Cyclopropane-Containing Fatty Acids from the Marine Bacterium Labrenzia sp. 011 with Antimicrobial and GPR84 Activity

 , , and
, , and
Abstract
:1. Introduction
2. Results
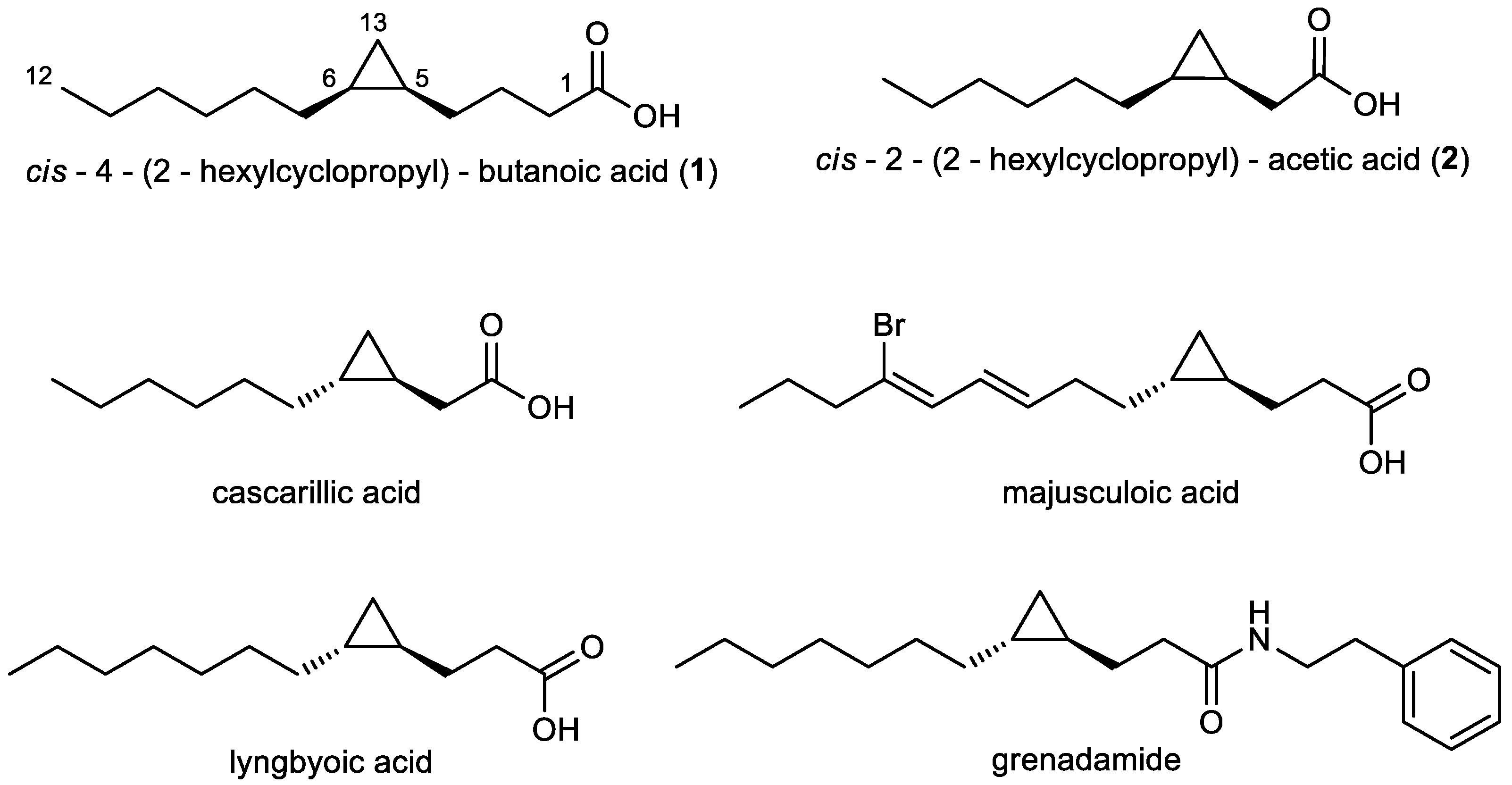
2.1. Isolation and Structural Elucidation of Compounds 1 and 2
2.2. Antimicrobial Assessment of 1 and 2
2.3. Effects of Compounds 1 and 2 on the Orphan G-Protein Coupled Receptor GPR84
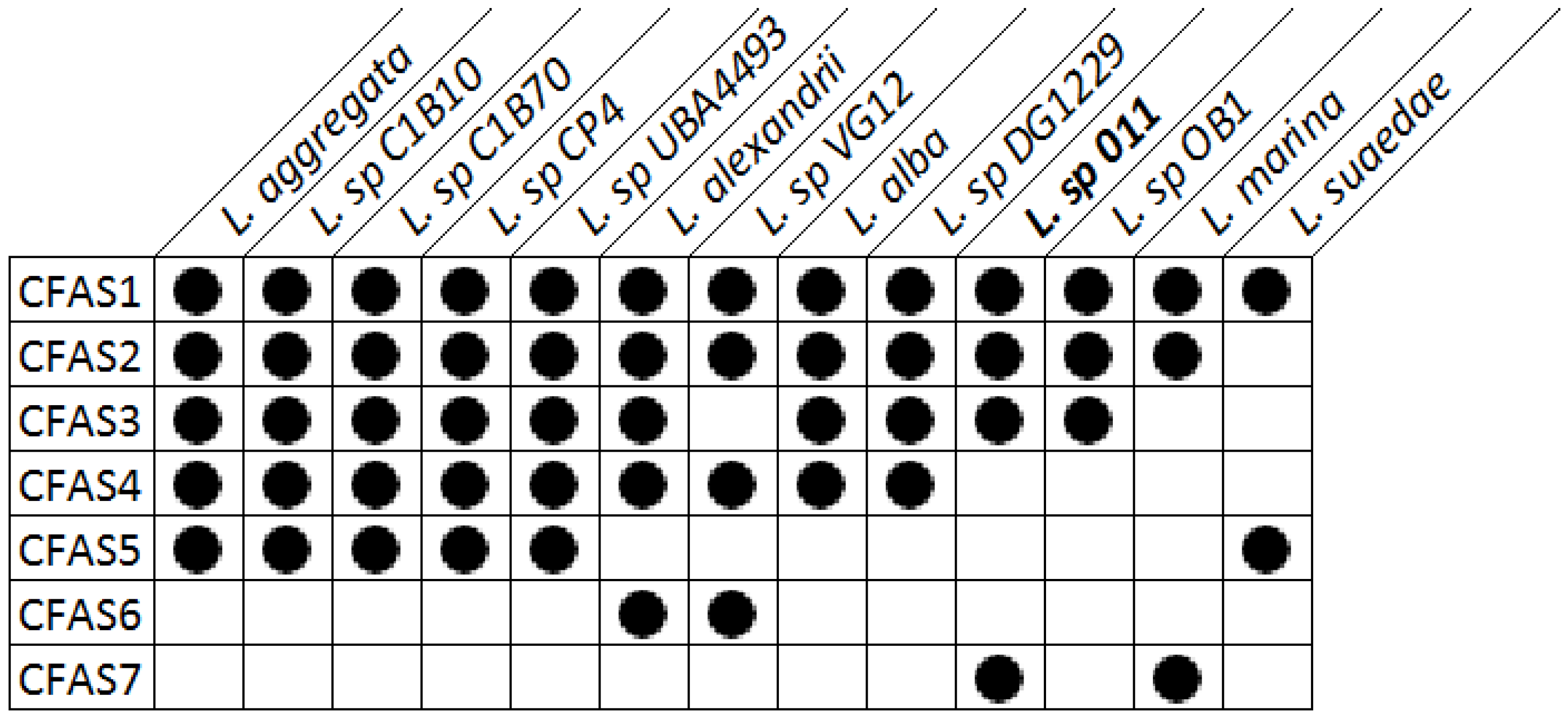
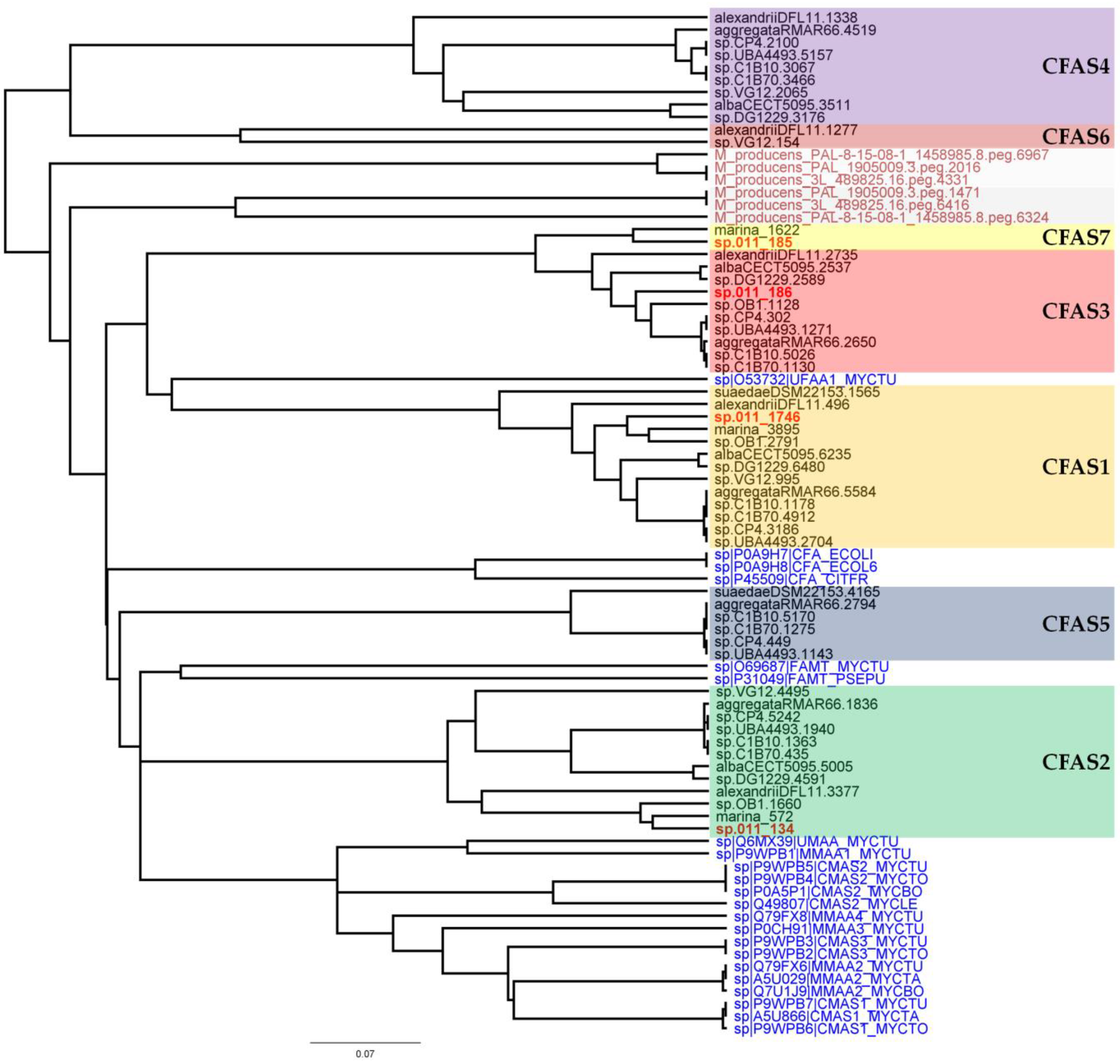
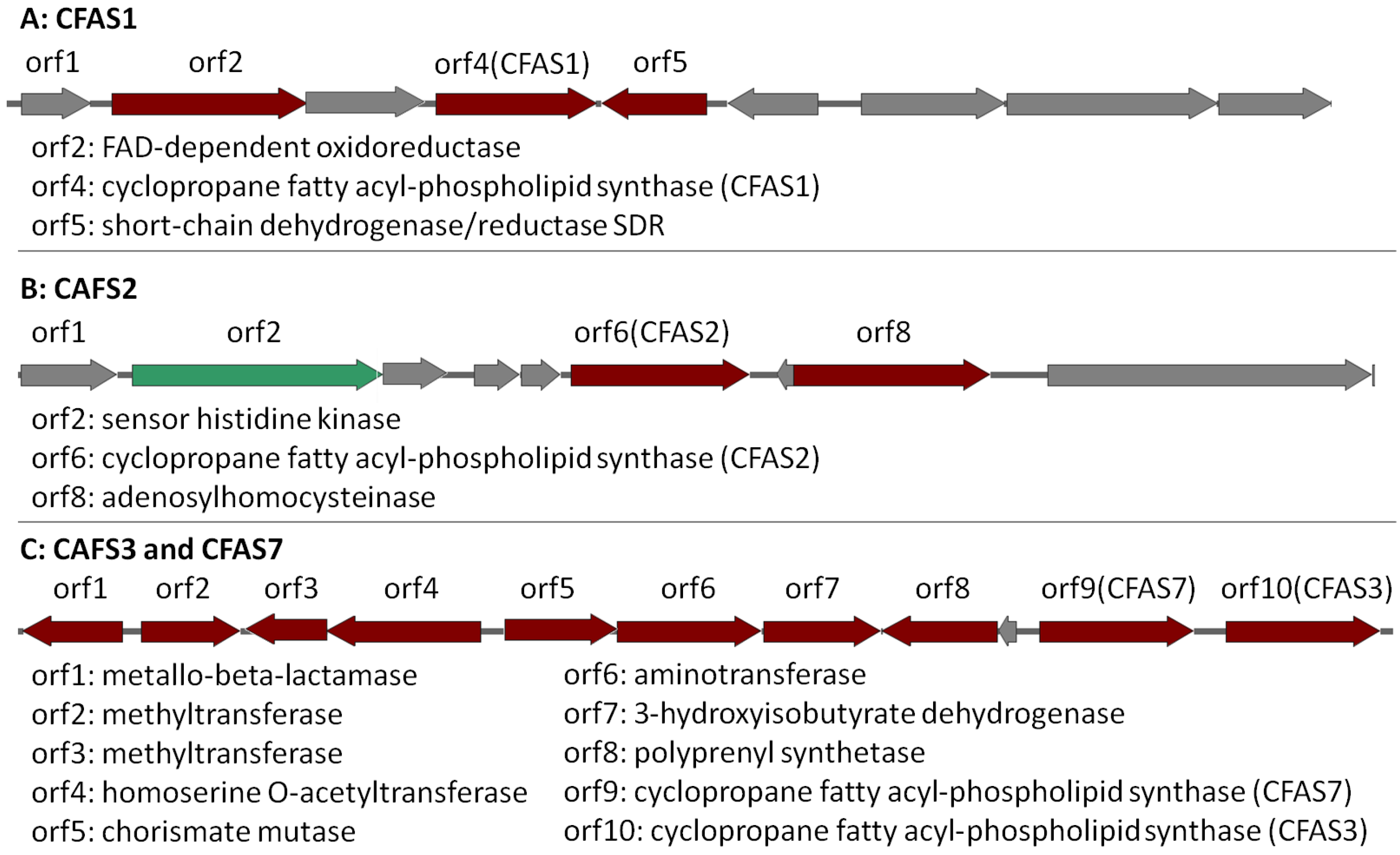
2.4. Bioinformatic Analysis of the Genome of Labrenzia sp. 011 for the Identification of the Putative Biosynthetic Genes of 1 and 2
3. Discussion
4. Experimental Section
4.1. General Procedures
4.2. Isolation and Taxonomy of the Bacterial Strain
4.3. Cultivation and Isolation Protocol
4.4. Genome Sequencing and Assembly
4.5. Prediction of CFAS Coding Regions
4.6. Genome Comparison and Phylogeny of CFAS
4.7. Antimicrobial Assays
4.8. Biological Assays at the GPR84 Receptor
4.9. cAMP Accumulation Assays
4.10. β-Arrestin Recruitment Assays
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Zinger, L.; Amaral-Zettler, L.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Huse, S.M.; Welch, D.B.M.; Martiny, J.B.H.; Sogin, M.; Boetius, A.; Ramette, A. Global patterns of bacterial beta-diversity in seafloor and seawater ecosystems. PLoS ONE 2011, 6, e24570. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2004, 21, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.R.; Fenical, W. Strategies for the discovery of secondary metabolites from marine bacteria: Ecological perspectives. Annu. Rev. Microbiol. 1994, 48, 559–584. [Google Scholar] [CrossRef] [PubMed]
- Boyd, K.G.; Adams, D.R.; Burgess, J.G. Antibacterial and repellent activities of marine bacteria associated with algal surfaces. Biofouling 1999, 14, 227–236. [Google Scholar] [CrossRef]
