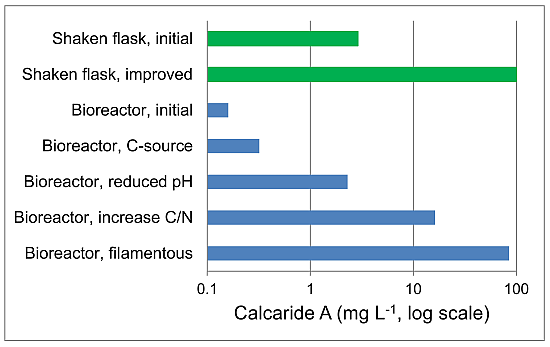
KF525 was cultivated in casamino acid glucose medium (pH 6.8) with low agitation, as previously reported [
10]. Calcaride A was extracted from the mycelium (1.1 mg·g
−1, equivalent to 2.9 ± 0.2 mg·L
−1), but essentially no calcaride A was found in the culture supernatant (0.04 ± 0.04 mg·L
−1) after incubation for 21 days, even though Silber
et al. [
10] reported purification of 17.7 mg from 12 L culture supernatant (1.5 mg·L
−1) after 24 days growth in the same medium. Biomass production in the flasks was low (5.2 ± 1.4 g·L
−1 after 10 days of incubation), limiting the amount of calcaride A produced. Biomass production was improved by cultivating KF525 in a bioreactor (7.1 g·L
−1 after 11 days) in the same medium (pH 6.8), but production of calcaride A was considerably reduced (0.02 mg·g
−1 biomass, equivalent to 0.15 mg·L
−1). No calcaride A was detected in the culture supernatant of the bioreactor culture and the related compounds were produced in proportionally small amounts, based on the UPLC chromatograms. We were thus interested to determine under which conditions calcaride A could be produced and whether it would be feasible to produce calcaride A in the bioreactor.
2.1. Calcaride A Production Is Affected by the Carbon Source
Casamino acids provide both carbon and nitrogen source. To assess whether the carbon source affected calcaride A production, KF525 was grown in defined medium with glucose, sucrose, fructose, maltose, lactose, malt extract or starch as the carbon source, with ammonium as the nitrogen source. KF525 grew on all carbon sources, producing the most biomass from starch (10.2 g·L
−1) and the least from lactose (4.4 ± 0.1 g·L
−1Table 1) after 25 days incubation.
Specific calcaride A production from glucose in defined medium (0.7 mg·g
−1) was similar to that observed in casamino acid medium (1.1 mg·g
−1), but the volumetric production (4.7 ± 0.4 mg·L
−1) was higher because the biomass concentration (6.3 ± 0.4 mg·L
−1) remained high in the culture (
Table 1). Calcaride A production was slightly higher with sucrose (0.9 ± 0.1 mg·g
−1, 7.2 ± 0.9 mg·L
−1) or fructose (1.5 ± 0.1 mg·g
−1, 8.1 ± 0.3 mg·L
−1) as the carbon source than with glucose and was significantly (
p < 0.05) reduced with other carbon sources (
Table 1). A similar trend of slightly higher calcaride A production from sucrose than from glucose was also observed in casamino acid medium (data not shown) and sucrose was provided as the carbon source for further studies.
The calcarides are related to the 15G256 compounds from
Hypoxylon oceanicum, another marine fungus, described by Schlingmann
et al. [
17], who suggested that the precursors were 6-hydroxymellein and β-hydroxybutyric acid, presumably via malonyl-CoA. 6-Methoxymellein would also be needed for the synthesis of the calcarides [
10]. Mellein, hydroxymelleins and methoxymelleins are produced by a variety of fungi (e.g., [
18]). Interestingly, sucrose has been shown to be a good substrate for production of mellein by the fungus
Aspergillus ochraceous, although maltose and starch were found to be equally good or better substrates for producing 4-hydroxymellein [
19]. The preference for synthesis of calcaride A on sucrose or fructose suggests that glucose may repress the synthetic pathway. Production was not enhanced on maltose or longer chains of glucose. Efficient hydrolysis of the maltose, dextin or starch, however, may also have resulted in glucose repression. The hydrolytic enzymes produced by
Calcarisporia species have not been studied.
Table 1.
Biomass and Calcaride A (specific and volumetric) produced by Calcarisporium sp. KF525 grown in defined medium for ca. 25 days in flasks (100 rpm, 22 °C) with various carbon sources.
Table 1.
Biomass and Calcaride A (specific and volumetric) produced by Calcarisporium sp. KF525 grown in defined medium for ca. 25 days in flasks (100 rpm, 22 °C) with various carbon sources.
