Quorum Sensing Inhibitors from Marine Microorganisms and Their Synthetic Derivatives
by
 and
and
Jianwei Chen
1,†, 
Bixia Wang
1,†,
Yaojia Lu
1,
Yuqi Guo
1,
Jiadong Sun
2,
Bin Wei
1,
Huawei Zhang
1 and
Hong Wang
1,* 1
College of Pharmaceutical Science & Collaborative Innovation Center of Yangtze River Delta Region Green Pharmaceuticals, Zhejiang University of Technology, Hangzhou 310014, China
2
Laboratory of Bioorganic Chemistry, National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK), National Institutes of Health, Bethesda, MD 20878, USA
*
Author to whom correspondence should be addressed.
†
These authors contribute equally to this work.
Mar. Drugs 2019, 17(2), 80; https://doi.org/10.3390/md17020080
Submission received: 16 December 2018
/
Revised: 19 January 2019
/
Accepted: 23 January 2019
/
Published: 28 January 2019
(This article belongs to the Collection Papers from “Sino–Italian Symposium on Bioactive Natural Products”)
Abstract
:Quorum sensing inhibitors (QSIs) present a promising alternative or potent adjuvants of conventional antibiotics for the treatment of antibiotic-resistant bacterial strains, since they could disrupt bacterial pathogenicity without imposing selective pressure involved in antibacterial treatments. This review covers a series of molecules showing quorum sensing (QS) inhibitory activity that are isolated from marine microorganisms, including bacteria, actinomycetes and fungi, and chemically synthesized based on QSIs derived from marine microorganisms. This is the first comprehensive overview of QSIs derived from marine microorganisms and their synthetic analogues with QS inhibitory activity.
1. Introduction
We have found ourselves facing a significant problem in modern healthcare settings where many anti-infective drugs have lost their effectiveness against life-threatening and debilitating diseases [1,2]. The pathogens have outplaced our abilities to sustainably manage them. Thus, there is an urgent need to discover new types of antimicrobial compounds and novel mechanisms for disease prevention and treatment. One competitive antimicrobial advantage proposed in targeting quorum sensing (QS) is that the treatment of quorum sensing inhibitors (QSIs) does not inhibit bacterial growth and does not also exert a selective pressure to develop bacterial resistance to this treatment. Granted, interference of QS will likely decrease bacterial fitness for survival under certain conditions, but if a delicate control is performed on pathogenic QS-regulated genes, then developing resistance mechanisms against QS-inhibiting therapies may be a difficult proposition in pathogenic bacteria.
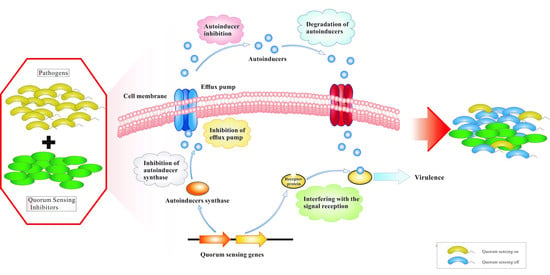
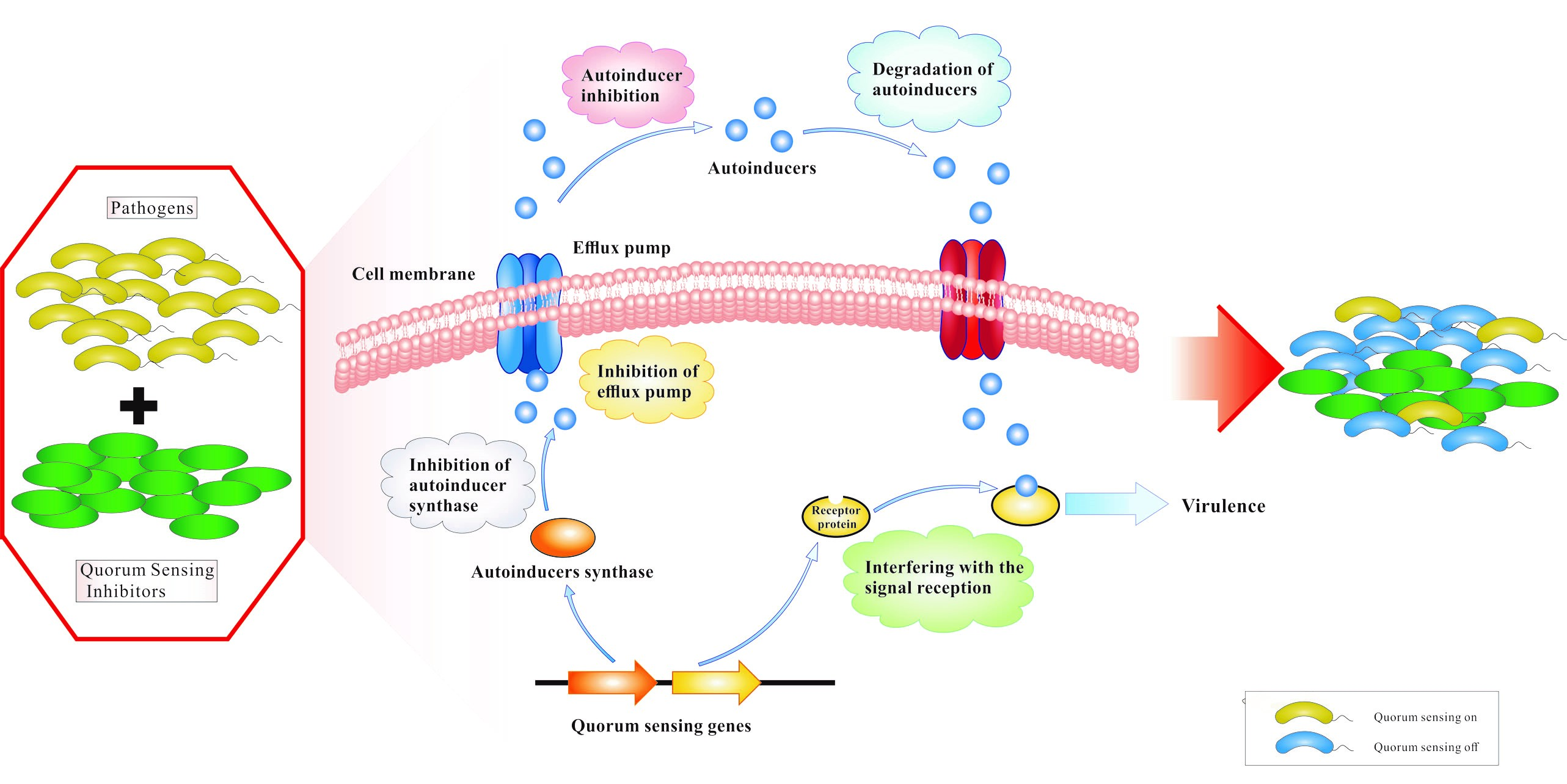
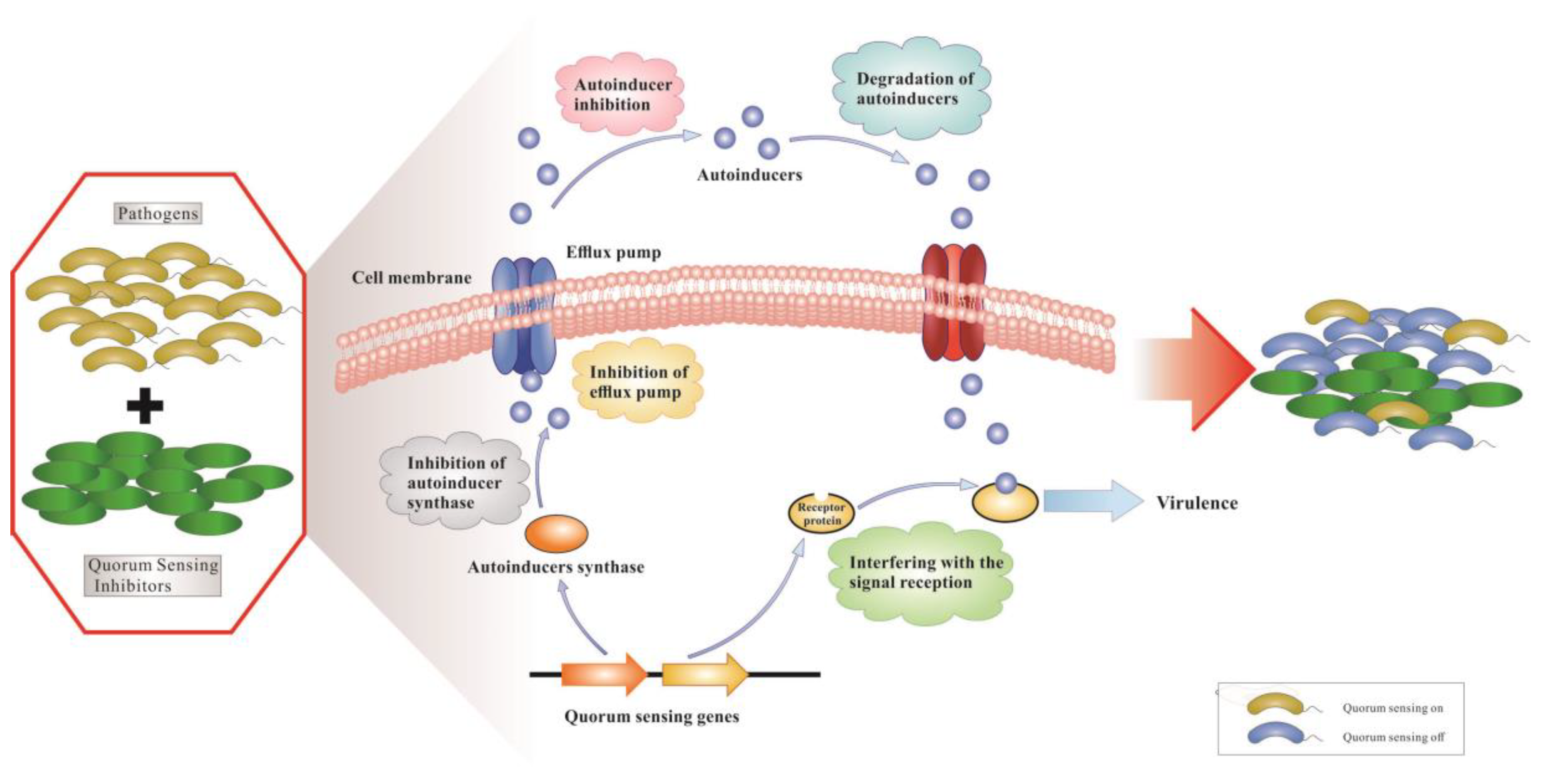
QS is a cell–cell communication process that enables bacteria to regulate their collective behaviors in response to population density and species composition changes in the surrounding environment. It allows bacteria to synchronize gene expression by virtue of extracellular signaling molecules called autoinducers. These autoinducers are released into the surrounding environment where they could be recognized by specific receptors that reside either in the cytoplasm or in the membrane. When autoinducers reach a certain threshold concentration, a signal cascade is triggered that promotes synchronous gene expression in the population of bacteria, such as bioluminescence, the secretion of virulence factors, the formation of biofilm, and other biological behaviors. Commonly, gram-negative bacteria use the cytoplasmic transcription factors, LuxR-type QS receptors, to detect N-acyl-homoserine lactones (AHLs) produced by partner LuxI-type synthases. AHLs are most commonly QS autoinducers used by gram-negative bacteria. Exceptions to this are other QS signals, including autoinducer-2, diffusible signal factor, Pseudomonas quinolone signal and new molecules [2,3]. Gram-positive bacteria do not harbor LuxI/R homologues and instead utilize unmodified or modified oligopeptides as autoinducers. At present, many known autoinducers are bound by a membrane-bound sensor kinase located in the cell inner membrane, which switches its phosphatase and kinase activity in response to interaction with peptides, which changes the phosphorylation state of bacterial cognate response regulators and finally leads to activation or inhibition of QS target genes [4]. Both gram-positive and gram-negative bacteria use the QS system, and interfering with QS has been