- Ganapiriya, V.; Maharajan, A.; Kumarasamy, P. Antifouling effect of bioactive compounds from marine sponge Acanthella elongata and different species of bacterial film on larval attachment of Balanus amphitrite (cirripedia, crustacea). Braz. Arch. Biol. Technol. 2012, 55, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Clare, A. Marine natural product antifoulants: Status and potential. Biofouling 1996, 9, 211–229. [Google Scholar] [CrossRef]
- Paul, V.J.; Puglisi, M.P. Chemical mediation of interactions among marine organisms. Nat. Prod. Rep. 2004, 21, 189–209. [Google Scholar] [CrossRef] [PubMed]
- Gil-Turnes, M.S.; Hay, M.E.; Fenical, W. Symbiotic marine bacteria chemically defend crustacean embryos from a pathogenic fungus. Science (New York, N.Y.) 1989, 246, 116–118. [Google Scholar] [CrossRef]
- Gil-Turnes, M.S.; Fenical, W. Embryos of Homarus americanus are Protected by Epibiotic Bacteria. Biol. Bull. 1992, 182, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Arfken, A.; Song, B.; Bowman, J.S.; Piehler, M. Denitrification potential of the eastern oyster microbiome using a 16S rRNA gene based metabolic inference approach. PLoS ONE 2017, 12, e0185071. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Li, T.; Chen, M.; Huang, G. Cross-ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl. Environ. Microbiol. 2008, 74, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Keats, K.F.; Rivkin, R.B.; Lang, A.S. Water mass and depth determine the distribution and diversity of Rhodobacterales in an Arctic marine system. FEMS Microbiol. Ecol. 2013, 84, 564–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pujalte, M.J.; Lucena, T.; Ruvira, M.A.; Arahal, D.R.; Macián, M.C. The Family Rhodobacteraceae. In Alphaproteobacteria and Betaproteobacteria: With 263 Tables, 4th ed.; Rosenberg, E., Ed.; Springer Reference: Berlin, Germany, 2014; pp. 439–512. [Google Scholar]
- Biebl, H.; Pukall, R.; Lunsdorf, H.; Schulz, S.; Allgaier, M.; Tindall, B.J.; Wagner-Dobler, I. Description of Labrenzia alexandrii gen. nov., sp. nov., a novel alphaproteobacterium containing bacteriochlorophyll a, and a proposal for reclassification of Stappia aggregata as Labrenzia aggregata comb. nov., of Stappia marina as Labrenzia marina comb. nov. and of Stappia alba as Labrenzia alba comb. nov., and emended descriptions of the genera Pannonibacter, Stappia and Roseibium, and of the species Roseibium denhamense and Roseibium hamelinense. Int. J. Syst. Evol. Microbiol. 2007, 57, 1095–1107. [Google Scholar]
