
Trophic Diversification Out of Ancestral Specialization: An Example from a Radiating African Cyprinid Fish (Genus Garra)


1
Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences, 152742 Borok, Russia
2
Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, 119071 Moscow, Russia
3
Eco-Analytical Laboratory, Cherepovets State University, 162600 Cherepovets, Russia
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Diversity 2022, 14(8), 629; https://doi.org/10.3390/d14080629
Submission received: 9 June 2022
/
Revised: 1 August 2022
/
Accepted: 3 August 2022
/
Published: 6 August 2022
(This article belongs to the Special Issue Aquatic Biodiversity: Evolution, Taxonomy and Conservation)
Abstract
:Trophic resource partitioning is one of the main ecological mechanisms of adaptive radiation. The Garra is a highly specialized periphyton feeder that has widened jaws equipped with a horny cutting scraper. In a river located in the Ethiopian Highlands in East Africa, a diversification of Garra composed of six sympatric ecomorphs which were strikingly diverse in trophic morphology was revealed. A hypothesis on trophic resource partitioning was tested using data on diet composition, gut length, and stable isotopes. The obtained results confirmed the trophic diversification of Garra ecomorphs. Three feeding modes were revealed: (i) periphytonophagy, (ii) mixed periphytonophagy and zoophagy, and (iii) zoophagy. The periphyton feeders had a long gut and were enriched in δ13C values compared to the shorter gut and lowered δ13C values in the zoophagous ecomorphs. Therefore, Garra could respecialize out of its ancestral specialization. This finding does not support the generalists-to-specialists hypothesis on the origin of specializations, and suggests that Liem’s paradox is a more common phenomenon. In the case of specialists, we assume that new ecological opportunities can be ‘visible’ to specialists if they are preceded by relaxed selection constrains that lead to the widening of the ecological/morphological plasticity to jump out of a canalized mode of ancestral specialization.
1. Introduction
Adaptive radiation is the evolution of ecological diversity within a rapidly multiplying lineage [1]. Fishes, being one of the most diverse groups of vertebrates, display numerous patterns of adaptive radiation, e.g., [2,3,4,5,6,7,8,9,10,11]. Trophic resource partitioning is one of the main ecological mechanisms of adaptive radiation. Trophic diversification is accommodating of many fishes’ adaptive radiation. For instance, the sympatric diversification of neotropical cichlids from Central and South America is based on trophic resource partitioning [12,13,14]. It has been suggested that trophic radiation preceded the diversification of African cichlids in Lake Tanganyika [15,16]. Adaptive radiation and diversification bursts based on trophic divergence were found not only in cichlids but also in other fish groups. Among these, white fishes and Arctic charrs are some of the best known examples [8,9,17,18,19,20,21]. Adaptive radiation driven by trophic resource partitioning was revealed also among cyprinid fishes [3,4,22,23,24,25,26,27,28]. The brightest example of trophic adaptive divergence among cyprinids falls to the genus Labeobarbus Rüppell, 1835, in Lake Tana (East Africa, Ethiopia), which was represented by 15 sympatric species/ecomorphs [3,4,29,30,31]. Remarkably, the majority of adaptive radiations of fishes have been documented in a lacustrine environment. The riverine environment has been commonly considered inappropriate for adaptive radiation due to various reasons (e.g., the instable hydrological regime, less diversified habitats, increased gene flow, etc.). At the same time, during the last two decades, novel examples of fish adaptive radiations have been revealed in rivers [14,27,32,33,34,35,36,37,38]. Among others, the Ethiopian Highlands seem to be a hotspot of riverine diversifications, being a home for several young fish radiations, i.e., Labeobarbus with four independently evolved riverine radiations, each represented by four–six sympatric ecomorphs [33], and the genus Garra Hamilton, 1822, represented by six young species/ecomorphs radiated in the Sore River, White Nile basin [34,39].
