Site-Specific Variability in the Chemical Diversity of the Antarctic Red Alga Plocamium cartilagineum


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
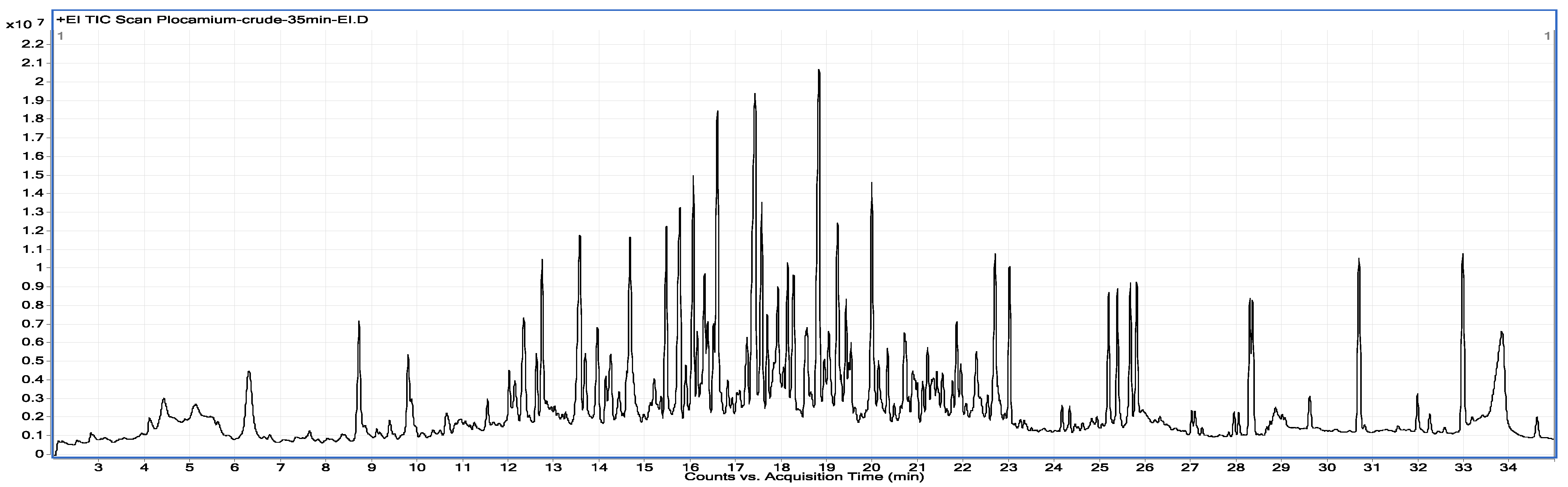
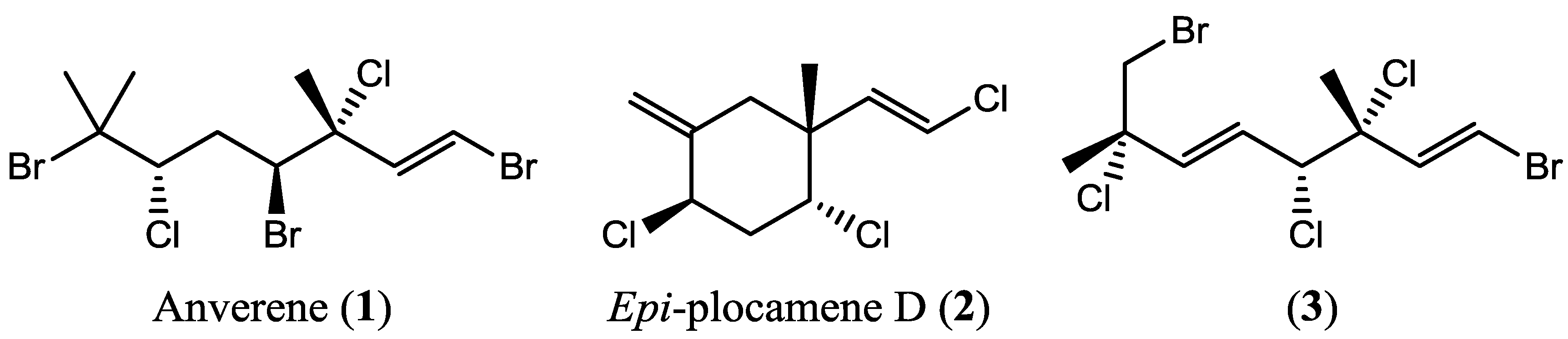
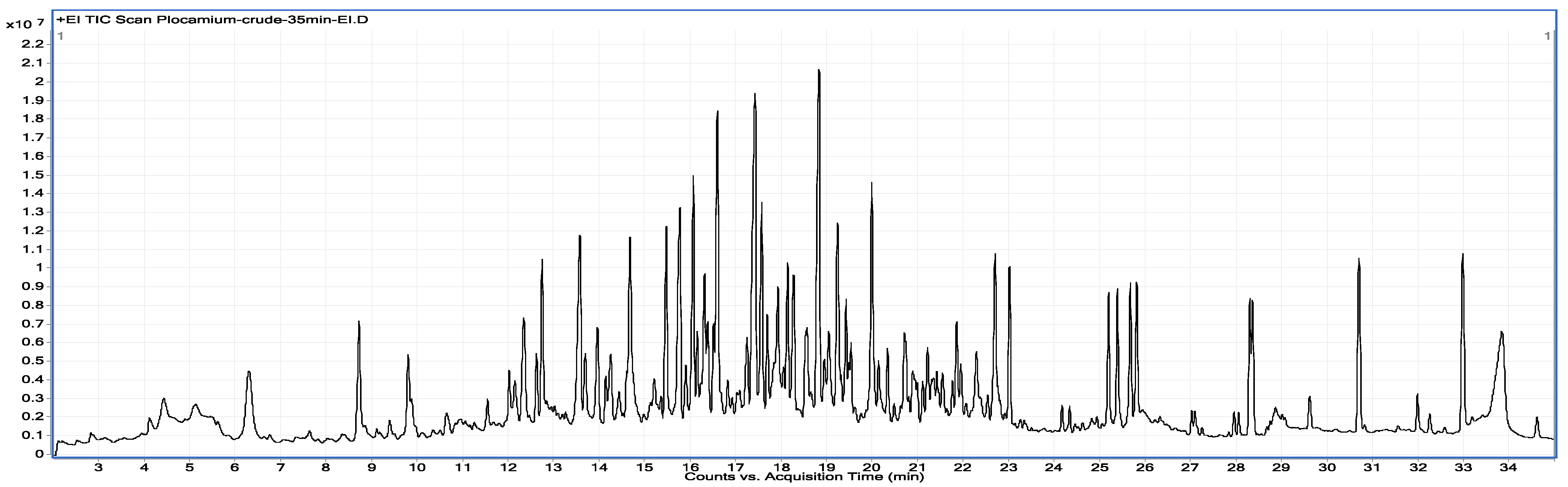
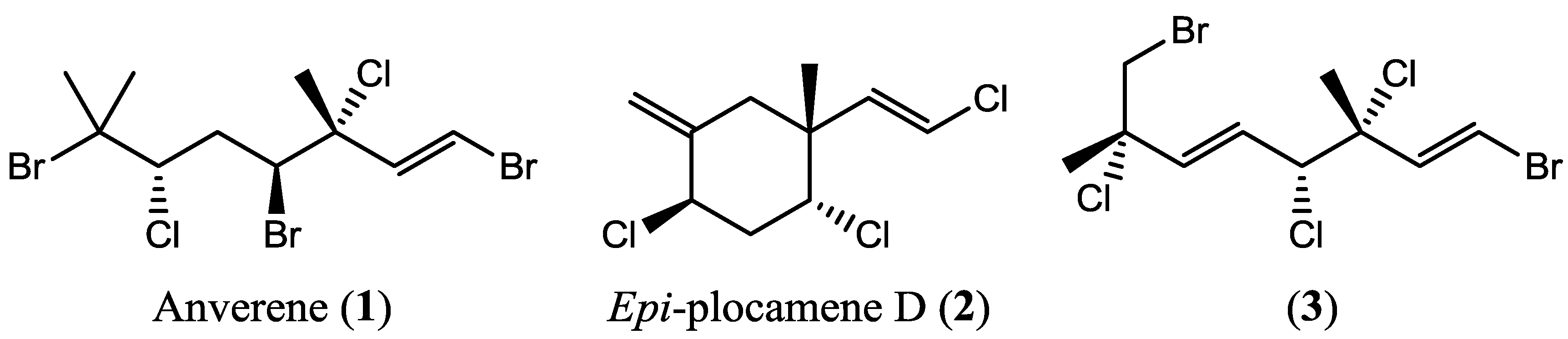
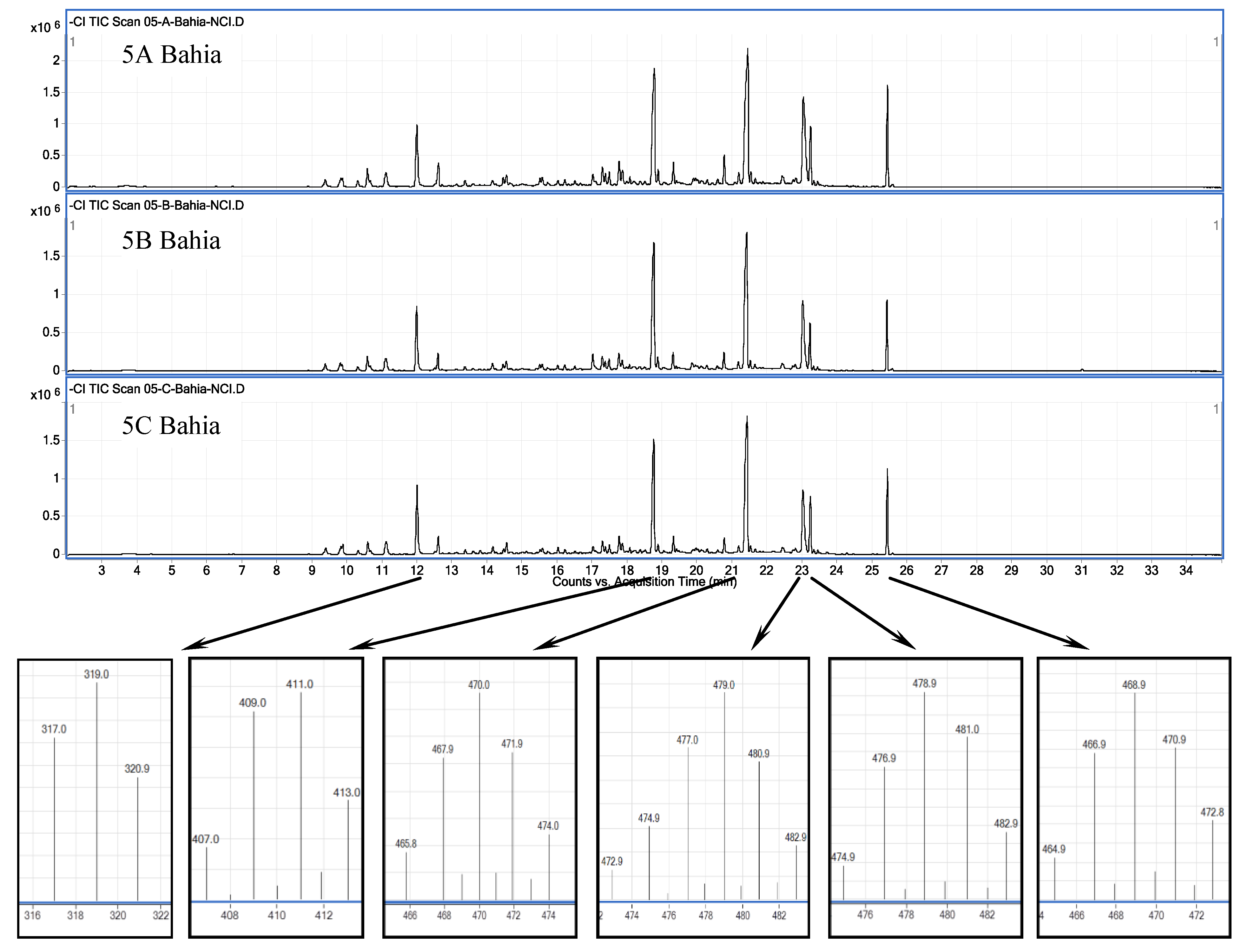
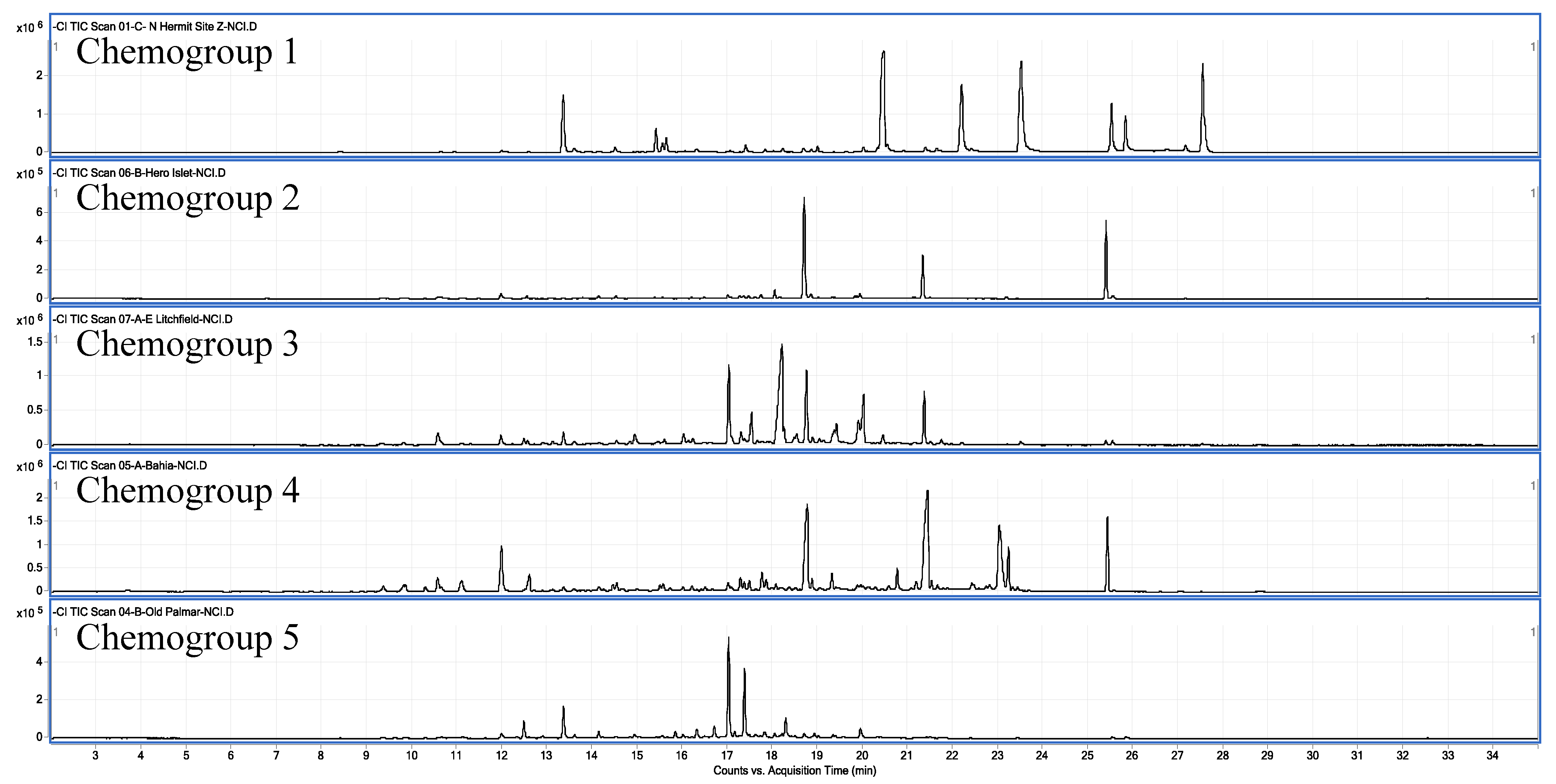
2.1. Chemical Analysis


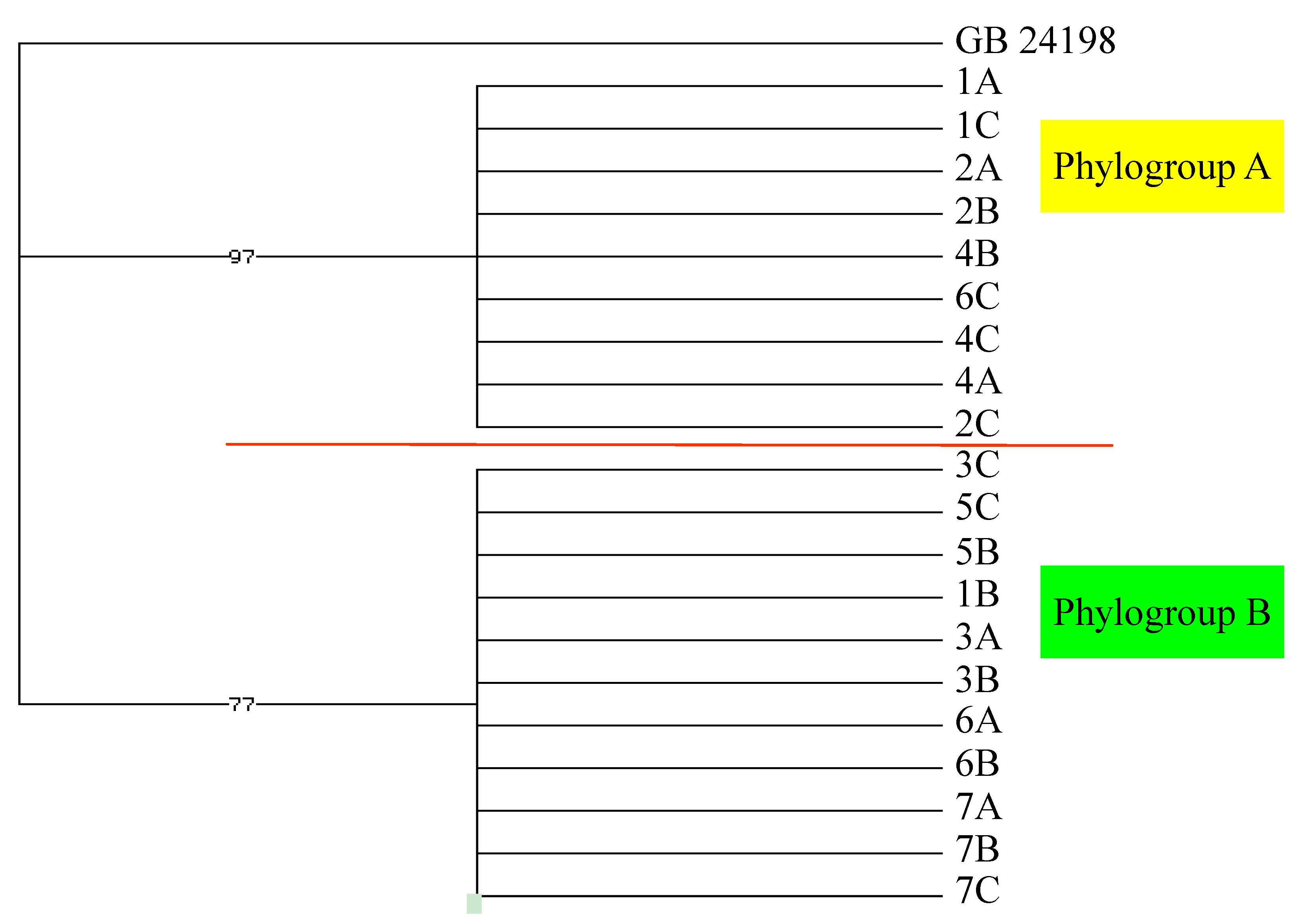
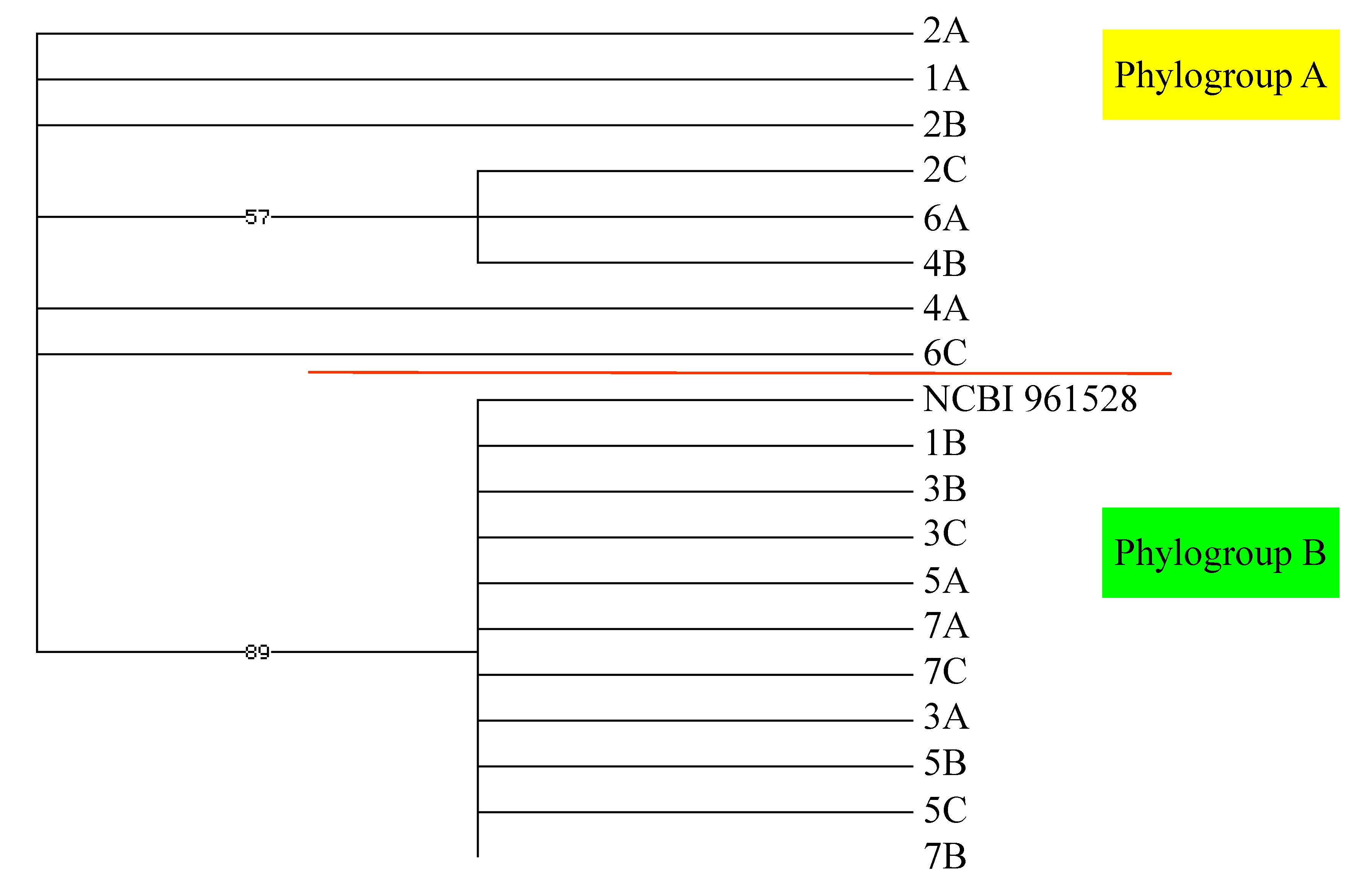
2.2. Genetic Analysis

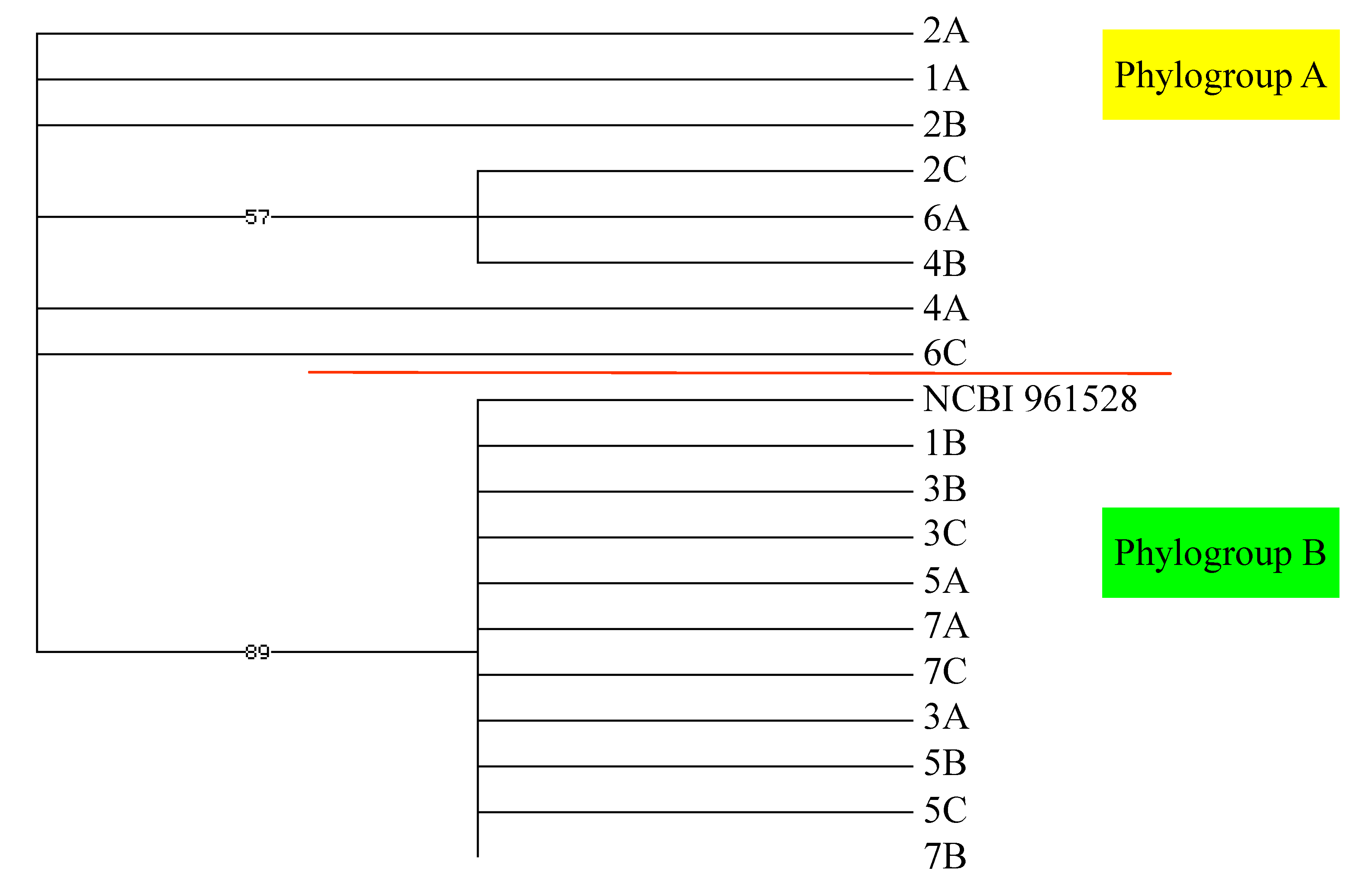
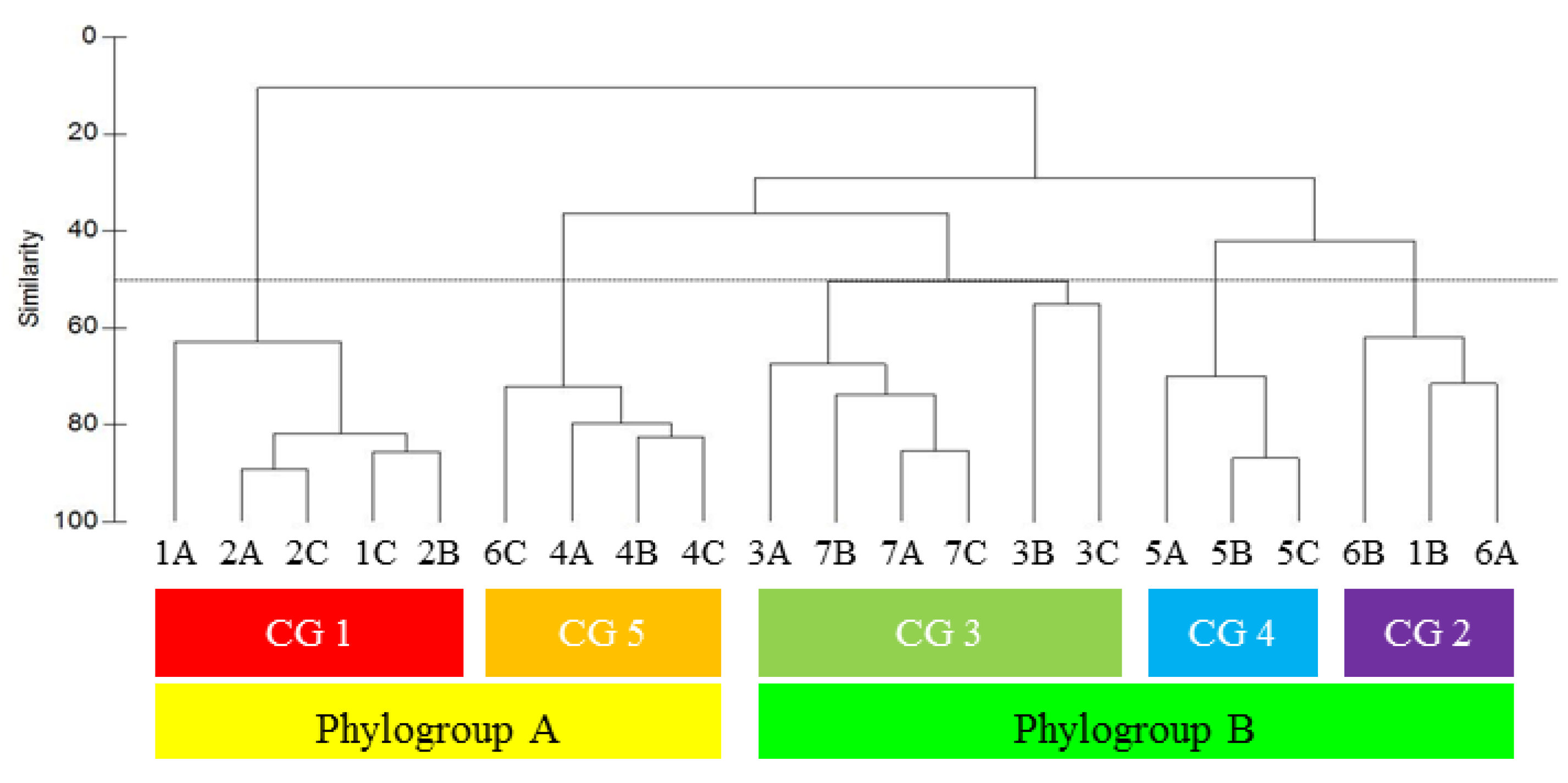
2.3. Metabogenetic Characterization of Plocamium cartilagineum

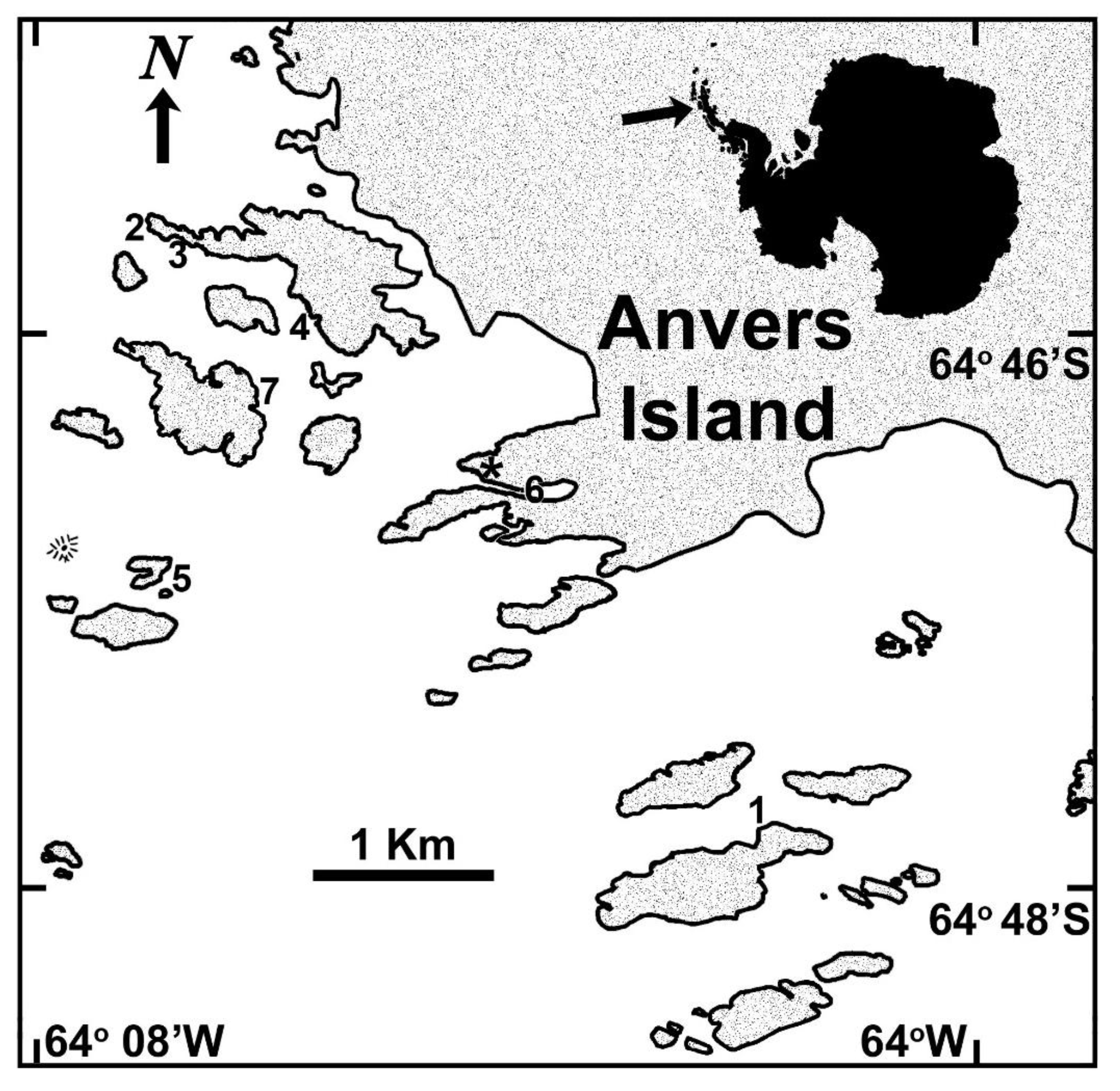
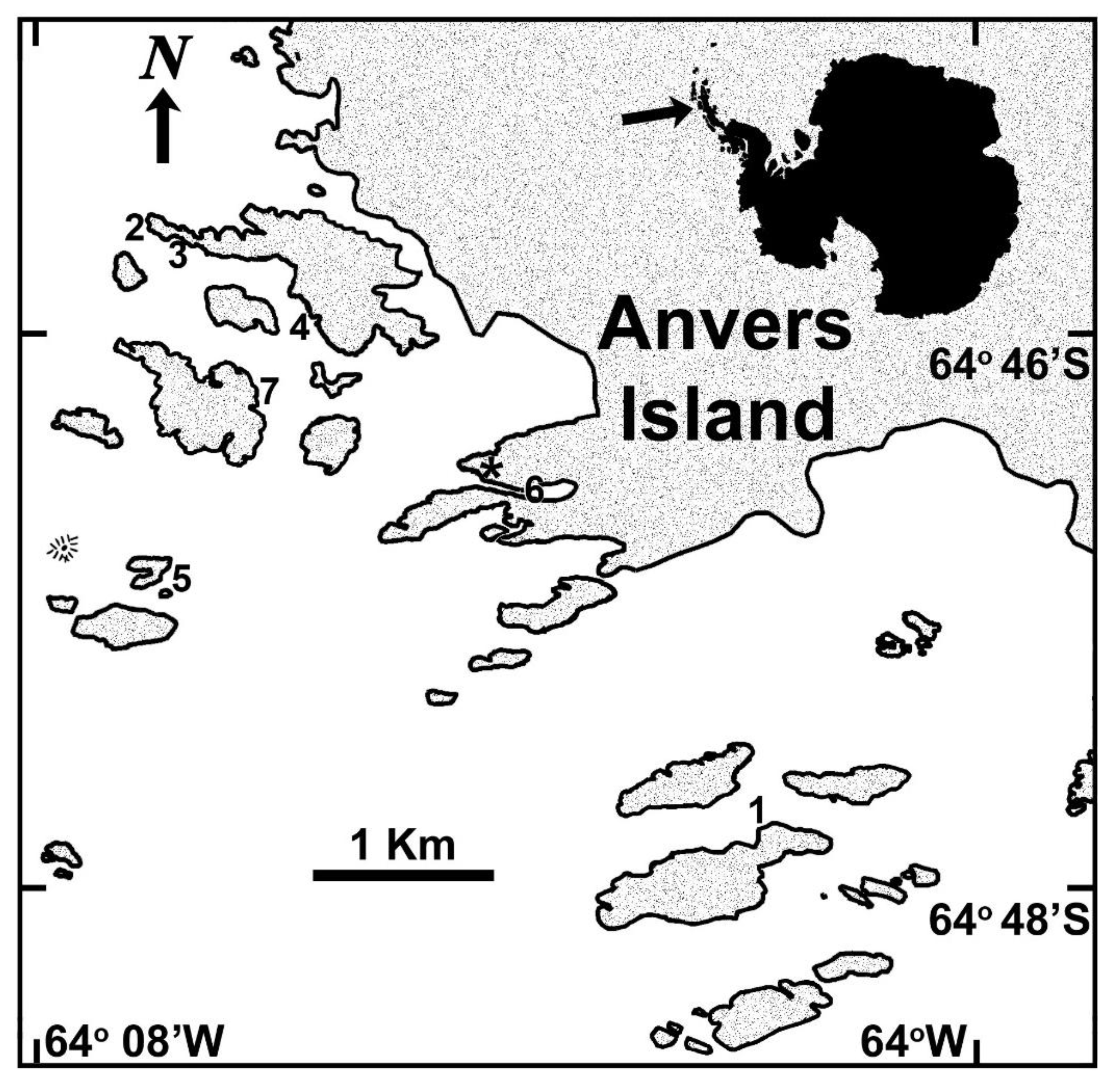
2.4. Site Specificity among Plocamium cartilagineum Metabogenetic Groups
3. Experimental Section
3.1. Biological Material
3.2. Chemical Analysis
3.2.1. Algal Extraction and Monoterpene Purification
3.2.2. GC/MS Analysis
3.3. Phylogenetic Analysis
3.4. Metabolomic Data Processing and Multivariate Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- AlgaeBase. Available online: http://www.algaebase.org (accessed on 12 January 2013).