- Boettcher, K.J.; Barber, B.J.; Singer, J.T. Additional Evidence that Juvenile Oyster Disease Is Caused by a Member of the Roseobacter Group and Colonization of Nonaffected Animals by Stappia stellulata-Like Strains. Appl. Environ. Microbiol. 2000, 66, 3924–3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloy, A.P.; Ford, S.E.; Karney, R.C.; Boettcher, K.J. Roseovarius crassostreae, the etiological agent of Juvenile Oyster Disease (now to be known as Roseovarius Oyster Disease) in Crassostrea virginica. Aquaculture 2007, 269, 71–83. [Google Scholar] [CrossRef]
- Pujalte, M.J.; Carmen Macián, M.; Arahal, D.R.; Garay, E. Stappia alba sp. nov., isolated from Mediterranean oysters. Sys. Appl. Microbiol. 2005, 28, 672–678. [Google Scholar] [CrossRef]
- Romalde, J.L.; Barja, J.L. Bacteria in molluscs: Good and bad guys. In Current Research, Technology and Education Topics In Applied Microbiology And Microbial Biotechnology; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2010; pp. 136–147. ISBN 978-84-614-6194-3. [Google Scholar]
- Schleissner, C.; Cañedo, L.M.; Rodríguez, P.; Crespo, C.; Zúñiga, P.; Peñalver, A.; de La Calle, F.; Cuevas, C. Bacterial Production of a Pederin Analogue by a Free-Living Marine Alphaproteobacterium. J. Nat. Prod. 2017, 80, 2170–2173. [Google Scholar] [CrossRef] [PubMed]
- Carballeira, N.M. New advances in fatty acids as antimalarial, antimycobacterial and antifungal agents. Prog. Lipid Res. 2008, 47, 50–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Han, R.; Wang, Y.; Li, X.; Zhang, M.; Yan, Y. Fungicidal Activity of a Medium-chain Fatty Acids Mixture Comprising Caprylic, Pelargonic and Capric Acids. Plant Pathol. J. 2014, 13, 65–70. [Google Scholar] [CrossRef]
- Pohl, C.; Kock, J.; Thibane, V. Antifungal free fatty acids: A review. In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2011; pp. 61–71. [Google Scholar]
