
High Megabenthic Complexity and Vulnerability of a Mesophotic Rocky Shoal Support Its Inclusion in a Mediterranean MPA

 ,
,  , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
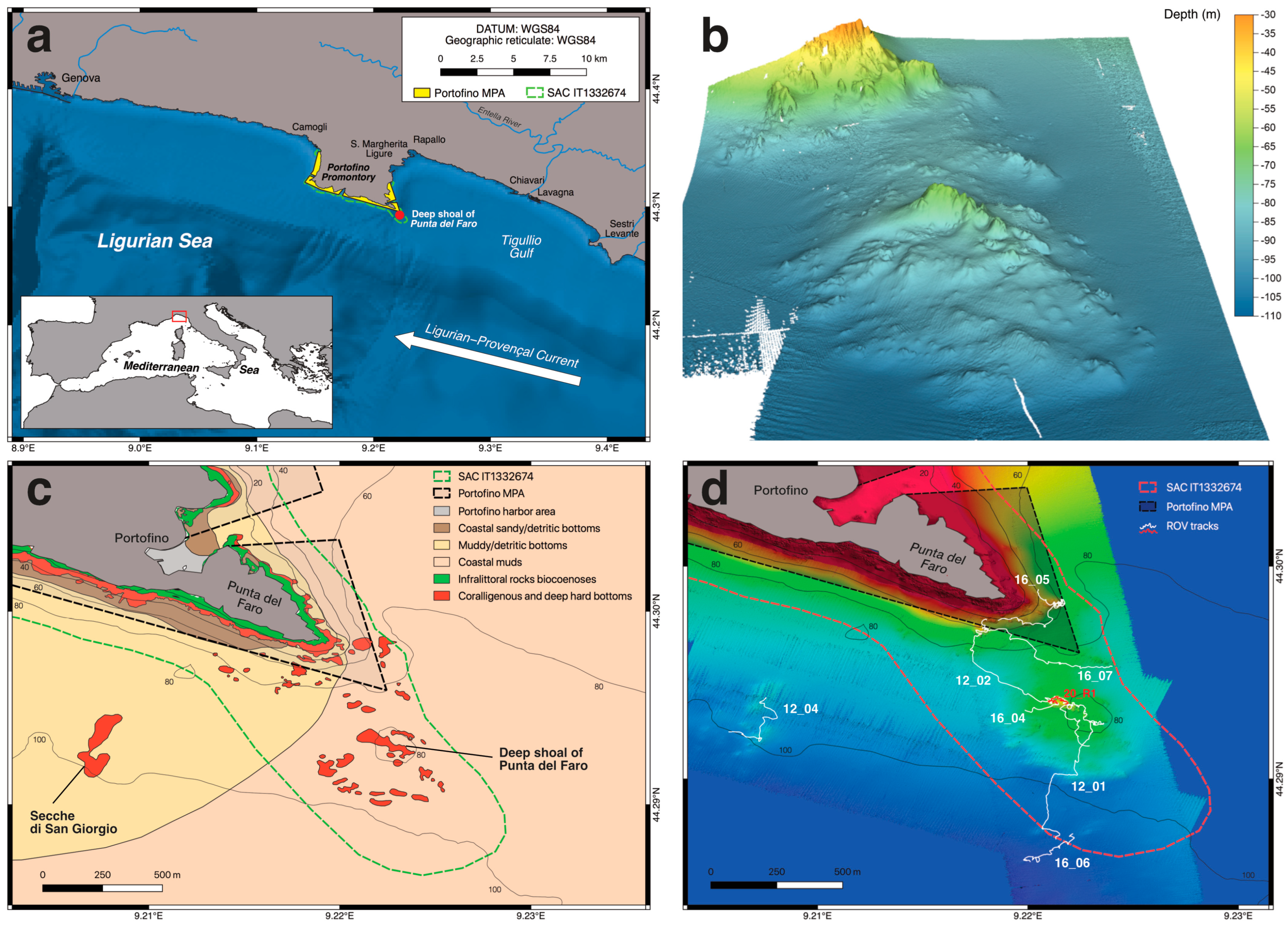
2. Materials and Methods
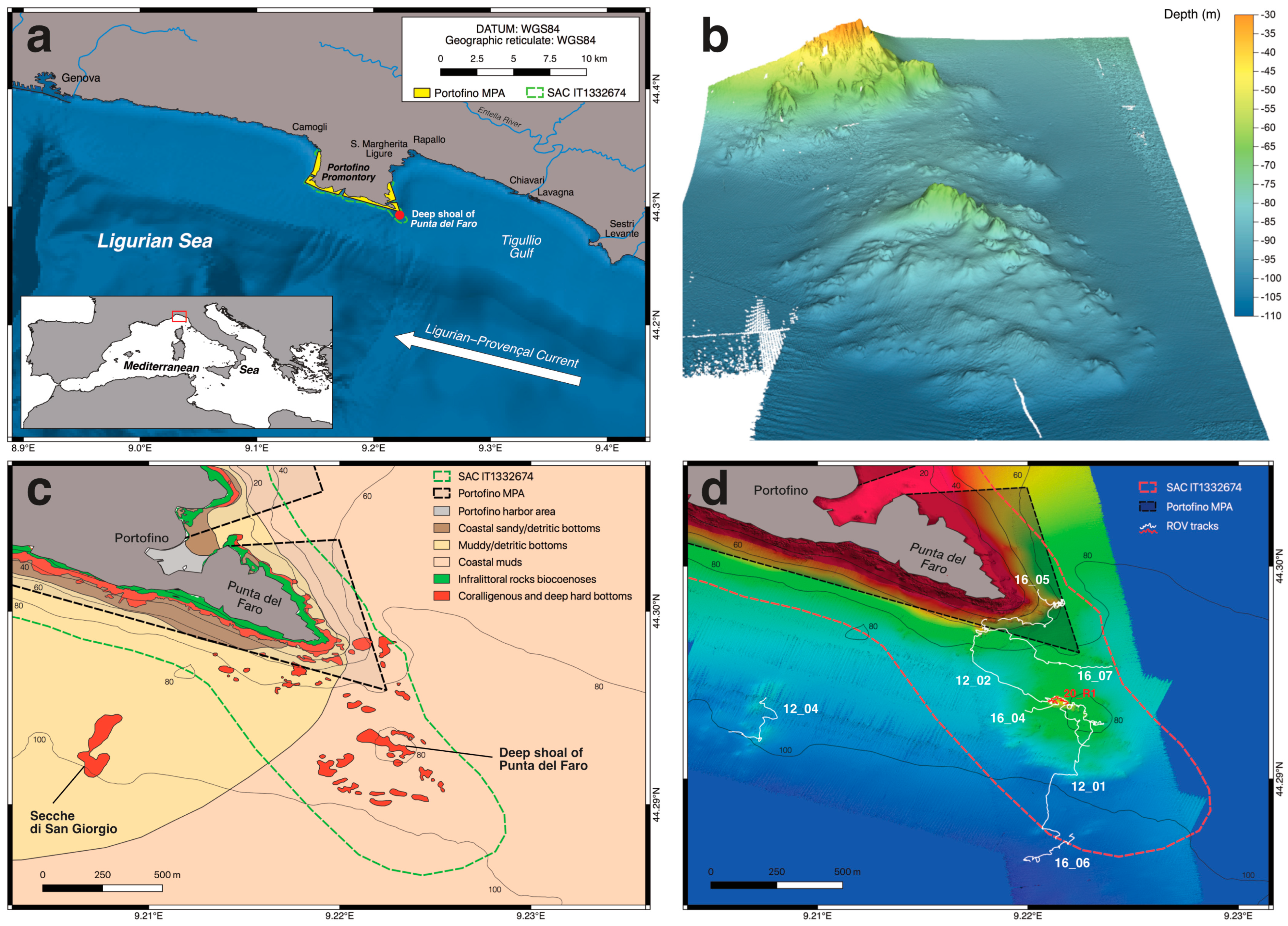
2.1. Study Area
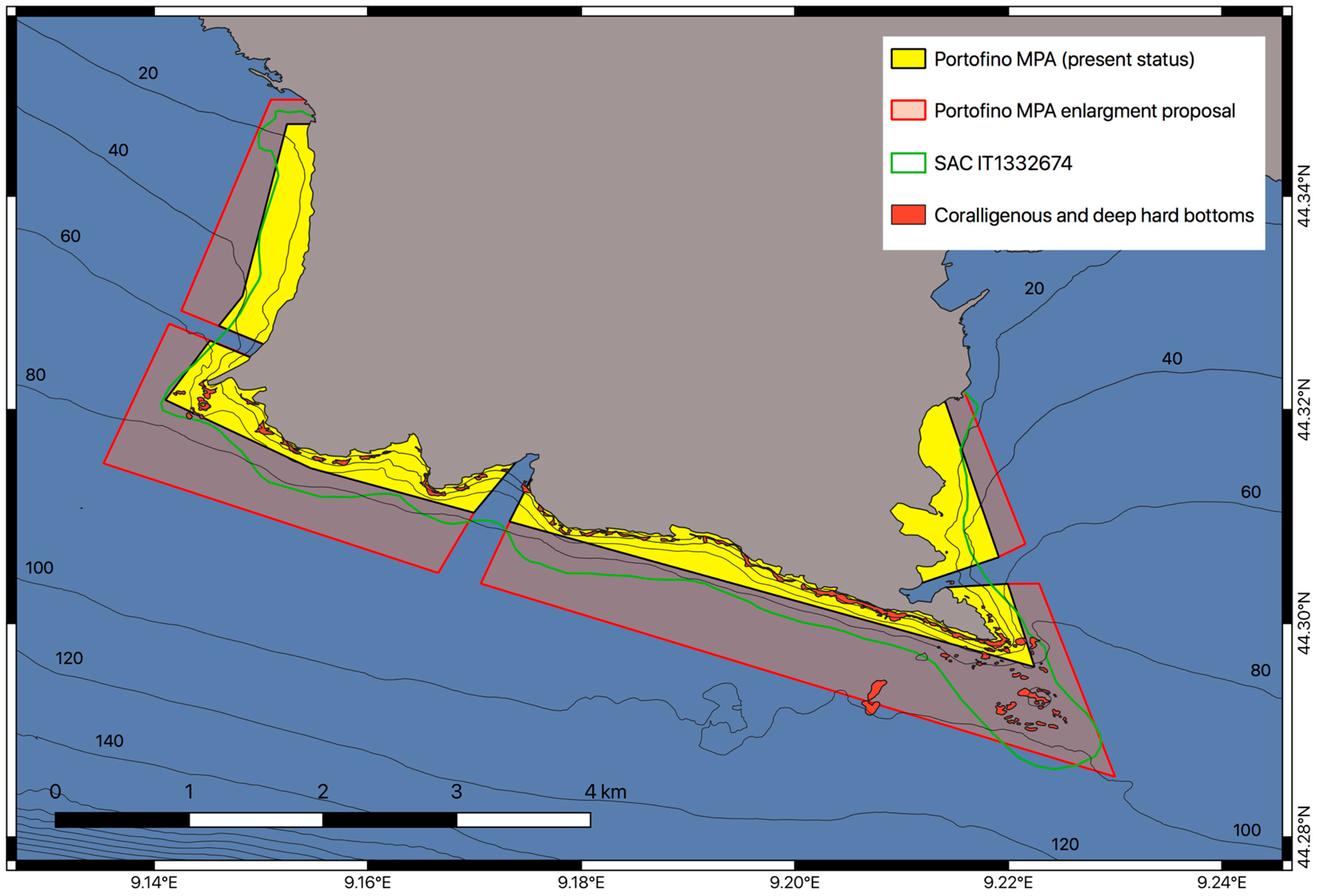
2.2. Legal Framework
2.3. Fishing Fleet
2.4. Biocoenoses Characterization and Health-Status Assessment
2.5. Habitat Modeling
2.6. Fishing-Pressure Quantification
3. Results
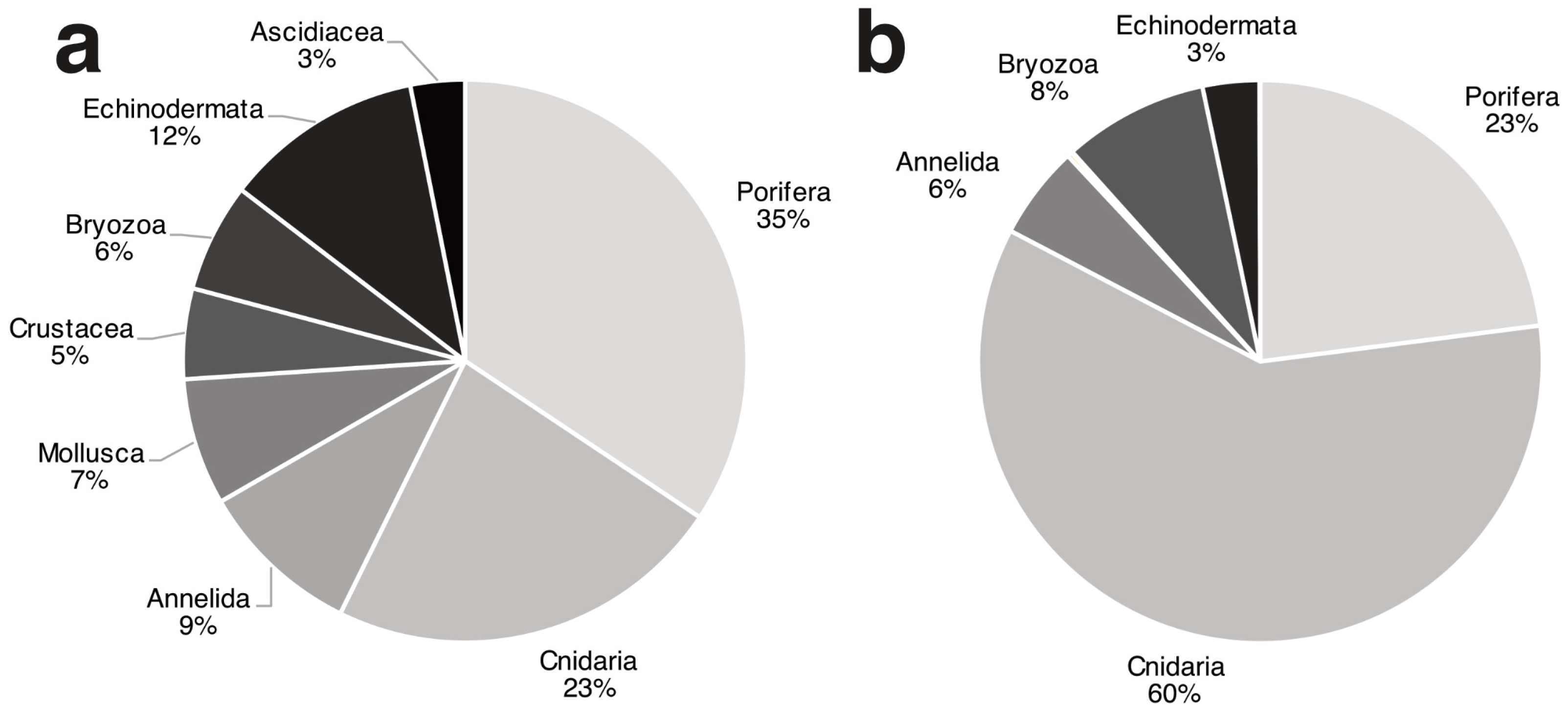
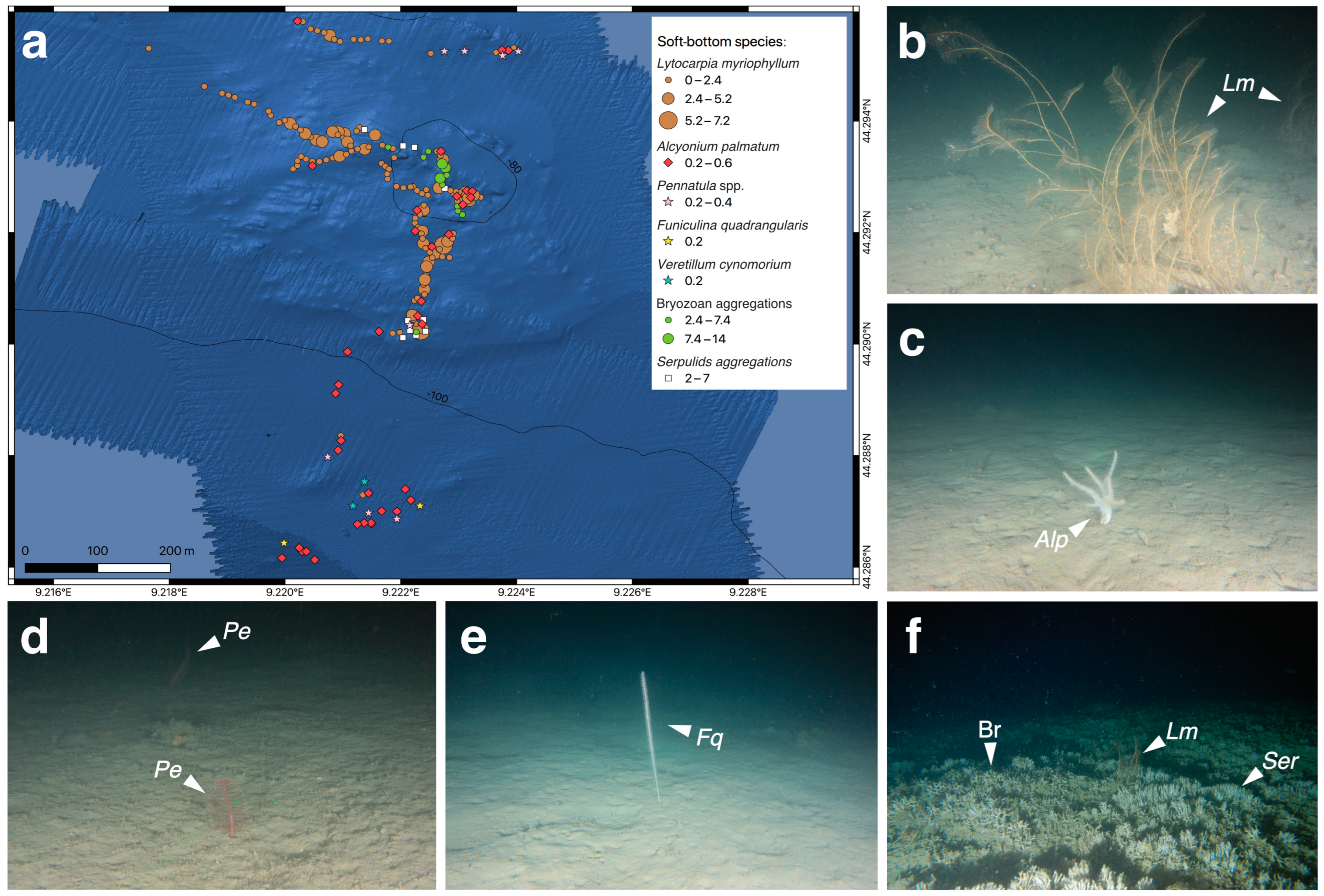
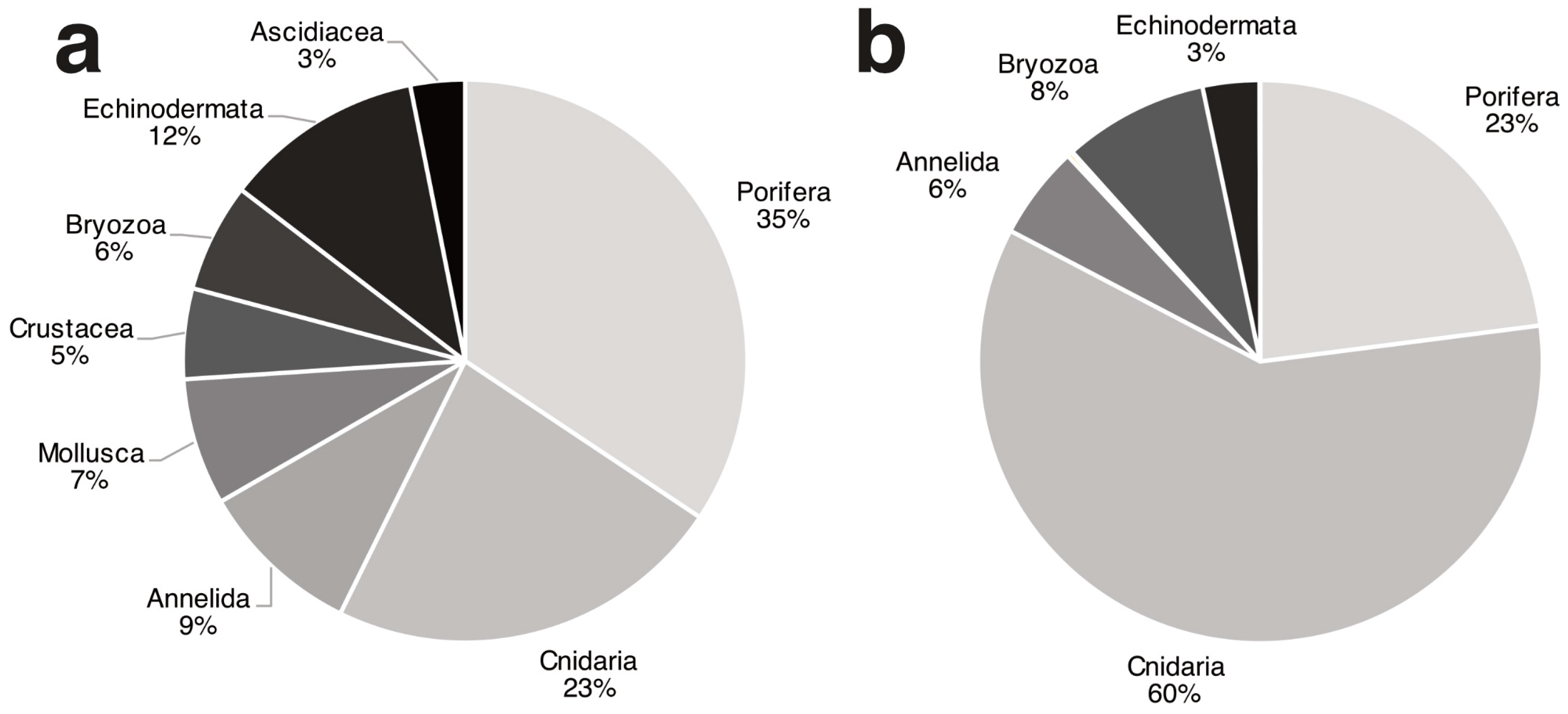
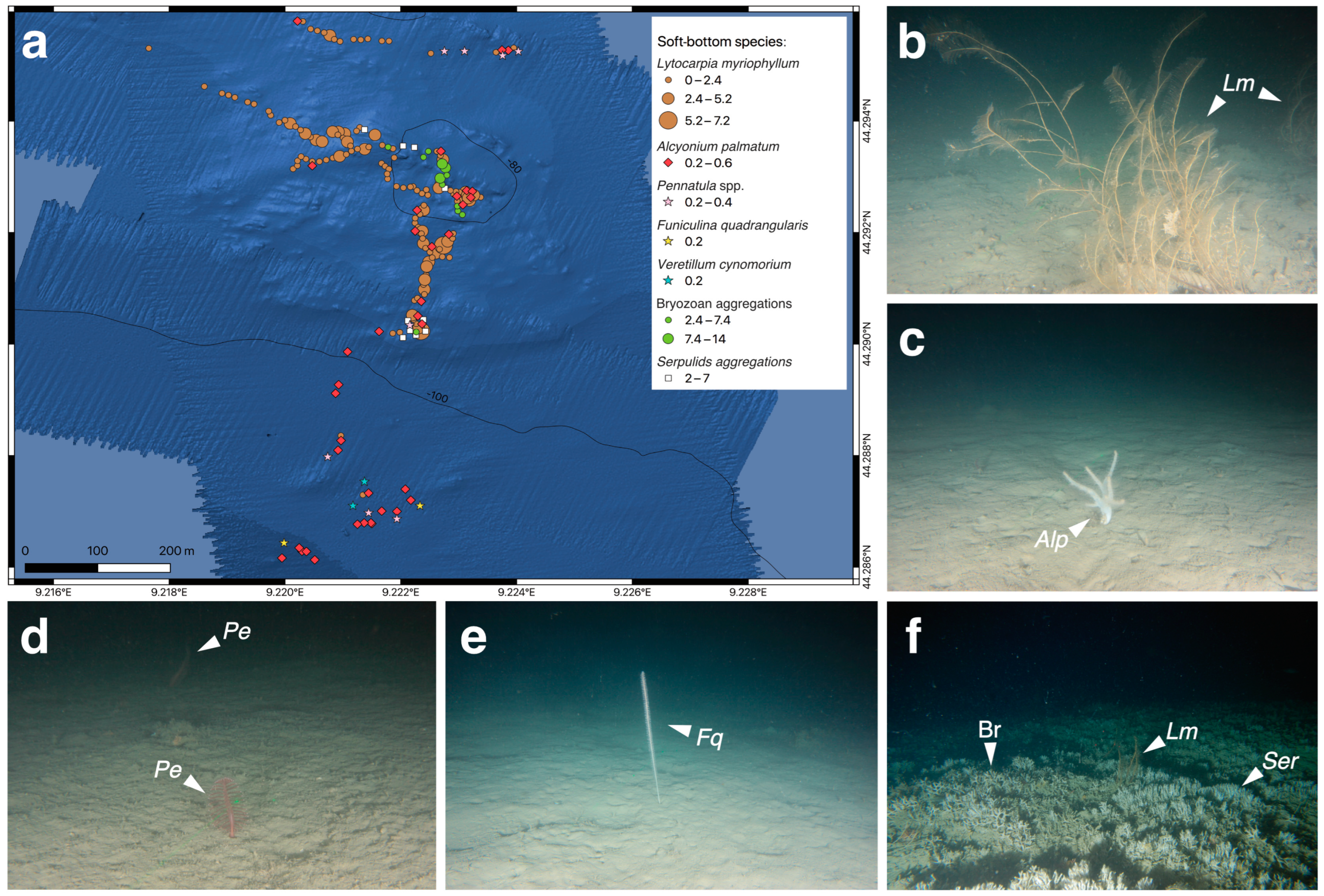
3.1. Biocoenotic Characterization
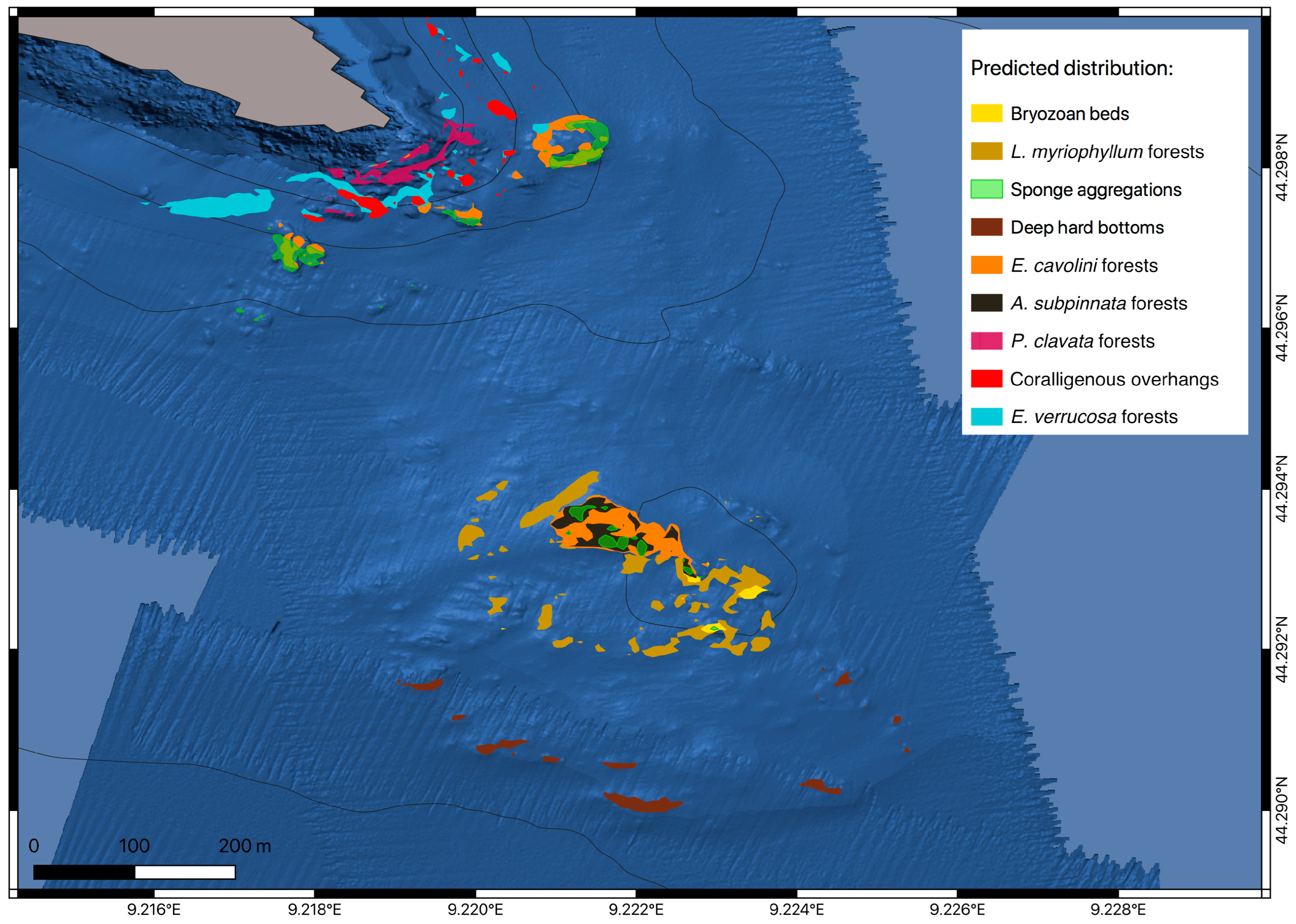
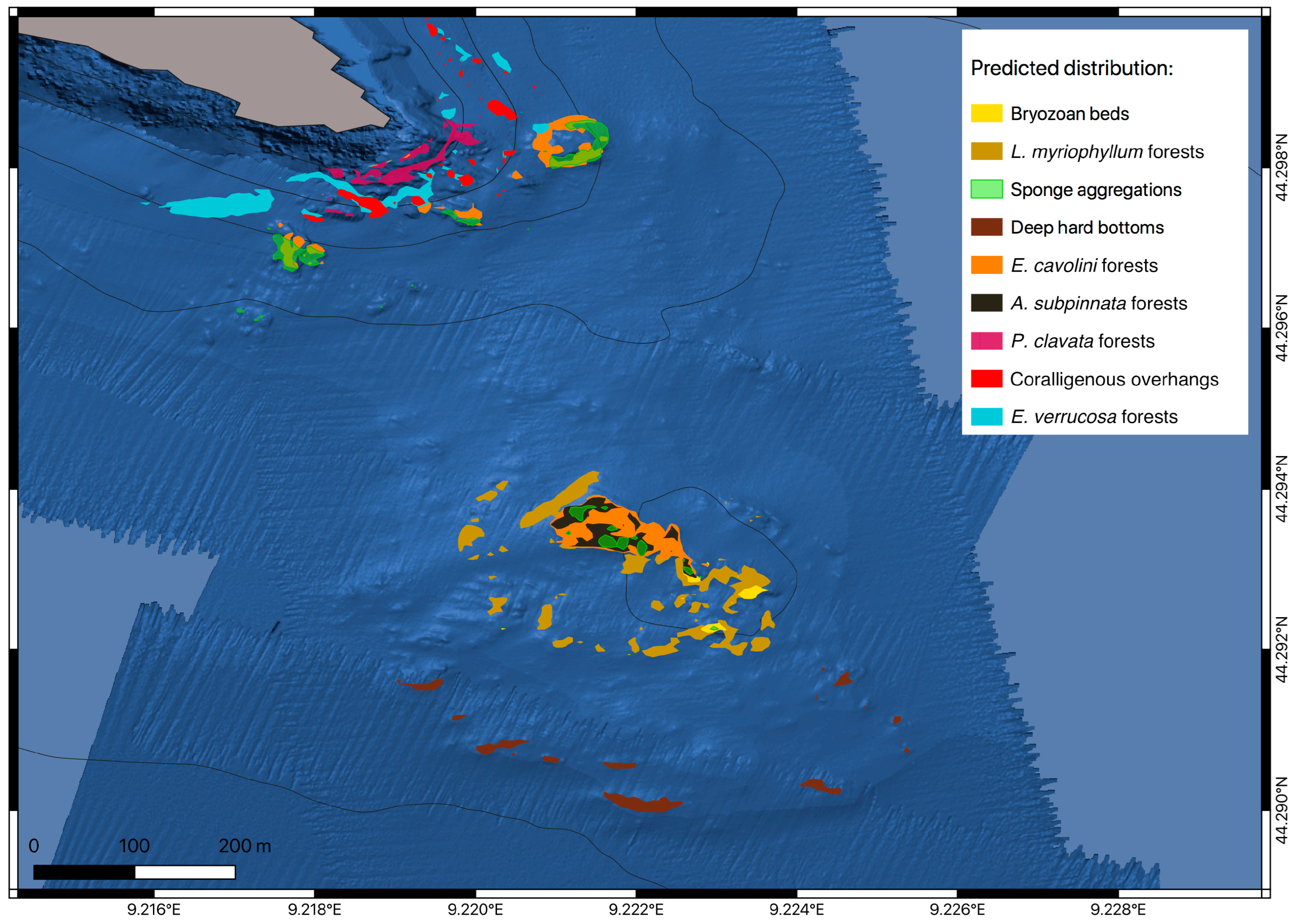
3.2. Predictive Habitat Mapping
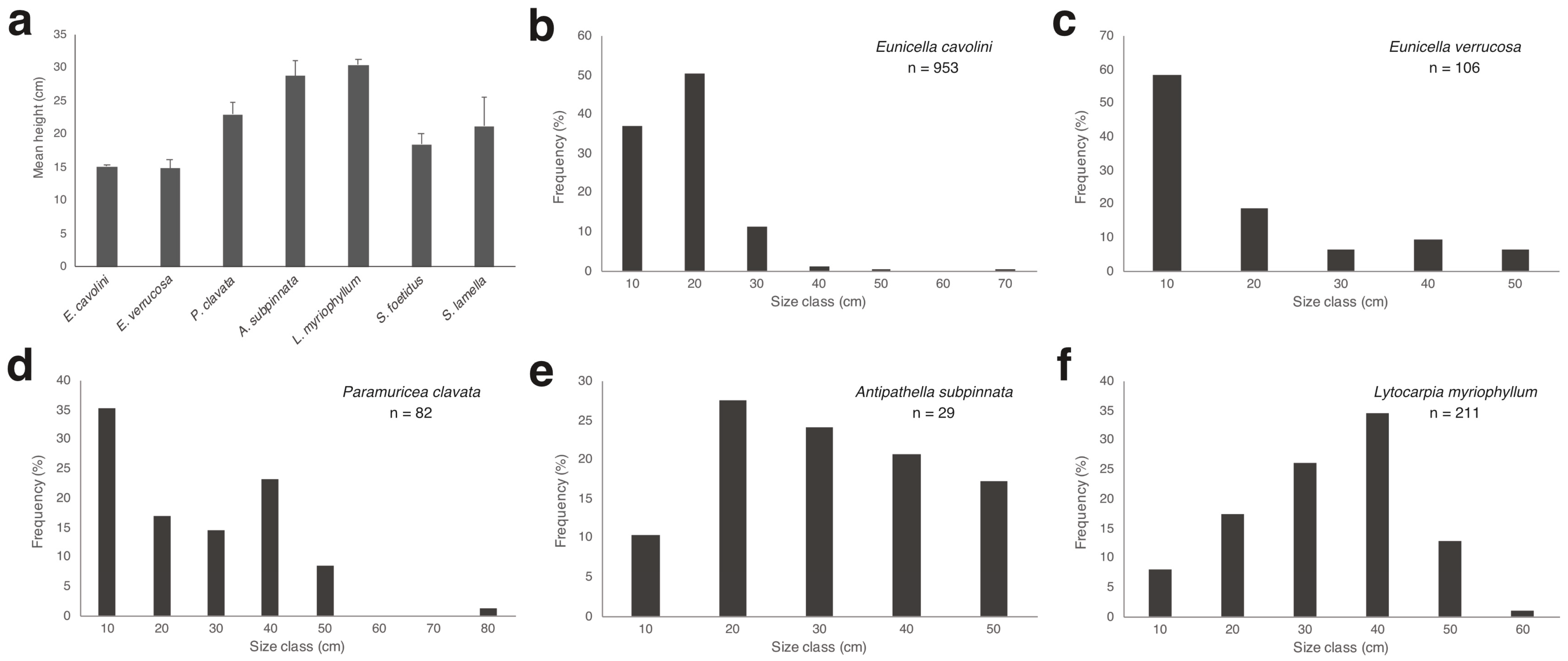
3.3. Structuring Species’ Morphometry and Size Distribution
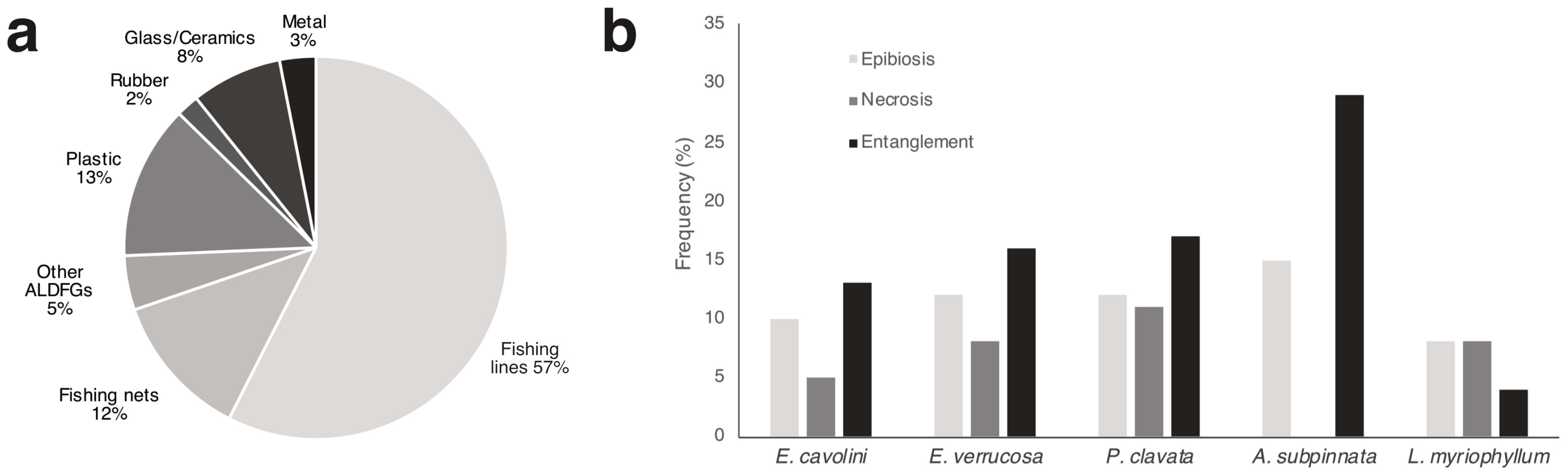
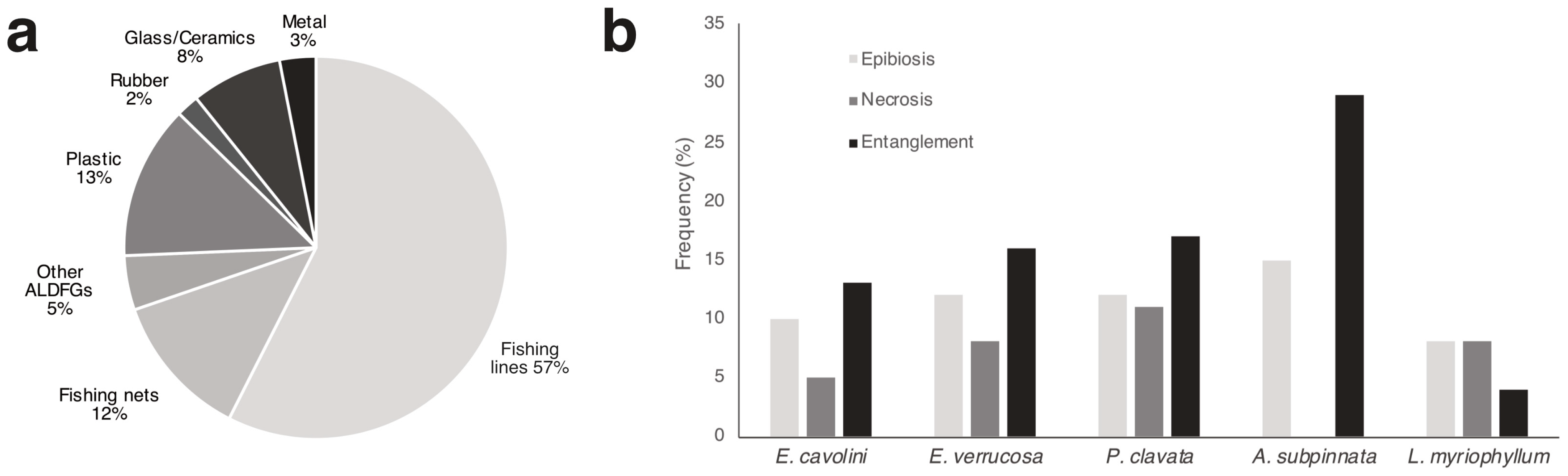
3.4. Environmental Status of the Benthic Biocoenoses
3.5. Fishing Pressure
4. Discussion
4.1. The Deep Shoal of Punta del Faro in the Context of the Portofino MPA
4.2. Peculiar Attributes of the Shoal at a Regional Scale
4.3. Fishing Pressure on and Environmental Status of the Shoal
4.4. Vulnerability and Conservation Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cattaneo-Vietti, R.; Bavestrello, G.; Cerrano, C. Prime osservazioni sul corallo rosso profondo lungo il Promontorio di Portofino. In Seconda Ricerca sul Corallo Rosso Nelle Acque Italiane; Ministero per le Risorse Agricole, Alimentari e Forestali; Centro Lubrense Esplorazioni Marine: Massa Lubrense, Italy, 1995; pp. 104–113. [Google Scholar]
- Diviacco, G.; Tunesi, L. Cartografia dei fondali antistanti la punta di Portofino (Mar Ligure) e osservazioni sui popolamenti bentici. Atti Congr. AIOL 1999, 13, 233–239. [Google Scholar]
- Cerrano, C.; Danovaro, R.; Gambi, C.; Pusceddu, A.; Riva, A.; Schiaparelli, S. Gold coral (Savalia savaglia) and gorgonian forests enhance benthic biodiversity and ecosystem functioning in the mesophotic zone. Biodivers. Conserv. 2010, 19, 153–167. [Google Scholar] [CrossRef]
- Cerrano, C.; Bianchelli, S.; Di Camillo, C.G.; Torsani, F.; Pusceddu, A. Do colonies of Lytocarpia myriophyllum, L. 1758 (Cnidaria, Hydrozoa) affect the biochemical composition and the meiofaunal diversity of surrounding sediments? Chem. Ecol. 2015, 31, 1–21. [Google Scholar] [CrossRef]
- Di Camillo, C.G.; Boero, F.; Gravili, C.; Previati, M.; Torsani, F.; Cerrano, C. Distribution, ecology and morphology of Lytocarpia myriophyllum (Cnidaria: Hydrozoa), a Mediterranean Sea habitat former to protect. Biodivers. Conserv. 2013, 22, 773–787. [Google Scholar] [CrossRef]
- Enrichetti, F.; Dominguez-Carrió, C.; Toma, M.; Bavestrello, G.; Betti, F.; Canese, S.; Bo, M. Megabenthic communities of the Ligurian deep continental shelf and shelf break (NW Mediterranean Sea). PLoS ONE 2019, 14, e0223949. [Google Scholar] [CrossRef] [PubMed]
- Coppari, M.; Mestice, F.; Betti, F.; Bavestrello, G.; Castellano, L.; Bo, M. Fragmentation, re-attachment ability and growth rate of the Mediterranean black coral Antipathella subpinnata. Coral Reefs 2019, 38, 1–14. [Google Scholar] [CrossRef]
- Coppari, M.; Ferrier-Pages, C.; Castellano, M.; Massa, F.; Olivari, E.; Bavestrello, G.; Povero, P.; Bo, M. Seasonal variation of the stable C and N isotopic composition of the mesophotic black coral Antipathella subpinnata (Ellis & Solander, 1786). Estuar. Coast. Shelf Sci. 2020, 233, 106520. [Google Scholar]
- Van de Water, J.A.; Coppari, M.; Enrichetti, F.; Ferrier-Pagès, C.; Bo, M. Local conditions influence the prokaryotic communities associated with the mesophotic black coral Antipathella subpinnata. Front. Microbiol. 2020, 11, 537813. [Google Scholar] [CrossRef]
- Terzin, M.; Paletta, M.G.; Matterson, K.; Coppari, M.; Bavestrello, G.; Abbiati, M.; Bo, M.; Costantini, F. Population genomic structure of the black coral Antipathella subpinnata in Mediterranean Vulnerable Marine Ecosystems. Coral Reefs 2021, 40, 751–766. [Google Scholar] [CrossRef]
- Enrichetti, F.; Dominguez-Carrió, C.; Toma, M.; Bavestrello, G.; Canese, S.; Bo, M. Assessment and distribution of seafloor litter on the deep Ligurian continental shelf and shelf break (NW Mediterranean Sea). Mar. Pollut. Bull. 2020, 151, 110872. [Google Scholar] [CrossRef]
- Bavestrello, G.; Cerrano, C.; Zanzi, D.; Cattaneo-Vietti, R. Damage by fishing activities to the Gorgonian coral Paramuricea clavata in the Ligurian Sea. Aquat. Conserv. Mar. Freshw. Ecosyst. 1997, 7, 253–262. [Google Scholar] [CrossRef]