- Saunders, G.W.; Lehmkuhl, K.V. Molecular divergence and morphological diversity among four cryptic species of Plocamium (Plocamiales, Florideophyceae) in northern Europe. Eur. J. Phycol. 2005, 40, 293–312. [Google Scholar] [CrossRef]
- MarinLit, version 2013; University of Canterbury: Christchurch, New Zealand, 2013.
- Kladi, M.; Vagias, C.; Roussis, V. Volatile halogenated metabolites from marine red algae. Phytochem. Rev. 2004, 3, 337–366. [Google Scholar] [CrossRef]
- Neushul, M. Benthic marine algae. In Antarctic Map Folio Series 10; Balech, E., El-Sayed, S.Z., Hasle, G., Neushul, M., Zaneveld, J.S., Eds.; American Geographical Society: New York, NY, USA, 1968; pp. 14–15. [Google Scholar]
- Wiencke, C.; Clayton, M.H. Antarctic Seaweeds: Synopsis of the Antarctic Benthos; Gantner, A.R.G., Ruggell, K.G., Eds.; Springer-Verlag: Berlin, Germany, 2002; Volume 9. [Google Scholar]
- Neushul, M. Diving observation of sub-tidal Antarctic marine vegetation. Bot. Mar. 1965, 8, 234–243. [Google Scholar] [CrossRef]
- Moe, R.L.; DeLaca, T.E. Occurrence of macroscopic algae along the Antarctic Peninsula. Antarctic J. 1976, 11, 20–24. [Google Scholar]