| Carbon Source | Biomass (g·L−1) | Calcaride A (mg·g−1) | Calcaride A (mg·L−1) |
|---|
| sucrose | 7.7 ± 0.6 bcd | 0.9 ± 0.1 c | 7.2 ± 0.8 b |
| fructose | 5.4 ± 0.04 ab | 1.5 ± 0.1 d | 8.1 ± 0.3 b |
| glucose | 6.3 ± 0.4 abc | 0.7 ± 0.02 bc | 4.7 ± 0.4 ab |
| maltose | 5.1 ± 0.2 a | 0.05 ± 0.02 a | 0.3 ± 0.1 a |
| malt extract | 9.7 cd | 0.4 ab | 3.9 ab |
| starch | 10.2 d | 0.2 a | 1.9 a |
| xylose | 5.3 ab | 0.2 a | 0.9 a |
| lactose | 4.4 ± 0.1 a | 0.1 ± 0.03 a | 0.4 ± 0.1 a |
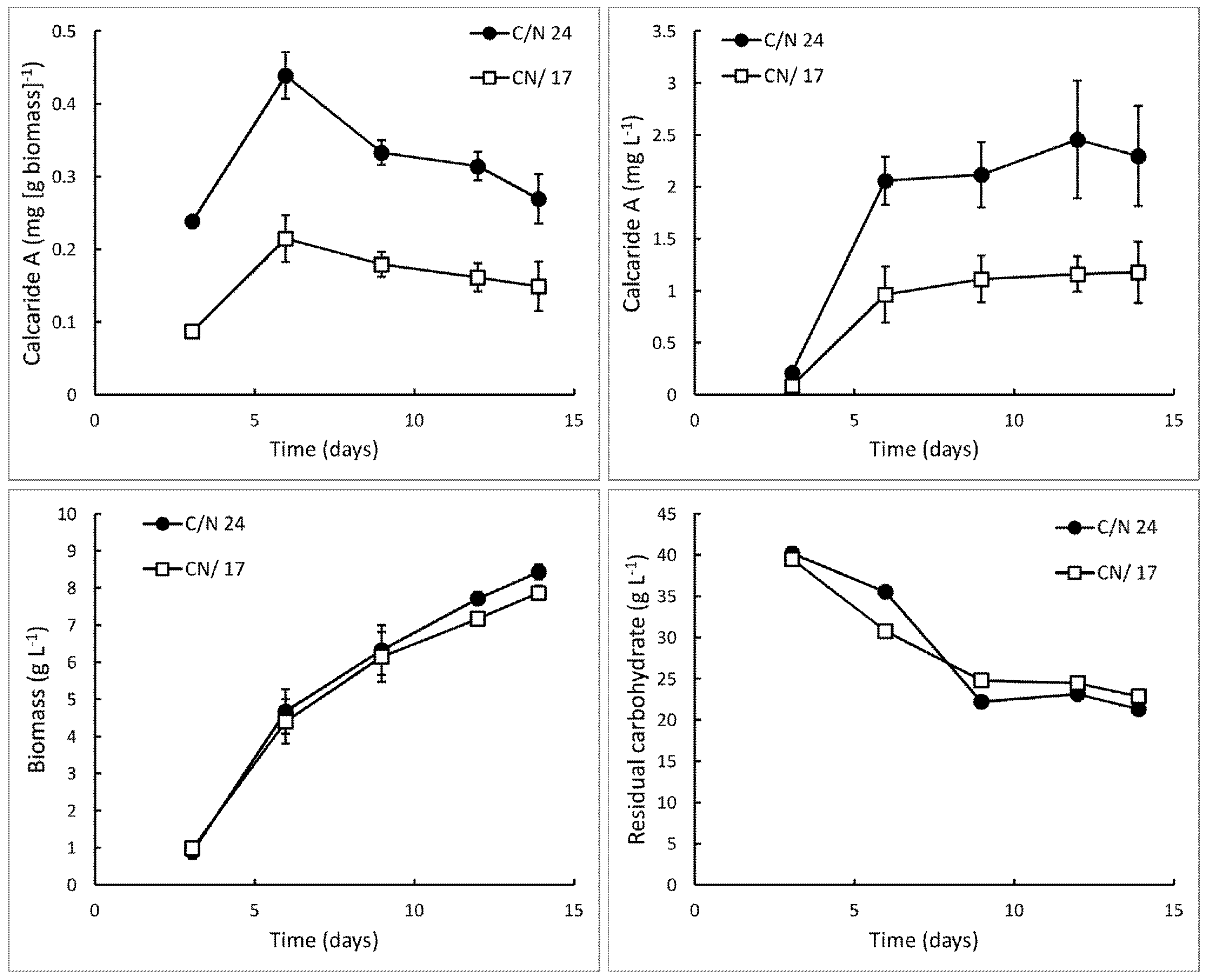
When grown in bioreactors for 21 days, with sucrose as the carbon source and yeast extract as the nitrogen source and a constant pH of 6.8 ± 0.2, KF525 produced more calcaride A from sucrose (0.02 mg·g−1 biomass, equivalent to 0.32 mg·L−1) than from glucose (0.01 mg·g−1 biomass, equivalent to 0.16 mg·L−1), but production was still very low and comparable to that previously observed in the bioreactor with casamino acids glucose medium. Biomass production was slightly higher on sucrose (13.4 ± 1.6 g·L−1) than on glucose (11.6 ± 1.2 g·L−1).