- Macmillan, J.B.; Molinski, T.F. Majusculoic acid, a brominated cyclopropyl fatty acid from a marine cyanobacterial mat assemblage. J. Nat. Prod. 2005, 68, 604–606. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Köse, M.; Namasivayam, V.; Sylvester, K.; Borges, G.; Thimm, D.; von Kügelgen, I.; Müller, C.E. 6-(Ar)Alkylamino-Substituted Uracil Derivatives: Lipid Mimetics with Potent Activity at the Orphan G Protein-Coupled Receptor 84 (GPR84). ACS Omega 2018, 3, 3365–3383. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Curto, E.; Milligan, G. Metabolism meets immunity: The role of free fatty acid receptors in the immune system. Biochem. Pharmacol. 2016, 114, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Köse, M.; Sylvester, K.; Weighardt, H.; Thimm, D.; Borges, G.; Förster, I.; von Kügelgen, I.; Müller, C.E. Diindolylmethane Derivatives: Potent Agonists of the Immunostimulatory Orphan G Protein-Coupled Receptor GPR84. J. Med. Chem. 2017, 60, 3636–3655. [Google Scholar] [CrossRef] [PubMed]
- Roberts, I.O.; Baird, M.S.; Liu, Y. The absolute stereochemistry of cascarillic acid. Tetrahedron Lett. 2004, 45, 8685–8686. [Google Scholar] [CrossRef]
- Kwan, J.C.; Meickle, T.; Ladwa, D.; Teplitski, M.; Paul, V.; Luesch, H. Lyngbyoic acid, a “tagged” fatty acid from a marine cyanobacterium, disrupts quorum sensing in Pseudomonas aeruginosa. Mol. Biosyst. 2011, 7, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Sitachitta, N.; Gerwick, W.H. Grenadadiene and grenadamide, cyclopropyl-containing fatty acid metabolites from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 1998, 61, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Knothe, G. NMR characterization of dihydrosterculic acid and its methyl ester. Lipids 2006, 41, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, C.; Page, J.; Bedard, A.; Tremblay, P.; Vallieres, L. G protein-coupled receptor 84, a microglia-associated protein expressed in neuroinflammatory conditions. Glia 2007, 55, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, X.; Simonavicius, N.; Tian, H.; Ling, L. Medium-chain fatty acids as ligands for orphan G protein-coupled receptor GPR84. J. Biol. Chem. 2006, 281, 34457–34464. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, C.; Kuo, F. The G-protein coupled receptor, GPR84 regulates IL-4 production by T lymphocytes in response to CD3 crosslinking. Immunol. Lett. 2005, 101, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Takaishi, S.; Nagasaki, M.; Onozawa, Y.; Iino, I.; Maeda, H.; Komai, T.; Oda, T. Medium-chain fatty acid-sensing receptor, GPR84, is a proinflammatory receptor. J. Biol. Chem. 2013, 288, 10684–10691. [Google Scholar] [CrossRef] [PubMed]
- Amiri Moghaddam, J.; Dávila-Céspedes, A.; Alanjary, M.; Blom, J.; König, G.M.; Schäberle, T.F. Draft Genome Sequence of the Marine Bacterium Labrenzia sp. 011, a Potential Protective Agent of Mollusks. Data in Brief (in press)
- Courtois, F.; Guérard, C.; Thomas, X.; Ploux, O. Escherichia coli cyclopropane fatty acid synthase. Eur. J. Biochem. 2004, 271, 4769–4778. [Google Scholar] [CrossRef] [PubMed]
- Guangqi, E.; Lesage, D.; Ploux, O. Insight into the reaction mechanism of the Escherichia coli cyclopropane fatty acid synthase: Isotope exchange and kinetic isotope effects. Biochimie 2010, 92, 1454–1457. [Google Scholar] [PubMed]
- Engene, N.; Rottacker, E.C.; Kaštovský, J.; Byrum, T.; Choi, H.; Ellisman, M.H.; Komárek, J.; Gerwick, W.H. Moorea producens gen. nov., sp. nov. and Moorea bouillonii comb. nov., tropical marine cyanobacteria rich in bioactive secondary metabolites. Int. J. Syst. Evol. Microbiol. 2012, 62, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Machida, S.; Bakku, R.K.; Suzuki, I. Expression of Genes for a Flavin Adenine Dinucleotide-Binding Oxidoreductase and a Methyltransferase from Mycobacterium chlorophenolicum Is Necessary for Biosynthesis of 10-Methyl Stearic Acid from Oleic Acid in Escherichia coli. Front. Microbiol. 2017, 8, 2061. [Google Scholar] [CrossRef] [PubMed]