- Lamb, I.M.; Zimmerman, M.H. Benthic marine algae of the Antarctic Peninsula. In Biology of the Antarctic Seas V; American Geophysical Union: Washington, DC, USA, 1977; Volume 23, pp. 130–229. [Google Scholar]
- Hommersand, M.H.; Moe, R.L.; Amsler, C.D.; Fredericq, S. Notes on the systematics of Antarctic and sub-Antarctic Rhodophyta with descriptions of four new genera and five new species. Bot. Mar. 2009, 52, 509–534. [Google Scholar]
- Amsler, C.D.; Iken, K.B.; McClintock, J.B.; Amsler, M.O.; Peters, K.J.; Hubbard, J.M.; Furrow, F.B.; Baker, B.J. Comprehensive evaluation of the palatability and chemical defenses of subtidal macroalgae from the Antarctic Peninsula. Mar. Ecol. Prog. Ser. 2005, 294, 141–159. [Google Scholar] [CrossRef]
- Ankisetty, S.; Nandiraju, S.; Win, H.; Park, Y.-C.; Amsler, C.D.; McClintock, J.B.; Baker, J.A.; Diyabalanage, T.; Pasaribu, A.; Singh, M.P.; et al. Chemical investigations of predator deterred macroalgae from the Antarctic Peninsula. J. Nat. Prod. 2004, 67, 1295–1302. [Google Scholar] [CrossRef]
