
Anti-Restenotic Roles of Dihydroaustrasulfone Alcohol Involved in Inhibiting PDGF-BB-Stimulated Proliferation and Migration of Vascular Smooth Muscle Cells

 , , ,
, , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
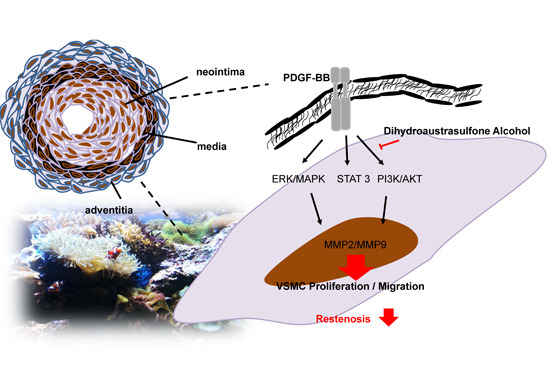
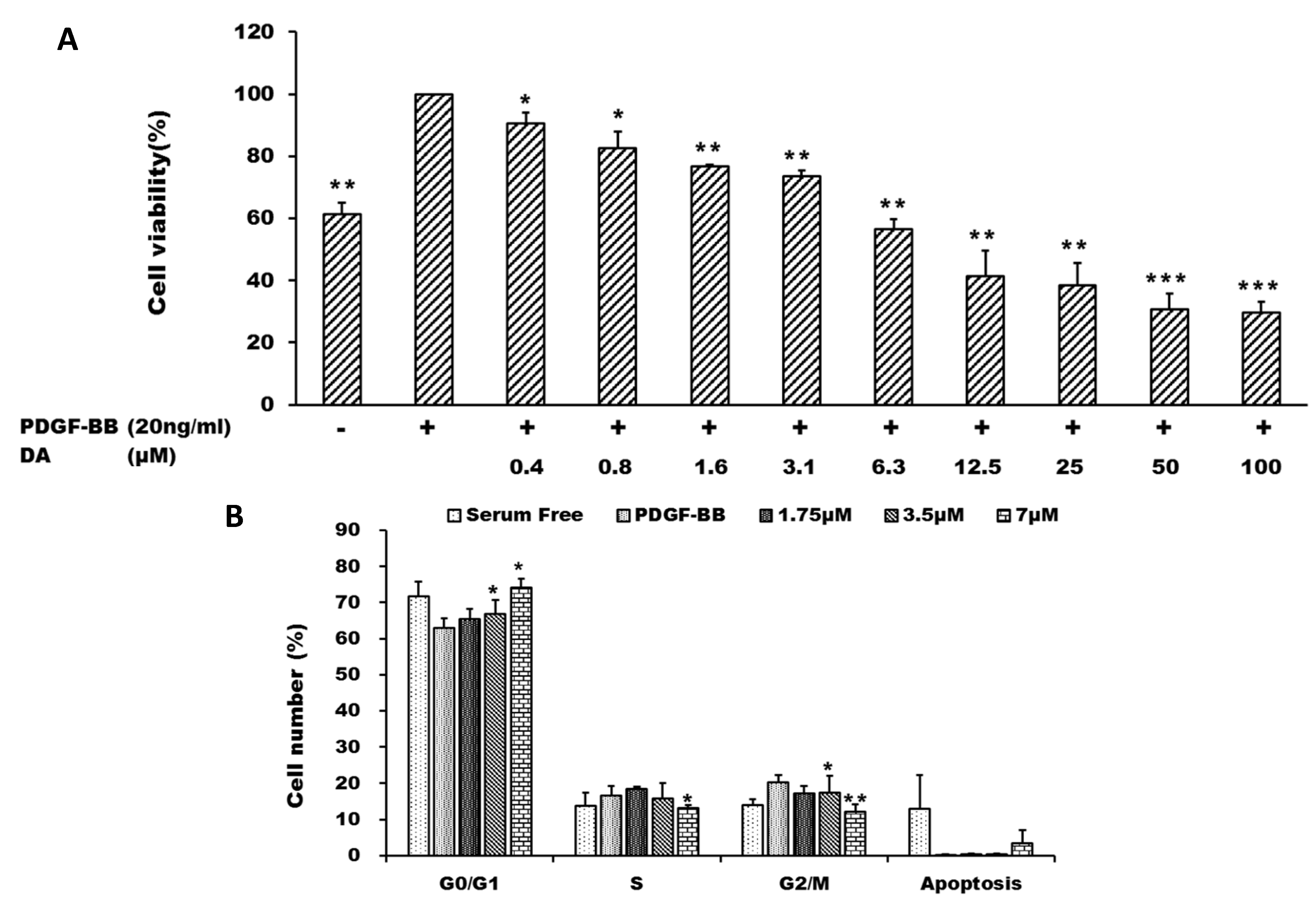
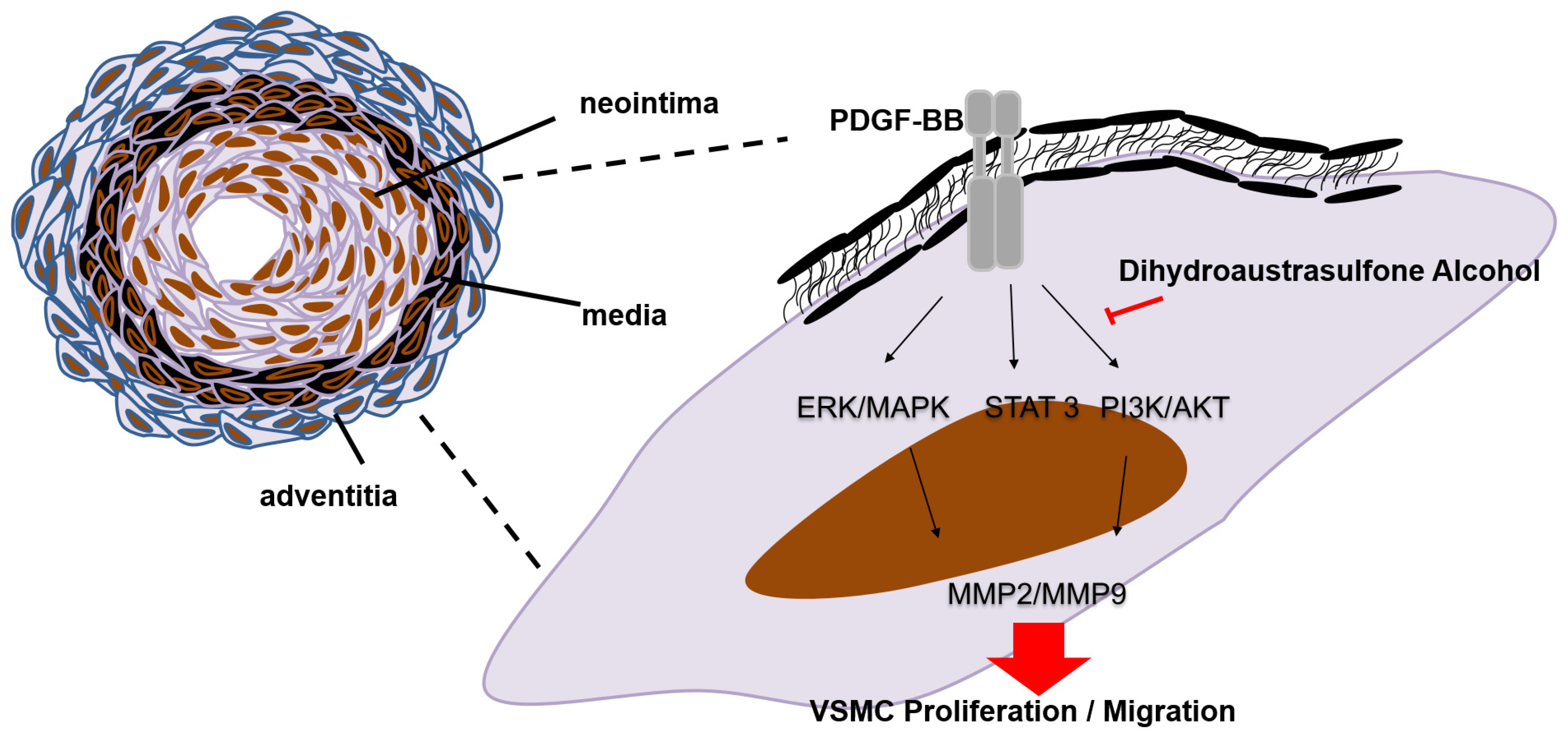
2.1. Dihydroaustrasulfone Alcohol Inhibited Platelet-Derived Growth Factor (PDGF)-BB-Induced Vascular Smooth Muscle Cell (VSMC) Proliferation
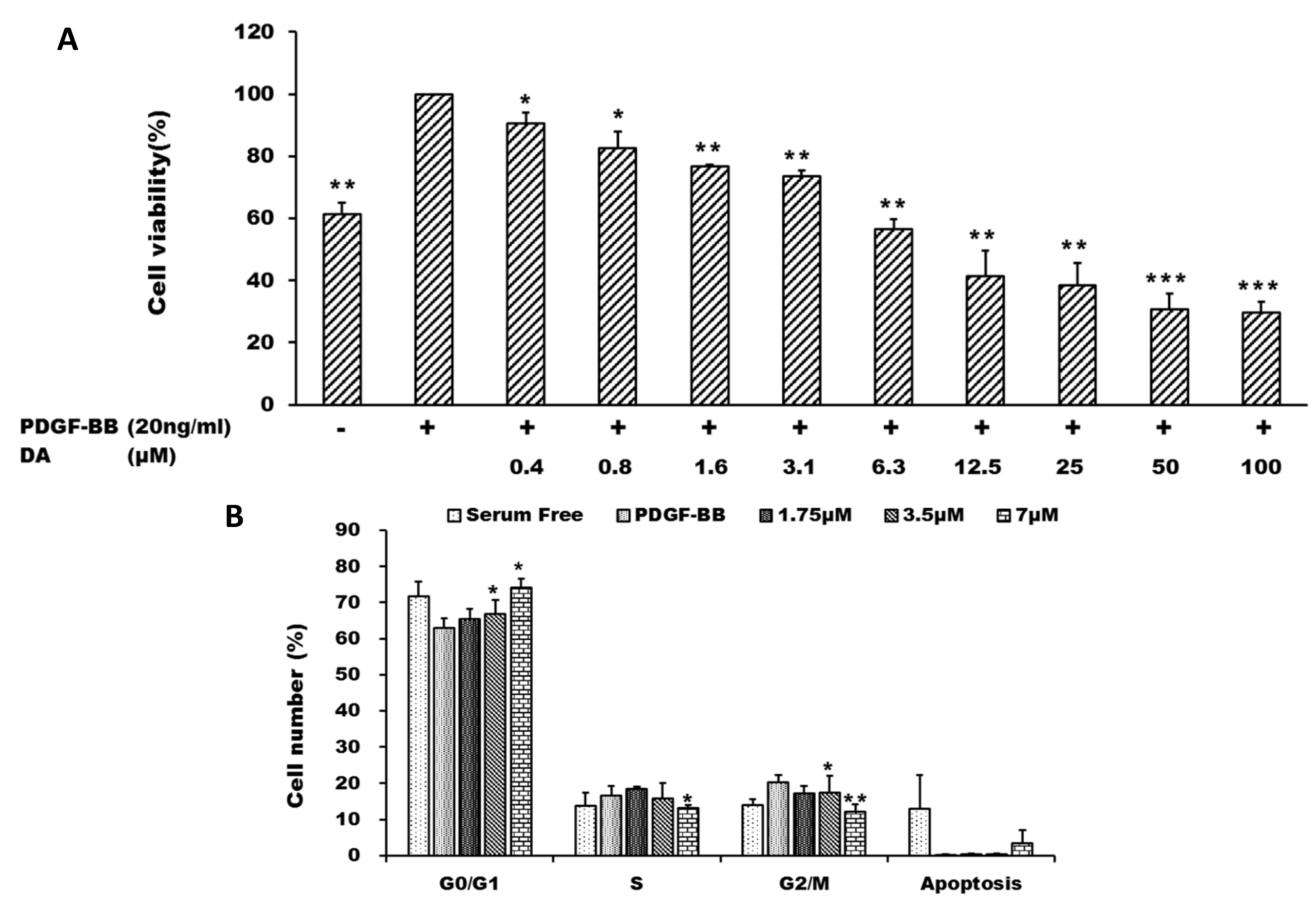
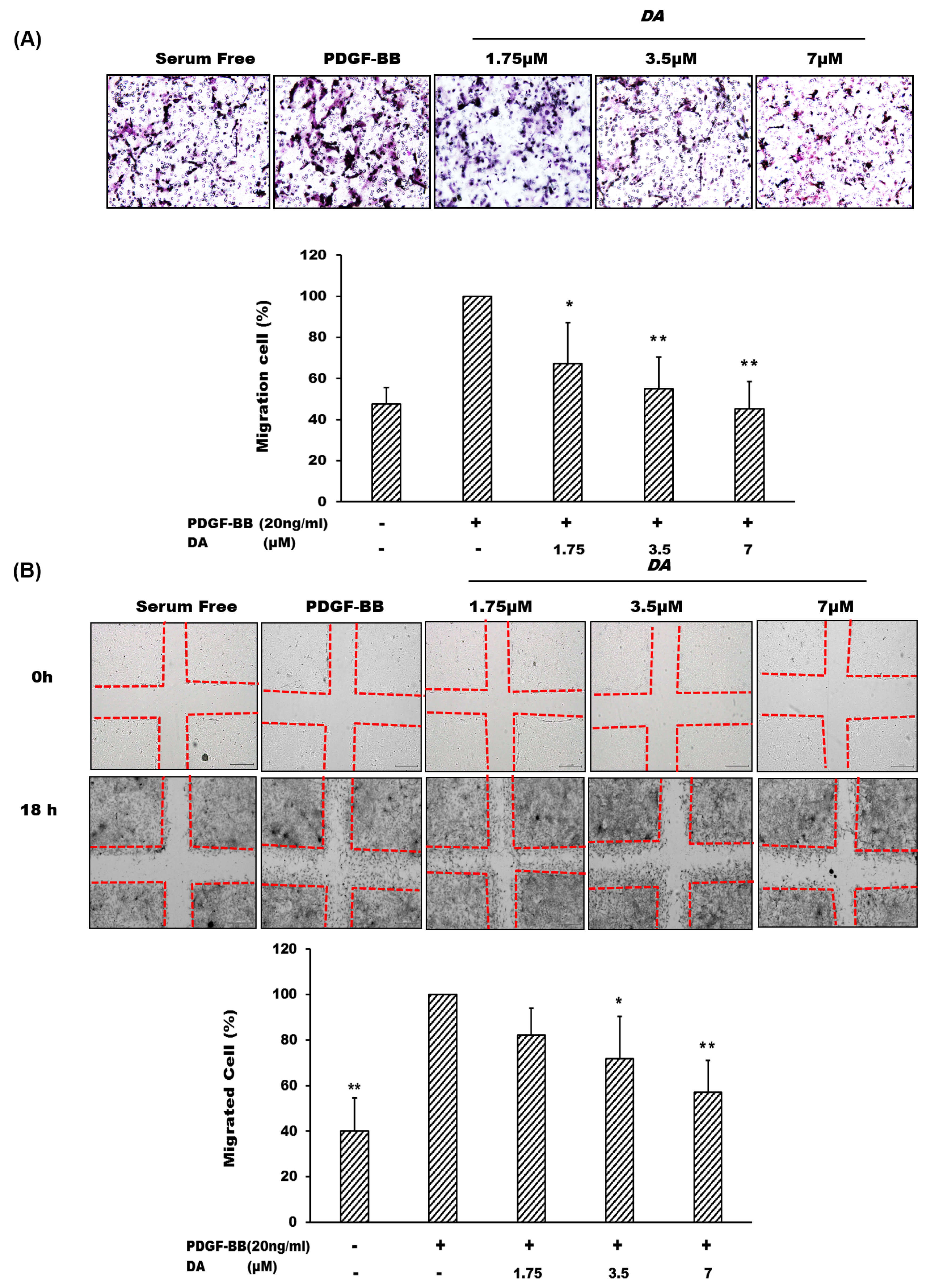
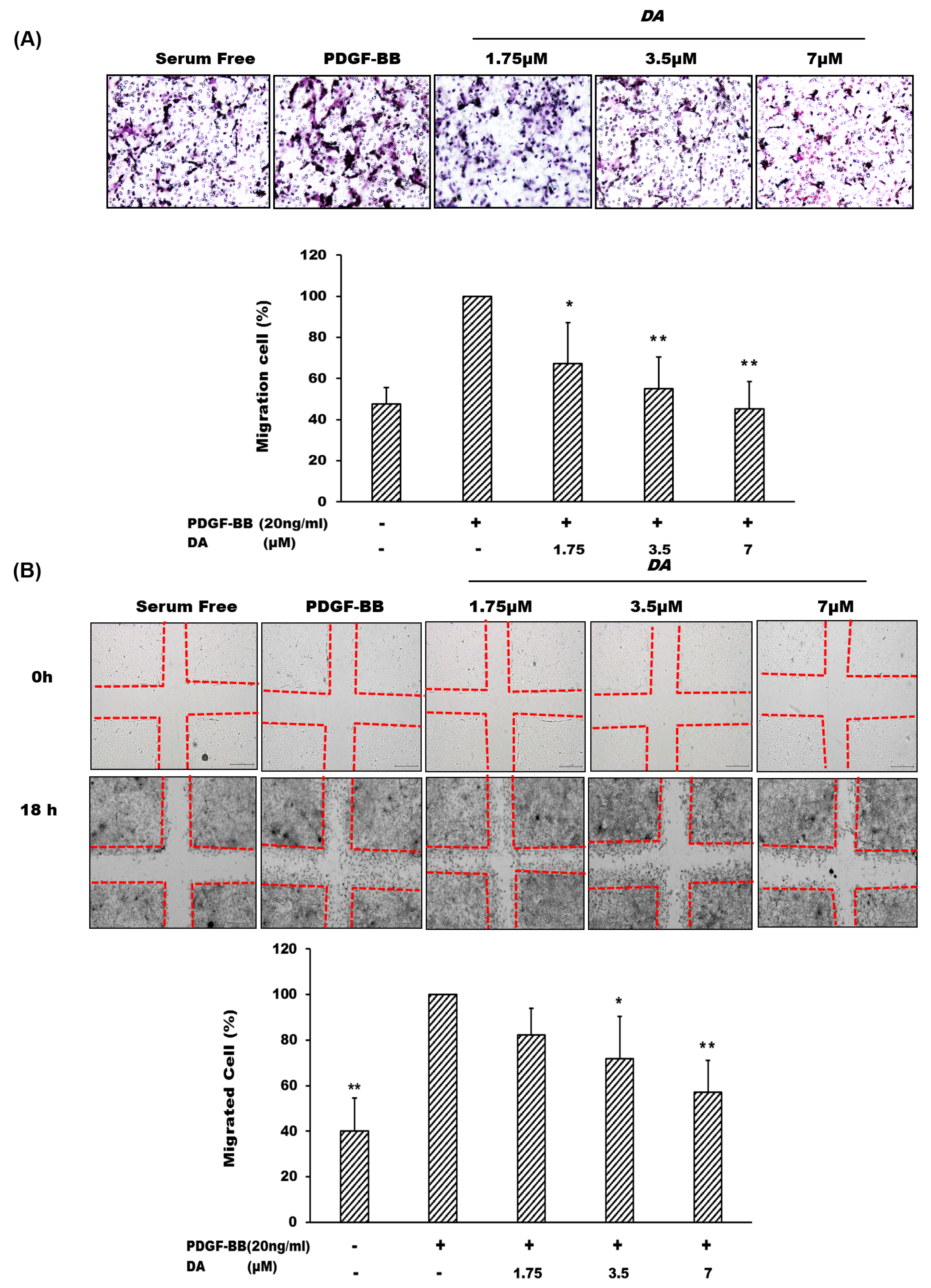
2.2. Dihydroaustrasulfone Alcohol Inhibits PDGF-BB-Induced VSMC Migration
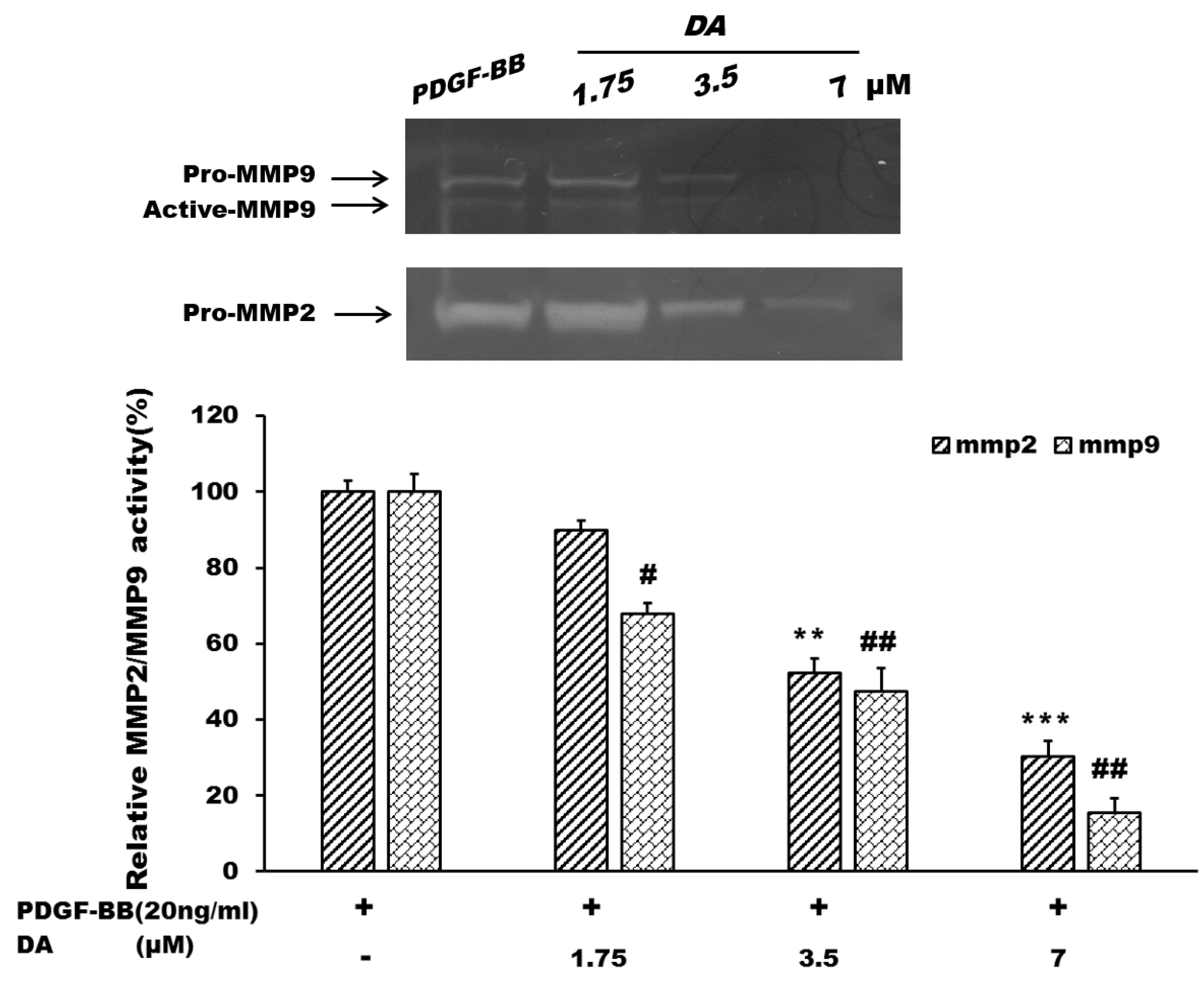
2.3. Effects of Dihydroaustrasulfone Alcohol on the Proliferative and Migration-Associated Proteins in VSMCs

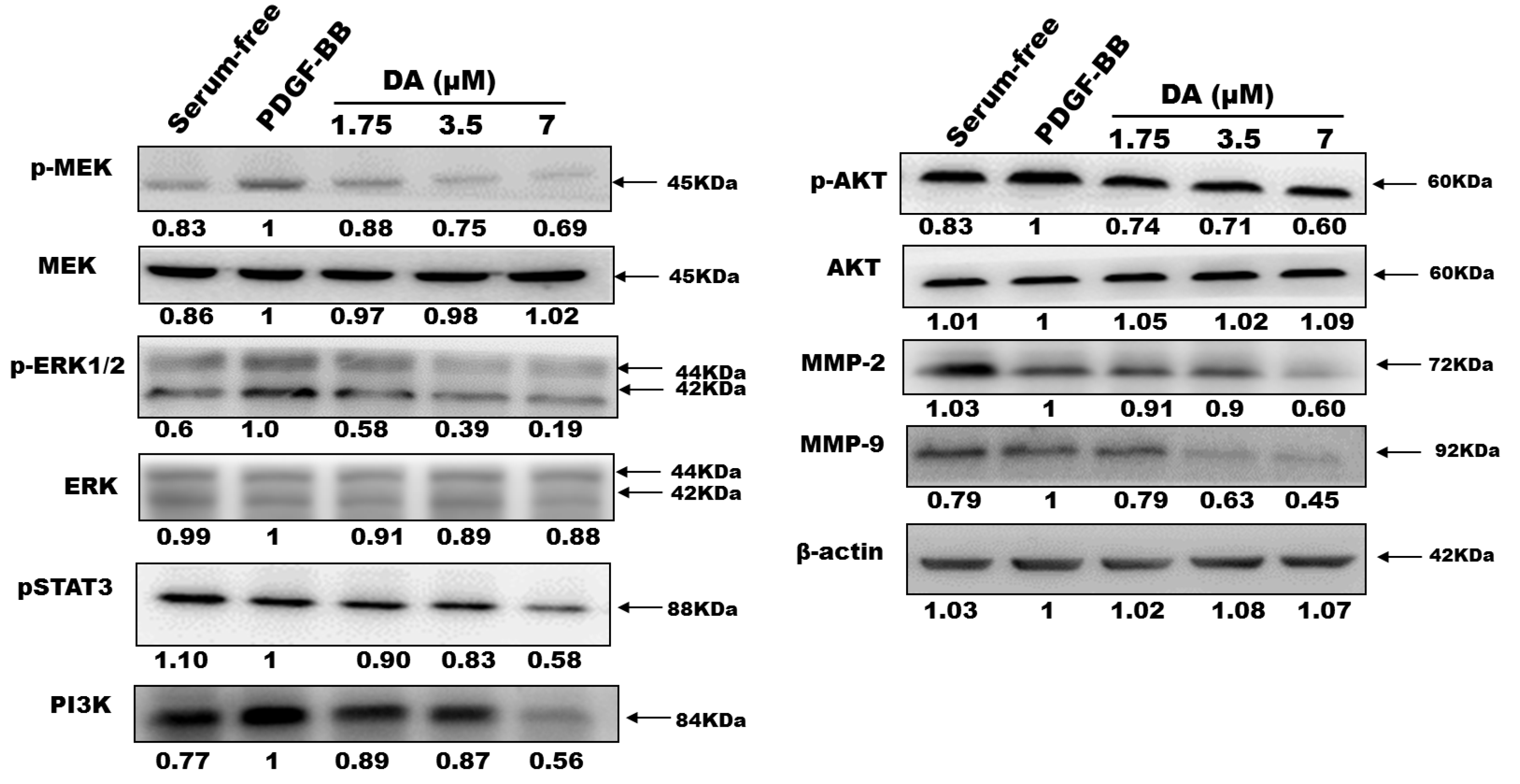
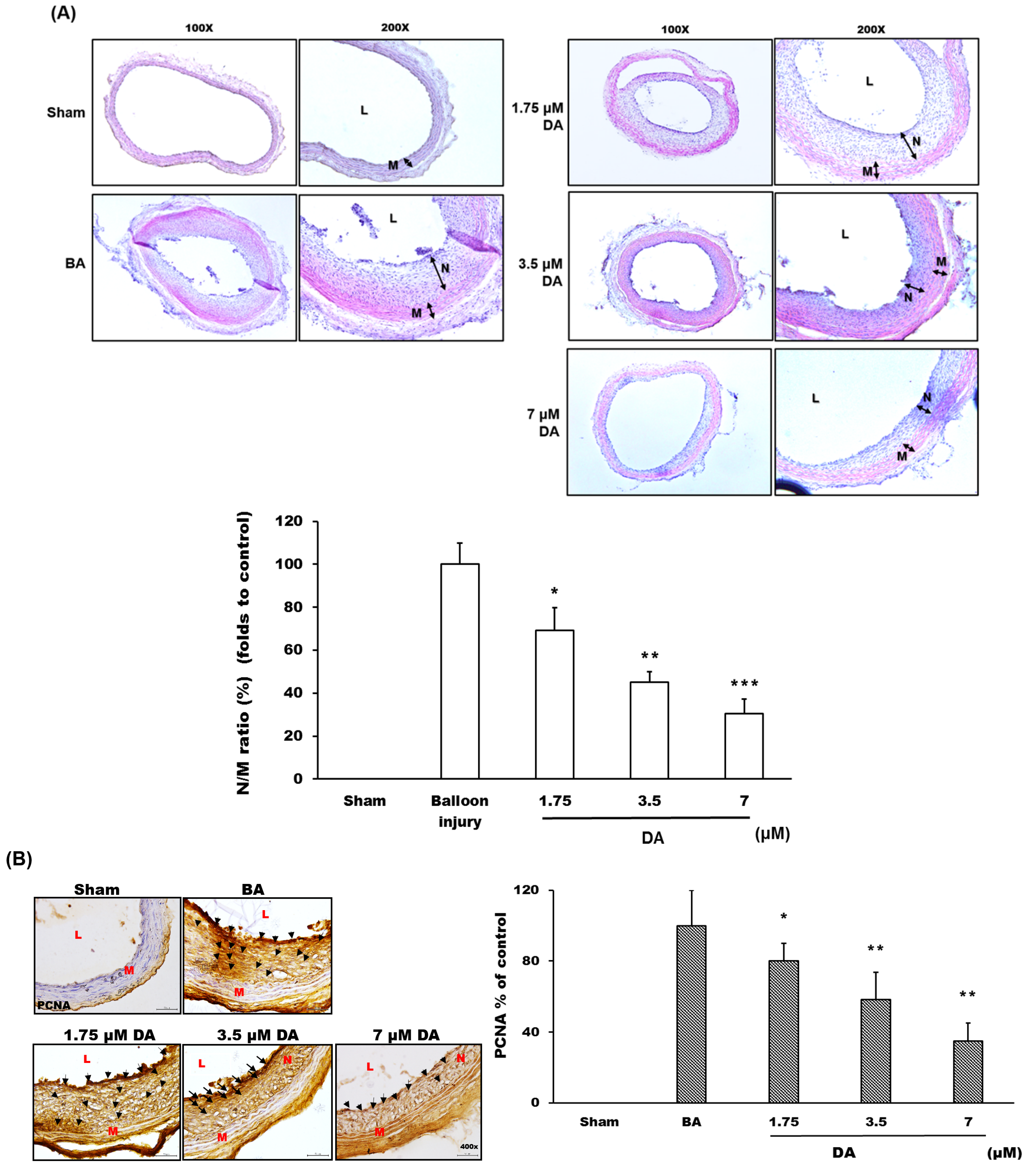
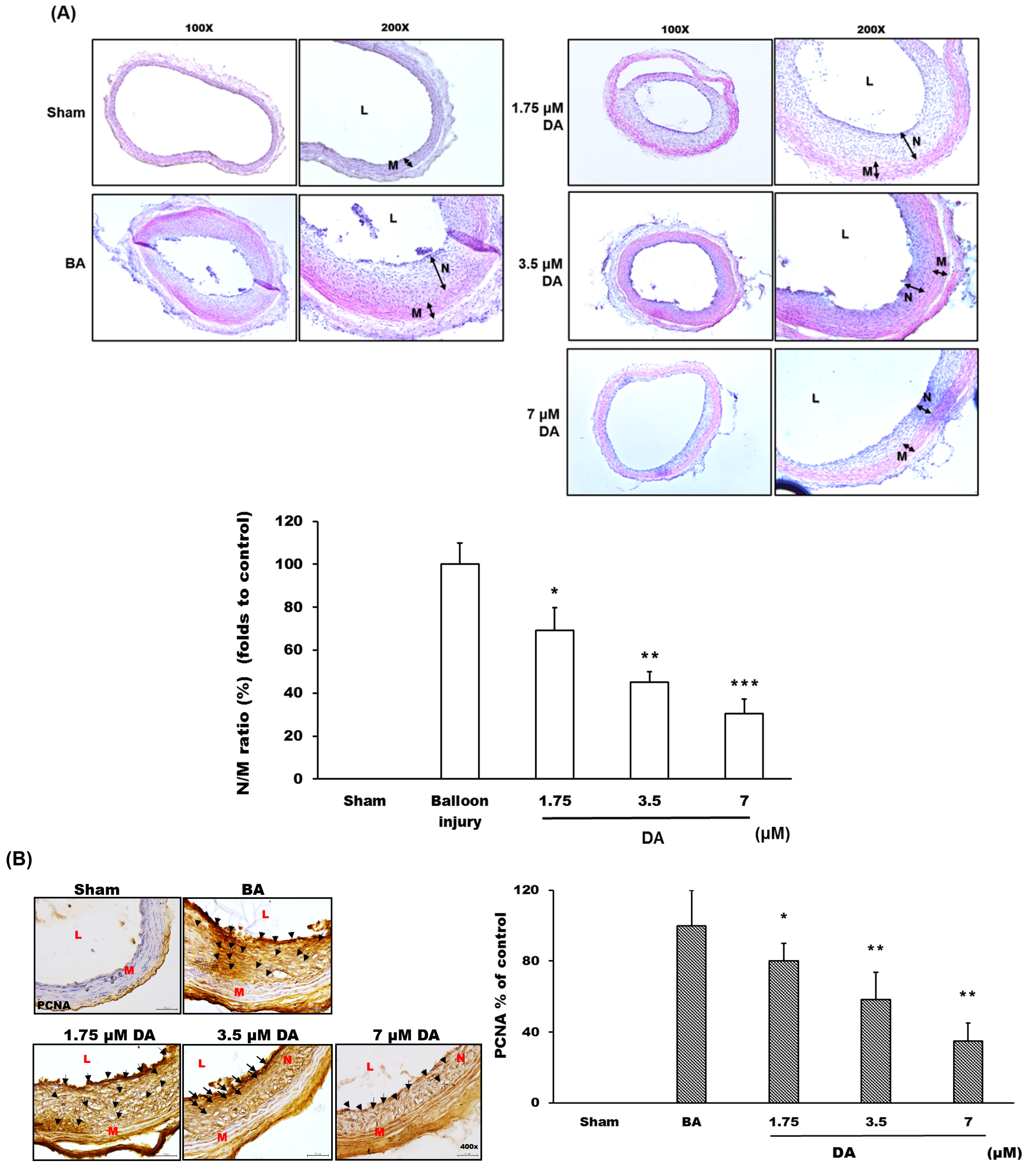
2.4. Dihydroaustrasulfone Alcohol Inhibited Neointimal Hyperplasia

3. Experimental Section
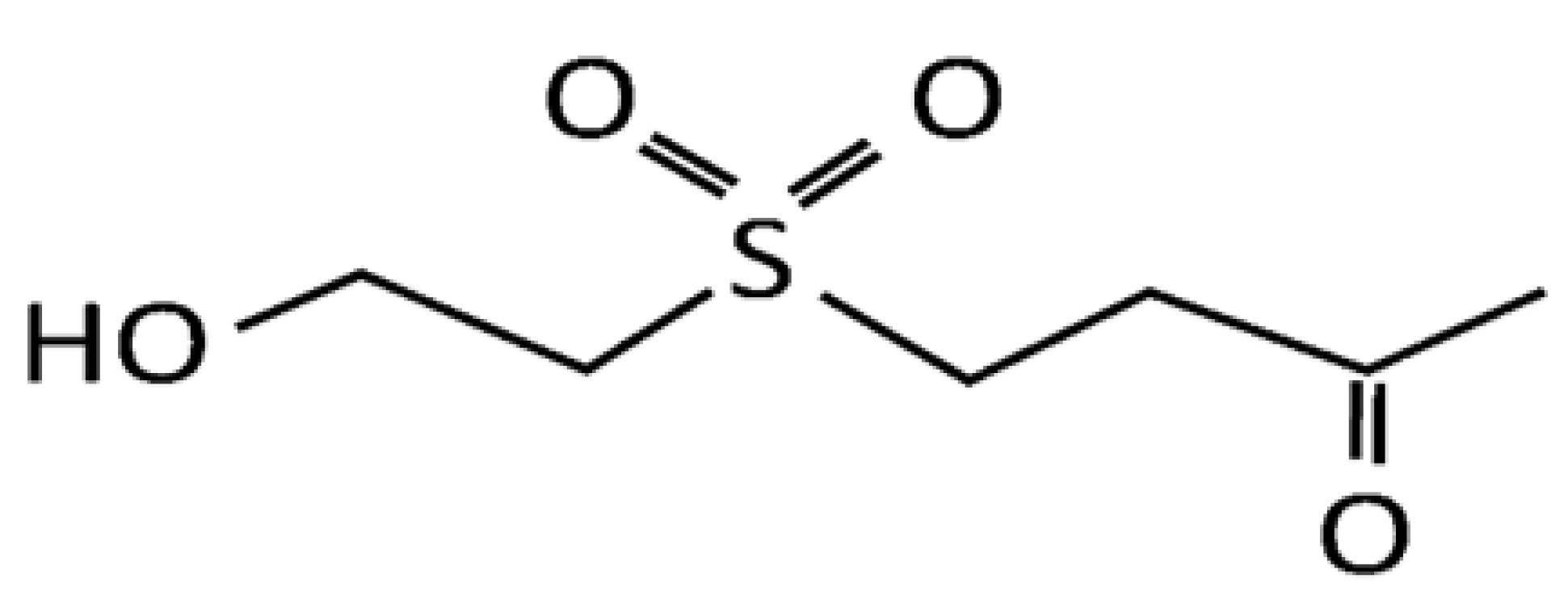
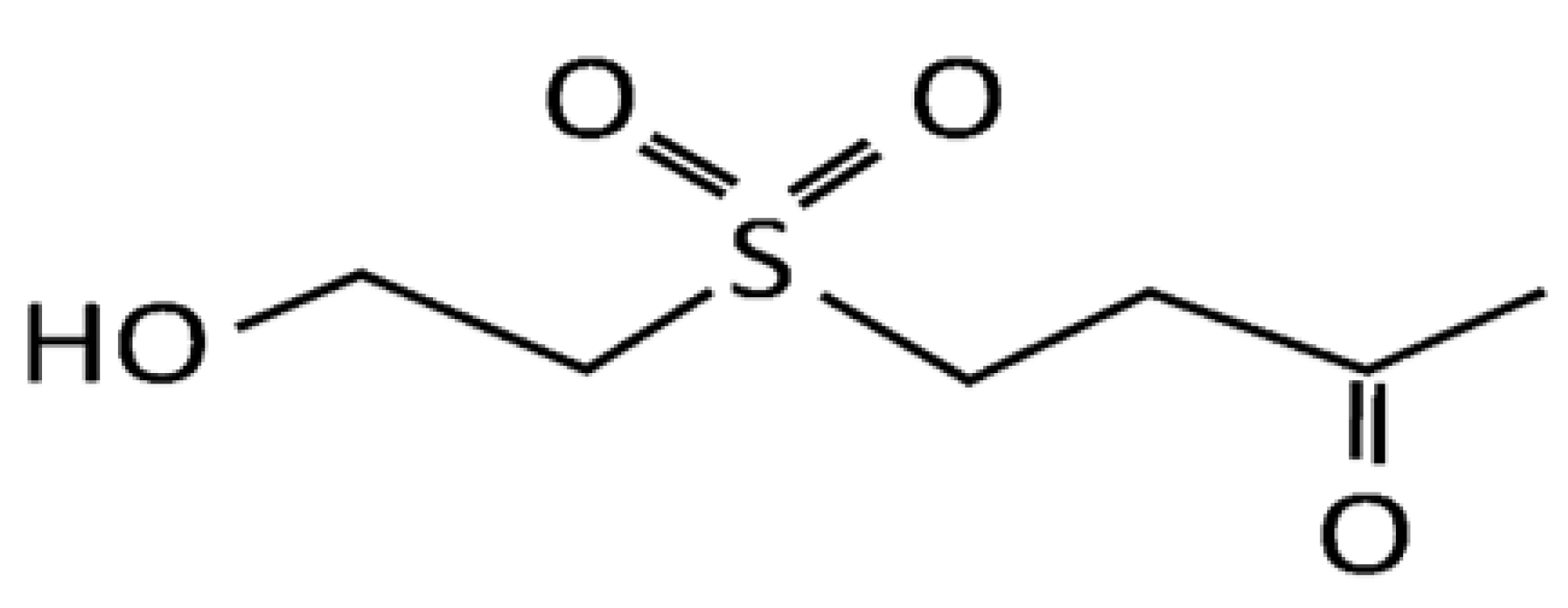
3.1. Materials
3.2. Cell Culture and MTT Cell Proliferation Assay
3.3. Flow Cytometry
