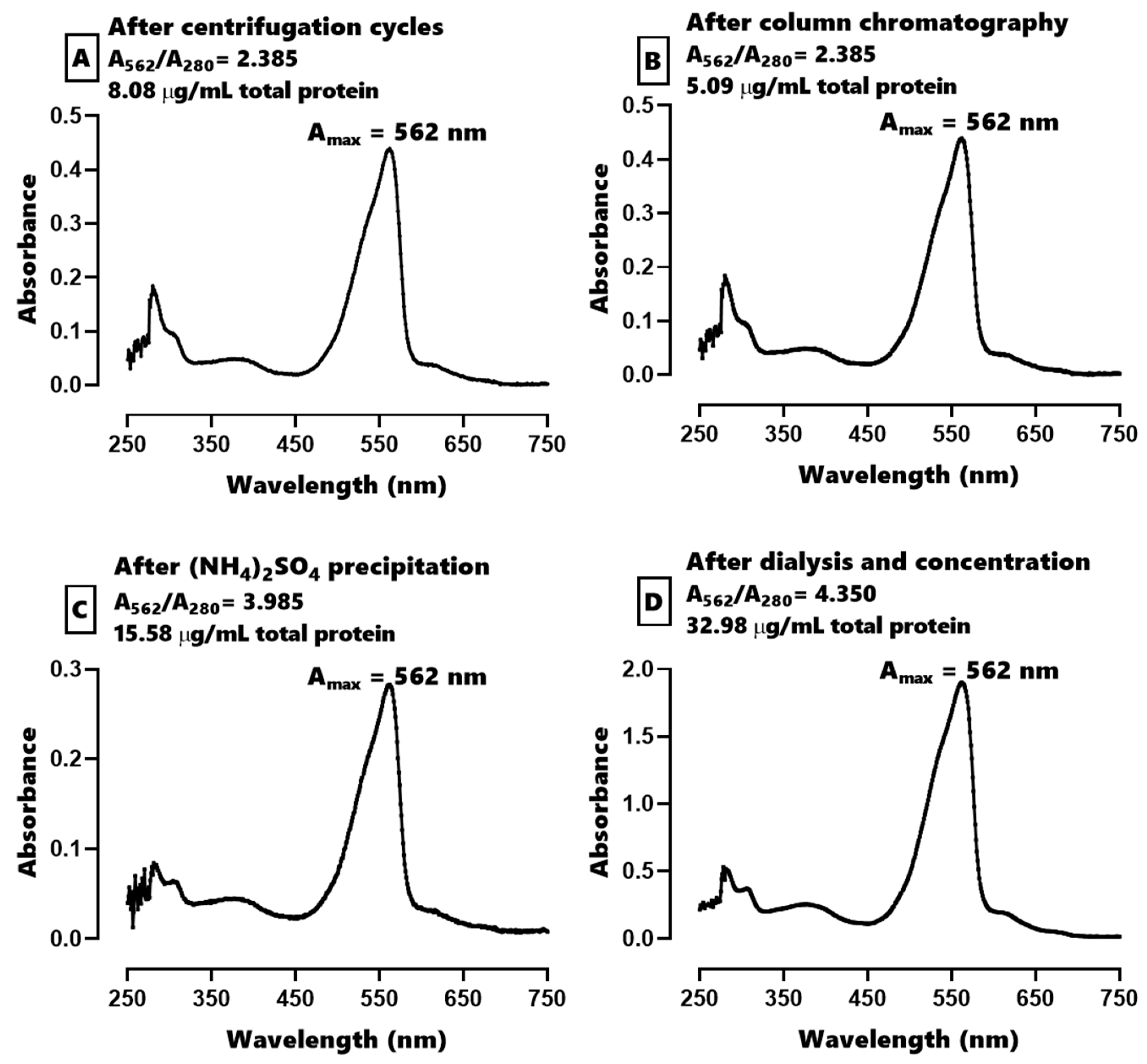
Regarding the purification of C-PE, the cyanobacterial biomass was centrifuged at 10,000×
g for 1 min and 5–10 g of the resulting cell pellet was re-suspended in 20 mL of distilled water. Subsequently, three freeze–thaw cycles were performed, freezing at −20 °C and thawing at 4 °C during 24 h. The resulting slurry was centrifuged in 4 cycles at 21,400×
g for 10 min at 4 °C to remove the cell debris. An aliquot of 20 mL of the phycobiliprotein-rich extract was injected into a column (33 cm long × 4.7 cm in diameter) containing Sephadex G-250 gel previously equilibrated with 10 mM of PB (pH 7.4). The pink fractions were obtained and precipitated with a saturated solution of (NH
4)
2SO
4 at 4 °C for 24 h in the dark. This mixture was centrifuged at 21,400×
g for 2 min at 4 °C, and the resulting pellet was resuspended in 100 mM of PB at pH 7.4. The membrane was then dialyzed with PB for 24 h, after which time an aliquot of C-PE was immediately lyophilized to construct a calibration curve, obtain an absorption spectrum, and characterize the extract fluorometrically with an EEM. The C-PE extract was solubilized in PB and 5 mM of sucrose and frozen at −20 °C to await administration to the animals [
36].