- Barkan, D.; Rao, V.; Sukenick, G.D.; Glickman, M.S. Redundant function of cmaA2 and mmaA2 in Mycobacterium tuberculosis cis cyclopropanation of oxygenated mycolates. J. Bacteriol. 2010, 192, 3661–3668. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, P.; Casadevall, A. Quorum sensing in fungi—A review. Med. Mycol. 2012, 50, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Boettcher, K.J.; Barber, B.J.; Singer, J.T. Additional evidence that juvenile oyster disease is caused by a member of the Roseobacter group and colonization of nonaffected animals by Stappia stellulata-like strains. Appl. Environ. Microbiol. 2000, 66, 3924–3930. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-H.; Kuo, J.; Sung, P.-J.; Chang, Y.-C.; Lu, M.-C.; Wong, T.-Y.; Liu, J.-K.; Weng, C.-F.; Twan, W.-H.; Kuo, F.-W. Isolation of marine bacteria with antimicrobial activities from cultured and field-collected soft corals. World J. Microbiol. Biotechnol. 2012, 28, 3269–3279. [Google Scholar] [CrossRef] [PubMed]
- Graca, A.P.; Bondoso, J.; Gaspar, H.; Xavier, J.R.; Monteiro, M.C.; de La Cruz, M.; Oves-Costales, D.; Vicente, F.; Lage, O.M. Antimicrobial activity of heterotrophic bacterial communities from the marine sponge Erylus discophorus (Astrophorida, Geodiidae). PLoS ONE 2013, 8, e78992. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Timms-Wilson, T.M.; Bailey, M.J. Identification of conserved traits in fluorescent pseudomonads with antifungal activity. Environ. Microbiol. 2000, 2, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Poger, D.; Mark, A.E. A ring to rule them all: The effect of cyclopropane Fatty acids on the fluidity of lipid bilayers. J. Phys. Chem. B 2015, 119, 5487–5495. [Google Scholar] [CrossRef] [PubMed]
- Motl, O.; Amin, M.; Sedmera, P. The structure of cascarillic acid from cascarilla essential oil. Phytochemistry 1972, 11, 407–408. [Google Scholar] [CrossRef]
- Wilson, S.R.; Prodan, K.A. The synthesis and stereochemistry of cascarillic acid. Tetrahedron Lett. 1976, 17, 4231–4234. [Google Scholar] [CrossRef]
- To, T.M.H.; Grandvalet, C.; Tourdot-Maréchal, R. Cyclopropanation of membrane unsaturated fatty acids is not essential to the acid stress response of Lactococcus lactis subsp. cremoris. Appl. Environ. Microbiol. 2011, 77, 3327–3334. [Google Scholar] [CrossRef] [PubMed]