- Bo, M.; Bava, S.; Canese, S.; Angiolillo, M.; Cattaneo-Vietti, R.; Bavestrello, G. Fishing impact on deep Mediterranean rocky habitats as revealed by ROV investigation. Biol. Conserv. 2014, 171, 167–176. [Google Scholar] [CrossRef]
- Hinz, H. Impact of bottom fishing on animal forests: Science, conservation, and fisheries management. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 1041–1059. [Google Scholar]
- Galgani, F.; Pham, C.K.; Claro, F.; Consoli, P. Marine animal forests as useful indicators of entanglement by marine litter. Mar. Pollut. Bull. 2018, 135, 735–738. [Google Scholar] [CrossRef]
- Angiolillo, M.; Fortibuoni, T. Impacts of marine litter on Mediterranean reef systems: From shallow to deep waters. Front. Mar. Sci. 2020, 7, 581966. [Google Scholar] [CrossRef]
- Canals, M.; Pham, C.K.; Bergmann, M.; Gutow, L.; Hanke, G.; Van Sebille, E.; Angiolillo, M.; Buhl-Mortensen, L.; Cau, A.; Ioakeimidis, C.; et al. The quest for seafloor macrolitter: A critical review of background knowledge, current methods and future prospects. Environ. Res. Lett. 2021, 16, 023001. [Google Scholar] [CrossRef]
- Prato, G.; Barrier, C.; Francour, P.; Cappanera, V.; Markantonatou, V.; Guidetti, P.; Mangialajo, L.; Cattaneo-Vietti, R.; Gascuel, D. Assessing interacting impacts of artisanal and recreational fisheries in a small Marine Protected Area (Portofino, NW Mediterranean Sea). Ecosphere 2016, 7, e01601. [Google Scholar] [CrossRef]
- Venturini, S.; Campodonico, P.; Cappanera, V.; Fanciulli, G.; Cattaneo Vietti, R. Recreational fisheries in Portofino Marine Protected Area, Italy: Some implications for the management. Fish. Manag. Ecol. 2017, 24, 382–391. [Google Scholar] [CrossRef]
- Venturini, S.; Merotto, L.; Campodonico, P.; Cappanera, V.; Fanciulli, G.; Cattaneo-Vietti, R. Recreational fisheries within the Portofino MPA and surrounding areas (Ligurian Sea, western Mediterranean Sea). Mediterr. Mar. Sci. 2019, 20, 142–150. [Google Scholar] [CrossRef]
- Betti, F.; Bavestrello, G.; Bo, M.; Ravanetti, G.; Enrichetti, F.; Coppari, M.; Cappanera, V.; Venturini, S.; Cattaneo-Vietti, R. Evidences of fishing impact on the coastal gorgonian forests inside the Portofino MPA (NW Mediterranean Sea). Ocean Coast. Manag. 2020, 187, 105105. [Google Scholar] [CrossRef]
- Coppo, S.; Diviacco, G.; Montepagano, E. Nuovo Atlante degli Habitat Marini della Liguria: Cartografia delle Praterie di Posidonia oceanica e dei Principali Popolamenti Marini Costieri. Regione Liguria. 2020. Available online: https://geoportal.regione.liguria.it/archivio-focus/item/605-nuovo-atlante-habitat-marini-2020.html (accessed on 6 August 2023).
- Tortonese, E. Bionomia marina della regione costiera fra Punta della Chiappa e Portofino (Riviera Ligure di Levante). Arch. Oceanogr. E Limnol. 1958, 11, 167–210. [Google Scholar]
- Tortonese, E. Nuovo contributo alla conoscenza del benthos della scogliera ligure. Arch. Oceanogr. E Limnol. 1961, 12, 163–183. [Google Scholar]
- Coratza, P.; Bollati, I.M.; Panizza, V.; Brandolini, P.; Castaldini, D.; Cucchi, F.; Deiana, G.; Del Monte, M.; Faccini, F.; Finocchiaro, F.; et al. Advances in geoheritage mapping: Application to iconic geomorphological examples from the italian landscape. Sustainability 2021, 13, 11538. [Google Scholar] [CrossRef]
- Tortonese, E. Recenti ricerche sul bentos in ambienti litorali del mare Ligure. Pubbl. Stn. Zool. Napoli 1962, 32, 99–116. [Google Scholar]
- Rossi, L. Idroidi viventi sulle scogliere del promontorio di Portofino (Golfo di Genova). Ann. Mus. Civ. Stor. Nat. Genova 1961, 72, 68–85. [Google Scholar]
- Sarà, M.; Balduzzi, A.; Boero, F.; Pansini, M.; Pessani, D.; Pronzato, R. Analisi di un popolamento bentonico di falesia del Promontorio di Portofino: Dati preliminari. Boll. Musei E Degli Ist. Biol. Dell’università Genova 1978, 46, 119–137. [Google Scholar]
- Morri, C.; Bianchi, C.N.; Damiani, V.; Peirano, A.; Romeo, G.; Tunesi, L. L’ambiente marino tra Punta della Chiappa e Sestri Levante (Mar Ligure): Profilo ecotipologico e proposta di carta bionomica. Boll. Musei E Degli Ist. Biol. Dell’università Genova 1986, 52, 213–231. [Google Scholar]
- Diviacco, G.; Tunesi, L.; Bianchi, C.N.; Morri, C.; Cattaneo-Vietti, R. Carta dei Principali Popolamenti Bentonici dell’Area Marina Protetta di Portofino (Scala 1:10.000); AMP Portofino: Santa Margherita Ligure, Italy, 2004. [Google Scholar]
- Millot, C. Circulation in the western Mediterranean Sea. J. Mar. Syst. 1999, 20, 423–442. [Google Scholar] [CrossRef]
- Doglioli, A.M.; Griffa, A.; Magaldi, M.G. Numerical study of a coastal current on a steep slope in presence of a cape: The case of the Promontorio di Portofino. J. Geophys. Res. Ocean. 2004, 109, 1–19. [Google Scholar] [CrossRef]
