α-Conotoxin TxIB: A Uniquely Selective Ligand for α6/α3β2β3 Nicotinic Acetylcholine Receptor Attenuates Nicotine-Induced Conditioned Place Preference in Mice


Abstract
:1. Introduction
2. Results
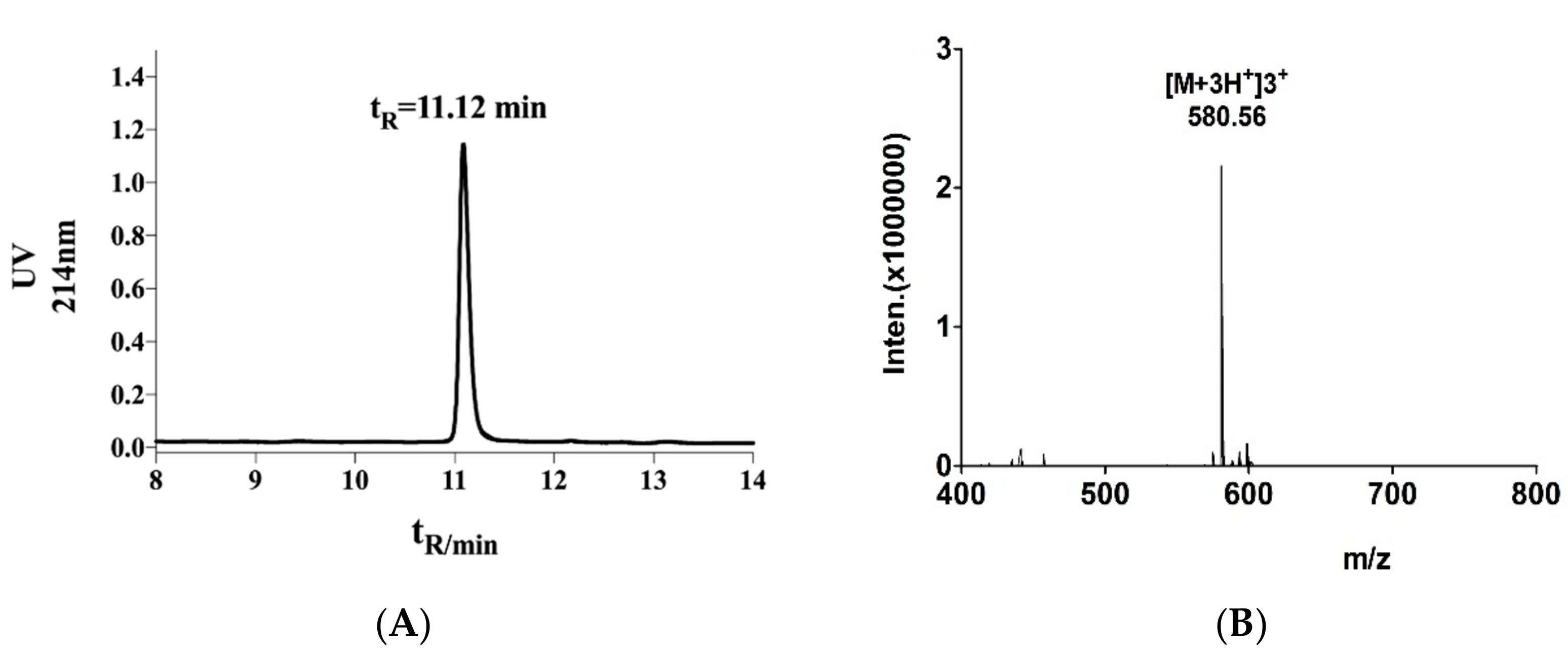
2.1. Structural Identification of α-Conotoxin TxIB