identified as a potential novel targeted therapeutic strategy to treat bacterial infections [5,6,7,8]. For example, gram-negative bacterial QS inhibition by QSIs is depicted in Figure 1. We display different mechanisms of action against a QS system; (a) inhibition of autoinducer synthases or decrease of activity of receptor proteins; (b) inhibition of autoinducer biosynthesis; (c) degradation of autoinducers; and (d) interference with binding of autoinducers and receptor proteins by competitive binding of autoinducer analogues and receptor proteins. For pathogens that regulate virulence via signaling molecules, QS interference also renders bacterial infections more benign and promotes the host innate immune system to more effectively eradicate the pathogens.
Marine microbial species, due to marine chemodiversity, have been considered as an untapped source for unique chemical leads with numerous biological activities [9,10,11]. These compounds provided a wide range of valuable drug candidates for treating various diseases in plants, animals and humans. However, marine microbial species have not been fully exploited like their terrestrial counterparts; according to the statistics, valuable compounds derived from marine environments have been discovered to a much lower extent (1%) than terrestrial environments so far, suggesting the very low percentage of metabolites isolated from marine microbial species [12]. Also, some evidence suggests that QS is a frequent phenomenon in marine environments [13]; QSIs have been found in diverse marine microbial species, such as marine bacteria, actinomycetes and fungi. The aim of this review is to give a comprehensive overview of QSIs from marine microbial species and their synthetic derivatives with QS inhibitory activity.
2. QSIs from Marine Bacteria and Their Derivatives with QS Inhibitory Activity
2.1. QSIs from Marine Gram-Positive Bacteria and Their Derivatives with QS Inhibitory Activity
Halophilic microorganisms possess a multitude of bioactive secondary metabolites due to their unique physiological and genetic properties. Halobacillus salinus C42 from a sea grass sample collected in the Point Judith Salt Pond, South Kingstown, RI afforded two phenethylamide metabolites, 2,3-methyl-N-(2’-phenylethyl)-butyramide (1) and N-(2’-phenylethyl)-isobutyramide (2), which were proven nontoxic to a panel of bacteria, fungi and microalgae [14,15]. These compounds inhibited QS-regulated violacein biosynthesis of Chromobacterium violaceum CV026 and green fluorescent protein production of Escherichia coli JB525. They acted as antagonists of bacterial QS by competing with AHL for receptor binding. The cyclo(L-Pro-L-Val) (3) isolated by Haloterrigena hispanica SK-3 could promote the expression of QS-regulated genes in bacterial AHL reporters, suggesting that archaea have the ability to interact with AHL-producing bacteria in syntrophic communities [16]. In contrast, four different diketopiperazines (DKPs): cyclo(L-Pro-L-Phe) (4), cyclo(L-Pro-L-Leu) (5), cyclo(L-Pro-L-isoLeu) (6), and cyclo(L-Pro-D-Phe) (7) isolated from Marinobacter sp. SK-3 demonstrated their QS-inhibitory activities based on the test of C. violaceum CV017 and E. coli [17]. This indicated that DKPs from microorganisms could activate or inhibit bacterial QS, pointing to a vital role of these molecules within microbial communities.
Three active metabolites isolated from Oceanobacillus sp. XC22919 were identified as 2-methyl-N-(2′-phenylethyl)-butyramide (8), 3-methyl-N-(2′-phenylethyl)-butyramide (9) and benzyl benzoate (10), and were first reported to exhibit the apparent QS inhibitory activities against C. violaceum 026 and Pseudomonas aeruginosa [18]. These molecules could inhibit violacein production in C. violaceum 026, as well as pyocyanin production, elastase and proteolytic enzymes, and biofilm formation in P. aeruginosa. Among them, Compound 8 significantly inhibited the formation of biofilm of P. aeruginosa, with a maximum of 50.6% inhibition, at 100 μg/mL. Saurav et al. [19] performed bioassay-guided isolation from three bacterial isolates of sponges (Nautella sp., Erythrobacter sp. CUA-870, and Dietzia maris IHBB 9296). The isolates Cc27, Pv86 and Pv91were found to be positive for QS inhibitory activity and inhibited the formation of biofilm by over 50% in tested strains (E. coli, P. aeruginosa PAO1, and Bacillus subtilis). Finally, nine main secondary metabolites (11–19) were identified in Cc27 (11–13), Pv86 (14–16), and Pv91 (17–19) using LC–HRMS/MS.