- Ruggieri, N.; Castellano, M.; Misic, C.; Gasparini, G.; Cattaneo-Vietti, R.; Povero, P. Seasonal and interannual dynamics of a coastal ecosystem (Portofino, Ligurian Sea) in relation to meteorological constraints. Geophys. Res. Abstr. 2006, 8, 07774. [Google Scholar]
- d’Ortenzio, F.; Ribera d’Alcalà, M. On the trophic regimes of the Mediterranean Sea: A satellite analysis. Bioscience 2009, 6, 139–148. [Google Scholar] [CrossRef]
- Vassallo, P.; Bellardini, D.; Castellano, M.; Dapueto, G.; Povero, P. Structure and functionality of the mesozooplankton community in a coastal marine environment: Portofino marine protected area (Liguria). Diversity 2021, 14, 19. [Google Scholar] [CrossRef]
- Bertolino, M.; Betti, F.; Bo, M.; Cattaneo-Vietti, R.; Pansini, M.; Romero, J.; Bavestrello, G. Changes and stability of a Mediterranean hard bottom benthic community over 25 years. J. Mar. Biol. Assoc. U. K. 2016, 96, 341–350. [Google Scholar] [CrossRef]
- Misic, C.; Castellano, M.; Harriague, A.C. Organic matter features, degradation and remineralisation at two coastal sites in the Ligurian Sea (NW Mediterranean) differently influenced by anthropogenic forcing. Mar. Environ. Res. 2011, 72, 67–74. [Google Scholar] [CrossRef]
- UNEP. Report of the 14th Ordinary Meeting of the Contracting Parties to the Convention for the Protection of the Marine Environment and Coastal Region of the Mediterranean and Its Protocols; UNEP (DEP)/MED IG: Nairobi, Kenya, 2005; pp. 13–16. [Google Scholar]
- Radicioli, M.; Angiolillo, M.; Giusti, M.; Proietti, R.; Fortibuoni, T.; Silvestri, C.; Tunesi, L. Monitoring coralligenous reefs in Italian coastal waters within the Marine Strategy Framework Directive. In Proceedings of the 4th Mediterranean Symposium on the Conservation of Coralligenous & Other Calcareous Bio-Concretions, Genoa, Italy, 20–21 September 2022; pp. 96–101. [Google Scholar]
- Markantonatou, V.; Marconi, M.; Cappanera, V.; Campodonico, P.; Bavestrello, A.; Cattaneo-Vietti, R.; Papadopoulou, N.; Smith, C.; Cerrano, C. Spatial allocation of fishing activity on coralligenous habitats in Portofino MPA (Liguria, Italy). In Proceedings of the Second Mediterranean Symposium on the Conservation of Coralligenous and Other Calcareous Bio-Concretions, Portorož, Slovenia, 29–30 October 2014; Bouafif, C., Langar, H., Ouerghi, A., Eds.; RAC/SPA: Tunis, Tunisia, 2014; Volume 247, pp. 123–188. [Google Scholar]
- Cappanera, V.; Venturini, S.; Campodonico, P.; Blini, V.; Ortenzi, C. Valutazione Dell’impatto Antropico sul Sistema Costiero, con Particolare Riferimento alla Pressione Antropica All’interno dell’Area Marina Protetta del Promontorio di Portofino; Portofino MPA Report; CONISMA: Santa Margherita Ligure, Italy, 2006; pp. 1–267. [Google Scholar]
- Venturini, S.; Massa, F.; Castellano, M.; Costa, S.; Lavarello, I.; Olivari, E.; Povero, P. Recreational boating in Ligurian Marine Protected Areas (Italy): A quantitative evaluation for a sustainable management. Environ. Manag. 2016, 57, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Bava, S.; Cappanera, V.; Fanciulli, G.; Povero, P.; Tunesi, L.; Cattaneo-Vietti, R. Stima dell’impatto antropico nell’AMP di Portofino: Proposta di uno strumento di sintesi delle pressioni relative alla fruizione. Biol. Mar. Mediterr. 2007, 14, 70–71. [Google Scholar]
- Enrichetti, F.; Bo, M.; Morri, C.; Montefalcone, M.; Toma, M.; Bavestrello, G.; Tunesi, L.; Canese, S.; Giusti, M.; Salvati, E.; et al. Assessing the environmental status of temperate mesophotic reefs: A new, integrated methodological approach. Ecol. Indic. 2019, 102, 218–229. [Google Scholar] [CrossRef]
- Parravicini, V.; Rovere, A.; Vassallo, P.; Micheli, F.; Montefalcone, M.; Morri, C.; Paoli, C.; Albertelli, G.; Fabiano, M.; Bianchi, C.N. Understanding relationships between conflicting human uses and coastal ecosystems status: A geospatial modeling approach. Ecol. Indic. 2012, 19, 253–263. [Google Scholar] [CrossRef]
- Dapueto, G.; Massa, M.; Pergent-Martini, C.; Povero, P.; Rigo, I.; Vassallo, P.; Venturini, S.; Paoli, C. Sustainable management accounting model of recreational boating anchoring in Marine Protected Areas. J. Clean. Prod. 2022, 342, 130905. [Google Scholar] [CrossRef]
- Breiman, L. Random Forest. Mach. Learn 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Vassallo, P.; Bianchi, C.N.; Paoli, C.; Holon, F.; Navone, A.; Bavestrello, G.; Cattaneo-Vietti, R.; Morri, C. A predictive approach to benthic marine habitat mapping: Efficacy and management implications. Mar. Pollut. Bull. 2018, 131, 218–232. [Google Scholar] [CrossRef]