2.2. Nicotine-Induced Conditioned Place Preference in Mice
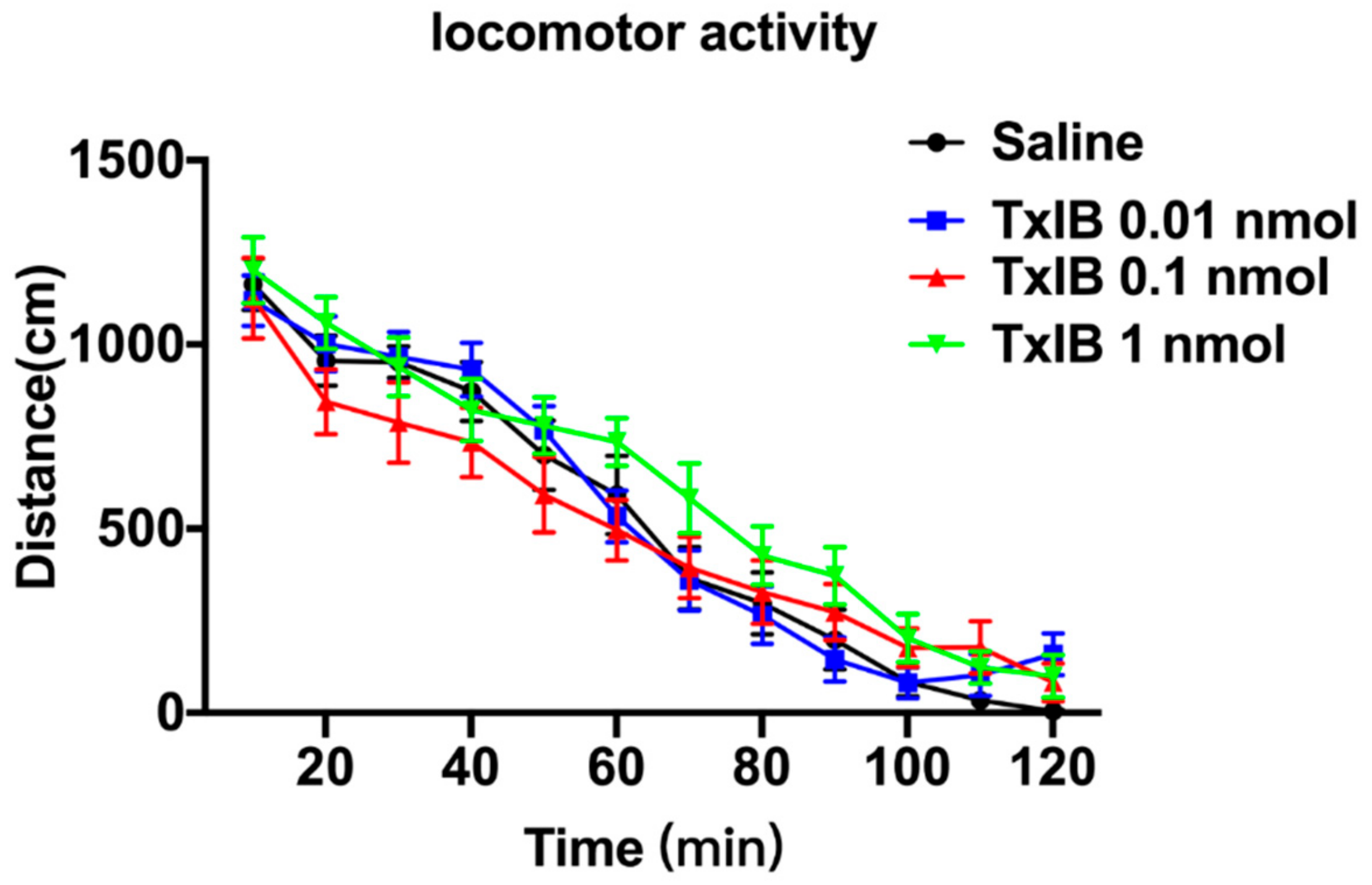
2.3. Effect of Different Doses of TxIB on Locomotor Activity
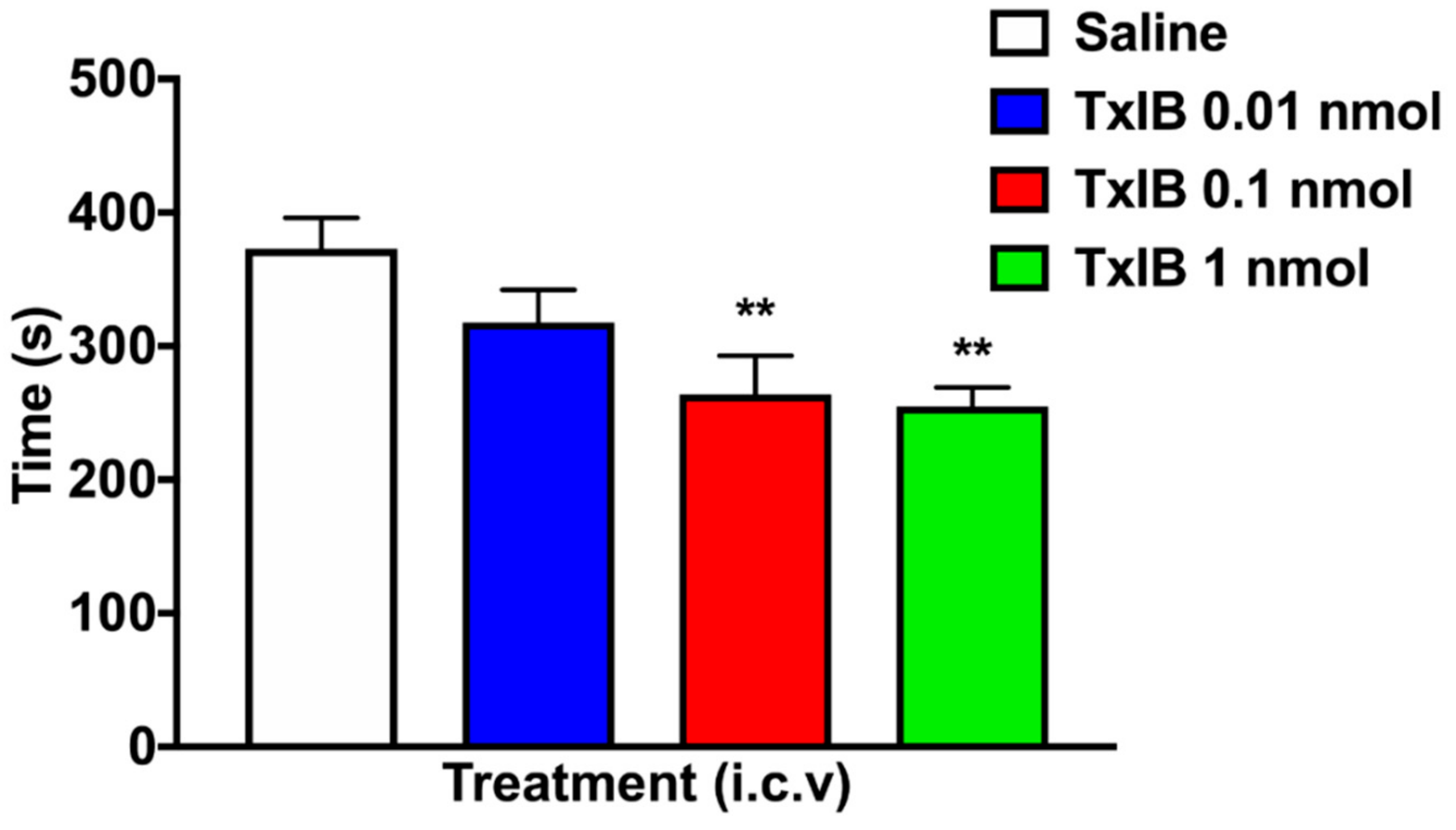
2.4. Effect of Different Doses of TxIB on Inhibition of CPP Expression
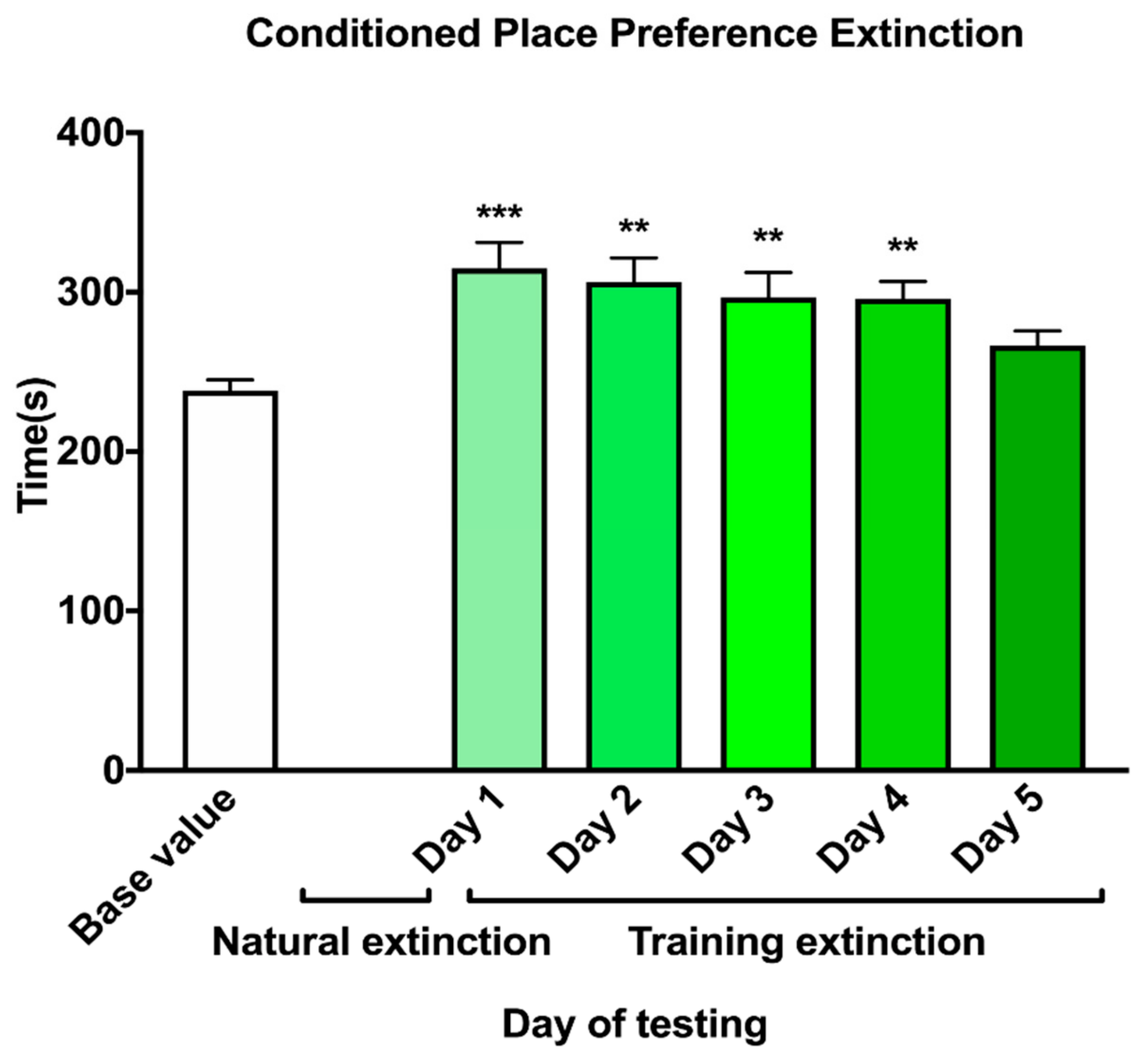