3.4. Cell Migration Assay
3.5. In-Gel Gelatinase Zymography
3.6. Western Blot
3.7. Rat Carotid Artery Balloon Angioplasty
3.8. Histopathological Analysis
3.9. Statistical Analysis
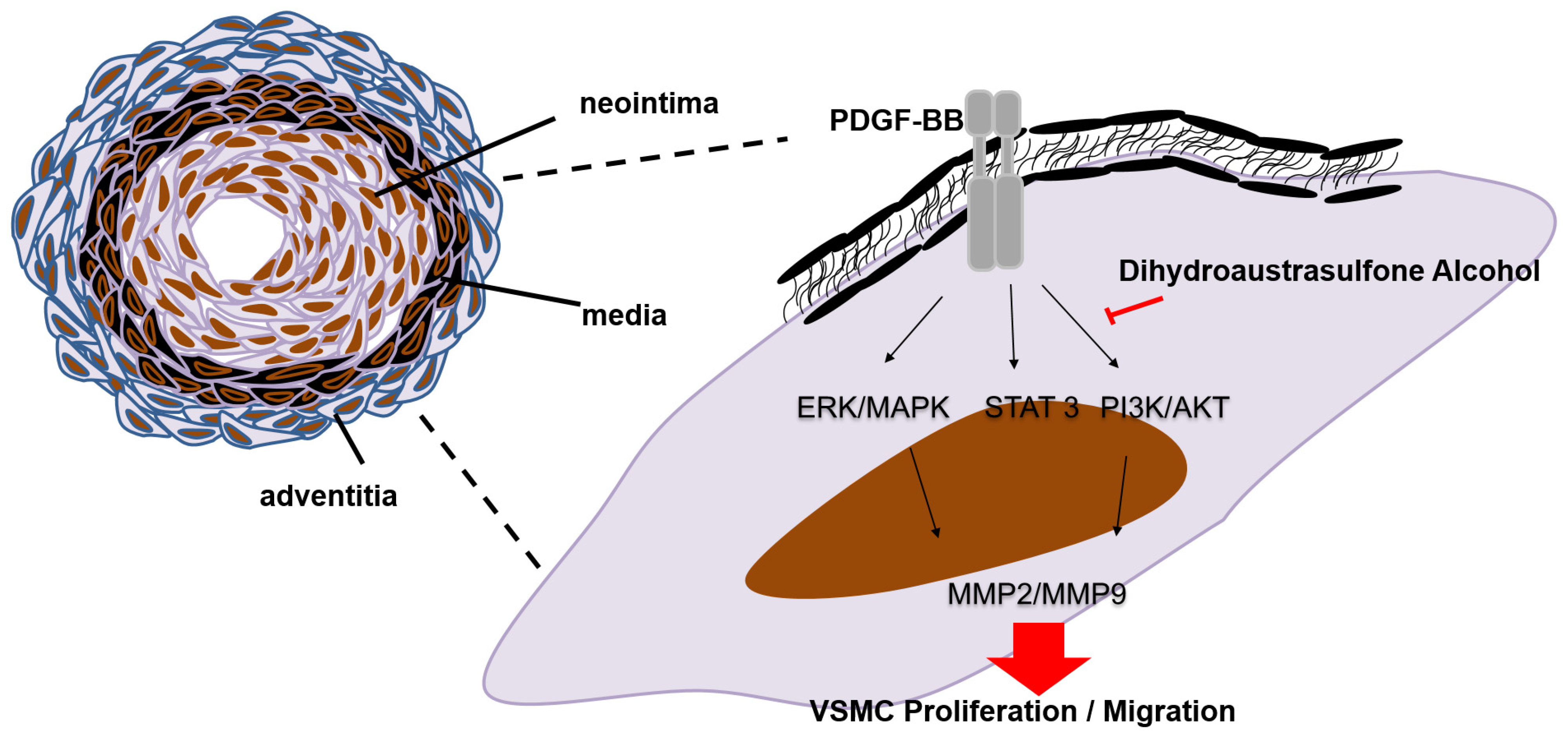
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Weintraub, W.S. The pathophysiology and burden of restenosis. Am. J. Cardiol. 2007, 100, K3–K9. [Google Scholar] [CrossRef]
- Raines, E.W. PDGF and cardiovascular disease. Cytokine Growth Factor Rev. 2004, 15, 237–254. [Google Scholar] [CrossRef] [PubMed]
- Dzau, V.J.; Braun-Dullaeus, R.C.; Sedding, D.G. Vascular proliferation and atherosclerosis: New perspectives and therapeutic strategies. Nat. Med. 2002, 8, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Sachinidis, A.; Locher, R.; Vetter, W.; Tatje, D.; Hoppe, J. Different effects of platelet-derived growth factor isoforms on rat vascular smooth muscle cells. J. Biol. Chem. 1990, 265, 10238–10243. [Google Scholar] [PubMed]
- Chan, C.M.; Fang, J.Y.; Lin, H.H.; Yang, C.Y.; Hung, C.F. Lycopene inhibits PDGF-BB-induced retinal pigment epithelial cell migration by suppression of PI3K/Akt and MAPK pathways. Biochem. Biophys. Res. Commun. 