- Zamzow, J.P.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Habitat choice and predator avoidance by Antarctic amphipods: The roles of algal chemistry and morphology. Mar. Ecol. Prog. Ser. 2010, 400, 155–163. [Google Scholar] [CrossRef]
- Aumack, C.F.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Chemically mediated resistance to mesoherbivory in finely branched macroalgae along the western Antarctic Peninsula. Eur. J. Phycol. 2010, 45, 19–26. [Google Scholar] [CrossRef]
- Aumack, C.F.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Impacts of mesograzers on epiphyte and endophyte growth associated with chemically defended macroalgae from the western Antarctic Peninsula: A mesocosm experiment. J. Phycol. 2011, 47, 36–41. [Google Scholar] [CrossRef]
- Stierle, D.B.; Sims, J.J. Polyhalogenated cyclic monoterpenes from the red alga Plocamium cartilagineum of Antarctica. Tetrahedron 1979, 35, 1261–1265. [Google Scholar] [CrossRef]
- Dougherty, R.C.; Dalton, J.; Biros, F.J. Negative chemical ionization mass spectra of polycyclic chlorinated insecticides. J. Mass Spectrom. 1972, 11, 1171–1181. [Google Scholar]
- ezanka, T.; Dembitsky, V.M. Poly halogenated homosesquiterpenic fatty acids from Plocamium cartilagineum. Phytochemistry 2001, 57, 607–611. [Google Scholar]