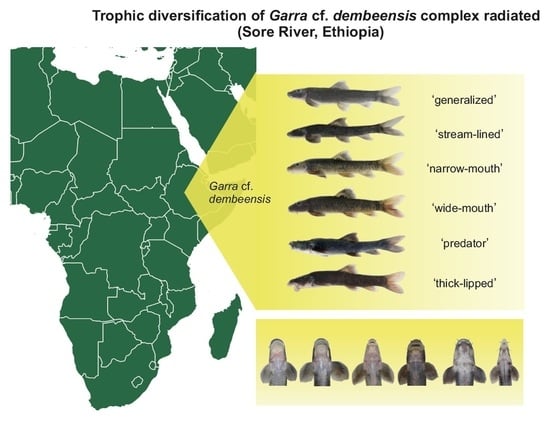
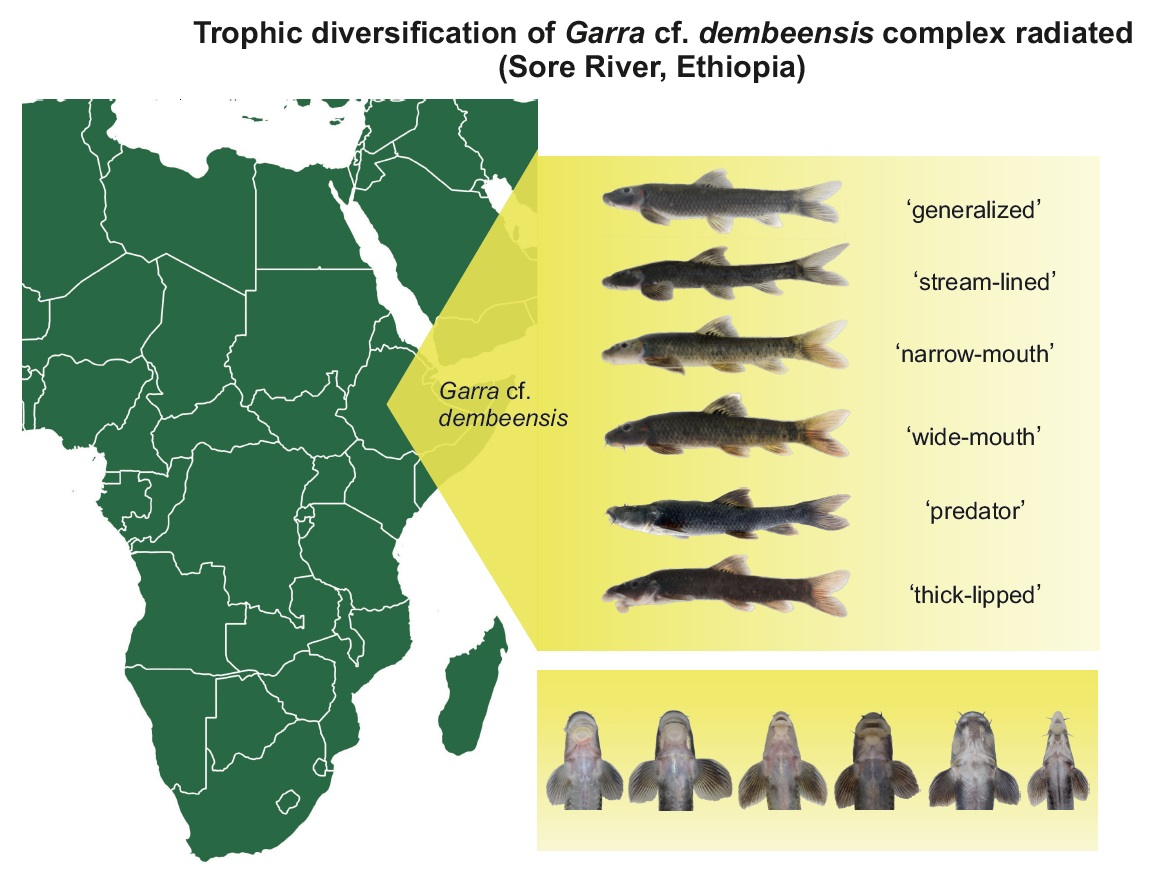
Garra are fish of moderate size (commonly up to 20 cm) with a sucking gular disc, and mostly inhabit the rhithron zone of river systems [40]. The genus Garra is species rich (ca. 90 species—[41]) and widely distributed from Southeastern Asia to West Africa. Garra species are highly specialized algae-scrapers that graze periphyton from rocks and stones using broadened jaws equipped with horny scrapers [42,43,44]. However, a recently discovered riverine assemblage of Garra cf. dembeensis from the Sore River (Ethiopia) composed of six sympatric ecomorphs displayed strikingly diverse trophic morphologies (Figure 1) that suggest trophic diversification and resource partitioning [39]. Genetic study confirmed a scenario of the recent evolution of this radiation, given the lack of mtDNA-divergences but the presence of genomic distinctness for all of the sympatric ecomorphs revealed in the genome-wide SNP study [34]. At the same time, little is known on the ecological drivers of this adaptive radiation, i.e., whether Garra ecomorphs partition trophic resources as it might be proposed based on phenotypic divergence. Therefore, our study aimed to check a hypothesis on the trophic diversification of six co-occurring ecomorphs of Garra cf. dembeensis from the Sore R. (White Nile basin) which evolved during recent adaptive radiation. For this purpose we studied the diet, gut length, and stable isotope (δ13C and δ15N) signatures of sympatric ecomorphs.