2.2. The Effect of pH on Calcaride A Production
Since production in defined medium in agitated flasks was as high as in complex (casamino acid glucose) medium in flasks,
Calcarisporium sp. KF525 was also grown in defined medium in the bioreactor. The pH of defined medium is lower than that of the casamino acid glucose medium described by Silber
et al. [
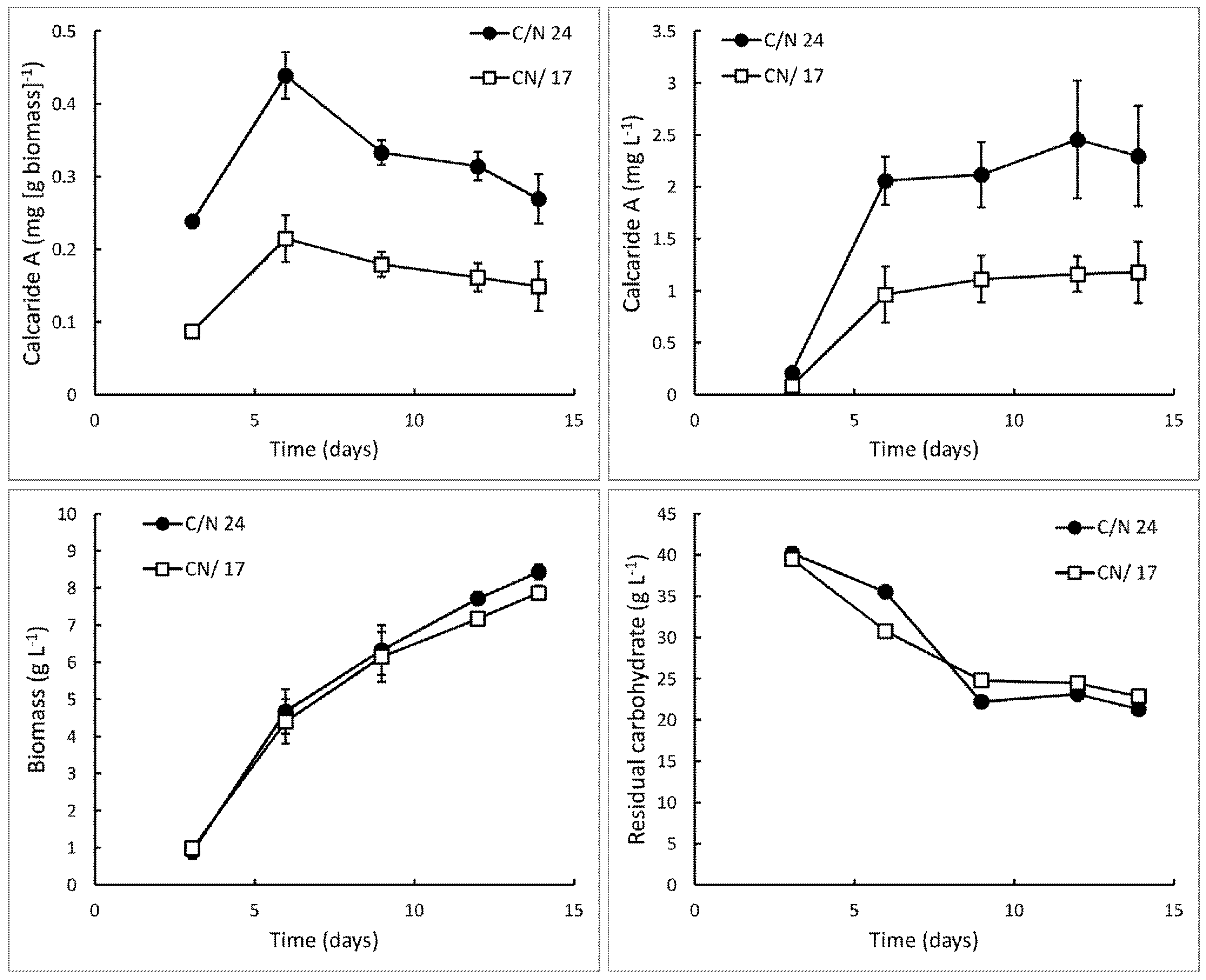
10], and these cultures were grown at pH 5.2 ± 0.3, the initial pH of the medium. Sucrose was provided as the carbon source. Within only 14 days, 0.3 ± 0.0 mg·g
−1 (equivalent to 2.3 ± 0.5 mg·L
−1;
Figure 2) calcaride A was produced, which was approximately ten times higher than previously obtained in a bioreactor. However, the carbohydrate was not completely consumed (9.0 ± 0.5 g·L
−1 glucose and 13.0 ± 0.8 g·L
−1 fructose remaining after 12–14 days), and only 8.4 ± 0.7 g·L
−1 biomass was produced (
Figure 3, C/N 24).
Figure 2.
Production of calcaride A and biomass and total residual carbohydrate (i.e., glucose, fructose and sucrose) in bioreactor cultures of KF525 in defined medium with sucrose as the carbon source. The pH was maintained at 5.2 ± 0.3. Nitrogen was supplied as (NH4)2SO4 (C/N 24) or (NH4)2SO4 supplemented with glutamine (C/N 17). Error bars represent ± sem (n = 2).
Figure 2.
Production of calcaride A and biomass and total residual carbohydrate (i.e., glucose, fructose and sucrose) in bioreactor cultures of KF525 in defined medium with sucrose as the carbon source. The pH was maintained at 5.2 ± 0.3. Nitrogen was supplied as (NH4)2SO4 (C/N 24) or (NH4)2SO4 supplemented with glutamine (C/N 17). Error bars represent ± sem (n = 2).