- Efron, B. Bootstrap methods: Another look at the jack- knife. Ann. Stat. 1979, 7, 569–593. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Cerrano, C.; Bastari, A.; Calcinai, B.; Di Camillo, C.; Pica, D.; Puce, S.; Valisano, L.; Torsani, F. Temperate mesophotic ecosystems: Gaps and perspectives of an emerging conservation challenge for the Mediterranean Sea. Eur. Zool. J. 2019, 86, 370–388. [Google Scholar] [CrossRef]
- Bell, J.J.; Micaroni, V.; Harris, B.; Strano, F.; Broadribb, M.; Rogers, A. Global status, impacts, and management of rocky temperate mesophotic ecosystems. Conserv. Biol. 2022, e13945. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Bramanti, L.; Gori, A.; Orejas, C. Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Springer International Publishing: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Gori, A.; Bavestrello, G.; Grinyó, J.; Dominguez-Carrió, C.; Ambroso, S.; Bo, M. Animal Forests in deep coastal bottoms and continental shelf of the Mediterranean Sea. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 207–233. [Google Scholar]
- Bo, M.; Canese, S.; Spaggiari, C.; Pusceddu, A.; Bertolino, M.; Angiolillo, M.; Giusti, M.; Loreto, M.F.; Salvati, E.; Greco, S.; et al. Deep coral oases in the south Tyrrhenian sea. PLoS ONE 2012, 7, e49870. [Google Scholar] [CrossRef]
- Bo, M.; Bavestrello, G.; Angiolillo, M.; Calcagnile, L.; Canese, S.; Cannas, R.; Cau, A.; D’Elia, M.; D’Oriano, F.; Follesa, M.C.; et al. Persistence of pristine deep-sea coral gardens in the Mediterranean Sea (SW Sardinia). PLoS ONE 2015, 10, e0119393. [Google Scholar] [CrossRef]
- Montefalcone, M.; Tunesi, L.; Ouerghi, A. A review of the classification systems for marine benthic habitats and the new updated Barcelona Convention classification for the Mediterranean. Mar. Environ. Res. 2021, 169, 105387. [Google Scholar] [CrossRef]
- Pérès, J.M.; Picard, J. Noveau Manuel de Bionomie Bentique de la Mer Méditerranée; Station Marine d’Eudoume: Marseille, France, 1964; p. 137. [Google Scholar]
- Canessa, M.; Betti, F.; Bo, M.; Enrichetti, F.; Toma, M.; Bavestrello, G. Possible Population Growth of Astrospartus mediterraneus (Risso, 1826)(Ophiuroidea, Gorgonocephalidae) in the Mediterranean Sea. Diversity 2023, 15, 122. [Google Scholar] [CrossRef]
- Cerrano, C.; Bavestrello, G.; Bianchi, C.N.; Cattaneo-Vietti, R.; Bava, S.; Morganti, C.; Morri, C.; Picco, P.; Sara, G.; Schiaparelli, S.; et al. A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (North-western Mediterranean), summer 1999. Ecol. Lett. 2000, 3, 284–293. [Google Scholar] [CrossRef]
- Cánovas-Molina, A.; Montefalcone, M.; Bavestrello, G.; Masmoudi, M.B.; Haguenauer, A.; Hammami, P.; Chaoui, L.; Kara, M.H.; Aurelle, D. From depth to regional spatial genetic differentiation of Eunicella cavolini in the NW Mediterranean. Comptes Rendus Biol. 2018, 341, 421–432. [Google Scholar] [CrossRef]
- Bo, M.; Tazioli, S.; Spanò, N.; Bavestrello, G. Antipathella subpinnata (Antipatharia, Myriopathidae) in Italian seas. Ital. J. Zool. 2008, 75, 185–195. [Google Scholar] [CrossRef]
- Gaino, E.; Scoccia, F. Gamete spawning in Antipathella subpinnata (Anthozoa, Antipatharia): A structural and ultrastructural investigation. Zoomorphology 2010, 129, 213–219. [Google Scholar] [CrossRef]
- Coppari, M.; Fumarola, L.; Bramanti, L.; Romans, P.; Pillot, R.; Bavestrello, G.; Bo, M. Unveiling asexual reproductive traits in black corals: Polyp bail-out in Antipathella subpinnata. Coral Reefs 2020, 39, 1517–1523. [Google Scholar] [CrossRef]
- Costantini, F.; Gori, A.; Lopez-González, P.; Bramanti, L.; Rossi, S.; Gili, J.M.; Abbiati, M. Limited genetic connectivity between gorgonian morphotypes along a depth gradient. PLoS ONE 2016, 11, e0160678. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L. Contributo allo studio della fauna di profondità vivente presso la Riviera ligure di Levante. Ann. Mus. Civ. Stor. Nat. Genova Suppl. Doriana 1958, 2, 1–13. [Google Scholar]
- Boero, F.; Fresi, E. Zonation and evolution of a rocky bottom hydroid community. Mar. Ecol. 1986, 7, 123–150. [Google Scholar] [CrossRef]
- Dominguez-Carrió, C.; Riera, J.L.; Robert, K.; Zabala, M.; Requena, S.; Gori, A.; Orejas, C.; Iacono, C.L.; Estournel, C.; Corbera, G.; et al. Diversity, structure and spatial distribution of megabenthic communities in Cap de Creus continental shelf and submarine canyon (NW Mediterranean). Prog. Oceanogr. 2022, 208, 102877. [Google Scholar] [CrossRef]
- Enrichetti, F.; Bava, S.; Bavestrello, G.; Betti, F.; Lanteri, L.; Bo, M. Artisanal fishing impact on deep coralligenous animal forests: A Mediterranean case study of marine vulnerability. Ocean. Coast. Manag. 2019, 177, 112–126. [Google Scholar] [CrossRef]