2. Materials and Methods
2.1. Study Area and Sampling
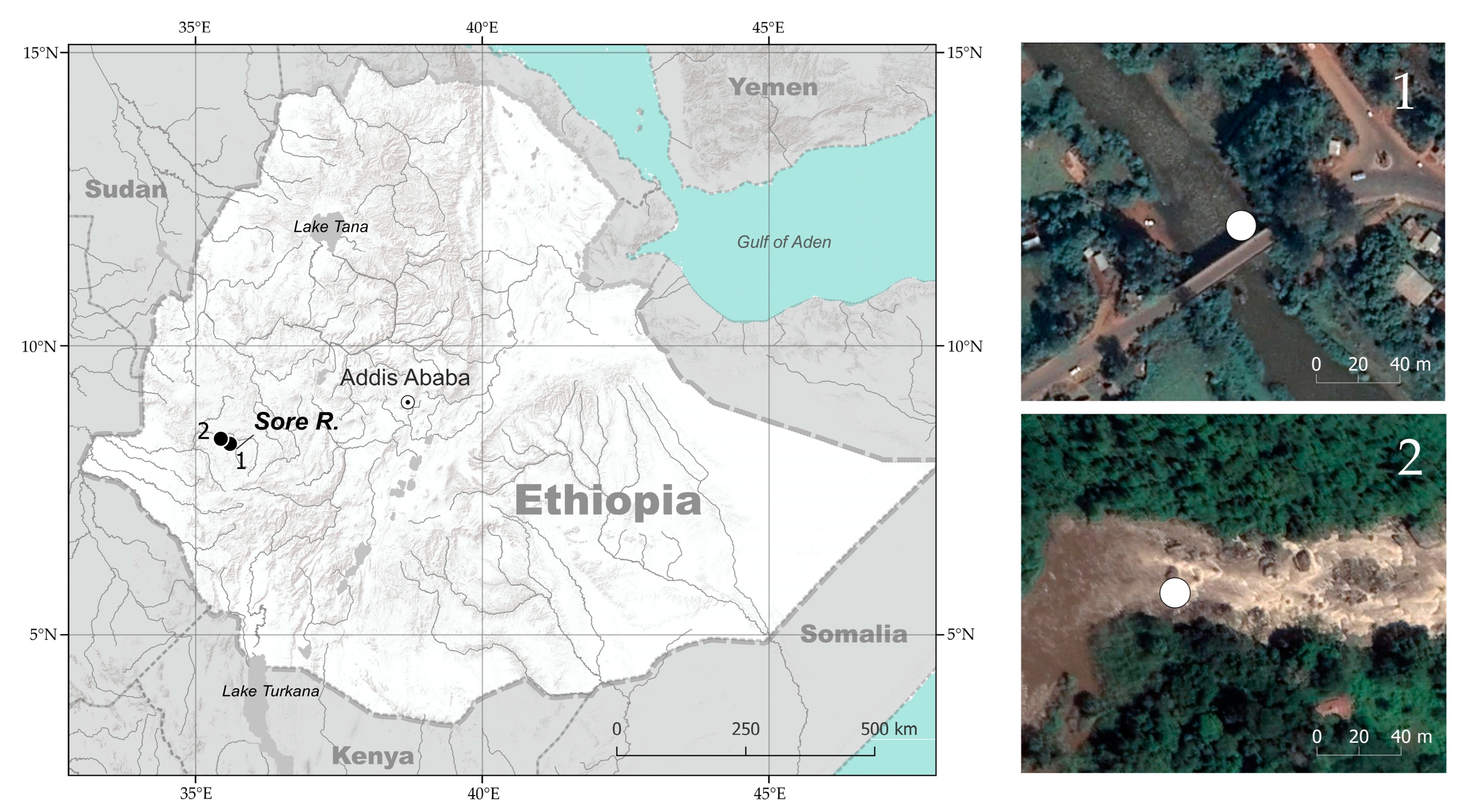
The Sore River is a tributary of the Baro-Akobo-Sobat drainage in the White Nile basin (Ethiopia, East Africa). The region is covered by moist Afromontane forest that has been rapidly shrinking in recent decades due to agricultural development [45]. The Sore has a length of ~160 km; its catchment area is ~2000 km2, and is characterized by a substantial seasonal variation of rainfall (a dry season from December to March) [46]. The elevation difference between the Sore source (an altitude of ~2215 m above sea level) and its confluence with the Gabba River (alt. 963 m asl) is >1.2 km. The Sore River basin shares drainage boundaries with the Blue Nile in the northeast and Omo-Turkana in the southeast. We sampled the middle reaches of the Sore River at two sites: (i) at the City of Metu (8.3116° N, 35.5983° E, alt. 1550 m asl) and (ii) ~35 km downstream along the river course (8.3988° N, 35.4383° E, alt. 1310 m asl). The river width at the rapids sampled was 20–40 m at the beginning of the rainy season, with a depth < 1 m, and the bottom consisted of rocks and large boulders. Garra samples from the Sore River were collected using an electrofishing device (LR-24 Combo Backpack, Smith-Root), a cast, and frame nets in June 2012 and April 2014. The fish fauna of the river segment under consideration included (apart from Garra spp.) Labeobarbus cf. intermedius Rüppell 1835, Enteromius cf. pleurogramma (Boulenger 1902), Labeo cf. cylindricus Peters 1852, Labeo forskalii Rüppell 1835, Chiloglanis niloticus Boulenger 1900 (at the lower site only), and introduced Coptodon zillii (Gervais 1848) [34]. The fish sampling was conducted under the umbrella of the Joint Ethiopian–Russian Biological Expedition (JERBE) with the permissions of the National Fisheries and Aquatic Life Research Center (NFALRC) under the Ethiopian Institute of Agricultural Research (EIAR) and the Ethiopian Ministry of Science and Technology. The fish were killed with an overdose of MS-222 anaesthetic [47], and were first preserved in 10% formalin and then transferred to 70% ethanol for the study of their gut content. In total, 83–182 specimens were analyzed, depending on the type of analysis (Table 1). The designation of ecomorphs is taken from [34]: No. 1, «generalized»; No. 2, «stream-lined»; No. 3, «narrow-mouth»; No. 4, «wide-mouth»; No. 5, «predator»; No. 6, «thick-lipped». The map of the sampling sites (Figure 2) was created in QGIS Version 3.16.4-Hannover; the satellite images were taken from Google Maps [48].