2.5. Extinction of Nicotine-Induced CPP
2.6. Effect of Different Doses of TxIB on Inhibition of CPP Reinstatement
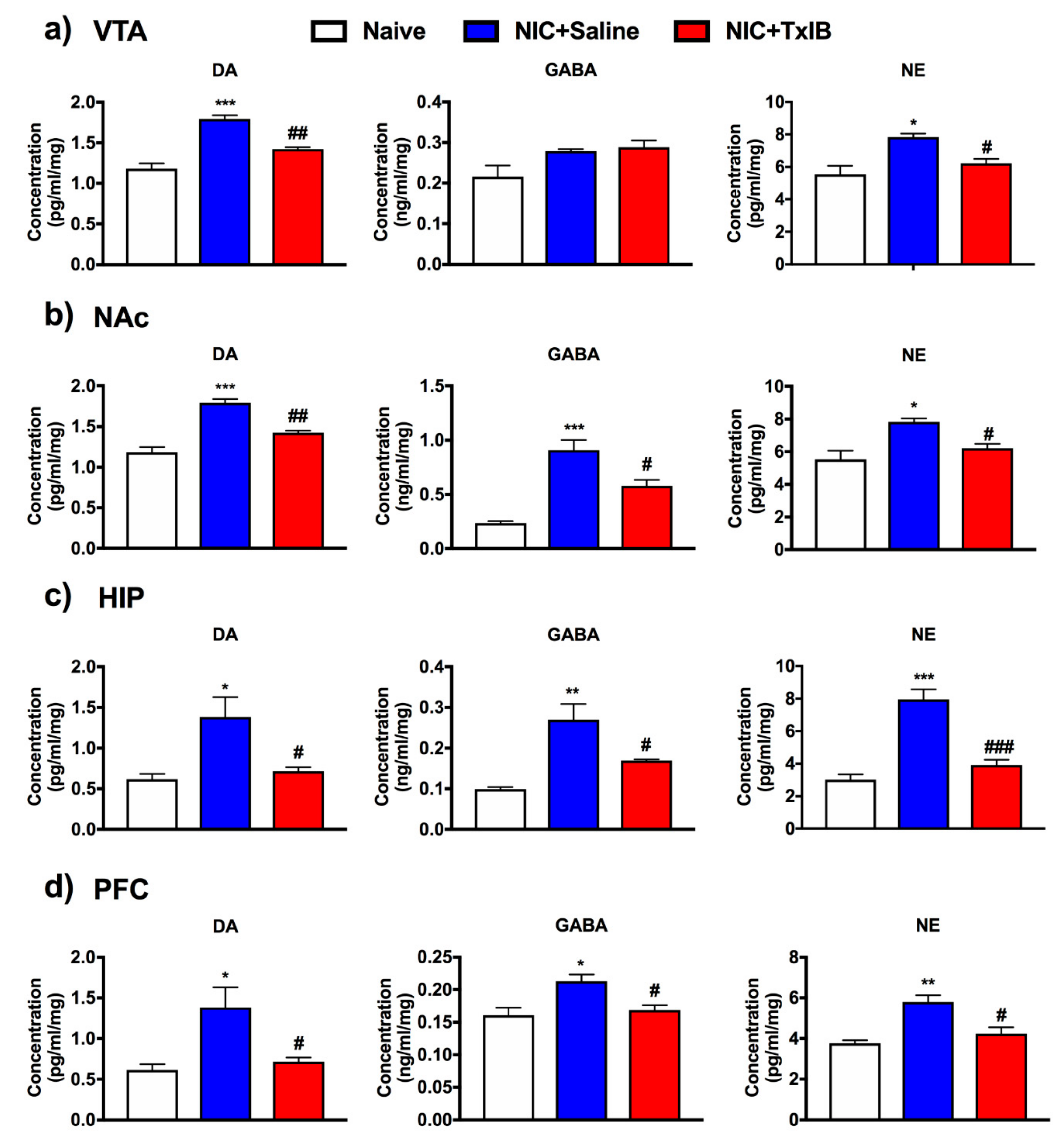
2.7. Changes of Different Neurotransmitters in Several Brain Regions
3. Discussion
4. Materials and Methods
4.1. Chemical Synthesis of α-Conotoxin TxIB