- Grinyó, J.; Gori, A.; Greenacre, M.; Requena, S.; Canepa, A.; Iacono, C.L.; Ambroso, S.; Purroy, A.; Gili, J.M. Megabenthic assemblages in the continental shelf edge and upper slope of the Menorca Channel, Western Mediterranean Sea. Prog. Oceanogr. 2018, 162, 40–51. [Google Scholar] [CrossRef]
- De la Torriente, A.; Serrano, A.; Fernández-Salas, L.M.; García, M.; Aguilar, R. Identifying epibenthic habitats on the Seco de los Olivos Seamount: Species assemblages and environmental characteristics. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2018, 135, 9–22. [Google Scholar] [CrossRef]
- Gili, J.M.; Coma, R. Benthic suspension feeders: Their paramount role in littoral marine food webs. Trends Ecol. Evol. 1998, 13, 316–321. [Google Scholar] [CrossRef]
- Buhl-Mortensen, L.; Vanreusel, A.; Gooday, A.J.; Levin, L.A.; Priede, I.G.; Buhl-Mortensen, P.; Gheerardyn, H.; King, N.J.; Raes, M. Biological structures as a source of habitat heterogeneity and biodiversity on the deep ocean margins. Mar. Ecol. 2010, 31, 21–50. [Google Scholar] [CrossRef]
- Rossi, S.; Coppari, M.; Viladrich, N. Benthic-pelagic coupling: New perspectives in the animal forests. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 855–885. [Google Scholar]
- Fourt, M.; Goujard, A.; Pérez, T.; Chevaldonné, P. Guide de la Faune Profonde de la Mer Méditerranée. Explorations des Roches et Canyons Sous-Marins des Côtes Françaises; Publications Scientifiques du Muséum d’Histoire Naturelle, Patrimoines Naturels: Paris, France, 2017; Volume 75. [Google Scholar]
- Bo, M.; Canese, S.; Bavestrello, G. Discovering Mediterranean black coral forests: Parantipathes larix (Anthozoa: Hexacorallia) in the Tuscan Archipelago, Italy. Ital. J. Zool. 2014, 81, 112–125. [Google Scholar] [CrossRef]
- Turicchia, E.; Abbiati, M.; Sweet, M.; Ponti, M. Mass mortality hits gorgonian forests at Montecristo Island. Dis. Aquat. Org. 2018, 131, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Sini, M.; Kipson, S.; Linares, C.; Koutsoubas, D.; Garrabou, J. The yellow gorgonian Eunicella cavolini: Demography and disturbance levels across the Mediterranean Sea. PLoS ONE 2015, 10, e0126253. [Google Scholar] [CrossRef]
- Grinyó, J.; Gori, A.; Ambroso, S.; Purroy, A.; Calatayud, C.; Dominguez-Carrió, C.; Coppari, M.; Iacono, C.L.; López-González, P.J.; Gili, J.M. Diversity, distribution and population size structure of deep Mediterranean gorgonian assemblages (Menorca Channel, Western Mediterranean Sea). Prog. Oceanogr. 2016, 145, 42–56. [Google Scholar] [CrossRef]
- Carugati, L.; Moccia, D.; Bramanti, L.; Cannas, R.; Follesa, M.C.; Salvadori, S.; Cau, A. Deep-Dwelling Populations of Mediterranean Corallium rubrum and Eunicella cavolini: Distribution, Demography, and Co-Occurrence. Biology 2022, 11, 333. [Google Scholar] [CrossRef]
- Bavestrello, G.; Bertone, S.; Cattaneo-Vietti, R.; Cerrano, C.; Gaino, E.; Zanzi, D. Mass mortality of Paramuricea clavata (Anthozoa, Cnidaria) on Portofino Promontory cliffs, Ligurian Sea, Mediterranean Sea. Mar. Life 1994, 4, 15–19. [Google Scholar]
- Cerrano, C.; Arillo, A.; Azzini, F.; Calcinai, B.; Castellano, L.; Muti, C.; Valisano, L.; Zega, G.; Bavestrello, G. Gorgonian population recovery after a mass mortality event. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 147–157. [Google Scholar] [CrossRef]
- Schiaparelli, S.; Castellano, M.; Povero, P.; Sartoni, G.; Cattaneo-Vietti, R. A benthic mucilage event in North-Western Mediterranean Sea and its possible relationships with the summer 2003 European heatwave: Short term effects on littoral rocky assemblages. Mar. Ecol. 2007, 28, 341–353. [Google Scholar] [CrossRef]
- Garrabou, J.; Coma, R.; Bensoussan, N.; Bally, M.; Chevaldonné, P.; Cigliano, M.; Díaz, D.; Harmelin, J.G.; Gambi, M.C.; Kersting, D.K.; et al. Mass mortality in Northwestern Mediterranean rocky benthic communities: Effects of the 2003 heat wave. Glob. Chang. Biol. 2009, 15, 1090–1103. [Google Scholar] [CrossRef]
- Otero, M.D.M.; Numa, C.; Bo, M.; Orejas, C.; Garrabou, J.; Cerrano, C.; Kružić, P.; Antoniadou, C.; Aguilar, R.; Kipson, S.; et al. Overview of the Conservation Status of Mediterranean Anthozoans; International Union for Conservation of Nature and Natural Resources (IUCN): Gland, Switzerland, 2017. [Google Scholar]
- Bo, M.; Bavestrello, G. Mediterranean Black Coral Communities. In Mediterranean Cold-Water Corals: Past, Present and Future; Orejas, C., Jiménez, C., Eds.; Coral Reefs of the World; Springer: Cham, Switzerland, 2019; Volume 9, pp. 249–251. [Google Scholar]
- Enrichetti, F.; Toma, M.; Bavestrello, G.; Betti, F.; Giusti, M.; Canese, S.; Moccia, D.; Quarta, G.; Calcagnile, L.; Andaloro, F.; et al. Facies created by the yellow coral Dendrophyllia cornigera (Lamarck, 1816): Origin, substrate preferences and habitat complexity. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2023, 195, 104000. [Google Scholar] [CrossRef]