Two novel depsipeptides, solonamide A (20) and B (21), from a marine Photobacterium were identified by bioassay-guided isolation [20]. They interfere with agr QS activity in the highly virulent, community-acquired strain USA300 and Staphylococcus aureus 8325-4. This is the first report of the agr QS inhibitors from the marine bacteria. Generally, the S. aureus agr QS system includes at least four agr subclasses, and the autoinducing peptide from each class could induce agr in strains of its own class rather than repress agr of other subclasses [21,22,23]. However, solonamide B reduced agr QS expression significantly in three of four known agr classes (agr group I, agr group II, agr group IV), as well as having a minor effect against agr group III in the S. aureus agr system. Moreover, solonamide B significantly decreased the expression of phenol-soluble modulins, directly controlled AgrA and the transcription of agrA, as well as dramatically reduced the overall toxicity of supernatants towards human neutrophils. This indicated that solonamide B not only interfered with the expression of AgrA and agrA, but also repressed biosynthesis of virulence factors controlled by the agr QS system [24]. Further analysis demonstrated that solonamide B interfered with agr QS activation by preventing interactions between AgrC sensor histidine kinase and S. aureus autoinducing peptides. Structural comparison of solonamide B and autoinducing peptides suggests that the ability to interfere with different agr QS classes is related to the cyclic structure of solonamide B, and the differences observed may correlate with the temporal RNAIII induction pattern or the individual structures of autoinducing peptides [25].
In order to further elaborate structure–function relationships for AgrC QS antagonists, an array of 27 lactam hybrid analogues based on solonamide B and autoinducing peptides were designed and tested for AgrC-inhibitory activity [26]. Among them, 21 compounds (22–41) showed inhibition on the S. aureus QS system. However, there was considerable difference for their inhibitory activity. Hybrid analogues with all-L stereochemistry of the amino acids (22) were equipotent to AgrC inhibitors, solonamides A and B; however, compounds 25, 27 and 33–35 were 20- to 40-fold higher in the inhibition of AgrC than the starting hit compound 22. The structure–activity relationship indicates several structural features are very important determinants for AgrC inhibition (Figure 2); (a) ring size must be identical to the known autoinducing peptides; (b) the tail should be preferably selected from short fatty acid moieties; (c) a Phe residue shows more potent inhibition than other aliphatic or aromatic residues in residue no. 2; (d) residues no. 3 and 4 need to be further studied as there are no clear-cut conclusions; (e) the Leu in residue no. 5 is also crucial for activity.
Morever, Kajerulff et al. [27] reported four novel agr QS-interfering cyclodepsipeptides, ngercheumicin F–I (42–45) from a marine Photobacterium halotolerans. All four ngercheumicins enhanced expression of spa and decreased transcription of hla and rnaIII in the S. aureus lacZ reporter assays. Further studies showed that ngercheumicins reduce expression of rnaIII in the CA-MRSA strain USA300 by Northern blot analysis, suggesting that ngercheumicins interfere with agr QS activation. It can be speculated that these compounds could interfere with QS pathways that exist in the marine environment or even act as a class of novel alternative QS molecules.
2.2. QSIs from Marine Gram-Negative Bacteria
Marine gram-negative bacteria have been discovered to produce QS inhibitory compounds. For example, Vibrio alginolyticus G16 from seaweed Gracilaria gracilis could disrupt QS signaling pathways and reduce biofilm formation in Serratia marcescens. An active compound, phenol, 2,4-bis(1,1-dimethylethyl) (46) was obtained and identified [28]. It could inhibit the QS-mediated virulence factor biosynthesis in S. marcescens and lead to a significant reduction in biofilm (85%), lipase (84%), haemolysin (70%), protease (42%), and extracellular polysaccharide (85%) without affecting bacterial growth. Quantitative PCR analysis confirmed that the N-butanoyl-L-homoserine lactone (C4-HSL)-mediated bsmA gene was obviously downregulated in S. marcescens. Reduction in expression level of bsmA could be related to the ability of phenol, 2,4-bis(1,1-dimethylethyl) to affect QS-regulated biofilm formation. In addition to anti-QS-mediated biofilm inhibition, phenol, 2,4-bis(1,1-dimethylethyl) simultaneously induced hydration of the microbial cell wall, which made it a potential anti-biofilm agent by dual approaches. At the same time, the compound also increased the susceptibility of S. marcescens to gentamicin, which opened another avenue for combination therapy to improve the effectiveness of clinical antibiotics.
Through bioassay-guided fractionation, Sun and coworkers [29] have recently obtained an active diketopiperazine, cyclo(Trp–Ser) (47) from the marine bacterium, Rheinheimera aquimaris QSI02. It decreased QS-regulated violacein biosynthesis (67%) of C. violaceum CV026 and pyocyanin biosynthesis (65%), elastase activity (40%) and biofilm formation (60%) of P. aeruginosa PAO1. The analysis of molecular dynamics suggested that cyclo(Trp–Ser) binds more easily to the LasR receptor than natural QS-signaling molecules (AHLs), but the opposite is true in the LasR receptor. These results demonstrated that cyclo(Trp–Ser) not only efficiently inhibited the biosynthesis of violacein in C. violaceum CV026, but also reduced the formation of biofilm and other QS-mediated phenotypes in P. aeruginosa PAO1. In addition to these small molecules, MomL (48), a novel AHL lactonase derived from marine Muricauda olearia Th120, also significantly attenuated the virulence factor production (protease and pyocyanin) of P. aeruginosa PAO1. The growth of PAO1 was almost not affected, whereas AHL accumulations in the cultures were obviously reduced, suggesting that the reduction of production of virulence factors was due to AHL degradation [30]. Although QS processes were widely distributed in marine microorganisms and QSIs were discovered as a frequent phenomenon in the marine environments [31], QSIs from marine gram-negative bacteria are still very scarce when compared to marine gram-positive bacteria, and there may be drawbacks to probing QSIs in marine gram-negative bacteria using traditional bioassay-guided isolation.
3. QSIs from Marine Actinomycetes and Their Derivatives with QS Inhibitory Activity
As recently reported by Fu and coworkers [32], three new α-pyrones, nocapyrones H (49), I (50) and M (51) from marine-derived actinomycete Nocardiopsis dassonvillei subsp. dassonvillei XG-8-1 exhibited inhibitory activities on QS-controlled gene expression in both C. violaceum CV026 and P. aeruginosa QSIS-lasI biosensors at a concentration of 100 μg/mL. This is the first report of α-pyrones inhibiting QS-regulated gene expression in pathogenic bacteria. Recently, four novel α-pyrones and eight known analogues were also detected in the secondary metabolites of Streptomyces sp. OUCMDZ-3436 isolated from the marine green alga Enteromorpha prolifera [33]. The results of bioassays suggested that these α-pyrones did not exhibit any QS inhibitory activity. However, the skeleton of α-pyrone could be easily transformed into pyridine-2(1H)-one, which had been proved to have a variety of biological activities [34,35,36,37,38,39,40,41,42]. Therefore, based on a diversity-enhanced extracts approach [43,44], four novel α-pyridones (52–55) were obtained and exhibited the inhibitory effect on gene expression regulated by QS in P. aeruginosa QSIS-lasI biosensors at 6.35 μg/well.
In order to further obtain potent QSIs from pyrone-derived compounds, Park and coworkers [45] designed and synthesized several novel pyrone-derived QSIs (56–65) to inhibit the binding of N-(3-oxododecanoyl)-L-homoserine lactone (OdDHL) to the LasR of P. aeruginosa. Among the 10 novel pyrone-derived QSIs, compound 63 exhibited the most potent in-vitro inhibitory activity against biofilm formation. Furthermore, all of the selected QS-inducible genes, including synthase genes (lasI, rhlI, pqsC, pqsD, pqsH, and phnB), and auto-inducer receptor genes (lasR, mvfR, and rhlR), were also significantly downregulated by compound 63. The modeling studies indicated that it mostly interacted with residues in the binding pocket of LasR that was highly similar to the crystal ligand OdDHL. The structure–activity relationship indicated that the pyrone derivatives with more than nine alkyl chains would significantly reduce biofilm formation.