4.2. Animals
4.3. Drugs
4.4. Intracerebroventricular Surgery
4.5. Nicotine-Induced Conditioned Place Preference (CPP)
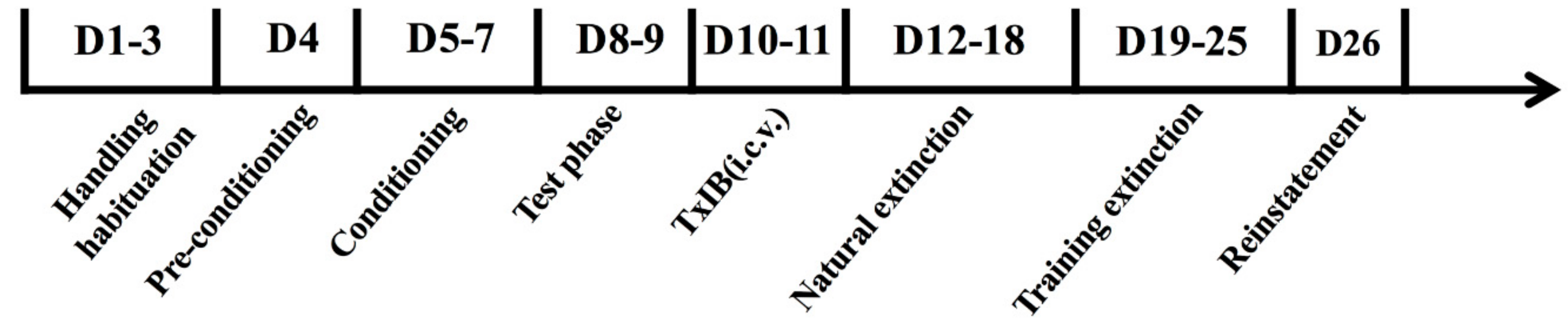
4.6. Treatment Protocol in Animals
4.7. Sample Preparation
4.8. Enzyme-Linked Immunosorbent Assay
4.9. Locomotor Activity
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- National Center for Chronic Disease Prevention and Health Promotion (US) Office on Smoking and Health. The Health Consequences of Smoking—50 Years of Progress: A Report of the Surgeon Heneral; US Department of Health and Human Services: Atlanta, GA, USA, 2014.