- Enrichetti, F.; Bavestrello, G.; Betti, F.; Coppari, M.; Toma, M.; Pronzato, R.; Canese, S.; Bertolino, M.; Costa, G.; Pansini, M.; et al. Keratose-dominated sponge grounds from temperate mesophotic ecosystems (NW Mediterranean Sea). Mar. Ecol. 2020, 41, e12620. [Google Scholar] [CrossRef]
- Relini, G.; Peirano, A.; Tunesi, L. Osservazioni sulle comunità dei fondi strascicabili del Mar Ligure Centro-Orientale. Boll. Musei E Degli Ist. Biol. Dell’università Genova 1986, 52, 139–161. [Google Scholar]
- Cocito, S.; Bedulli, D.; Sgorbini, S. Distribution patterns of the sublittoral epibenthic assemblages on a rocky shoal in the Ligurian Sea (NW Mediterranean). Sci. Mar. 2002, 66, 175–181. [Google Scholar] [CrossRef]
- Lombardi, C.; Taylor, P.D.; Cocito, S. Bryozoan constructions in a changing Mediterranean Sea. In The Mediterranean Sea; Springer: Dordrecht, The Netherlands, 2014; pp. 373–384. [Google Scholar]
- Asch, R.G.; Collie, J.S. Changes in a benthic megafaunal community due to disturbance from bottom fishing and the establishment of a fishery closure. Fish. Bull. 2008, 106, 438–456. [Google Scholar]
- de Juan, S.; Demestre, M.; Thrush, S. Defining ecological indicators of trawling disturbance when everywhere that can be fished is fished: A Mediterranean case study. Mar. Policy 2009, 33, 472–478. [Google Scholar] [CrossRef]
- Cataudella, S.; Spagnolo, M. Lo Stato della Pesca e Dell’acquacoltura nei Mari Italiani; Ministero delle Politiche Agricole Alimentari e Forestali: Rome, Italy, 2011. [Google Scholar]
- Falsone, F.; Scannella, D.; Geraci, M.L.; Vitale, S.; Colloca, F.; Di Maio, F.; Milisenda, G.; Gancitano, V.; Bono, G.; Fiorentino, F. Identification and characterization of trammel net métiers: A case study from the southwestern Sicily (Central Mediterranean). Reg. Stud. Mar. Sci. 2020, 39, 101419. [Google Scholar] [CrossRef]
- Erzini, K.; Monteiro, C.C.; Ribeiro, J.; Santos, M.N.; Gaspar, M.; Monteiro, P.; Borges, T.C. An experimental study of gill net and trammel net ‘ghost fishing’ off the Algarve (southern Portugal). Mar. Ecol. Prog. Ser. 1997, 158, 257–265. [Google Scholar] [CrossRef]
- Gonçalves, J.M.S.; Stergiou, K.I.; Hernando, J.A.; Puente, E.; Moutopoulos, D.K.; Arregi, L.; Soriguer, M.C.; Vilas, C.; Coelho, R.; Erzini, K. Discards from experimental trammel nets in southern European small-scale fisheries. Fish. Resour. 2007, 88, 5–14. [Google Scholar] [CrossRef]
- Batista, M.I.; Teixeira, C.M.; Cabral, H.N. Catches of target species and bycatches of an artisanal fishery: The case study of a trammel net fishery in the Portuguese coast. Fish. Resour. 2009, 100, 167–177. [Google Scholar] [CrossRef]
- Catanese, G.; Hinz, H.; del Mar Gil, M.; Palmer, M.; Breen, M.; Mira, A.; Pastor, E.; Grau, A.; Campos-Candela, A.; Koleva, E.; et al. Comparing the catch composition, profitability and discard survival from different trammel net designs targeting common spiny lobster (Palinurus elephas) in a Mediterranean fishery. PeerJ 2018, 6, e4707. [Google Scholar] [CrossRef] [PubMed]
- Erzini, K.; Gonçalves, J.M.; Bentes, L.; Moutopoulos, D.K.; Casal, J.A.H.; Soriguer, M.C.; Puente, E.; Errazkin, L.A.; Stergiou, K.I. Size selectivity of trammel nets in southern European small-scale fisheries. Fish. Res. 2006, 79, 183–201. [Google Scholar] [CrossRef]
- Stergiou, K.I.; Moutopoulos, D.K.; Soriguer, M.C.; Puente, E.; Lino, P.G.; Zabala, C.; Monteiro, P.; Errazkin, L.A.; Erzini, K. Trammel net catch species composition, catch rates and métiers in southern European waters: A multivariate approach. Fish. Res. 2006, 79, 170–182. [Google Scholar] [CrossRef]
- Gil, M.M.; Catanese, G.; Palmer, M.; Hinz, H.; Pastor, E.; Mira, A.; Grau, A.; Koleva, E.; Grau, A.M.; Morales-Nin, B. Commercial catches and discards of a Mediterranean small-scale cuttlefish fishery: Implications of the new EU discard policy. Sci. Mar. 2018, 82, 155–164. [Google Scholar] [CrossRef]
- Sartor, P.; Veli, D.L.; De Carlo, F.; Ligas, A.; Massaro, A.; Musumeci, C.; Sartini, M.; Rossetti, I.; Sbrana, M.; Viva, C. Reducing unwanted catches of trammel nets: Experimental results of the “guarding net” in the caramote prawn, Penaeus kerathurus, small-scale fishery of the Ligurian Sea (western Mediterranean). Sci. Mar. 2018, 82, 131–140. [Google Scholar] [CrossRef]
- Montseny, M.; Linares, C.; Viladrich, N.; Biel, M.; Gracias, N.; Baena, P.; Quintanilla, E.; Ambroso, S.; Grinyó, J.; Santín, A.; et al. Involving fishers in scaling up the restoration of cold-water coral gardens on the Mediterranean continental shelf. Biol. Conserv. 2021, 262, 109301. [Google Scholar] [CrossRef]