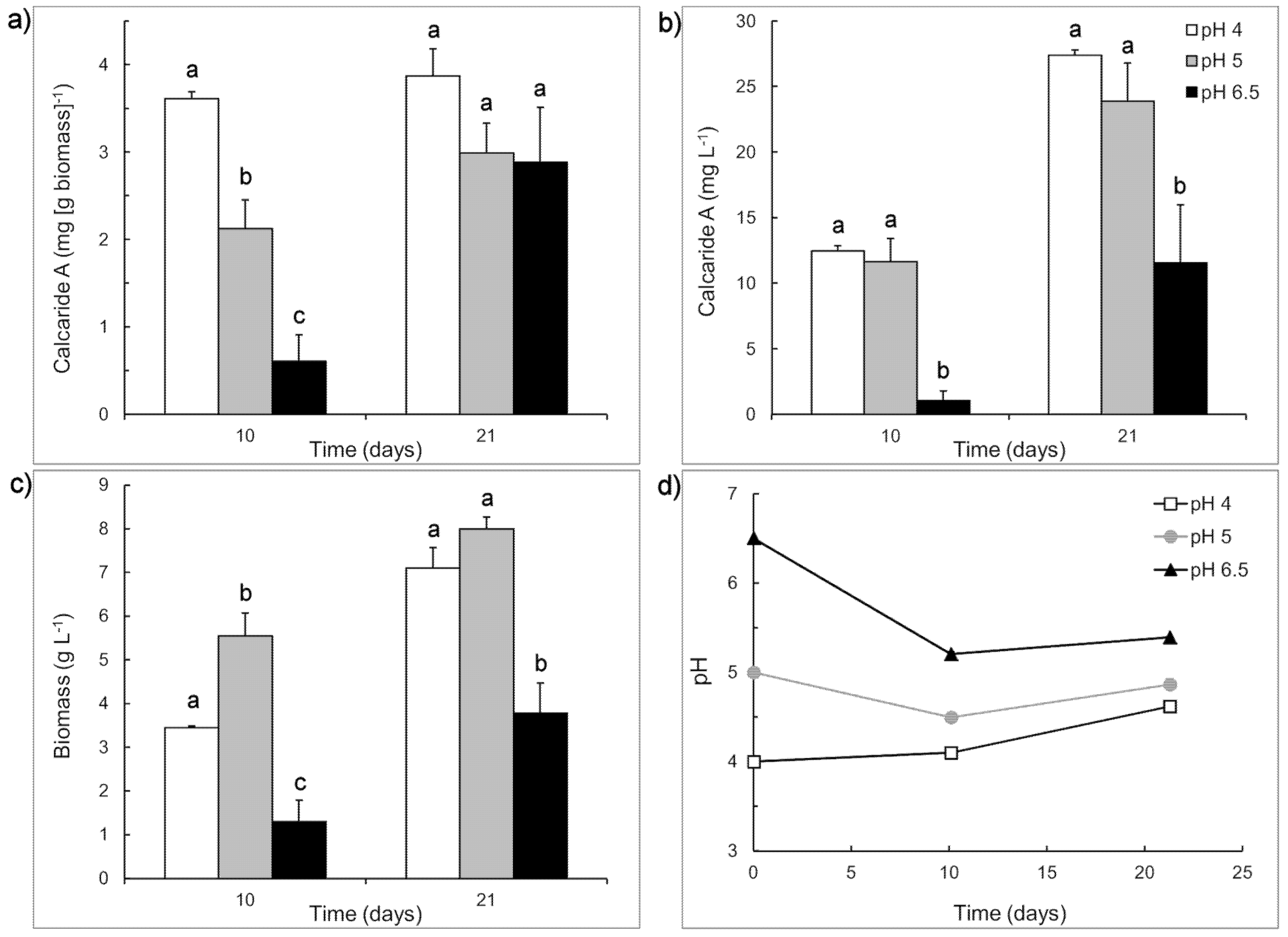
To assess the importance of the shift from pH 6.8 to pH 5.2 for obtaining calcaride A production in the bioreactor, we compared the production of calcaride A at different initial pH values in flasks using the casamino acid medium described by Silber
et al. [
10] and adjusting the initial pH to either 6.8 (pH 6.5 after sterilization) or 5.2 (pH 5.0 after sterilization). The comparison was made with sucrose as the carbon source and a lower pH (4.0) was included. During growth, the pH of cultures with initial pH of 6.5 decreased to 5.4 ± 0.0, whereas the pH of cultures with initial pH of 4 increased to 4.6 ± 0.1 and the pH of cultures with initial pH of 5 changed very little (
Figure 3).
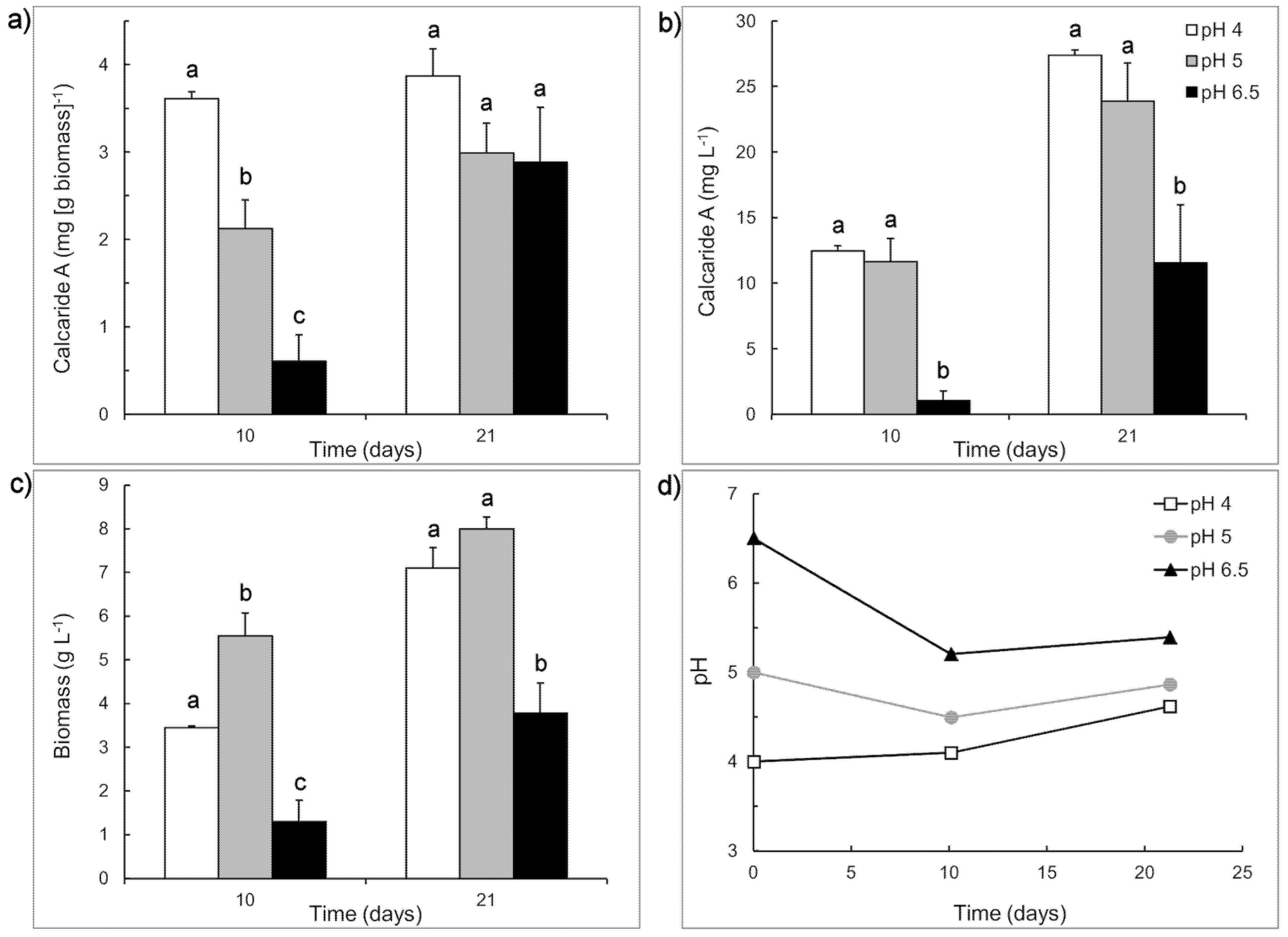
Production of calcaride A (volumetric) was similar at pH 4 and 5 and significantly (
p < 0.05) lower at pH 6.5 after 10 and 21 days incubation, reflecting both higher initial (
i.e., at day 10) specific concentrations of calcaride A in the mycelia at pH 4 or 5, compared to pH 6.5, and higher biomass production (
Figure 3). The amount of calcaride A produced in casamino acid sucrose medium (
Figure 3) was higher than that previously observed in defined medium with sucrose as the carbon source (
Table 1), with up to 3.9 ± 0.3 mg·g
−1 biomass (equivalent to 27.4 ± 0.4 mg·L
−1) calcaride A produced at pH 4 after 21 days incubation. Glucose and/or fructose, but no sucrose, were still present after 21 days of incubation in cultures with initial pH 4 (3.9 ± 0.0 g·L
−1 glucose), pH 5 (6.5 ± 0.1 g·L
−1 fructose) and pH 6.5 (9.7 ± 0.9 g·L
−1 glucose, 16.2 ± 0.4 g·L
−1 fructose). Since the pH of cultures with initial pH 4 increased during incubation, and volumetric calcaride A production was similar at pH 4 and 5, pH values around 5 were used for subsequent investigations.
Figure 3.
Production of (a) specific and (b) volumetric calcaride A and (c) biomass by KF525 in sucrose-casamino acid medium with initial pH of 4 (white), 5 (grey) or 6.5 (black). (d) Changes in pH during the cultivation. Error bars represent + sem (n = 2 or 4) and are smaller than the size of the symbol for pH. Letters above the bars (a, b or c) indicate significant differences (p < 0.05) for values measured at the same time (Day 10 or Day 21).
Figure 3.