- Cippitelli, A.; Wu, J.; Gaiolini, K.A.; Mercatelli, D.; Schoch, J.; Gorman, M.; Ramirez, A.; Ciccocioppo, R.; Khroyan, T.V.; Yasuda, D.; et al. AT-1001: A high-affinity alpha3beta4 nAChR ligand with novel nicotine-suppressive pharmacology. Br. J. Pharmacol. 2015, 172, 1834–1845. [Google Scholar] [CrossRef] [PubMed]
- Corringer, P.J.; Novère, N.L.; Changeux, J.P. Nicotinic receptors at the amino acid level. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 431–458. [Google Scholar] [CrossRef] [PubMed]
- Changeux, J.P.; Corringer, P.J.; Maskos, U. The nicotinic acetylcholine receptor: From molecular biology to cognition. Neuropharmacology 2015, 96, 135–136. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.S.; Layer, R.T.; McCabe, R.T. Conopeptides: From deadly venoms to novel therapeutics. Drug Discov. Today 2000, 5, 98–106. [Google Scholar] [CrossRef]
- Perry, D.C.; Mao, D.; Gold, A.B.; McIntosh, J.M.; Pezzullo, J.C.; Kellar, K.J. Chronic nicotine differentially regulates alpha6- and beta3-containing nicotinic cholinergic receptors in rat brain. J. Pharmacol. Exp. Ther. 2007, 322, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.J.; Martin, B.R.; Changeux, J.P.; Damaj, M.I. Differential Role of Nicotinic Acetylcholine Receptor Subunits in Physical and Affective Nicotine Withdrawal Signs. J. Pharmacol. Exp. Ther. 2008, 325, 302–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, J.M.; Azam, L.; Staheli, S.; Dowell, C.; Lindstrom, J.M.; Kuryatov, A.; Garrett, J.E.; Marks, M.J.; Whiteaker, P. Analogs of alpha-conotoxin MII are selective for alpha6-containing nicotinic acetylcholine receptors. Mol. Pharmacol. 2004, 65, 944–952. [Google Scholar] [CrossRef]
- Berry, J.N.; Engle, S.E.; McIntosh, J.M.; Drenan, R.M. α6-Containing nicotinic acetylcholine receptors in midbrain dopamine neurons are poised to govern dopamine-mediated behaviors and synaptic plasticity. Neuroscience 2015, 304, 161–175. [Google Scholar] [CrossRef]
- Quik, M.; Perez, X.A.; Grady, S.R. Role of alpha6 nicotinic receptors in CNS dopaminergic function: Relevance to addiction and neurological disorders. Biochem. Pharmacol. 2011, 82, 873–882. [Google Scholar] [CrossRef]
- Picciotto, M.R.; Zoli, M.; Rimondini, R.; Lena, C.; Marubio, L.M.; Pich, E.M.; Fuxe, K.; Changeux, J.P. Acetylcholine receptors containing the beta2 subunit are involved in the reinforcing properties of nicotine. Nature 1998, 391, 173–177. [Google Scholar] [CrossRef]
- Epping-Jordan, M.P.; Picciotto, M.R.; Changeux, J.P.; Pich, E.M. Assessment of nicotinic acetylcholine receptor subunit contributions to nicotine self-administration in mutant mice. Psychopharmacology 1999, 147, 25–26. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Jones, A.; Lewis, R.J. Remarkable inter- and intra-species complexity of conotoxins revealed by LC/MS. Peptides 2009, 30, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Rivier, J.; Clark, C.; Ramilo, C.A.; Corpuz, G.P.; Abogadie, F.C.; Mena, E.E.; Woodward, S.R.; Hillyard, D.R.; Cruz, L.J. Diversity of Conus neuropeptides. Science 1990, 249, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus venom peptide pharmacology. Pharmacol. Rev. 2012, 64, 259–298. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Jin, A.H.; Kaas, Q.; Jones, A.; Alewood, P.F.; Lewis, R.J. Deep venomics reveals the mechanism for expanded peptide diversity in cone snail venom. Mol. Cell. Proteom. 2013, 12, 312–329. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Wu, Y.; Zhu, X.; Hu, Y.; McIntyre, M.; Christensen, S.; Akcan, M.; Craik, D.J.; McIntosh, J.M. Characterization of a novel alpha-conotoxin from conus textile that selectively targets alpha6/alpha3beta2beta3 nicotinic acetylcholine receptors. J. Biol. Chem. 2013, 288, 894–902. [Google Scholar] [CrossRef]
- Baumeister, R.; Heatherton, T.; Tice, D. How and why people fail at self-regulation. In Losing Control; Academic Press: New York, NY, USA, 1994; p. 307. [Google Scholar]
- Koob, G.F.; Le Moal, M. Drug addiction, dysregulation of reward, and allostasis. Neuropsychopharmacology 2001, 24, 97–129. [Google Scholar] [CrossRef]