2009, 388, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Vij, N.; Sharma, A.; Thakkar, M.; Sinha, S.; Mohan, R.R. PDGF-driven proliferation, migration, and IL8 chemokine secretion in human corneal fibroblasts involve JAK2-STAT3 signaling pathway. Mol. Vis. 2008, 14, 1020–1027. [Google Scholar] [PubMed]
- Chan, C.M.; Chang, H.H.; Wang, V.C.; Huang, C.L.; Hung, C.F. Inhibitory effects of resveratrol on PDGF-BB-induced retinal pigment epithelial cell migration via PDGFRβ, PI3K/Akt and MAPK pathways. PLoS ONE 2013, 8, e56819. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.C.; Chien, Y.C.; Pan, C.H.; Sheu, J.H.; Chen, C.Y.; Wu, C.H. Inhibitory effect of dihydroaustrasulfone alcohol on the migration of human non-small cell lung carcinoma A549 cells and the antitumor effect on a Lewis lung carcinoma-bearing tumor model in C57BL/6J mice. Mar. Drugs 2014, 12, 196–213. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.H.; Chao, C.H.; Wu, M.H.; Sheu, J.H. A neuroprotective sulfone of marine origin and the in vivo anti-inflammatory activity of an analogue. Eur. J. Med. Chem. 2010, 45, 5998–6004. [Google Scholar] [CrossRef] [PubMed]