- Velly, H.; Bouix, M.; Passot, S.; Penicaud, C.; Beinsteiner, H.; Ghorbal, S.; Lieben, P.; Fonseca, F. Cyclopropanation of unsaturated fatty acids and membrane rigidification improve the freeze-drying resistance of Lactococcus lactis subsp. lactis TOMSC161. Appl. Environ. Microbiol. 2015, 99, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Mukherjee, S.; Ning, Y.; Hsu, F.-F.; Zhang, K. Cyclopropane fatty acid synthesis affects cell shape and acid resistance in Leishmania mexicana. Int. J. Parasitol. 2018, 48, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Grogan, D.W.; Cronan, J.E. Cyclopropane ring formation in membrane lipids of bacteria. Microbiol. Mol. Biol. Rev. 1997, 61, 429–441. [Google Scholar] [PubMed]
- Banerjee, R.; Vats, P.; Dahale, S.; Kasibhatla, S.M.; Joshi, R. Comparative genomics of cell envelope components in mycobacteria. PLoS ONE 2011, 6, e19280. [Google Scholar] [CrossRef] [PubMed]
- Harms, H.; Poehlein, A.; Thürmer, A.; König, G.M.; Schäberle, T.F. Draft Genome Sequence of Zobellia sp. Strain OII3, Isolated from the Coastal Zone of the Baltic Sea. Genome Announ. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genomics 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; de Los Santos, E.L.C.; Kim, H.U.; Nave, M.; et al. antiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 45, W36–W41. [Google Scholar] [CrossRef] [PubMed]
- Blom, J.; Kreis, J.; Spänig, S.; Juhre, T.; Bertelli, C.; Ernst, C.; Goesmann, A. EDGAR 2.0: An enhanced software platform for comparative gene content analyses. Nucleic Acids Res. 2016, 44, W22–W28. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Takano, E.; Breitling, R. Detecting sequence homology at the gene cluster level with MultiGeneBlast. Mol. Biol. Evol. 2013, 30, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Schäberle, T.F.; Goralski, E.; Neu, E.; Erol, O.; Hölzl, G.; Dörmann, P.; Bierbaum, G.; König, G.M. Marine myxobacteria as a source of antibiotics—Comparison of physiology, polyketide-type genes and antibiotic production of three new isolates of Enhygromyxa salina. Mar. Drugs 2010, 8, 2466–2479. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Antimicrobial Activity | ||