Production of (a) specific and (b) volumetric calcaride A and (c) biomass by KF525 in sucrose-casamino acid medium with initial pH of 4 (white), 5 (grey) or 6.5 (black). (d) Changes in pH during the cultivation. Error bars represent + sem (n = 2 or 4) and are smaller than the size of the symbol for pH. Letters above the bars (a, b or c) indicate significant differences (p < 0.05) for values measured at the same time (Day 10 or Day 21).
Both calcaride A and biomass were preferentially produced at low pH, explaining why production in defined medium, with initial pH ~5.2, was generally better than in medium with organic nitrogen sources (casamino acids or yeast extract), with initial pH 6.5 to 7. It also explained why production in complex medium in flasks with initial pH 6.5–6.8 was better than in bioreactors at pH 6.8 (
Section 2.1), since the pH in the flasks decreased to levels at which calcaride A production would be enhanced, whereas the pH of the bioreactors had been kept constant. While many filamentous fungi prefer to grow at pH values between 4 and 6, marine fungi are often grown in media with initial pH values between 6 and 7 when screening for compounds, presumably because the marine environment would be slightly alkaline (pH 7.5–8.4). Some compounds are optimally produced in this range (e.g., [
6,
8]), but the optimum is likely to be strain and product specific.
2.3. The Effect of Nitrogen and Carbon Concentrations on Calcaride A Production
The casamino acid glucose medium described in [
10] does not supply sufficient nitrogen for all of the carbon to be consumed unless carbon is incorporated into storage compounds or secondary metabolites. The carbon–nitrogen ratio (C/N) of this medium was ~67 (g/g), which was much higher than the level at which nitrogen is expected to limit growth (C/N > 10; [
7,
20]). A high C/N ratio may be useful to trigger production of secondary metabolites such as the calcarides that do not contain nitrogen, and we continued to use medium with high C/N ratios. However, sucrose was not completely consumed in complex (C/N ~67,
Section 2.2) or defined (C/N ~24,
Figure 2) medium, suggesting that the C/N ratio could be reduced, either by increasing the amount of nitrogen supplied or by reducing the carbon concentration. When the concentration of nitrogen in the defined medium was increased (C/N ~17), supplied as either more ammonium or as the amino acid glutamine, sucrose consumption and biomass production were essentially unaffected, indicating that nitrogen remained limiting, while calcaride A production was reduced by about 50% (
Figure 2). We therefore considered C/N ratios between ~24 and ~67.