- Li, P.C.; Pan, C.H.; Sheu, M.J.; Wu, C.C.; Ma, W.F.; Wu, C.H. Deep sea water prevents balloon angioplasty-induced hyperplasia through MMP-2: An in vitro and in vivo study. PLoS ONE 2014, 9, e96927. [Google Scholar] [CrossRef] [PubMed]
- Jean, Y.H.; Chen, W.F.; Sung, C.S.; Duh, C.Y.; Huang, S.Y.; Lin, C.S.; Tai, M.H.; Tzeng, S.F.; Wen, Z.H. Capnellene, a natural marine compound derived from soft coral, attenuates chronic constriction injury-induced neuropathic pain in rats. Br. J. Pharmacol. 2009, 158, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Jean, Y.H.; Chen, W.F.; Duh, C.Y.; Huang, S.Y.; Hsu, C.H.; Lin, C.S.; Sung, C.S.; Chen, I.M.; Wen, Z.H. Inducible nitric oxide synthase and cyclooxygenase-2 participate in anti-inflammatory and analgesic effects of the natural marine compound lemnalol from Formosan soft coral Lemnalia cervicorni. Eur. J. Pharmacol. 2008, 578, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.J.; Wen, Z.H.; Dai, C.F.; Chiang, M.Y.; Sheu, J.H. Nanolobatolide, a new C18 metabolite from the Formosan soft coral Sinularia nanolobata. Org. Lett. 2009, 11, 5030–5032. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.H.; Wen, Z.H.; Wu, Y.C.; Yeh, H.C.; Sheu, J.H. Cytotoxic and anti-inflammatory cembranoids from the soft coral Lobophytum crassum. J. Nat. Prod. 2008, 71, 1819–1824. [Google Scholar] [CrossRef] [PubMed]