Cyclo(Pro–Gly) (66) and cinnamic acid (67) from marine invertebrate-derived Streptomyces sp. were reported to attenuate P. aeruginosa virulence as QS inhibitors [46]. The in-vivo study suggested that cinnamic acid protected Caenorhabditis elegans from the virulence of P. aeruginosa resulting in reduced mortality [47]. This protective mechanism is likely to be a consequence of competitive suppression of RhlR and LasR receptor proteins by cinnamic acid. A significant reduction in colonization of the bacteria treated with cinnamic acid was also further observed in the nematode. These data were consistent with past results that curcumin increased the survival rate of Caenorhabditis elegans ~28% by decreasing the expression of genes involved in biofilm formation and attenuating the biosynthesis of signal molecules in P. aeruginosa PAO1 [48]. The in-silico analysis showed that it could act as a competitive inhibitor for the natural signal molecules towards active pockets of LasR and RhlR QS circuits in P. aeruginosa.
Several derivatives structurally related to cinnamic acid were also synthesized and assessed for their effects on the QS process. Cinnamyl alcohol (68), methyl trans-cinnamate (69) and allyl cinnamate (70) were able to completely inhibit C. violaceum QS at 1 mM, and cinnamamide (71), 4-chlorocinnamic acid (72), α-methylcinnamic acid (73) and 3,4-(methylenedioxy)cinnamic acid (74) at 5 mM [49]. These data were consistent with past results that 71 and 72 inhibited Vibrio spp. virulence factor biosynthesis in vitro and in vivo [50,51]. However, by contrast, 68 and 69 were devoid of inhibitory activity as QSIs against Vibrio spp. [50]. This suggests that they are selective QSIs against different pathogens. The structure–activity relationship of 67 indicated that the replacement of the carboxylic acid moiety by an aldehyde group (cinnamaldehyde) or the lack of the double bond (3-phenylpropionic acid) leads to inactivity. The replacement of the vinyl acid function by a vinyl sulfone resulted in a more active compound, methyl-styryl sulfone (75). Substituents with electron withdrawing properties increased QS activity. The inhibitory activity decreased in the order 4-trifluoromethyl cinnamic acid (76) > 72 > 2,3,4,5,6-pentafluoro-cinnamic acid (77). The aromatic ring of cinnamic acid was replaced by an alkyl group, and a carboxylic acid moiety was replaced by an acrolein moiety, and these changes still led to active autoinducer-2 signal QS inhibitors, such as (E)-2-pentenal (78), (E)-2-tridecenal (79), methyl-(E)-2-nonenoate (80), and (E)-2-heptenal (81) [50].
The research group of Miao et al. [52] obtained and identified a secondary metabolite, actinomycin D (82) derived Streptomyces parvulus HY026 isolated from a seawater sample, which showed remarkable anti-QS activity. It significantly inhibited the violacein biosynthesis of C. violaceum (65%) at 12.5 μg/mL and prodigiosin production (the pigment inhibition zone of 13.5 mm) of Serratia proteamaculans 657 at 25 μg/disc without affecting bacterial growth. These findings indicated that researchers not only pay attention to the discovery of novel compounds, but also point to known antibiotics for the discovery of new valuable bioactivity.
4. QSIs from Marine Fungi and Their Derivatives with QS Inhibitory Activity
Marine fungi were targeted as potent producers of QSIs, since they possess the ability to synthesize and secrete diverse secondary metabolites, such as peptides, terpenes, polyketide-derived alkaloids, and mixed biosynthesis metabolites [53]. A γ-pyrone derivative, kojic acid (83) from marine-derived fungus Altenaria sp. isolated from marine green alga Ulva pertusa of Pyoseon Beach, Jeju Island, inhibited QS-dependent luminescence of the reporter E. coli pSB401 induced by N-hexanoyl-L-homoserine lactone (C6-HSL) at >36 μM. However, the molecule only interfered with LuxR reporters [54]. Equisetin (84), from a marine fungus Fusarium sp. Z10, inhibited biofilm formation and swarming motility of P. aeruginosa. Further studies showed that the compound inhibited elastase of P. aeruginosa PAO1 and transcriptional activation of lasB in E. coli MG4/pKDT17, attenuated pyocyanin biosynthesis of P. aeruginosa PAO1 and transcriptional activation of PqsA in E. coli pEAL08-2, and declined rhamnolipid biosynthesis, swarming motility and transcriptional activation of rhlA in E. coli pDSY. These data indicated that equisetin could inhibit las, rhl and the PQS system [55]. Asteltoxin (85), a known QS inhibitor from a marine fungus Penicillium sp. QF046, exhibited more potent inhibition of violacein than positive control, (Z-)-4-bromo-5-(bromomethylene)-2(5H)-furanone, and decreased the expression of multiple QS-related genes (lasA, lasB, vioB, vioI, cynS, and hcnB) [56]. A new bacterial enoyl-acyl carrier protein reductase inhibitor, meleagrin (86), from the marine fungus Penicillum chrysogenium isolated from seashore slime of Daechun beach, Chungcheongnam-do, Korea, inhibited QS of C. violaceum CV017 with minimum inhibitory concentration (MIC) of 138.42 mM [57,58]. The fermentation broth of Penicillium sp. SCS-KFD08 isolated from the marine animal Sipunculus nudus collected from Haikou Bay, China led to the isolation of six QSIs (87–92), based on the test of biosensor C. violaceum CV026. These secondary metabolites exhibited obvious anti-QS activity against C. violaceum CV026 at a dosage of 50 μg/well. Among them, compounds 91 and 92 inhibited violacein biosynthesis in C. violaceum CV026 cultures induced by signal molecule C6-HSL by up to 46% and 49% at subminimal inhibitory concentration (sub-MIC) of 300 μM, respectively [59].
In the screening of QS–disrupting molecules, 75 marine fungal isolates were obtained from saline lakes, mangrove rhizosphere and reef organisms. Their QS inhibitory activity was evaluated using C. violaceum CV026. Four strains of endophytic fungi belonging to Sarocladium (LAEE06), Fusarium (LAEE13), Epicoccum (LAEE14), and Khuskia (LAEE21) exhibited potent activity at concentrations ranging from 50 to 500 μg/mL. LC–HRMS analysis of these fungal bioactive metabolites resulted in the identification of several major compounds whose QS inhibitory properties had been known or unknown so far (93–108) [60]. For example, fusaric acid (96) and linoleic acid (108) were isolated from the samples of Fusarium (LAEE13) and Khuskia (LAEE21), respectively, and their ability to interfere with the QS system had been previously reported [61,62,63,64]. Two major components isolated from Epicoccum (LAEE14) were tentatively classified as two DKPs, variecolorin N (104) and phenylahistin (105). They may act as QS agonists or antagonists due to the wide recognition of DKPs as QSIs [65].
Facile and expeditious synthetic strategies have proven to be an efficient tool in obtaining valuable QSIs. Taking advantage of microwave–assisted synthesis, 39 fusaric acid analogues were obtained and tested for their QS inhibitory activity in three QS screening models, luxI-gft, lasB-gft, and rhlA-gft [66]. In the luxI-gft QS system, compounds 109–114 revealed QS inhibition at concentrations from 6.25 to 100 μg/mL. Compound 115 exhibited a little QS inhibition at 3.13 μg/mL whereas at higher concentrations (6.25–100 μg/mL) its QS inhibition was obviously enhanced. In the lasB-gft QS system, compound 116 showed good QS inhibition from 125 μg/mL. A structure–activity relationship for QS inhibition is depicted in Figure 3; (a) the C-2 ester group is essential for inhibition of QS, since it can simulate the intermolecular interaction solicited by the lactone moiety of QS signal molecules; (b) the carboxylic acid substituent at C-2 shows no QS inhibitory activity and inhibits the growth of bacteria; and (c) the alkyl substituent at C-5 is not mandatory, it can be swapped by an aromatic/heterocyclic aromatic ring or alkoxy, even if the compounds exhibit modest QS inhibitory activity.
5. Conclusions and Perspectives
Through reviewing the literature of QSIs from marine microorganisms and their synthetic analogues, it is very clear that 116 QSIs are derived from marine microorganisms and their synthetic analogues (Table 1). Therefore, more effort needs to be made towards the promising strains from marine microorganisms for the discovery of novel QSIs. Moreover, the majority of QSIs from marine microorganisms and their synthetic analogues identified to date function as useful chemical probes for mechanistic or structural studies rather than lead-like compounds for further anti-infective drug development.