2.2. Diet and Stable Isotopes
Intestines were taken from the body cavity of 182 preserved specimens of all of the ecomorphs, and were measured using a ruler to the nearest 1 mm. Gut length is a reliable indicator of diet. Carnivorous fishes generally have a shorter gut compared to the herbivorous ones, e.g., [49]. The sample size for each ecomorph is provided in Table 1. The SL of the examined individuals varied from 39 to 193 mm; one individual of ecomorph 5 had an extreme length of 193 mm. The ratio of the gut length (GL) to SL was used for subsequent analyses. Diet was assessed for the same individuals whose gut length was measured. Gut content was dried on filter paper and weighed using balance Pioneer PX84/E with 0.0001 g accuracy. The diet particles were identified using a Motic DMW-143-N2GG stereomicroscope (100–400× magnification) and Olympus CX41 microscope (100–1000× magnification). The diet components were grouped into: (i) periphyton, (ii) invertebrates, (iii) macrophytes, and (iv) others. Here, we follow the periphyton definition from Ewart-Smith [50]: periphyton consists mainly of freshwater algae, cyanobacteria and prokaryotes such as bacteria, fungi and protozoa that grow on rock substratum in all lotic systems. The group ‘Invertebrates’ included the larvae of amphibiotic insects, arachnids (Hydrachnidia), and their fragments. The group ‘Macrophytes’ included any fragments of plants—such as leaves, stems or seeds. The group ‘Others’ was composed of unidentified remnants of detritus and mineral particles (e.g., sand). A composite measure of diet, an index of relative importance (IR) [51,52], was used to assess the contribution of different components to a diet. The IR index was calculated as follows: , where Fi = the frequency of occurrence of each food species, and Pi = a part by weight; the value of i itself changes from 1 to n (n = the part of food organisms in the food bolus).
For stable isotope analyses, white muscle tissue from the dorsal side of the body under the dorsal fin was sampled from freshly collected specimens. White muscle samples were dried at 60 °C for subsequent stable isotope analyses. The samples were weighed using a Mettler Toledo MX5 analytical balance (Mettler Toledo, Columbus, OH, United States) with 2 μg accuracy, and were wrapped in tin capsules. The weight of the fish tissue samples varied from 250 to 500 μg. Stable isotope analysis was conducted at the Joint Usage Center of the Severtsov Institute of Ecology and Evolution (RAS), Moscow. Briefly, a Thermo Delta V Plus continuous-flow IRMS was coupled with an elemental analyzer (Flash 1112) and equipped with a Thermo No-Blank device. The isotopic composition of N and C was expressed in a δ notation relative to the international standards (atmospheric nitrogen and VPDB, respectively): δX (‰) = [(Rsample/Rstandard) − 1] × 1000, where R is the ratio of the heavier isotope to the lighter. The samples were analyzed with a reference gas calibrated against the International Atomic Energy Agency (IAEA) reference materials USGS 40 and USGS 41 (glutamic acid). The measurement accuracy was ± 0.2 δ units. Along with the isotopic analysis, the nitrogen and carbon content (as %) and C/N ratios were determined in all of the samples. In total, 83 white-muscle samples were analyzed.
For the lipid normalization of the samples with C/N (mass/mass) values higher than 3.50, we applied the equation δ13C corrected = δ13C untreated −3.32 + 0.99 C:N, as suggested by Post et al. [53]. In order to avoid the influence of seasonal or other factors, we used only samples collected in 2014 for the stable isotope analyses.
2.3. Statistical Analyses
The comparison of multiple samples obtained from different Garra ecomorphs in terms of gut length and stable isotope values (δ13C and δ15N) was performed using the Kruskal–Wallis test with a post-hoc Dunn’s test. Several R packages realized in R-studio v. 4.1.1 [54] were used for the statistical analyses and plot construction: the summarytools library [55] was used to obtain basal descriptive statistics; the ggplot2 library [56] was used to calculate the Pearson correlation, and to build the violin boxplots; and the kwAllPairsDunnTest function in the PMCMRplus library [57] was applied for the Kruskal–Wallis post hoc Dunn’s test. The package SIBER v.2.1.6 [58] was used to assess the differences in the isotopic trophic niche features. The total convex hull areas (TA), core trophic niche breadths, and correction for variable sample sizes (SEAc) were estimated. The total trophic overlap values for 95% TA were estimated using nicheROVER, a method that is insensitive to the sample size and incorporates statistical uncertainty using Bayesian methods [59].