- Newby, A.C. An overview of the vascular response to injury: A tribute to the late Russell Ross. Toxicol. Lett. 2000, 112–113, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Heldin, C.H.; Westermark, B. Mechanism of action and in vivo role of platelet-derived growth factor. Phys. Rev. 1999, 79, 1283–1316. [Google Scholar]
- Lee, K.P.; Lee, K.; Park, W.H.; Kim, H.; Hong, H. Piperine inhibits platelet-derived growth factor-BB-induced proliferation and migration in vascular smooth muscle cells. J. Med. Food 2015, 18, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Indolfi, C.; Coppola, C.; Torella, D.; Arcucci, O.; Chiariello, M. Gene therapy for restenosis after balloon angioplasty and stenting. Cardiol. Rev. 1999, 7, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.; Wang, H.; Lee, W.S.; Jain, M.K.; Lee, M.E.; Haber, E. Mitogen-activated protein kinase phosphatase-1 in rat arterial smooth muscle cell proliferation. J. Clin. Investig. 1996, 98, 1560–1567. [Google Scholar] [CrossRef] [PubMed]
- Gennaro, G.; Menard, C.; Michaud, S.E.; Deblois, D.; Rivard, A. Inhibition of vascular smooth muscle cell proliferation and neointimal formation in injured arteries by a novel, oral mitogen-activated protein kinase/extracellular signal-regulated kinase inhibitor. Circulation 2004, 110, 3367–3371. [Google Scholar] [CrossRef] [PubMed]
- Daniel, J.M.; Dutzmann, J.; Bielenberg, W.; Widmer-Teske, R.; Gunduz, D.; Hamm, C.W.; Sedding, D.G. Inhibition of STAT3 signaling prevents vascular smooth muscle cell proliferation and neointima formation. Basic. Res. Cardiol. 2012, 107, 261. [Google Scholar] [CrossRef] [PubMed]
- Shibata, R.; Kai, H.; Seki, Y.; Kato, S.; Wada, Y.; Hanakawa, Y.; Hashimoto, K.; Yoshimura, A.; Imaizumi, T. Inhibition of STAT3 prevents neointima formation by inhibiting proliferation and promoting apoptosis of neointimal smooth muscle cells. Hum. Gene Ther. 2003, 14, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Grote, K.; Luchtefeld, M.; Schieffer, B. JANUS under stress—Role of JAK/STAT signaling pathway in vascular diseases. Vascul. Pharmacol. 2005, 43, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Kai, H.; Shibata, R.; Nagata, T.; Yasukawa, H.; Yoshimura, A.; Imaizumi, T. Role of the JAK/STAT pathway in rat carotid artery remodeling after vascular injury. Circ. Res. 2000, 87, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Sasaguri, Y.; Murahashi, N.; Sugama, K.; Kato, S.; Hiraoka, K.; Satoh, T.; Isomoto, H.; Morimatsu, M. Development-related changes in matrix metalloproteinase expression in human aortic smooth muscle cells. Lab. Investig. 1994, 71, 261–269. [Google Scholar] [PubMed]
- Bendeck, M.P.; Conte, M.; Zhang, M.; Nili, N.; Strauss, B.H.; Farwell, S.M. Doxycycline modulates smooth muscle cell growth, migration, and matrix remodeling after arterial injury. Am. J. Surg. Pathol. 2002, 160, 1089–1095. [Google Scholar] [CrossRef]
- Cho, A.; Reidy, M.A. Matrix metalloproteinase-9 is necessary for the regulation of smooth muscle cell replication and migration after arterial injury. Circ. Res. 2002, 91, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.; Galis, Z.S. Matrix metalloproteinase-2 and -9 differentially regulate smooth muscle cell migration and cell-mediated collagen organization. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yang, J.; Lu, F.; Xu, D.; Zhou, L.; Shi, A.; Cao, K. Platelet-derived growth factor BB modulates PCNA protein synthesis partially through the transforming growth factor β signalling pathway in vascular smooth muscle cells. Biochem. Cell Biol. 2007, 85, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Stadius, M.L.; Gown, A.M.; Kernoff, R.; Collins, C.L. Cell proliferation after balloon injury of iliac arteries in the cholesterol-fed New Zealand White rabbit. Arterioscler. Thromb. Vasc. Biol. 1994, 14, 727–733. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.-C.; Sheu, M.-J.; Ma, W.-F.; Pan, C.-H.; Sheu, J.-H.; Wu, C.-H. Anti-Restenotic Roles of Dihydroaustrasulfone Alcohol Involved in Inhibiting PDGF-BB-Stimulated Proliferation and Migration of Vascular Smooth Muscle Cells. Mar. Drugs 2015, 13, 3046-3060. https://doi.org/10.3390/md13053046
Li P-C, Sheu M-J, Ma W-F, Pan C-H, Sheu J-H, Wu C-H. Anti-Restenotic Roles of Dihydroaustrasulfone Alcohol Involved in Inhibiting PDGF-BB-Stimulated Proliferation and Migration of Vascular Smooth Muscle Cells. Marine Drugs. 2015; 13(5):3046-3060. https://doi.org/10.3390/md13053046
Chicago/Turabian StyleLi, Pei-Chuan, Ming-Jyh Sheu, Wei-Fen Ma, Chun-Hsu Pan, Jyh-Horng Sheu, and Chieh-Hsi Wu. 2015. "Anti-Restenotic Roles of Dihydroaustrasulfone Alcohol Involved in Inhibiting PDGF-BB-Stimulated Proliferation and Migration of Vascular Smooth Muscle Cells" Marine Drugs 13, no. 5: 3046-3060. https://doi.org/10.3390/md13053046