- Sampaio, I.; Braga-Henriques, A.; Pham, C.; Ocaña, O.; De Matos, V.; Morato, T.; Porteiro, F.M. Cold-water corals landed by bottom longline fisheries in the Azores (north-eastern Atlantic). J. Mar. Biol. Assoc. U. K. 2012, 92, 1547–1555. [Google Scholar] [CrossRef]
- Mytilineou, C.; Smith, C.J.; Anastasopoulou, A.; Papadopoulou, K.N.; Christidis, G.; Bekas, P.; Kavadas, S.; Dokos, J. New cold-water coral occurrences in the Eastern Ionian Sea: Results from experimental long line fishing. Deep Sea Res. Part II 2014, 99, 146–157. [Google Scholar] [CrossRef]
- Kaiser, M.J.; Hormbrey, S.; Booth, J.R.; Hinz, H.; Hiddink, J.G. Recovery linked to life history of sessile epifauna following exclusion of towed mobile fishing gear. J. Appl. Ecol. 2018, 55, 1060–1070. [Google Scholar] [CrossRef]
- Biel-Cabanelas, M.; Santín, A.; Montasell, M.; Salazar, J.; Baena, P.; Viladrich, N.; Montseny, M.; Corbera, G.; Ambroso, S.; Grinyó, J. From emblematic to problematic: The case of Astrospartus mediterraneus (Risso, 1826)(Echinodermata: Ophiuroidea) in the artisanal fishing grounds of the Cap de Creus area (NW Mediterranean Sea). Cont. Shelf Res. 2023, 255, 104925. [Google Scholar] [CrossRef]
- Yıldız, T.; Karakulak, F.S. Types and extent of fishing gear losses and their causes in the artisanal fisheries of Istanbul, Turkey. J. Appl. Ichthyol. 2016, 32, 432–438. [Google Scholar] [CrossRef]
- Richardson, K.; Hardesty, B.D.; Vince, J.Z.; Wilcox, C. Global causes, drivers, and prevention measures for lost fishing gear. Front. Mar. Sci. 2021, 8, 690447. [Google Scholar] [CrossRef]
- Enrichetti, F.; Bavestrello, G.; Betti, F.; Rindi, F.; Tregrosso, A.; Bo, M. Fate of lost fishing gears: Experimental evidence of biofouling colonization patterns from the northwestern Mediterranean Sea. Environ. Pollut. 2021, 268, 115746. [Google Scholar] [CrossRef]
- Bavestrello, G.; Bo, M.; Canese, S.; Sandulli, R.; Cattaneo-Vietti, R. The red coral populations of the gulfs of Naples and Salerno: Human impact and deep mass mortalities. Ital. J. Zool. 2014, 81, 552–563. [Google Scholar] [CrossRef]
- Angiolillo, M.; di Lorenzo, B.; Farcomeni, A.; Bo, M.; Bavestrello, G.; Santangelo, G.; Cau, A.; Mastascusa, V.; Cau, A.; Sacco, F.; et al. Distribution and assessment of marine debris in the deep Tyrrhenian Sea (NW Mediterranean Sea, Italy). Mar. Pollut. Bull. 2015, 92, 149–159. [Google Scholar] [CrossRef]
- Cattaneo-Vietti, R.; Bavestrello, G.; Bo, M.; Canese, S.; Vigo, A.; Andaloro, F. Illegal ingegno fishery and conservation of deep red coral banks in the Sicily Channel (Mediterranean Sea). Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 604–616. [Google Scholar] [CrossRef]
- Dominguez-Carrió, C.; Sanchez-Vidal, A.; Estournel, C.; Corbera, G.; Riera, J.L.; Orejas, C.; Canals, M.; Gili, J.M. Seafloor litter sorting in different domains of Cap de Creus continental shelf and submarine canyon (NW Mediterranean Sea). Mar. Pollut. Bull. 2020, 161, 111744. [Google Scholar] [CrossRef]
- Aguilar, R.; Perry, L.A.; López, J. Conservation and management of vulnerable marine ecosystems. The ecology of benthic biodiversity hotspot. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 1165–1207. [Google Scholar]
- Turner, J.A.; Andradi-Brown, D.A.; Gori, A.; Bongaerts, P.; Burdett, H.L.; Ferrier-Pagès, C.; Voolstra, C.R.; Weinstein, D.K.; Bridge, T.C.; Costantini, F.; et al. Key questions for research and conservation of mesophotic coral ecosystems and temperate mesophotic ecosystems. Mesophotic Coral Ecosyst. 2019, 12, 989–1003. [Google Scholar]
- FAO. International Guidelines for the Management of Deep-Sea Fisheries in the High Seas; FAO: Rome, Italy, 2009; p. 73. [Google Scholar]
- Morato, T.; Pham, C.K.; Pinto, C.; Golding, N.; Ardron, J.A.; Duran Munoz, P.; Neat, F. A multi criteria assessment method for identifying Vulnerable Marine Ecosystems in the North-East Atlantic. Front. Mar. Sci. 2018, 5, 460. [Google Scholar] [CrossRef]
- Paoli, C.; Povero, P.; Burgos, E.; Dapueto, G.; Fanciulli, G.; Massa, F.; Scarpellini, P.; Vassallo, P. Natural capital and environmental flows assessment in marine protected areas: The case study of Liguria region (NW Mediterranean Sea). Ecol. Model. 2018, 368, 121–135. [Google Scholar] [CrossRef]
- EC. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions—EU Biodiversity Strategy for 2030 Bringing Nature Back into Our Lives; European Commission: Brussels, Belgium, 2020.
- Laffoley, D.; Baxter, J.M.; Amon, D.J.; Currie, D.E.; Downs, C.A.; Hall-Spencer, J.M.; Harden-Davies, H.; Page, R.; Reid, C.P.; Roberts, C.M.; et al. Eight urgent, fundamental and simultaneous steps needed to restore ocean health, and the consequences for humanity and the planet of inaction or delay. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 194–208. [Google Scholar] [CrossRef]