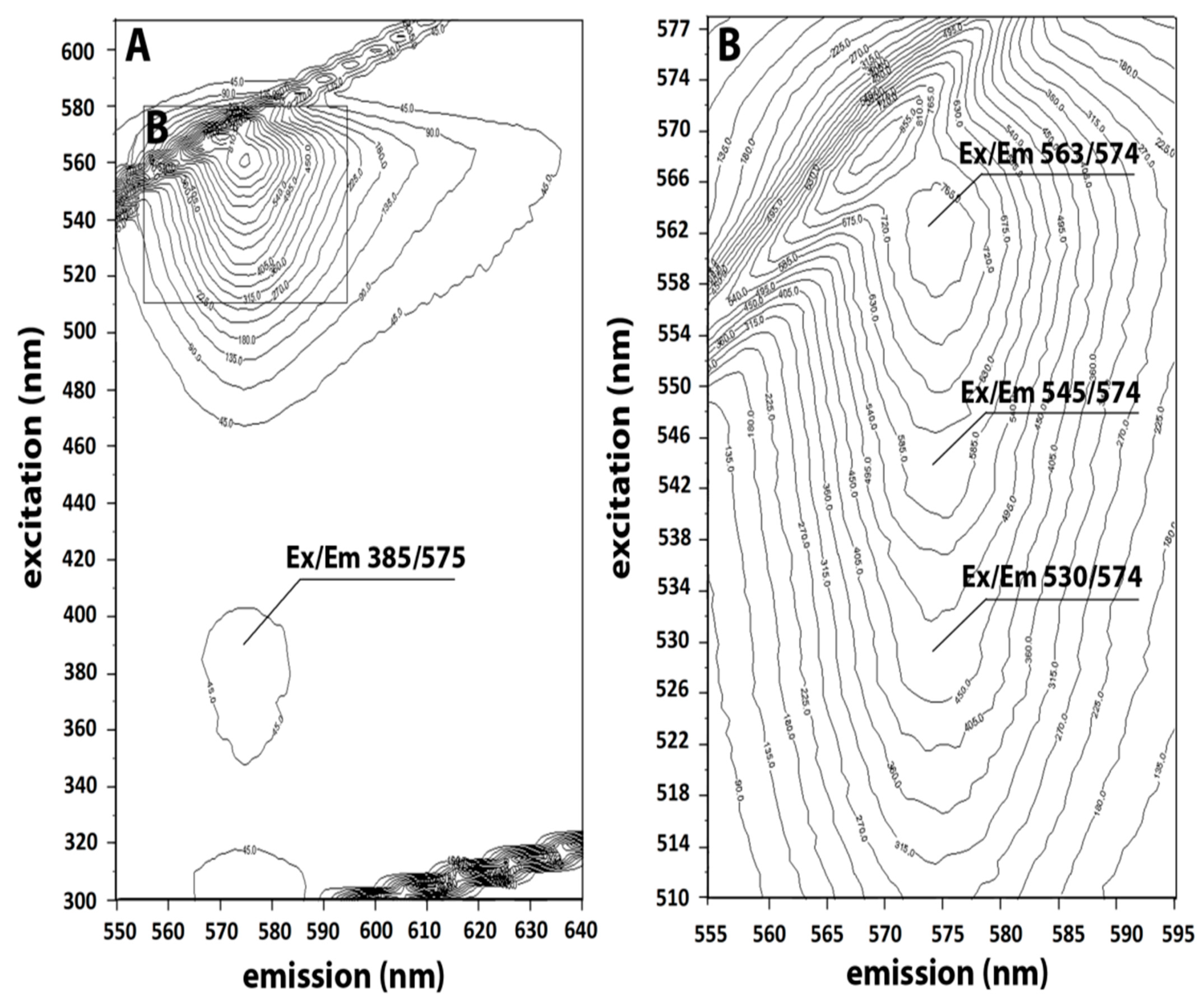
The EEM was recorded by scanning excitation and emission simultaneously in a Luminescent Spectrometer (Perkin Elmer LS 55) equipped with a Xenon discharge lamp and an excitation/emission slit 5/5. The scans were processed by 3D View Perkin Elmer software to produce 3D fingerprint contour maps by using fluorescence lines (with emission plotted on the
X-axis and excitation on the
Y-axis), as previously reported [
37].
The calibration curve of 0.6–6 mg/mL of C-PE solubilized in PB was calculated as follows:
The purity index was calculated as the ratio of the maximum absorbance peak to the absorbance peak of the proteins (A
562/A
280) [
38].
4.3. Evaluation of Oxidative Stress, the Redox Environment, and the Activity of Effector Caspases 3 and 9
Kidneys were homogenized in 3.5 mL of 10 mM PB for all assays. The quantification of oxidative stress, the redox environment, and the activity of effector caspases 3 and 9 was performed with a previously described method [
3,
22].
The lipid peroxidation technique employed an aliquot of 500 µL of homogenate, which was added to 4 mL of chloroform-methanol (2:1, v/v). The mixture was agitated and kept at 4 °C for 30 min (protected from light) to allow for the separation of the polar and nonpolar phases. Afterwards, the aqueous phase was aspirated and discarded. With an aliquot of 2 mL of the organic phase (chloroform), fluorescence was determined at 370 nm (excitation) and 430 nm (emission). The results were expressed as relative fluorescence units (RFU) per mg of protein.