- Gurgel, F.; Fredericq, S. Systematics of the Gracilariaceae (Gracilariales, Rhodophyta): A critical assessment based on rbcL sequence analysis. J. Phycol. 2004, 40, 138–159. [Google Scholar] [CrossRef]
- Robba, L.; Russell, S.; Barker, G.; Brodie, J. Assessing the use of the mitochondrial cox1 marker for use in DNA barcoding of red algae (Rhodophyta). Am. J. Bot. 2006, 93, 1101–1108. [Google Scholar] [CrossRef]
- Yang, E.; Kim, M.; Geraldino, P.; Sahoo, D.; Shin, J.; Boo, S.M. Mitochondrial cox1 and plastid rbcL genes of Gracilaria vermiculophylla (Gracilariaceae, Rhodophyta). J. Appl. Phycol. 2008, 20, 161–168. [Google Scholar] [CrossRef]
- Payo, D.A.; Colo, J.; Calumpong, H.; de Clerck, O. Variability of non-polar secondary metabolites in the red alga Portieria. Mar. Drugs 2011, 9, 2438–2468. [Google Scholar] [CrossRef] [Green Version]
- Puglisi, M.P.; Paul, V.J. Intraspecific variation in the red alga Portieria hornemannii: Monoterpene concentrations are not influenced by nitrogen or phosphorus enrichment. Mar. Biol. 1997, 128, 161–170. [Google Scholar] [CrossRef]