In addition, the current lack of methodological standardization in assessing effectiveness of QSI candidates limits the broad validity of any findings. The use of only laboratory-adapted strains may lead to a major pitfall in this field, since these strains may be distant from pathogens of relevant clinical infections. Therefore, further screening of QSIs in conditions that mimic in-vivo pathogenic infections will be vital for the future study and development of QSIs. It is worth noting that QSIs are most likely to be beneficial as potent adjuvants of conventional antibiotics for the treatment of clinical treatment rather than as standalone therapeutic agents, as QSIs allow for temporal control of virulence gene expression rather than result in selective pressures on bacterial survival [67,68,69,70]. For example, the QSI pyrizine–2–carboxylic acid significantly increased the susceptibility of antibiotics (tetracycline, doxycycline, erythromycin and chloramphenicol) against multidrug-resistant Vibrio cholerae [71]. Undoubtedly, simultaneous use of inhibitors for various targets including the QS system will contribute to combating multiantibiotic–resistant bacteria [72,73].
Author Contributions
J.C. and B.W. contributed equally to this work. J.C. and B.W. conceived and wrote the review; Y.G. edited the chemical structures; Y.L., B.W. and J.S. offered important advice to improve the review; H.Z. and H.W. conceived the review and revised the paper.
Funding
This project was supported by National Natural Science Foundation of China (No. 81773628, 81741165, and No. 41776139), National Key R & D Program of China (No. 2017YFE0103100 and 2018YFC0311003), China Postdoctoral Science Foundation (2017M620254), Zhejiang Postdoctoral Science Foundation (J.C.), and Foundation of Key Laboratory of Marine Ecosystem and Biogeochemistry, SOA (LMEB201501).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schillaci, D.; Spanò, V.; Parrino, B.; Carbone, A.; Montalbano, A.; Barraja, P.; Diana, P.; Cirrincione, G.; Cascioferro, S.M. Pharmaceutical approaches to target antibiotic resistance mechanisms. J. Med. Chem. 2017, 60, 8268–8297. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, C.; Parsek, M.R.; Greenberg, E.P. Regulation of gene expression by cell-to-cell communication: Acyl-homoserine lactone quorum sensing. Annu. Rev. Genet. 2001, 35, 439–468. [Google Scholar] [CrossRef] [PubMed]
- Breah, L.S.; Federle, M.J. Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar]
- Kaufmann, G.; Park, J.; Janda, K.D. Bacterial quorum sensing: A new target for anti-infective immunotherapy. Expert Opin. Biol. Ther. 2008, 8, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.A.; Hoven, A.D.; Cook, A.M. Therapeutic frontiers: Preventing and treating infectious diseases by inhibiting bacterial quorum sensing. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 27, 635–642. [Google Scholar] [CrossRef]
- Galloway, W.R.J.D.; Hodgkinson, J.T.; Bowden, S.D.; Welch, M.; Spring, D.R. Quorum sensing in Gram-negative bacteria: Small-molecule modulation of AHL and AI-2 quorum sensing pathways. Chem. Rev. 2011, 111, 28–67. [Google Scholar] [CrossRef] [PubMed]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A quorum-sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proc. Natl. Acad. Sci. USA 2013, 110, 17981–17986. [Google Scholar] [CrossRef]
- Lu, Y.; Dong, X.; Liu, S.; Bie, X. Characterization and identification of a novel marine Streptomyces sp. produced antibacterial substance. Mar. Biotechnol. 2009, 11, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.W.; Wu, Q.H.; Rowley, D.C.; Al-Kareef, A.M.; Wang, H. Anticancer agent-based marine natural products and related compounds. J. Asian Nat. Prod. Res. 2015, 17, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ashforth, E.; Ren, B.; Song, F.; Dai, H.; Liu, M.; Wang, J.; Xie, Q.; Zhang, L. Bioprospecting microbial natural product libraries from the marine environment for drug discovery. J. Antibiot. 2010, 63, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2016, 33, 382–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teasdale, M.E.; Donovan, K.A.; Forschner-Dancause, S.R.; Rowley, D.C. Gram-positive marine bacteria as a potential resource for the discovery of quorum sensing inhibitors. Mar. Biotechnol. 2011, 13, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Maskey, R.P.; Asolkar, R.N.; Kapaun, E.; Wagner-Dobler, I.; Laatsch, H. Phytotoxic arylethylamides from limnic bacteria using a screening with microalgae. J. Antibiot. 2002, 55, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Teasdale, M.E.; Liu, J.; Wallace, J.; Akhlaghi, F.; Rowley, D.C. Secondary metabolites produced by the marine bacterium Halobacillus salinus that inhibit quorum sensing-controlled phenotypes in Gram-negative bacteria. Appl. Environ. Microbiol. 2009, 75, 567–572. [Google Scholar] [CrossRef]
- Tommonaro, G.; Abbamondi, R.; Iodice, C.; Tait, K.; De Rosa, S. Diketopiperazines produced by the halophilic Archaeon, Haloterrigena hispanica, activate AHL bioreporters. Microb. Ecol. 2012, 63, 490–495. [Google Scholar] [CrossRef]
- Abed, R.M.M.; Dobretsov, S.; Al-Fori, M.; Gunasekera, S.P.; Sudesh, K.; Paul, V.J. Quorum-sensing inhibitory compounds from extremophilic microorganisms isolated from a hypersaline cyanobacterial mat. J. Ind. Microbiol. Biot. 2013, 40, 759–772. [Google Scholar] [CrossRef]
- Chen, X.C.; Chen, J.W.; Yan, Y.C.; Chen, S.; Xu, X.W.; Zhang, H.W.; Wang, H. Quorum sensing inhibitors from marine bacteria Oceanobacillus sp. XC22919. Nat. Prod. Res. 2018, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Saurav, K.; Bar-Shalom, R.; Haber, M.; Burgsdorf, I.; Oliviero, G.; Costantino, V.; Morgenstern, D.; Steindler, L. In search of alternative antibiotic drugs: Quorum-quenching activity in sponges and their bacterial isolates. Front. Microbiol. 2016, 7, 416. [Google Scholar] [CrossRef]
- Mansson, M.; Nielsen, A.; Kjærulff, L.; Gotfredsen, C.H.; Wietz, M.; Ingmer, H.; Gram, L.; Larsen, T.O. Inhibition of virulence gene expression in Staphylococcus aureus by novel depsipeptides from a marine Photobacterium. Mar. Drugs 2011, 9, 2537–2552. [Google Scholar] [CrossRef]
- Richard, P.N. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol. Microbiol. 2010, 48, 1429–1449. [Google Scholar]
- Ji, G.Y.; Beavis, R.; Novick, R.P. Bacterial interference caused by autoinducing peptide variants. Science 1997, 276, 2027–2030. [Google Scholar] [CrossRef] [PubMed]