The level of ROS was quantified by the formation of 2,7-dichlorofluorescein (DCF), and 10 μL of the homogenate was added to 1945 µL of TRIS-HEPES (18:1 v/v) and incubated in the presence of 50 µL of 2,7-dichlorofluorescin diacetate (DCFH-DA) at 37 °C for 1 h. The reaction was stopped by freezing, and the fluorescence was measured at 488 nm (excitation) and 525 nm (emission).
Nitrites were assessed as indirect markers of nitrergic stress. An aliquot of 500 µL of homogenate was added to 500 µL of concentrated chlorohydric acid and 500 µL of 20% zinc suspension. The mixture was stirred and incubated at 37 °C for 1 h, followed by centrifugation at 4000× g for 2 min. The supernatant (50 µL) was added to a 96-well polystyrene plate containing 50 µL of 0.6% sulfanilamide and 0.12% N-(naftyl)-ethylenediamine, and then incubated for 15 min at room temperature. The absorbance was measured at 530 nm in a Multiscan Go® plate spectrophotometer.
A determination was made of two redox environment markers, GSH and GSSG, in a sample of 300 µL, treated with 500 µL of 30% phosphoric acid and centrifuged at 10,000× g for 30 min at 4 °C. To analyze GSH, an aliquot of 30 µL of the supernatant was diluted in 1.9 mL of FEDTA (1:10, 100 mM phosphate and 5 mM EDTA), and the mixture was reacted with 100 µL of o-phthaldialdehyde. To assess GSSG, 130 µL of the supernatant was added to 60 µL of N-ethylmaleimide and left for 30 min. Subsequently, an aliquot of 60 µL of the mixture was combined with 1.84 mL of FEDTA and 100 µL of o-phthaldialdehyde. The two chemical species were measured at 350 nm (excitation) and 420 nm (emission).