3. Results
3.1. Gut Length
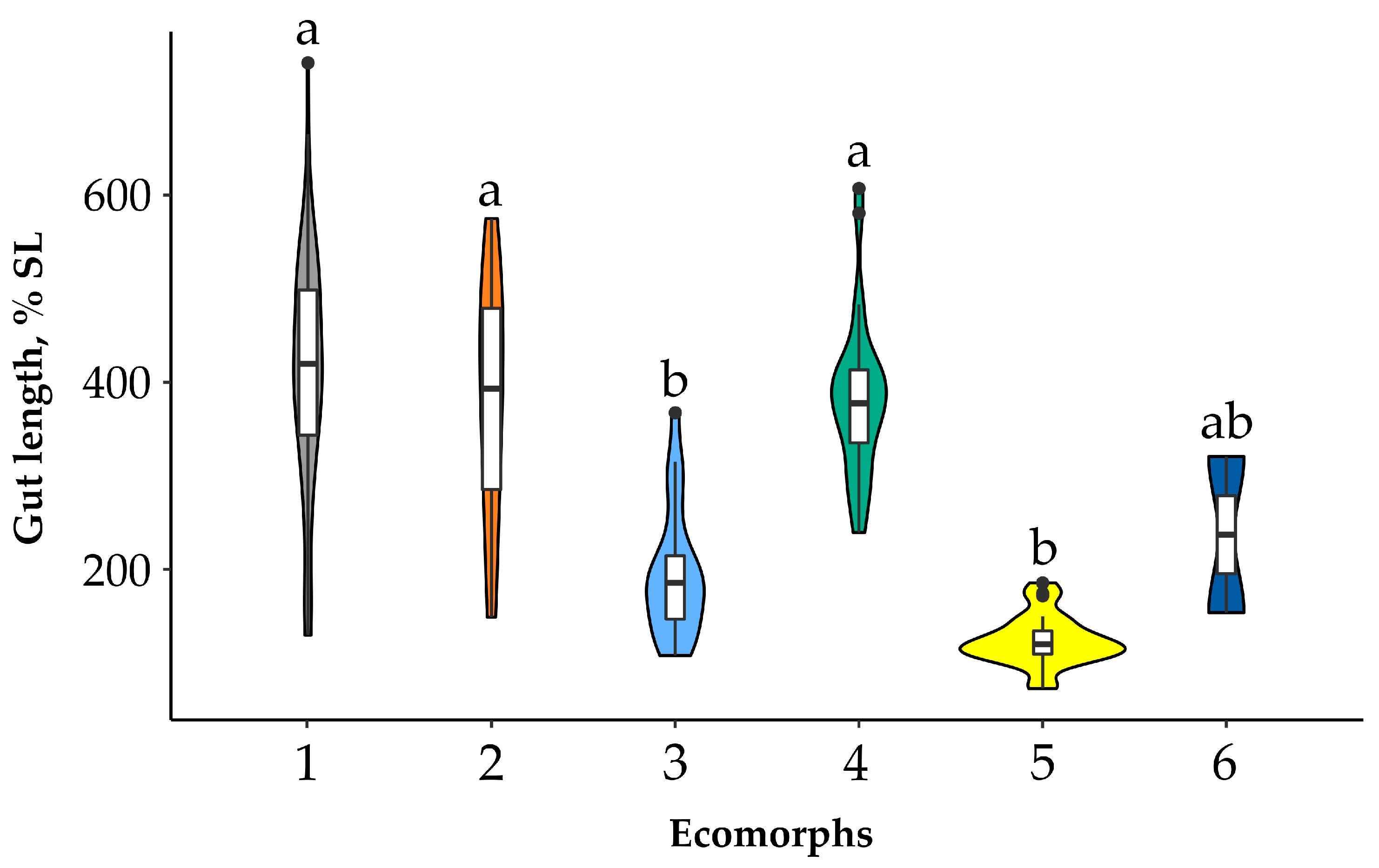
All of the Garra ecomorphs had a gut length varying from 72.5 to 741.4% SL (Figure 3). Based on gut length, ecomorphs can be divided for two groups: (i) long guts were detected in ecomorphs no. 1 (mean = 409.3% SL), no. 4 (359.6% SL), and no. 2 (379.8% SL), and (ii) short guts were detected in ecomorphs nos. 3 and 5 (194.7 и 123.8% SL, respectively). Ecomorph no. 6 had an intermediate gut length (237.1% SL). The ecomorphs with a longer gut significantly differed from the ecomorphs with a shorter gut (p < 0.001—Figure 3). Relative gut length was positively correlated with body length in ecomorphs nos. 1 and 4 (p < 0.05, Figure S1). The gut length of ecomorph no. 5 had a negative correlation with its body length (although non-significant—Figure S1).
3.2. Diet
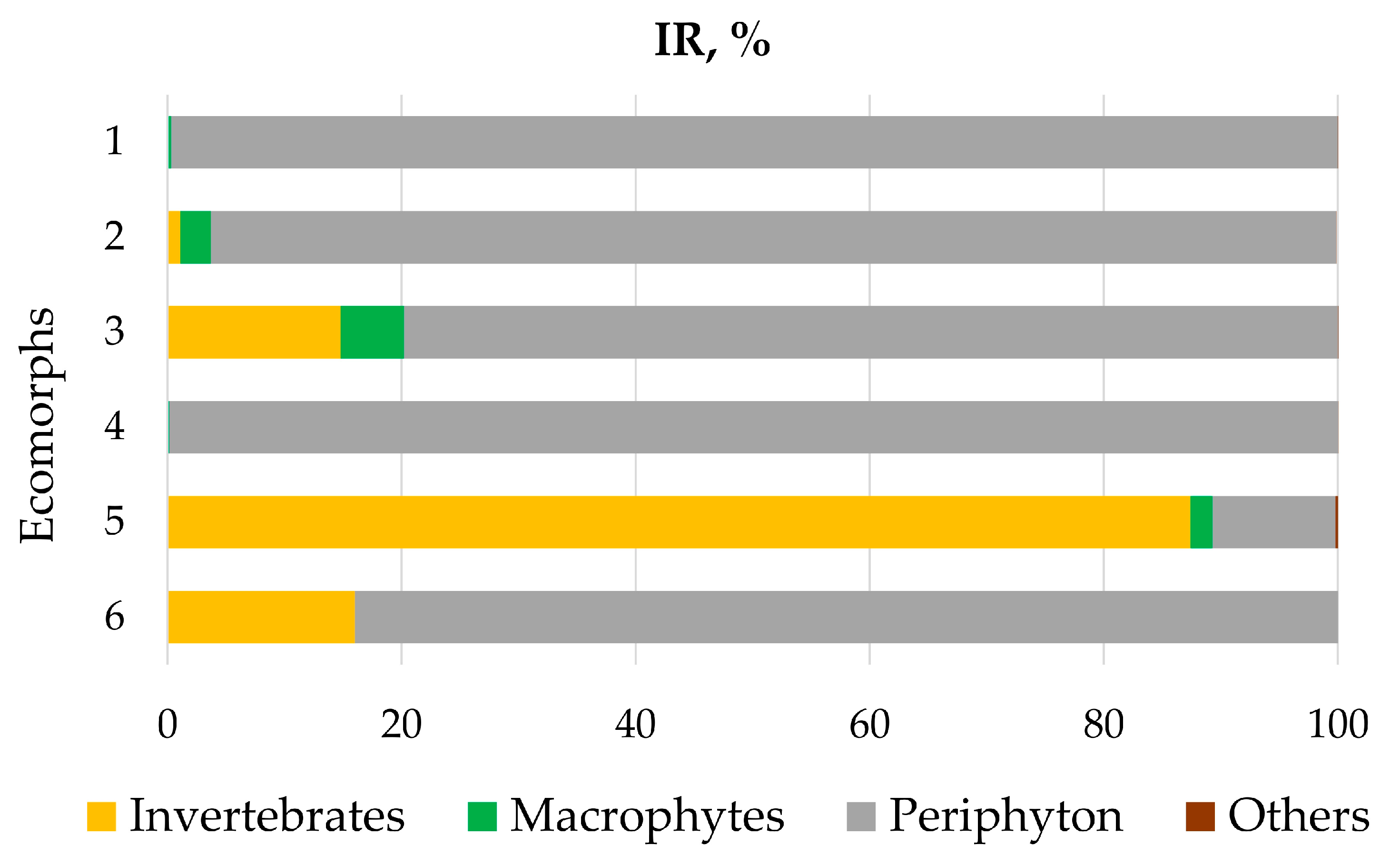
The diets of the six sympatric Garra ecomorphs are diverse, and include (i) periphyton represented by diatoms (Bacillariophyceae), green algae (Chlorophyta) and charophytes (Cosmarium); (ii) invertebrates (mainly insects: Diptera, Trichoptera, Ephemeroptera, Coleoptera); (iii) macrophytes; and (iv) others. According to the IR index, the ecomorphs were divergent in the composition of various food items (Figure 4). We will now consider the diet of certain ecomorphs in more detail.
Regarding ecomorph no. 1, 68 guts were filled (85%) while 12 (15%) were empty. The average gut fullness was 56%. Periphyton significantly predominated (IR = 99.7%), and was detected in 80% of the individuals studied. Macrophytes were detected in 16% of the examined guts but with a very low proportion in relation to periphyton (IR = 0.2%). The same was found for amphibiotic insects: their fragments were detected in 20% of the examined guts, but with IR = 0.1%. Among insects, larvae of Chironomidae and Trichoptera as well as imago of water beetles Coleoptera were recorded.
Regarding ecomorph no. 2, 21 guts (91%) were filled with food items and two (9%) were empty. The average gut fullness was 68%. Periphyton greatly dominated (IR = 96.2%) among the other items, and was detected in 87% of the individuals studied. Macrophytes were detected in 39% of the examined guts (IR = 2.6%). Larvae of amphibiotic insects were found in 60% of the guts (IR = 1.1%). Among these were Trichoptera (2–4 larvae per a gut) Ephemeroptera (up to 53 larvae), Chironomidae (up to 19 larvae), and Simulidae (up to 9 larvae).