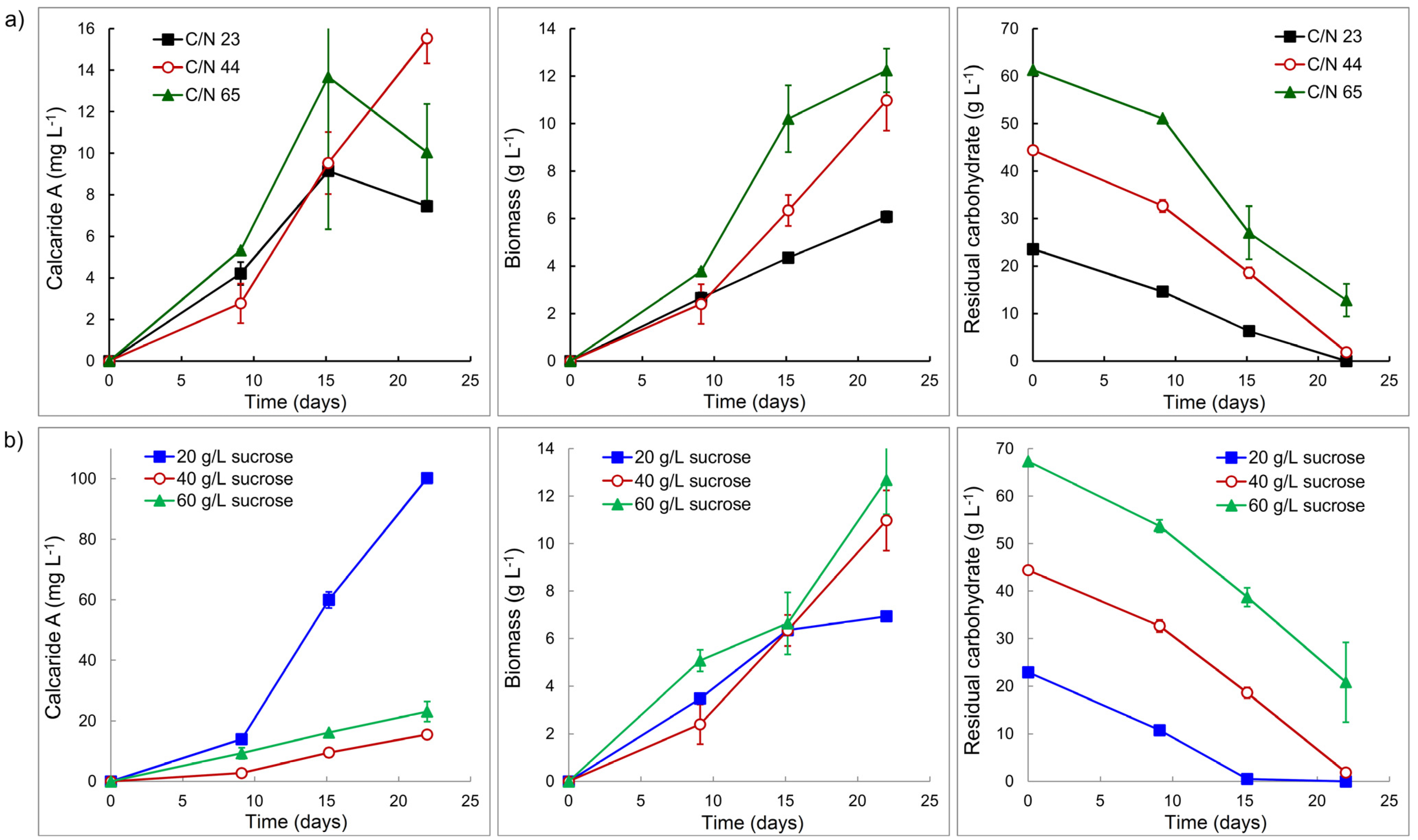
Two sets of conditions were investigated in flasks, either keeping the concentration of the nitrogen source (glutamine) constant but changing the concentration of sucrose to change the C/N ratio or adjusting the concentration of glutamine to maintain a constant C/N ratio for different concentrations of sucrose. Glutamine was used as the nitrogen source since results obtained while reviewing the protocol to generate conidia of KF525 indicated that glutamine supported better production of calcaride A than ammonium or casamino acids. Amino acids have also been shown to affect production of mullein and 4-hydroxymellein in
A. ochraceous [
19]. Production of calcaride A in flasks with a constant concentration of glutamine, but different concentrations of sucrose (20, 40 or 60 g·L
−1 to provide C/N 23, 44 or 65), was not affected (
p > 0.05) by the change in C/N (
Figure 4a). The yield of biomass from sucrose (0.26 g biomass per g sucrose consumed) was the same at all C/N ratios, but significantly (
p < 0.05) less biomass was produced from 20 g·L
−1 sucrose (C/N 23) than from 40 or 60 g·L
−1 sucrose. Carbohydrate (sucrose, glucose or fructose) was not completely consumed at C/N 65 with 60 g·L
−1 sucrose provided, but was consumed at C/N 44 and 23 (
Figure 4a). Since all carbohydrate was consumed at C/N 44, we used this ratio of carbon and nitrogen to assess whether providing more or less sucrose could be beneficial.