The activity of caspases 3 and 9 was evaluated using a commercial colorimetric assay kit as specified in the manufacturer’s instructions (Millipore, APT165 and APT173, respectively). Accordingly, p-nitroaniline (pNA) was cleaved from the substrate N-Acetyl-Asp-Glu-Val-Asp p-nitroaniline (DEVD-pNA, caspase 3) or N-Acetyl-Leu-Glu-His-Asp p-nitroaniline (LEHD-pNA, caspase 9), and then the samples were measured spectrophotometrically at 405 nm.
4.4. Examination of Kidney Damage
Histopathological analysis was performed to appraise kidney damage. The kidneys were fixed for 48 h in 4% paraformaldehyde in PBS. Afterwards, they were embedded in paraffin to obtain 5 µm slices on a standard microtome. Each section was stained with H&E, dehydrated, and mounted in resin. The presence or absence of kidney cell damage was evaluated with a histopathological graded scale [
6,
20,
22] as follows: 0, undamaged (indistinguishable from the controls); 1, minimal (affecting ≤25% of the tubules and glomerulus); 2, mild (affecting >25% and ≤50% of the tubules and glomerulus); 3, moderate (affecting >50% and ≤75% of the tubules and glomerulus); 4, severe (affecting >75% of the tubules and glomerulus).
4.5. Western Blot Analysis for Nephrin, Podocin, and ER Stress Markers
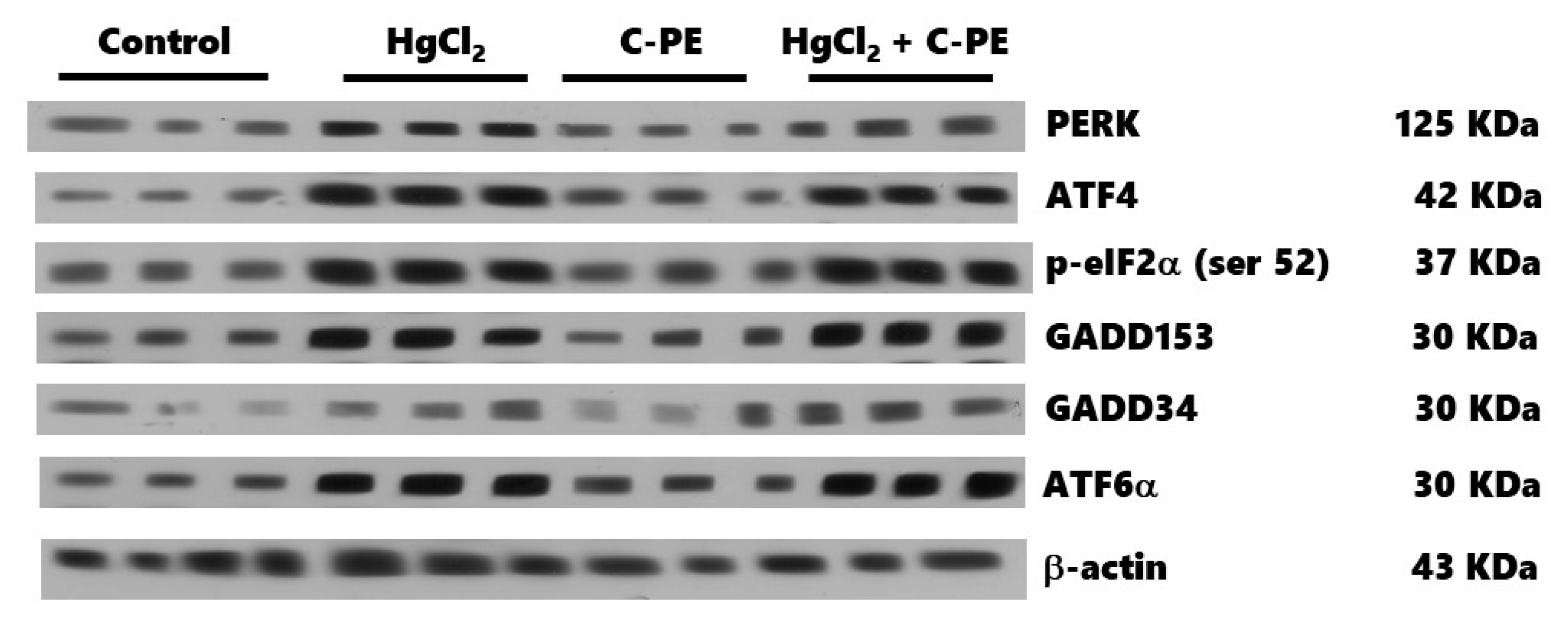
The expression of proteins was determined with Western blot assays. Briefly, the samples were prepared with 100 µL of the homogenate mixed with 50 μL of a complete protease inhibitor cocktail® (MilliporeSigma, Burlington, MA, USA) in lysis buffer, and then 150 μL of the 2× Laemmli sample buffer (Biorad, Hercules, CA, USA, 161-0737) was added. The samples were homogenized by vortex, placed in a boiling water bath for 3 min, and then kept at −20 °C to await processing. Fifty μg of protein samples were loaded in 15% polyacrylamide gel with sodium dodecyl sulfate (SDS-PAGE) and separated by electrophoresis (90 V for 60 min). Subsequently, the proteins were electrotransferred from the gels to PVDF membranes in a Trans-Blot Turbo system (Biorad) at 25 V and 2.05 A for 7 min. Upon completing this time, the