Regarding ecomorph no. 3, 18 guts (82%) were filled by food items and four (18%) were empty. Average gut fullness was 40%. Periphyton was still a major item (IR 79.8%); it was found in 13 guts (59%). However, portion of larvae of amphibiotic insects was notably heightened compared to ecomorphs 1–2. Insects recorded in 77% of guts (IR = 14.8%). The following taxonomic groups of insects were detected: Trichoptera (one to five larvae per a gut), Chironomidae (up to 235 larvae), Ceratopogonidae (sporadically), and Simuliidae (one to three larvae). Arachnids (Hydrachnidia, two to five imago) were recorded in two guts.
Regarding ecomorph no. 4, 28 guts (97%) were filled by food, while only one (3%) was empty. The average gut fullness was 38%. Periphyton greatly dominated (IR = 99.9%), being detected in 93% of the examined guts. The portion of larval insects (Trichoptera, Chironomidae, Ceratopogonidae) was exceptionally low (IR = 0.1%), with occurrence in 31% of the examined guts. Macrophyte fragments were found in 20% of the guts, and their portion was extremely low (IR < 0.1%).
Regarding ecomorph no. 5, 23 guts (88%) were filled by food and three (12%) were empty. The average gut fullness was minimal (20%) among all of the ecomorphs. Contrary to the previous ecomorphs, the diet was composed mainly of the larvae of amphibiotic insects (IR = 87.4%), which were recorded in 85% of the investigated guts. The following taxonomic groups of insects were detected: Ephemeroptera (up to 15 larvae per a gut) and Chironomidae (one to two larvae), and larvae of Trichoptera and Simuliidae were recorded sporadically. Arachnids (Hydrachnidia) were also found singly. One gut contained fragments of Hymenoptera imago. The portion of periphyton was low (IR = 10.5%), with occurrence in 23% of the investigated guts. Macrophytes were detected sporadically (IR = 1.9%), and were represented by seeds and fragments of plant tissues. Only 19% of the guts contained macrophytes.