Reducing the C/N from 65 to 44 with 60 g·L
−1 sucrose did not enable KF525 to utilize all the carbohydrate (
Figure 4b) and production of calcaride A and biomass was similar (
p > 0.05) at C/N 44 and C/N 65 (
Figure 4). However, increasing the C/N from 23 to 44 with 20 g·L
−1 sucrose resulted in a significant (
p < 0.05) increase in calcaride A production (
Figure 4b), although biomass production was not (
p > 0.05) affected. All carbohydrate was consumed within 15 days. High calcaride A production thus occurred during a carbon-limited stationary phase, as might be expected for a secondary metabolite. Calcaride A production was significantly higher (
p < 0.05, ANOVA) than in flasks with 40 or 60 g·L
−1 sucrose after the sucrose had been consumed (
Figure 4b). Biomass production after 21 days with 40 g·L
−1 sucrose and 2 g·L
−1 glutamine (C/N 44,
Figure 4) was comparable (
p > 0.05) to that obtained with 40 g·L
−1 sucrose and casamino acids at pH 4–5 (C/N ~67,
Figure 3), but calcaride A production was lower (
p < 0.05).
Figure 4.
(a) Production of calcaride A and biomass by KF525 and residual carbohydrate (sucrose, glucose and fructose) concentration in sucrose glutamine (2 g·L−1) medium with initial pH of 5 and variable C/N. (b) Production of calcaride A and biomass by KF525 and residual carbohydrate concentration in sucrose glutamine medium with initial pH of 5 and C/N ratio of 44. Error bars represent ± sem (n = 2).
Figure 4.
(a) Production of calcaride A and biomass by KF525 and residual carbohydrate (sucrose, glucose and fructose) concentration in sucrose glutamine (2 g·L−1) medium with initial pH of 5 and variable C/N. (b) Production of calcaride A and biomass by KF525 and residual carbohydrate concentration in sucrose glutamine medium with initial pH of 5 and C/N ratio of 44. Error bars represent ± sem (n = 2).
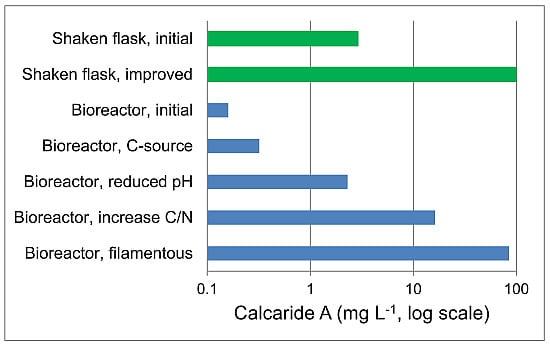
2.4. Improved Production of Calcaride A in Bioreactor Cultures
Reducing the pH from 7.0 to 5.2 had resulted in measureable calcaride A production in stirred tank reactors (
Figure 2), but production was lower than observed in the equivalent flasks. The best production in flasks was obtained in medium with 20 g·L
−1 sucrose and glutamine as the nitrogen source with a C/N ratio of 44. When this condition was applied in a bioreactor (pH 5.4), calcaride A production increased to 1.4 mg·g
−1 (16.2 mg·L
−1;
Figure 5, 20 g/L sucrose pellets). This was comparable to the production observed after 21 days in flasks with 40 or 60 g·L
−1 sucrose (
Figure 4). Calcaride A was produced during growth, and no further calcaride A production occurred after the carbohydrate had been consumed (11 days) (
Figure 5). Although production was lower than in flasks, the time required to produce calcaride A was reduced by nearly half. Production of calcaride A was similar at pH 4 as at pH 5.4 (data not shown). When the concentration of sucrose was increased to 40 g·L
−1, calcaride A production was similar to that observed with 20 g·L
−1 sucrose during the first eight days, but then decreased (
Figure 5, 40 g/L pellets).
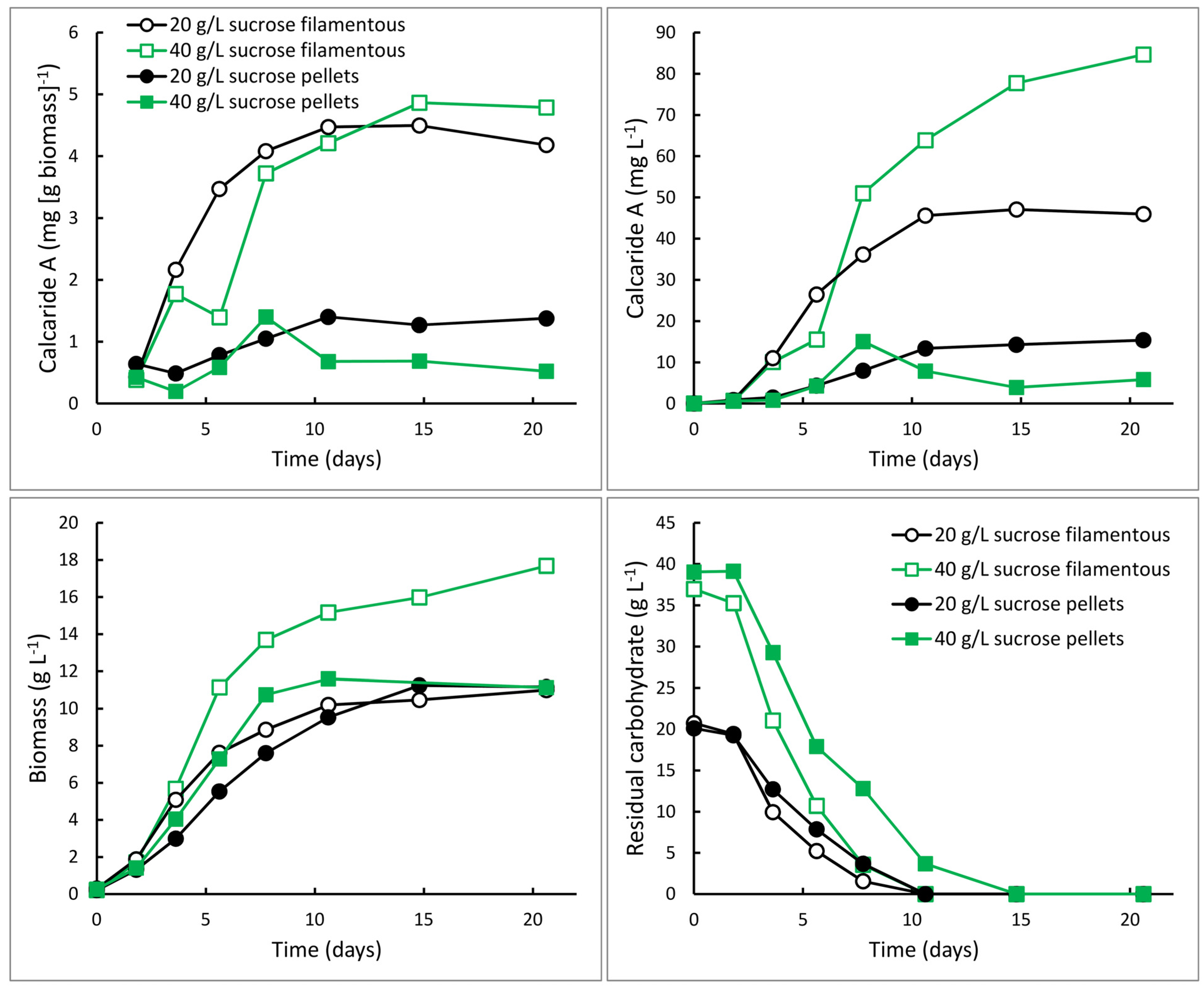
Figure 5.
Production of calcaride A and biomass and consumption of carbohydrate (sucrose and glucose and fructose derived from sucrose) by KF525 in bioreactors with 20 (circles) or 40 (squares) g·L−1 sucrose, and nitrogen supplied as glutamine (C/N 44, pH 5.4 ± 0.1). The mycelium grew as pellets (solid symbols) or filamentously (open symbols).
Figure 5.