- Castañé, A.; Valjent, E.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. Lack of CB1 cannabinoid receptors modifies nicotine behavioural responses, but not nicotine abstinence. Neuropharmacology 2002, 43, 857–867. [Google Scholar]
- Berrendero, F.; Mendizabal, V.; Robledo, P.; Galeote, L.; Bilkei-Gorzo, A.; Zimmer, A.; Maldonado, R. Nicotine-induced antinociception, rewarding effects, and physical dependence are decreased in mice lacking the preproenkephalin gene. J. Neurosci. 2005, 25, 1103–1112. [Google Scholar] [CrossRef]
- Castañé, A.; Soria, G.; Ledent, C.; Maldonado, R.; Valverde, O. Attenuation of nicotine-induced rewarding effects in A 2A knockout mice. Neuropharmacology 2006, 51, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.J.; McIntosh, J.M.; Brunzell, D.H.; Sanjakdar, S.S.; Damaj, M.I. The role of alpha6-containing nicotinic acetylcholine receptors in nicotine reward and withdrawal. J. Pharmacol. Exp. Ther. 2009, 331, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Sanjakdar, S.S.; Maldoon, P.P.; Marks, M.J.; Brunzell, D.H.; Uwe, M.; McIntosh, M.J.; Bowers, M.S.; Damaj, M.I. Differential roles of α6β2* and α4β2* neuronal nicotinic receptors in nicotine- and cocaine-conditioned reward in mice. Neuropsychopharmacology 2015, 40, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Pons, S.; Fattore, L.; Cossu, G.; Tolu, S.; Porcu, E.; McIntosh, J.M.; Changeux, J.P.; Maskos, U.; Fratta, W. Crucial role of alpha4 and alpha6 nicotinic acetylcholine receptor subunits from ventral tegmental area in systemic nicotine self-administration. J. Neurosci. 2008, 28, 12318–12327. [Google Scholar] [CrossRef] [PubMed]
- Benwell, M.E.; Balfour, D.J. The effects of acute and repeated nicotine treatment on nucleus accumbens dopamine and locomotor activity. Br. J. Pharmacol. 2012, 105, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Drenan, R.M.; Grady, S.R.; Whiteaker, P.; McClure-Begley, T.; McKinney, S.; Miwa, J.M.; Bupp, S.; Heintz, N.; McIntosh, J.M.; Bencherif, M.; et al. In vivo activation of midbrain dopamine neurons via sensitized, high-affinity alpha 6 nicotinic acetylcholine receptors. Neuron 2008, 60, 123–136. [Google Scholar] [CrossRef]
- Dani, J.A.; Heinemann, S. Molecular and cellular aspects of nicotine abuse. Neuron 1996, 16, 905–908. [Google Scholar] [CrossRef]
- Grenhoff, J.; Aston-Jones, G.; Svensson, T.H. Nicotinic effects on the firing pattern of midbrain dopamine neurons. Acta Physiol. Scand. 1986, 128, 351–358. [Google Scholar] [CrossRef]
- Pontieri, F.E.; Tanda, G.; Orzi, F.; Chiara, G.D. Effects of nicotine on the nucleus accumbens and similarity to those of addictive drugs. Nature 1996, 382, 255. [Google Scholar] [CrossRef]
- Quarta, D.; Smolders, I. Rewarding, reinforcing and incentive salient events involve orexigenic hypothalamic neuropeptides regulating mesolimbic dopaminergic neurotransmission. Eur. J. Pharm. Sci. 2014, 57, 2–10. [Google Scholar] [CrossRef]
- Siahposht-Khachaki, A.; Fatahi, Z.; Yans, A.; Khodagholi, F.; Haghparast, A. Involvement of AMPA/Kainate Glutamate receptor in the extinction and reinstatement of morphine-induced conditioned place preference: A behavioral and molecular study. Cell. Mol. Neurobiol. 2017, 37, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Polosa, R.; Benowitz, N.L. Treatment of nicotine addiction: Present therapeutic options and pipeline developments. Trends Pharmacol. Sci. 2011, 32, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Dani, J.A.; Ji, D.; Zhou, F.M. Synaptic plasticity and nicotine addiction. Neuron 2001, 31, 349–352. [Google Scholar] [CrossRef]
- Klink, R.; de Kerchove d’Exaerde, A.; Zoli, M.; Changeux, J.P. Molecular and physiological diversity of nicotinic acetylcholine receptors in the midbrain dopaminergic nuclei. J. Neurosci. 2001, 21, 1452–1463. [Google Scholar] [CrossRef]
- Le Novère, N.; Corringer, P.-J.; Changeux, J.-P. The diversity of subunit composition in nAChRs: Evolutionary origins, physiologic and pharmacologic consequences. J. Neurobiol. 2002, 53, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Champtiaux, N.; Han, Z.Y.; Bessis, A.; Rossi, F.M.; Zoli, M.; Marubio, L.; McIntosh, J.M.; Changeux, J.P. Distribution and pharmacology of α6-containing nicotinic acetylcholine receptors analyzed with mutant mice. Neuroscience 2002, 22, 1208–1217. [Google Scholar] [CrossRef] [PubMed]