- Secretariat of the United Nations Convention on Biological Diversity, First Draft of the Post-2020 Global Biodiversity Framework, 2021, Cbd/Wg2020/3/3. Available online: https://www.cbd.int/doc/c/abb5/591f/2e46096d3f0330b08ce87a45/wg2020-03-03-en.pdf (accessed on 6 August 2023).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dive CODE | Date | Location | Start Position (X) | Start Position (Y) | End Position (X) | End Position (Y) | Length (m) | Depth Range (m) | N° of OTUs |
|---|---|---|---|---|---|---|---|---|---|
| 12_01 | 02 06 2012 | DS (SE sector) | 9.2213 | 44.2872 | 9.2232 | 44.2926 | 923 | 73–101 | 46 |
| 12_02 | 02 06 2012 | DS and coastal cliff | 9.2231 | 44.2927 | 9.2180 | 44.2975 | 1494 | 55–90 | 69 |
| 12_04 | 03 06 2012 | San Giorgio shoals | 9.2058 | 44.2922 | 9.2075 | 44.2938 | 544 | 89–99 | 33 |
| 16_04 | 25 08 2016 | DS (crest) | 9.2199 | 44.2932 | 9.2228 | 44.2916 | 722 | 61–83 | 53 |
| 16_05 | 25 08 2016 | Coastal cliff | 9.2205 | 44.2989 | 9.2186 | 44.2975 | 908 | 59–61 | 51 |
| 16_06 | 25 08 2016 | DS (southern deep sector) | 9.2221 | 44.2874 | 9.2201 | 44.2862 | 503 | 101–104 | 22 |
| 16_07 | 25 08 2016 | DS (N sector) + coastal cliff | 9.2240 | 44.2953 | 9.2178 | 44.2971 | 747 | 60–86 | 52 |
| 20_R1 | 02 07 2020 | DS (crest) | 9.2213 | 44.2980 | 9.2204 | 44.2989 | 100 | 55–66 | 39 |
| Variables | Mean Decrease in Accuracy (%) |
|---|---|
| Slope | 26 |
| Roughness | 22 |
| Depth | 19 |
| Distance from coast | 13 |
| TRI | 7 |
| Aspect | 5 |
| TPI | 2 |
| Communities | Accuracy |
|---|---|
| A. subpinnata forests | 0.97 |
| E. cavolini forests | 0.96 |
| Coralligenous overhangs | 0.92 |
| Deep hard bottoms | 0.92 |
| Bryozoan beds | 0.91 |
| Sponge aggregations | 0.91 |
| E. verrucosa forests | 0.88 |
| L. myriophyllum forests | 0.85 |
| P. clavata forests | 0.70 |
| Transect ID | SR | BC | CC | DM | SSD | SSH | Status Index | SD | ENT | NCR | EBP | LD | LT | Impact Index | MACS Index |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2012_R1 | 67 | 33 | 33 | 33 | 100 | 100 | Good (56) | 100 | 33 | 33 | 33 | 100 | 100 | Very high (67) | Poor (45) |
| 2016_R2 | 67 | 33 | 33 | 33 | 100 | 67 | Moderate (50) | 100 | 33 | 33 | 33 | 67 | 100 | High (61) | Poor (44) |
| 2020_R3 | 67 | 33 | 33 | 33 | 100 | 100 | Good (56) | 100 | 33 | 33 | 67 | 100 | 100 | Very high (72) | Poor (42) |
| FAO Criteria | Evaluation |
|---|---|
| Uniqueness or rarity | Several biological features of the shoal are unique to the eastern Ligurian Sea, including E. cavolini and A. subpinnata forests. In addition, these species are listed as near-threatened on the IUCN Red List of Threatened Species. |
| Functional significance | The shoal is an important refuge site for commercial and non-commercial species. Its animal forests are dominated by filter-feeders known for their higher-level roles in the ecosystem, such as nutrient cycling and pelagic–benthic coupling. Some of them provide fundamental links between Mediterranean mesophotic populations. |
| Fragility | Emphasized by the modifications of the seafloor integrity due to the fishing gear and by the biological characteristics of the structuring species, which suffer from entanglements, breakages, necrosis, and detachment. |
| Peculiar life-history traits | Reflect the occurrence of slow-growing canopy-forming species, particularly gorgonians and black corals. |
| Structural complexity | Supported by the presence of complex topographic features and by the significant concentration of many canopy-forming organisms and commensal or closely associated species. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enrichetti, F.; Bavestrello, G.; Cappanera, V.; Mariotti, M.; Massa, F.; Merotto, L.; Povero, P.; Rigo, I.; Toma, M.; Tunesi, L.; et al. High Megabenthic Complexity and Vulnerability of a Mesophotic Rocky Shoal Support Its Inclusion in a Mediterranean MPA. Diversity 2023, 15, 933. https://doi.org/10.3390/d15080933
Enrichetti F, Bavestrello G, Cappanera V, Mariotti M, Massa F, Merotto L, Povero P, Rigo I, Toma M, Tunesi L, et al. High Megabenthic Complexity and Vulnerability of a Mesophotic Rocky Shoal Support Its Inclusion in a Mediterranean MPA. Diversity. 2023; 15(8):933. https://doi.org/10.3390/d15080933
Chicago/Turabian StyleEnrichetti, Francesco, Giorgio Bavestrello, Valentina Cappanera, Mauro Mariotti, Francesco Massa, Lorenzo Merotto, Paolo Povero, Ilaria Rigo, Margherita Toma, Leonardo Tunesi, and et al. 2023. "High Megabenthic Complexity and Vulnerability of a Mesophotic Rocky Shoal Support Its Inclusion in a Mediterranean MPA" Diversity 15, no. 8: 933. https://doi.org/10.3390/d15080933