- Matlock, D.B.; Ginsburg, D.W.; Paul, V.J. Spatial variability in secondary metabolite production by the tropical red alga Portieria hornemannii. Hydrobiologia 1999, 399, 267–273. [Google Scholar]
- Palma, R.; Edding, M.; Rovirosa, J.; San-Martín, A.; Argandoña, V.H. Effect of photon flux density and temperature on the production of halogenated monoterpenes by Plocamium cartilagineum (Plocamiaceae, Rhodophyta). Z. Naturforsch. C 2004, 59, 679–683. [Google Scholar]
- Wiencke, C.; Amsler, C.D. Seaweeds and Their Communities in Polar Regions. In Seaweed Biology: Novel Insights into Ecophysiology, Ecology and Utilization; Wiencke, C., Bischof, K., Eds.; Springer-Verlag: Berlin, Germany, 2012; pp. 265–294. [Google Scholar]
- Aumack, C.F.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Changes in amphipod densities among macroalgal habitats in day versus night collections along the Western Antarctic Peninsula. Mar. Biol. 2011, 158, 1879–1885. [Google Scholar] [CrossRef]
- Huang, Y.M.; Amsler, M.O.; McClintock, J.B.; Amsler, C.D.; Baker, B.J. Patterns of gammarid amphipod abundance and species composition associated with dominant subtidal macroalgae along the western Antarctic Peninsula. Polar Biol. 2007, 30, 1417–1430. [Google Scholar] [CrossRef]