- Queck, S.Y.; Jameson-Lee, M.; Villaruz, A.E.; Bach, T.H.L.; Khan, B.A.; Sturdevant, D.E.; Ricklefs, S.M.; Li, M.; Otto, M. RNAIII-independent target gene control by the agr quorum-sensing system: Insight into the evolution of virulence regulation in Staphylococcus aureus. Mol. Cell 2008, 32, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Anita, N.; Maria, M.N.; Bojer, M.S.; Lone, G.; Larsen, T.O.; Novick, R.P.; Dorte, F.; Hanne, F.K.R.; Hanne, I. Solonamide B inhibits quorum sensing and reduces Staphylococcus aureus mediated killing of human neutrophils. PLoS ONE 2014, 9, e84992. [Google Scholar] [CrossRef]
- Jarraud, S.; Lyon, G.J.; Figueiredo, A.M.; Lina, G.; Gérard, L.; Vandenesch, F.; Etienne, J.; Muir, T.W.; Novick, R.P. Exfoliatin-producing strains define a fourth agr specificity group in Staphylococcus aureus. J. Bacteriol. 2011, 182, 6517–6522. [Google Scholar] [CrossRef]
- Hansen, A.M.; Peng, P.; Baldry, M.; Perez-Gassol, I.; Christensen, S.B.; Vinther, J.M.O.; Ingmer, H.; Franzyk, H. Lactam hybrid analogues of solonamide B and autoinducing peptides as potent S. aureus AgrC antagonists. Eur. J. Med. Chem. 2018, 152, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Kjaerulff, L.; Nielsen, A.; Mansson, M.; Gram, L.; Larsen, T.O.; Ingmer, H.; Gotfredsen, C.H. Identification of four new agr quorum sensing-interfering cyclodepsipeptides from a marine Photobacterium. Mar. Drugs 2013, 11, 5051–5062. [Google Scholar] [CrossRef]
- Padmavathi, A.R.; Abinaya, B.; Pandian, S.K. Phenol, 2,4-bis(1,1-dimethylethyl) of marine bacterial origin inhibits quorum sensing mediated biofilm formation in the uropathogen Serratia marcescens. Biofouling 2014, 30, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Dai, X.; Sun, J.; Bu, X.; Weng, C.; Li, H.; Hu, Z. A diketopiperazine factor from Rheinheimera aquimaris QSI02 exhibits anti-quorum sensing activity. Sci. Rep. 2016, 6, 39637. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Su, Y.; Brackman, G.; Cui, F.Y.; Zhang, Y.H.; Shi, X.C.; Coenye, T.; Zhang, X.H. MomL, a novel marine-derived N-acyl homoserine lactonase from Muricauda olearia. Appl. Environ. Microbiol. 2015, 81, 774–782. [Google Scholar] [CrossRef]
- Saurav, K.; Costantino, V.; Venturi, V.; Steindler, L. Quorum sensing inhibitors from the sea discovered using bacterial N-acyl-homoserine lactone-based biosensors. Mar. Drugs 2017, 15, 53. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Liu, P.; Gong, Q.; Wang, Y.; Wang, P.; Zhu, W. α-Pyrones from the marine-derived actinomycete Nocardiopsis dassonvillei subsp. dassonvillei XG-8-1. RSC Adv. 2013, 3, 20726–20731. [Google Scholar] [CrossRef]
- Du, Y.; Sun, J.; Gong, Q.; Wang, Y.; Fu, P.; Zhu, W. New α-pyridones with quorum-sensing inhibitory activity from diversity-enhanced extracts of a Streptomyces sp. derived from marine algae. J. Agric. Food Chem. 2018, 66, 1807–1812. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.A.; Tam, W.H.J.; Krepinsky, J. The chemistry of some 6-methyl-4-hydroxy-2-pyridones. Can. J. Chem. 1978, 56, 613–616. [Google Scholar] [CrossRef] [Green Version]
- Abadi, A.; Al-Deeb, O.; Al-Afify, A.; El-Kashef, H. Synthesis of 4-alkyl(aryl)-6-aryl-3-cyano- 2(1H)-pyridinones and their 2-imino isosteres as nonsteroidal cardiotonic agents. Farmaco 1999, 54, 195–201. [Google Scholar] [CrossRef]
- Cocco, M.T.; Congiu, C.; Onnis, V. Synthesis and antitumour activity of 4-hydroxy-2-pyridone derivatives. Eur. J. Med. Chem. 2000, 35, 545–552. [Google Scholar] [CrossRef]
- Öztürk, G.; Erol, D.D.; Uzbay, T.; Aytemir, M.D. Synthesis of 4(1H)-pyridinone derivatives and investigation of analgesic and antiinflammatory activities. Farmaco 2001, 56, 251–256. [Google Scholar] [CrossRef]
- Cocco, M.T.; Congiu, C.; Onnis, V. New bis(pyridyl)methane derivatives from 4-hydroxy-2- pyridones: Synthesis and antitumoral activity. Eur. J. Med. Chem. 2003, 38, 37–47. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Böger, P. Phytotoxic sites of action for molecular design of modern herbicides (Part 2): Amino acid, lipid and cell wall biosynthesis, and other targets for future herbicides. Weed Biol. Manag. 2004, 4, 59–70. [Google Scholar] [CrossRef]
- Storck, P.; Aubertin, A.; Grierson, D.S. Tosylation/mesylation of 4-hydroxy-3-nitro-2-pyridinones as an activation step in the construction of dihydropyrido[3,4-b]benzo[f][1,4]thiazepin-1-one based anti-HIV agents. Tetrahedron Lett. 2005, 46, 2919–2922. [Google Scholar] [CrossRef]
- Evidente, A.; Fiore, M.; Bruno, G.; Sparapano, L.; Motta, A. Chemical and biological characterisation of sapinopyridione, a phytotoxic 3,3,6-trisubstituted-2,4-pyridione produced by Sphaeropsis sapinea, a toxigenic pathogen of native and exotic conifers, and its derivatives. Phytochemistry 2006, 67, 1019–1028. [Google Scholar] [CrossRef]
- Macdonald, G.E.; Puri, A.; Shilling, D.G. Interactive effect of photoperiod and fluridone on growth, reproduction, and biochemistry of dioecious hydrilla (Hydrilla Verticillata). Weed Sci. 2008, 56, 189–195. [Google Scholar] [CrossRef]
- Lopez, S.N.; Ramallo, I.A.; Sierra, M.G.; Zacchino, S.A.; Furlan, R.L.E. Chemically engineered extracts as an alternative source of bioactive natural product-like compounds. Proc. Natl. Acad. Sci. USA 2007, 104, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Ramallo, I.A.; Salazar, M.O.; Mendez, L.; Furlan, R.L.E. Chemically engineered extracts: Source of bioactive compounds. Acc. Chem. Res. 2011, 44, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, H.S.; Ok, K.; Kim, Y.H.; Park, H.D.; Byun, Y. Design, synthesis and biological evaluation of 4-(alkyloxy)-6-methyl-2H-pyran-2-one derivatives as quorum sensing inhibitors. Bioorg. Med. Chem. Lett. 2015, 25, 2913–2917. [Google Scholar] [CrossRef] [PubMed]
- Naik, D.N.; Wahidullah, S.; Meena, R.M. Attenuation of Pseudomonas aeruginosa virulence by marine invertebrate-derived Streptomyces sp. Lett. Appl. Microbiol. 2013, 56, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Rajkumari, J.; Borkotoky, S.; Murali, A.; Suchiang, K.; Mohanty, S.K.; Busi, S. Cinnamic acid attenuates quorum sensing associated virulence factors and biofilm formation in Pseudomonas aeruginosa PAO1. Biotechnol. Lett. 2018, 40, 1087–1100. [Google Scholar] [CrossRef] [PubMed]
- Thimmaraju, R.; Bais, H.P. Curcumin, a known phenolic from Curcuma longa, attenuates the virulence of Pseudomonas aeruginosa PAO1 in whole plant and animal pathogenicity models. J. Agric. Food Chem. 2008, 56, 1955–1962. [Google Scholar]