membranes were blocked for 1 h under constant stirring in PBST (PBS with 0.05% Tween 20 and 5% low-fat milkSvelty®), followed by incubation overnight at 4 °C in a blocking buffer containing the primary antibodies. The primary antibodies (Santa Cruz Biotechnology, Dallas, TX, USA), diluted 1:1000, were PERK (sc-377400), p-eIF2α (Ser 52, sc-12412), ATF-4 (sc-200), GADD153 (sc-56107), ATF-6α (sc-166659), IRE-1α (sc-390960), p-p53 (Thr 155, sc-377567), XBP1 (sc-7160), Bax (sc-20067), and Bcl2 (sc-7382). Podocin (orb337389), nephrin (orb11107), GADD34 (orb13417), and p53 (orb14498) were acquired from Byorbit (Cambridgeshire, Cambridge, UK) and diluted 1:500. After incubation, membranes were washed three times with fresh PBST (20 min/wash) and then incubated in a secondary antibody diluted 1:1500 (HPR-conjugated goat anti-rabbit; Life Technologies, Rockford, IL, USA, 65-6120) at room temperature for 1 h under constant stirring. Membranes were washed three times with fresh PBST. Finally, the protein bands were revealed on photographic plates (JUAMA, Mexico City, Mexico) by chemiluminescence, using Luminata TM Forte® (MilliporeSigma, Burlington, MA, USA). β-Actin protein expression served as the loading control and constitutive protein (Santa Cruz Biotechnology; sc-1615, dilution 1:4000). The optical density (OD) of all bands was quantified by the Image J program (NIH, Bethesda, MD, USA) and described as the protein/β-actin ratio.
4.6. Statistical Analysis
All data are expressed as the mean ± standard error, except for the kidney damage score. The latter is described as the median ± interquartile spaces, with the values analyzed with the Kruskal–Wallis method. The variables in the first protocol (to evaluate oxidative stress and the redox environment) were examined by one-way analysis of variance (ANOVA). Two-way ANOVA was utilized to assess ER stress, considering the treatment and absence/presence of AKI as factors. ANOVA was followed by the Student-Newman-Keuls post hoc test. Statistical significance was considered at p < 0.05.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}