- Zoli, M.; Moretti, M.; Zanardi, A.; McIntosh, J.M.; Clementi, F.; Gotti, C. Identification of the nicotinic receptor subtypes expressed on dopaminergic terminals in the rat striatum. J. Neurosci. 2002, 22, 8785–8789. [Google Scholar] [CrossRef] [PubMed]
- Reena, G.; Jayati, B.; Yi-Pin, C.; Phillips, A.M.; Lawrence, T.; Armishaw, C.J. Oxidative folding and preparation of α-conotoxins for use in high-throughput structure-activity relationship studies. J. Pept. Sci. 2013, 19, 16–24. [Google Scholar]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Varani, A.P.; Aso, E.; Moutinho, L.M.; Maldonado, R.; Balerio, G.N. Attenuation by baclofen of nicotine rewarding properties and nicotine withdrawal manifestations. Psychopharmacology 2014, 231, 3031–3040. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, R. Participation of noradrenergic pathways in the expression of opiate withdrawal: Biochemical and pharmacological evidence. Neurosci. Biobehav. Rev. 1997, 21, 91–104. [Google Scholar] [CrossRef]
- Grabus, S.D.; Martin, B.R.; Brown, S.E.; Damaj, M.I. Nicotine place preference in the mouse: Influences of prior handling, dose and strain and attenuation by nicotinic receptor antagonists. Psychopharmacology 2006, 184, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Kota, D.; Martin, B.R.; Robinson, S.E.; Damaj, M.I. Nicotine dependence and reward differ between adolescent and adult male mice. J. Pharmacol. Exp. Ther. 2007, 322, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Myers, K.M.; Carlezon, W.A., Jr. Extinction of drug- and withdrawal-paired cues in animal models: Relevance to the treatment of addiction. Neurosci. Biobehav. Rev. 2010, 35, 285–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, M.A.; Rodriguez-Arias, M.; Minarro, J. Neurobiological mechanisms of the reinstatement of drug-conditioned place preference. Brain Res. Rev. 2009, 59, 253–277. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Base Value before CPP | Base Value after CPP | Base Value after Surgery |
|---|---|---|---|
| Saline | 246.2 ± 7.881 | 252 ± 9.766 | 272.1 ± 17.06 |
| NIC | 236.3 ± 9.353 | 399.6 ± 16.83 **** | 399.6 ± 17.91 **** |
| Treatment Group | Total Distance in Drug-Paired Compartment |
|---|---|
| Naive | 730.8 ± 21.19 |
| NIC | 939 ± 42.86 * |
| NIC + Saline | 661.7 ± 47.47 |
| NIC + TxIB 0.01 nmol | 626.2 ± 82 |
| NIC + TxIB 0.1 nmol | 704.9 ± 46.57 |
| NIC + TxIB 1 nmol | 683 ± 65.21 |
| Neurotransmitters | DA | GABA | NE | ||||
|---|---|---|---|---|---|---|---|
| Brain Regions | F Value | p Value | F Value | p Value | F Value | p Value | |
| VTA | F(2, 6) = 42.05 | <0.001 | F(2, 8) = 5.074 | <0.05 | F(2, 6) = 10.25 | <0.05 | |
| NAc | F(2, 6) = 42.05 | <0.001 | F(2, 7) = 22.17 | <0.001 | F(2, 6) = 10.25 | <0.05 | |
| HIP | F(2, 6) = 7.267 | <0.05 | F(2, 9) = 14.18 | <0.01 | F(2, 9) = 36.01 | <0.0001 | |
| PFC | F(2, 7) = 9.148 | <0.05 | F(2, 7) = 8.468 | <0.05 | F(2, 8) = 12.25 | <0.01 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, S.; Li, X.; Xiong, J.; Zhu, X.; Zhangsun, D.; Zhu, X.; Luo, S. α-Conotoxin TxIB: A Uniquely Selective Ligand for α6/α3β2β3 Nicotinic Acetylcholine Receptor Attenuates Nicotine-Induced Conditioned Place Preference in Mice. Mar. Drugs 2019, 17, 490. https://doi.org/10.3390/md17090490
You S, Li X, Xiong J, Zhu X, Zhangsun D, Zhu X, Luo S. α-Conotoxin TxIB: A Uniquely Selective Ligand for α6/α3β2β3 Nicotinic Acetylcholine Receptor Attenuates Nicotine-Induced Conditioned Place Preference in Mice. Marine Drugs. 2019; 17(9):490. https://doi.org/10.3390/md17090490
Chicago/Turabian StyleYou, Shen, Xiaodan Li, Jian Xiong, Xiaoyu Zhu, Dongting Zhangsun, Xiaopeng Zhu, and Sulan Luo. 2019. "α-Conotoxin TxIB: A Uniquely Selective Ligand for α6/α3β2β3 Nicotinic Acetylcholine Receptor Attenuates Nicotine-Induced Conditioned Place Preference in Mice" Marine Drugs 17, no. 9: 490. https://doi.org/10.3390/md17090490