- Malheiro, J.F.; Maillard, J.Y.; Borges, F.; Simões, M. Evaluation of cinnamaldehyde and cinnamic acid derivatives in microbial growth control. Int. Biodeter. Biodegr. 2018. [Google Scholar] [CrossRef]
- Gilles, B.; Shari, C.; Ulrik, H.; Serge, V.C.; Paul, C.; Louis, M.; Hans, J.N.; Tom, C. Structure-activity relationship of cinnamaldehyde analogs as inhibitors of AI-2 based quorum sensing and their effect on virulence of Vibrio spp. PLoS ONE 2011, 6, e16084. [Google Scholar] [CrossRef]
- Joshi, J.R.; Burdman, S.; Lipsky, A.; Yariv, S.; Yedidia, I. Plant phenolic acids affect the virulence of Pectobacterium aroidearum and P. carotovorum ssp. brasiliense via quorum sensing regulation. Mol. Plant. Pathol. 2016, 17, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Xu, J.; Yao, Z.; Jiang, Y.; Zhou, H.; Jiang, W.; Dong, K. The anti-quorum sensing activity and bioactive substance of a marine derived Streptomyces. Biotechnol. Biotechnol. Equip. 2017, 31, 1007–1015. [Google Scholar] [CrossRef]
- Hasan, S.; Ansari, M.I.; Ahmad, A.; Mishra, M. Major bioactive metabolites from marine fungi: A Review. Bioinformation 2015, 11, 176–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Jeong, J.H.; Lee, K.T.; Rho, J.R.; Choi, H.D.; Kang, J.S.; Son, B.W. Gamma-pyrone derivatives, kojic acid methyl ethers from a marine-derived fungus Alternaria [correction of Altenaria] sp. Arch. Pharm. Res. 2003, 26, 532–534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, M.; Zhu, X.; Yu, W.; Gong, Q. Equisetin as potential quorum sensing inhibitor of Pseudomonas aeruginosa. Biotechnol. Lett. 2018, 40, 1–6. [Google Scholar] [CrossRef]
- Tang, Y.D.; Zhou, M.L.; Yu, W.G.; Gong, Q.H. Isolation and identification of quorum sensing inhibitors from a strain of marine fungus Penicillium sp. QF046. Prog. Mod. Biomed. 2014, 14, 6607–6621. [Google Scholar]
- Zheng, C.J.; Sohn, M.J.; Lee, S.; Kim, W.G. Meleagrin, a new FabI inhibitor from Penicillium chryosogenum with at least one additional mode of action. PLoS ONE 2013, 8, e78922. [Google Scholar] [CrossRef]
- Dobretsov, S.; Teplitski, M.; Bayer, M.; Gunasekera, S.; Proksch, P.; Paul, V.J. Inhibition of marine biofouling by bacterial quorum sensing inhibitors. Biofouling 2011, 27, 893–905. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.D.; Zhou, L.M.; Ma, Q.Y.; Huang, S.Z.; Wang, P.; Dai, H.F.; Zhao, Y.X. Metabolites with Gram-negative bacteria quorum sensing inhibitory activity from the marine animal endogenic fungus Penicillium sp. SCS-KFD08. Arch. Pharm. Res. 2016, 40, 25–31. [Google Scholar] [CrossRef]
- Martín-Rodríguez, A.J.; Reyes, F.; Martín, J.; Pérez-Yépez, J.; León-Barrios, M.; Couttolenc, A.; Espinoza, C.; Trigos, Á.; Martín, V.S.; Norte, M.; et al. Inhibition of bacterial quorum sensing by extracts from aquatic fungi: First report from marine endophytes. Mar. Drugs 2014, 12, 5503–5526. [Google Scholar]
- Van Rij, E.T.; Wesselink, M.; Chin-A.-Woeng, T.F.C.; Bloemberg, G.V.; Lugtenberg, B.J.J. Influence of environmental conditions on the production of phenazine-1-carboxamide by Pseudomonas chlororaphis PCL1391. Mol. Plant Microb. Interact. 2004, 17, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Van Rij, E.T.; Girard, G.; Lugtenberg, B.J.J.; Bloemberg, G.V. Influence of fusaric acid on phenazine-1-carboxamide synthesis and gene expression of Pseudomonas chlororaphis strain PCL1391. Microbiology 2005, 151, 2805–2814. [Google Scholar] [CrossRef] [PubMed]
- Widmer, K.W.; Soni, K.A.; Hume, M.E.; Beier, R.C.; Jesudhasan, P.; Pillai, S.D. Identification of poultry meat-derived fatty acids functioning as quorum sensing signal inhibitors to autoinducer-2 (AI-2). J. Food Sci. 2007, 72, M363–M368. [Google Scholar] [CrossRef] [PubMed]
- Soni, K.A.; Jesudhasan, P.; Cepeda, M.; Widmer, K.; Jayaprakasha, G.K.; Patil, B.S.; Hume, M.E.; Pillai, S.D. Identification of ground beef–derived fatty acid inhibitors of autoinducer-2-based cell signaling. J. Food Protect. 2008, 71, 134–138. [Google Scholar] [CrossRef]
- De Carvalho, M.P.; Abraham, W.R. Antimicrobial and biofilm inhibiting diketopiperazines. Curr. Med. Chem. 2012, 19, 3564–3577. [Google Scholar] [CrossRef] [PubMed]
- Tung, T.T.; Jakobsen, T.H.; Dao, T.T.; Fuglsang, A.T.; Givskov, M.; Christensen, S.B.; Nielsen, J. Fusaric acid and analogues as Gram-negative bacterial quorum sensing inhibitors. Eur. J. Med. Chem. 2016, 126, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Zhang, X.H. Quorum quenching agents: Resources for antivirulence therapy. Mar. Drugs 2014, 12, 3245–3282. [Google Scholar] [CrossRef]
- Ng, W.L.; Bassler, B.L. Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 2009, 43, 197–222. [Google Scholar] [CrossRef]
- Muhlen, S.; Dersch, P. Anti-virulence strategies to target bacterial infections. Curr. Top. Microbiol. 2015, 398, 147–183. [Google Scholar]
- Wagner, S.; Sommer, R.; Hinsberger, S.; Lu, C.; Hartmann, R.W.; Empting, M.; Titz, A. Novel strategies for the treatment of Pseudomonas aeruginosa infections. J. Med. Chem. 2016, 59, 5929–5969. [Google Scholar] [CrossRef]
- Hema, M.; Princy, S.A.; Sridharan, V.; Vinoth, P.; Balamurugan, P.; Sumana, M.N. Synergistic activity of quorum sensing inhibitor, pyrizine-2-carboxylic acid and antibiotics against multi-drug resistant V. cholerae. Rsc Adv. 2016, 6, 45938–45946. [Google Scholar] [CrossRef]
- Zhou, J.W.; Luo, H.Z.; Jiang, H.; Jian, T.K.; Chen, Z.Q.; Jia, A.Q. Hordenine, a novel quorum sensing inhibitor and anti-biofilm agent against Pseudomonas aeruginosa. J. Agric. Food Chem. 2018, 66, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Soukarieh, F.; Williams, P.; Stocks, M.J.; Cámara, M. Pseudomonas aeruginosa quorum sensing systems as drug discovery targets: Current position and future perspectives. J. Med. Chem. 2018, 61, 10385–10402. [Google Scholar] [CrossRef]
Figure 1.
The mechanisms of action of QSIs in gram-negative pathogens.

Figure 2.
Structure–activity relationships obtained lactam hybrid analogues with AgrC-inhibitory activity. Color coding stands for the effect of each of the structure features: red (high potency); yellow (low potency); blue (detrimental to potency).
Figure 2.
Structure–activity relationships obtained lactam hybrid analogues with AgrC-inhibitory activity. Color coding stands for the effect of each of the structure features: red (high potency); yellow (low potency); blue (detrimental to potency).

Figure 3.
Structure–activity relationships obtained for fusaric acid analogues.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
QSIs from marine microorganisms.