|---|---|---|---|
| Bacteria | Compound 1 | Compound 2 | Streptomycin (+) |
| Escherichia coli DSM 498 | 2 | 3 | 6 |
| Bacillus megaterium DSM 32 | NA | 10 | 9 |
| Pseudoroseovarius crassostreae DSM 16950T | 5 | 5 | 6 |
| *Escherichia coli I-11276b | NT | 2 | NT |
| *MRSA LT-1338 | NT | 2 | NT |
| *MRSA LT-1334 | NT | 3 | NT |
| Fungi | Compound 1 | Compound 2 | Miconazole (+) |
| Eurotium rubrum DSM 62631 | NA | 10 | 8 |
| Mycotypha microspora MB#271115 | 3 | 6 | 9 |
| Microbotryum violaceum MB#110229 | 8 | 10 | 6 |
| Compound | cAMP Assay a | β-Arrestin Assay | ||
|---|---|---|---|---|
| EC50 ± SD (μM) (Receptor Activation) b n = 3 | Efficacy c | EC50 ± SD (μM) (Receptor Activation) d n = 5 | Efficacy e | |
| Decanoic acid | 7.42 ± 0.80 | 100% [26] | 6.08 ± 0.63 | 92% [26] |
| Dodecanoic acid | 8.87 ± 4.46 | 94% | 2.84 ± 0.54 | 95% |
| 1 | >>100 (0%) b | - | >30 (17%) d | - |
| 2 | >100 (24%) b | - | 0.114 ± 0.135 | 54% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amiri Moghaddam, J.; Dávila-Céspedes, A.; Kehraus, S.; Crüsemann, M.; Köse, M.; Müller, C.E.; König, G.M. Cyclopropane-Containing Fatty Acids from the Marine Bacterium Labrenzia sp. 011 with Antimicrobial and GPR84 Activity. Mar. Drugs 2018, 16, 369. https://doi.org/10.3390/md16100369
Amiri Moghaddam J, Dávila-Céspedes A, Kehraus S, Crüsemann M, Köse M, Müller CE, König GM. Cyclopropane-Containing Fatty Acids from the Marine Bacterium Labrenzia sp. 011 with Antimicrobial and GPR84 Activity. Marine Drugs. 2018; 16(10):369. https://doi.org/10.3390/md16100369
Chicago/Turabian StyleAmiri Moghaddam, Jamshid, Antonio Dávila-Céspedes, Stefan Kehraus, Max Crüsemann, Meryem Köse, Christa E. Müller, and Gabriele Maria König. 2018. "Cyclopropane-Containing Fatty Acids from the Marine Bacterium Labrenzia sp. 011 with Antimicrobial and GPR84 Activity" Marine Drugs 16, no. 10: 369. https://doi.org/10.3390/md16100369