Regarding ecomorph no. 6, two individuals were studied. Gut fullness was 32 и 50%. One gut contained periphyton, with a predominant portion of diatoms (Bacillariophyceae). Another gut contained mainly insects (Diptera, n = 121 larvae), and arachnids (Hydrachnidia, n = 3 imago). Plant fragments were occasionally detected.
Briefly, ecomorphs 1, 2 and 4 may be considered periphyton feeders, ecomorphs 3 and 6 may be considered as periphyton feeders with a significant portion of benthic invertebrates, and ecomorph 5 may be considered to be zoophagous.
3.3. Stable Isotopes
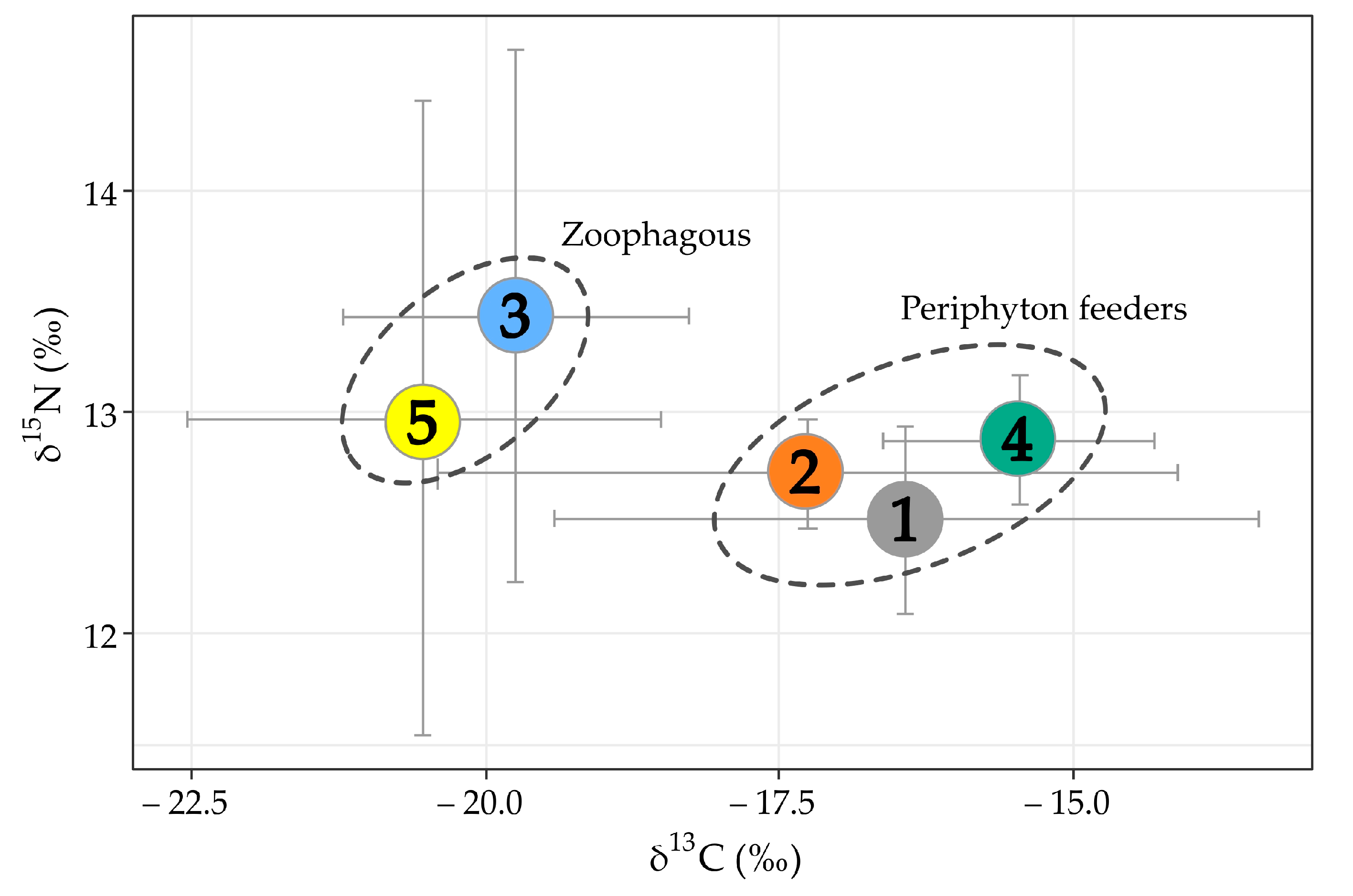
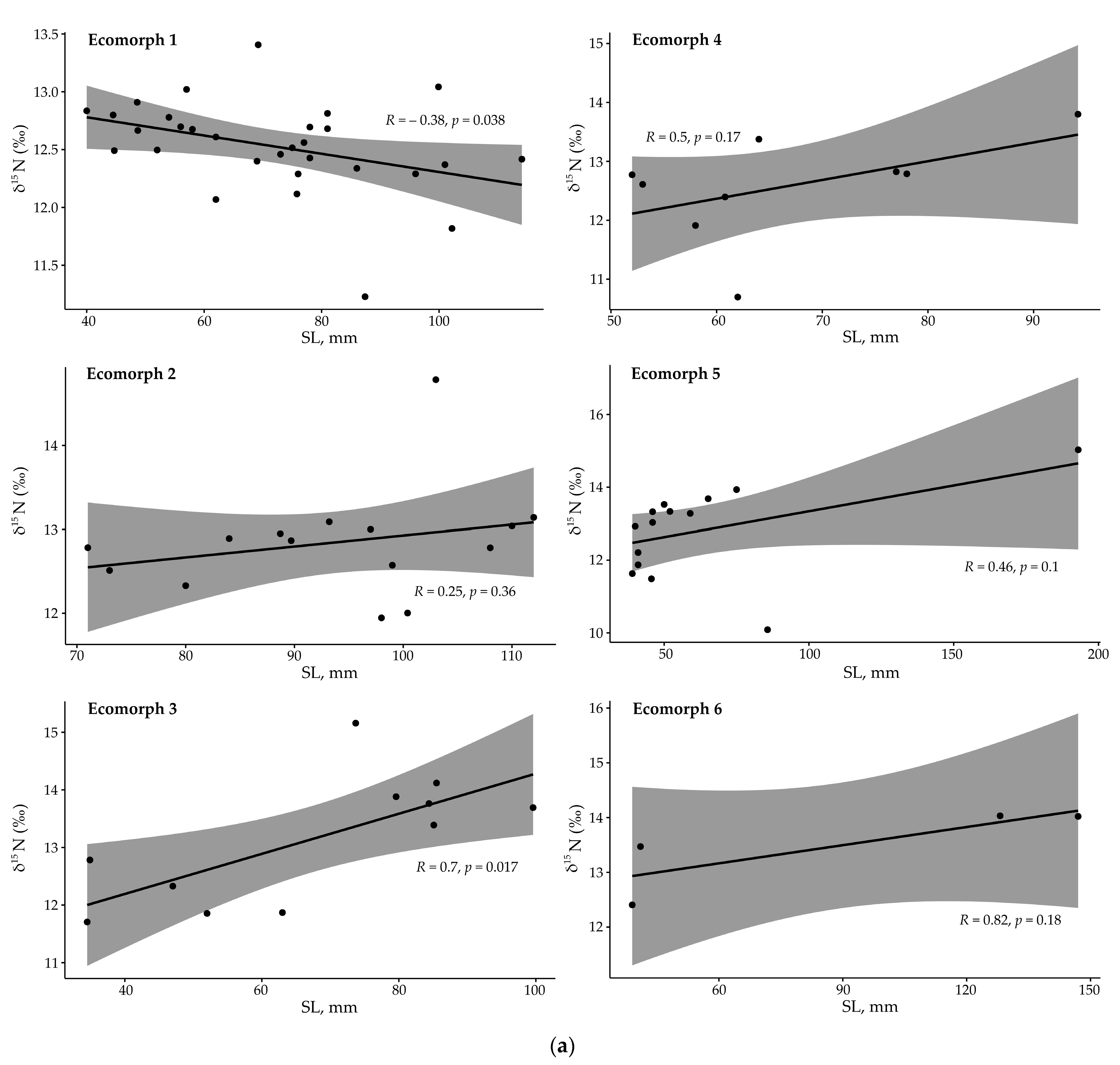
The first dataset based on wide range of body sizes (34.5–193.0 mm SL, n = 83) did not reveal significant divergence between the ecomorphs (Figure S2) due to high differentiation between small- and large-sized individuals. Upon excluding the too-small and too-large individuals, the second dataset included individuals from 50 to 90 мм SL (n = 44), and showed that some of the ecomorphs are significantly divergent in their δ13C values (Figure 5). In particular, periphyton feeders—ecomorphs nos. 1, 2, and 4—had larger δ13C values of –17.3‰, –16.4‰ и –15.5‰ (means), respectively, compared to ecomorphs nos. 3 and 5 (Figure 5). The latter ecomorphs with an increased (no. 3) or predominating (no. 5) portion of benthic organisms had lowered δ13C values: –20.5‰ and –19.7‰, respectively. Significant divergencies were revealed in