| No. | Structures of QSIs | Source | Biosensor Microorganisms | Specific Inhibitory Activity | Ref. |
|---|---|---|---|---|---|
| 1 |  | H. salinus C42 | V. harveyi BB120 | Inhibition of violacein of C. violaceum CV026 and green fluorescent protein of E. coli JB525 | [14,15] |
| 2 |  | ||||
| 3 |  | H. hispanica SK-3 | E. coli A. tumefaciens NTL4 | Inhibition of green fluorescent protein of V. anguillarum DM27 | [16] |
| 4 |  | Marinobacter sp. SK-3 | C. violaceum CV017 | Inhibition of luminescence of E. coli pSB401 | [17] |
| 5 |  | ||||
| 6 |  | ||||
| 7 |  | ||||
| 8 |  | Oceanobacillus sp.XC22919 | C. violaceum CV026 | Inhibition of violacein of C. violaceum ATCC12472 and pyocyanin, elastase and proteolytic activities, and biofilm formation of P. aeruginosa | [18] |
| 9 |  | ||||
| 10 |  | ||||
| 11 |  | Nautella sp. | C. violaceum CV026 | Inhibition of pyoyanin production of P. aeruginosa PAO1, and biofilm formation in E. coli, P. aeruginosa PAO1, and Bacillus subtilis | [19] |
| 12 |  | ||||
| 13 |  | ||||
| 14 |  | Erythrobacter sp. CUA-870 | C. violaceum CV026 | Inhibition of pyoyanin production of P. aeruginosa PAO1, and biofilm formation in E. coli, P. aeruginosa PAO1, and Bacillus subtilis | [19] |
| 15 |  | ||||
| 16 |  | ||||
| 17 |  | Dietzia maris IHBB 9296 | C. violaceum CV026 | Inhibition of pyoyanin production of P. aeruginosa PAO1, and biofilm formation in E. coli, P. aeruginosa PAO1, and Bacillus subtilis | [19] |
| 18 |  | ||||
| 19 |  | ||||
| 20 |  | Photobacterium | S. aureus 8325-4 | Inhibition of agrA and AgrA, and virulence factors α-hemolysin and Protein A in S. aureus | [20] |
| 21 |  | ||||
| 22: n = 1; R=H3C(CH2)5- 23: n = 1; R=H3C(CH2)8- 24: n = 2; R=H3C(CH2)5- |  | Chemical synthesis based on solonamide B and autoinducing peptides | P3-blaZ reporter strain | Inhibition of β-actamase activity controlled by S. aureus agr QS system | [26] |
| 25: R=H3C(CH2)5-; R1=H3C- 26: R=H3C(CH2)4-; R1=H3C- 27: R=H3C(CH2)6-; R1=H3C- 28: R=H3C(CH2)10-; R1=H3C- 29: R=H3C(CH2)5-; R1=H 30: R=H3C(CH2)5-; R1=(H3C)2CHCH2- |  | ||||
| 31: R=H3C-; R1=(H3C)2CHCH2- 32: R=(H3C)2CHCH2-; R1=H3C- 33: R=PhCH2-; R1=(H3C)2CHCH2- 34: R=(H3C)2CHCH2CH2- R1=(H3C)2CHCH2- 35: R=(H3C)2CHCH2- R1=(H3C)2CHCH2CH2- |  | ||||
| 36 |  | ||||
| 37 |  | ||||
| 38 |  | ||||
| 39 |  | ||||
40: R=  41: R=  |  | ||||
| 42: R=CH3(CH2)5CH=CH(CH2)3- 43: R=CH3(CH2)10- 44: R=CH3(CH2)5CH=CH(CH2)5- 45: R=CH3(CH2)12- |  | P. halotolerans | S. aureus lacZ reporter | Inhibition of expression of rnaIII in community-associated methicillin-resistant S. aureus USA300 | [27] |
| 46 |  | V. alginolyticus G16 | S. marcescens | Inhibition of biofilm, protease, haemolysin, lipase, prodigiosin and extracellular polysaccharide secretion in S. marcescens | [28] |
| 47 |  | R. aquimaris QSI02 | C. violaceum CV026 | Inhibition of pyocyanin production, elastase activity and biofilm formation in P. aeruginosa PA01 | [29] |
| 48 | N-acylhomoserine lactonase | Tenacibaculum sp. 20J | C. violaceum CV026 | Inhibition of SdiA in E. coli | [30] |
| 49 |  | Nocardiopsis dassonvillei subsp. dassonvillei XG-8-1 | C. violaceum CV026 and P. aeruginosa QSIS-lasI biosensors | Inhibition of C. violaceum CV026 and P. aeruginosa QSIS-lasI biosensors | [32] |
| 50 |  | ||||
| 51 |  | ||||
| 52 |  | Streptomyces sp. OUCMDZ-3436 | P. aeruginosa QSIS-lasI biosensors | Inhibition of gene expression controlled by QS in P. aeruginosa QSIS-lasI biosensors | [33] |
| 53 |  | ||||
| 54 |  | ||||
| 55 |  | ||||
| 56: n = 3; 57: n = 4; 58: n = 5; 59: n = 6; 60: n = 7; 61: n = 8; 62: n = 9; 63: n = 10; 64: n = 11; 65: n = 12 |  | Chemical synthesis based on pyrones | P. aeruginosa | Inhibition of biofilm formation of P. aeruginosa | [45] |
| 66 |  | Streptomyces sp. | C. violaceum ATCC 12472and P. aeruginosa ATCC 27853 | Inhibition of swarming, pyocyanin, biofilm formation, rhamnolipid production in P. aeruginosa | [46] |
| 67 |  | ||||
| 68 |  | Chemical synthesis based on cinnamic acid | C. violaceum | Inhibition of violacein of C. violaceum | [49] |
| 69 |  | ||||
| 70 |  | ||||
| 71 |  | ||||
| 72 |  | ||||
| 73 |  | ||||
| 74 |  | ||||
| 75 |  | Chemical synthesis based on cinnamic acid | Vibrio spp | Inhibition of biofilm formation, pigment production and protease production in Vibrio spp. | [50] |
| 76 |  | ||||
| 77 |  | ||||
| 78: R1=H; R2=C2H5- 79: R1=H; R2=C10H21- 80: R1=CH3O-; R2=C6H13- 81: R1=H; R2=C4H9- |  | ||||
| 82 |  | S. parvulus HY026 | C. violaceum ATCC 12472 | Inhibition of violaceim of C. violaceum and prodigiosin production of Serratia proteamaculans 657 | [52] |
| 83 |  | Altenaria sp. | E. coli pSB401 | Inhibition of luminescence of E. coli pSB401 | [54] |
| 84 |  | Fusarium sp. Z10 | P. aeruginosa QSIS- lasI biosensor | Inhibition of formation of biofilm, swarming motility, and the production of virulence factors in P. aeruginosa. | [55] |
| 85 |  | Penicillium sp. QF046 | C. violaceum CV026 | Inhibition of violacein of C. violaceum CV026 | [56] |
| 86 |  | P. chrysogenium | C. violaceum CV017 | Inhibition of S. aureus FabI | [57,58] |
| 87 |  | Penicillium sp. SCS-KFD08 | C. violaceum CV026 | Inhibition of violacein of C. violaceum CV026 | [59] |
| 88 |  | ||||
| 89 |  | ||||
| 90 |  | ||||
| 91 |  | ||||
| 92 |  | ||||
| 93 |  | Sarocladium | C. violaceum CV026 | Inhibition of violacein of C. violaceum CV026 | [60] |
| 94 |  | ||||
| 95 |  | ||||
| 96 |  | Fusarium | C. violaceum CV026 | Inhibition of violacein of C. violaceum CV026 | [60] |
| 97 |  | ||||
| 98 |  | ||||
| 99: R=CH3- 100: R=H |  | ||||
| 101 |  | ||||
| 102 |  | Epicoccum | C. violaceum CV026 | Inhibition of violacein of C. violaceum CV026 | [60] |
| 103 |  | ||||
| 104 |  | ||||
| 105 |  | ||||
| 106 |  | Khuskia | C. violaceum CV026 | Inhibition of violacein of C. violaceum CV026 | [60] |
| 107 |  | ||||
| 108 |  | ||||
109: R1=  ; R2=CH3O- ; R2=CH3O-110: R1=  ; R2=CH3CH2O- ; R2=CH3CH2O-111: R1=CH3CH2CH2-; R2=CH3O- 112: R1=  ; R2=CH3O- ; R2=CH3O-113: R1=  ; R2=CH3O- ; R2=CH3O-114: R1=CH3CH2CH2CH2- R2=CH3CH2O- 115: R1=CH3CH2CH2CH2-; R2=HO- 116: R1=Br-; R2=CH3O- |  | Chemical synthesis based on fusaric acid | P. aeruginosa and Vibrio fischeri | Inhibition of the las and rhl QS system in P. aeruginosa and the lux QS system in Vibrio fischeri | [66] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, J.; Wang, B.; Lu, Y.; Guo, Y.; Sun, J.; Wei, B.; Zhang, H.; Wang, H. Quorum Sensing Inhibitors from Marine Microorganisms and Their Synthetic Derivatives. Mar. Drugs 2019, 17, 80. https://doi.org/10.3390/md17020080
AMA Style
Chen J, Wang B, Lu Y, Guo Y, Sun J, Wei B, Zhang H, Wang H. Quorum Sensing Inhibitors from Marine Microorganisms and Their Synthetic Derivatives. Marine Drugs. 2019; 17(2):80. https://doi.org/10.3390/md17020080
Chicago/Turabian StyleChen, Jianwei, Bixia Wang, Yaojia Lu, Yuqi Guo, Jiadong Sun, Bin Wei, Huawei Zhang, and Hong Wang. 2019. "Quorum Sensing Inhibitors from Marine Microorganisms and Their Synthetic Derivatives" Marine Drugs 17, no. 2: 80. https://doi.org/10.3390/md17020080
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.