Production of calcaride A and biomass and consumption of carbohydrate (sucrose and glucose and fructose derived from sucrose) by KF525 in bioreactors with 20 (circles) or 40 (squares) g·L−1 sucrose, and nitrogen supplied as glutamine (C/N 44, pH 5.4 ± 0.1). The mycelium grew as pellets (solid symbols) or filamentously (open symbols).
KF525 generally grew as pellets in flasks and inclusion of agar to obtain filamentous growth (
cf. [
7]) appeared to suppress calcaride A production (data not shown). Therefore, bioreactors were inoculated with pre-cultures containing small pellets and the cultures continued to grow as small pellets (1–3 mm diameter). Unexpectedly, pre-cultures for two sucrose glutamine cultivations were filamentous and grew filamentously in the bioreactors. These cultivations grew more rapidly than the cultures that consisted of pellets and they consumed the substrate efficiently (
Figure 5), as expected for filamentous compared to pellet growth. Unexpectedly, they also produced more calcaride A (~4.5 mg·g
−1) than cultures with pellets (
Figure 5). All sucrose was consumed, even when 40 g·L
−1 sucrose was supplied, resulting in a higher biomass concentration and consequently higher volumetric production of calcaride A with 40 compared with 20 g·L
−1 sucrose. The amount of calcaride A produced in the filamentous bioreactor culture with 40 g·L
−1 sucrose (85 mg·L
−1) was comparable to that obtained from flasks with 20 g·L
−1 sucrose and 1 g·L
−1 glutamine (100 mg·L
−1). Neither bioreactor cultures with pellets nor bioreactor cultures with filamentous mycelia were oxygen limited, but nutrient (sucrose or oxygen) diffusion through the pellets may be limiting calcaride A production in stirred tank reactors. It was not clear why calcaride A was produced well by pellets in flasks, even when shaken at only 100 rpm, but not in bioreactors. Furthermore, additional study would be needed to develop a protocol to consistently generate inocula with the correct morphology for bioreactor production of calcaride A. None-the-less, the results demonstrate that production of substantial amounts of calcaride A in bioreactors is feasible.
A variety of strategies have been applied to obtain production of secondary metabolites from marine fungi in stirred tank bioreactors. Aspergiolide A production required a low shear environment, adequate oxygen [
5] and pH near 7 [
21]. Zhou
et al. [
6] and Tamminen
et al. [
7] found that conditions which promoted good biomass production promoted the production of 1403C (an anthracycline analogue) and scopularide A, respectively. The carbon–nitrogen ratio is important in the regulation of both fatty acid and polyketide synthesis and was optimized to enhance production of oxalicumone A [
22]. Both carbon and nitrogen sources may be important [
8,
22].
The C/N ratio and the source of carbon were both important in production of calcaride A. The nitrogen source may also be important, but was not systematically investigated. Because of the slow growth of Calcarisporium sp. KF525 and the need to maintain a relatively high C/N ratio, it was not possible to improve calcaride A production by increasing biomass production by KF525 or by using fed-batch strategies. The observation that filamentous mycelia were more productive than pellets suggests that agitation and the state of the pre-cultures are important and should be investigated further. Indeed it should be possible to further enhance calcaride A production in bioreactors now that detectable production has been achieved.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}