- Wiencke, C.; Clayton, M.N.; Gómez, I.; Iken, K.; Lüder, U.H.; Amsler, C.D.; Karsten, U.; Hanelt, D.; Bischof, K.; Dunton, K. Life strategy, ecophysiology and ecology of seaweeds in polar waters. Rev. Environ. Sci. Biotech. 2007, 6, 95–126. [Google Scholar] [CrossRef] [Green Version]
- Vergés, A.; Paul, N.A.; Steinberg, P.D. Sex and life-history stage alter herbivore responses to a chemically defended red alga. Ecology 2008, 89, 1334–1343. [Google Scholar] [CrossRef] [Green Version]
- Saunders, G.W.; McDevit, D.C. Methods for DNA Barcoding Photosynthetic Protists Emphasizing the Macroalgae and Diatoms. In DNA Barcodes: Methods and Protocols; Kress, W.J., Erickson, D.L., Eds.; Springer Science + Business Media: New York, NY, USA, 2012; pp. 207–222. [Google Scholar]
- Freshwater, D.W.; Rueness, J. Phylogenetic relationships of some European Gelidium (Gelidiales, Rhodophyta) species, based on rbcL nucleotide sequence analysis. Phycologia 1994, 33, 187–194. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8.
- Zwickl, D.J. Genetic Algorithm Approaches for the Phylogenetic Analysis of Large Biological Sequence Datasets under the Maximum Likelihood Criterion. Ph.D. Thesis, The University of Texas at Austin, Austin, TX, USA, 2006. [Google Scholar]
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Young, R.M.; Von Salm, J.L.; Amsler, M.O.; Lopez-Bautista, J.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Site-Specific Variability in the Chemical Diversity of the Antarctic Red Alga Plocamium cartilagineum. Mar. Drugs 2013, 11, 2126-2139. https://doi.org/10.3390/md11062126
Young RM, Von Salm JL, Amsler MO, Lopez-Bautista J, Amsler CD, McClintock JB, Baker BJ. Site-Specific Variability in the Chemical Diversity of the Antarctic Red Alga Plocamium cartilagineum. Marine Drugs. 2013; 11(6):2126-2139. https://doi.org/10.3390/md11062126
Chicago/Turabian StyleYoung, Ryan M., Jacqueline L. Von Salm, Margaret O. Amsler, Juan Lopez-Bautista, Charles D. Amsler, James B. McClintock, and Bill J. Baker. 2013. "Site-Specific Variability in the Chemical Diversity of the Antarctic Red Alga Plocamium cartilagineum" Marine Drugs 11, no. 6: 2126-2139. https://doi.org/10.3390/md11062126