δ13C values between ecomorphs nos. 1 and 5 (p < 0.01), and between nos. 1 and 3 (p < 0.05). It is noteworthy that the zoophagous ecomorphs (3 and 5) had slightly increased (although non-significant) δ15N values (13.0–13.4‰) compared to the periphyton feeders (12.5–12.9‰). Significant changes in δ15N values along the body length were revealed in ecomorph 1 (negative correlation; p < 0.05; Figure 5), and ecomorph 3 (positive correlation; p < 0.05; Figure 6a). Ecomorph 2 presented a significant negative correlation of δ13C values with body length (p < 0.05), while ecomorph 6 displayed a near-significant negative correlation of δ13C values.
Values of δ13C and δ15N correlate with body size and gut length in some ecomorphs (Figure 6, Figures S3 and S4). In particular, ecomorph 1 had a negative correlation of δ15N values with body size (p < 0.05), while ecomorph 3 had a positive correlation (p < 0.05). Only ecomorph 2 had a (negative) correlation of δ13C with body size (p < 0.05). In relation to gut length, ecomorph 3 and ecomorph 4 had negative (p < 0.01) and positive (p < 0.05) correlations of δ15N values, respectively. At the same time, ecomorphs 1, 2, and 4 (all are periphyton feeders) had a positive correlation of δ13C values with gut length (p < 0.01–0.05) (Figure 6).
The ‘thick-lipped’ ecomorph was represented by four individuals in the stable isotope analyses, and did not display significant differences or correlations. However, it had high δ15N values (Figure S3).
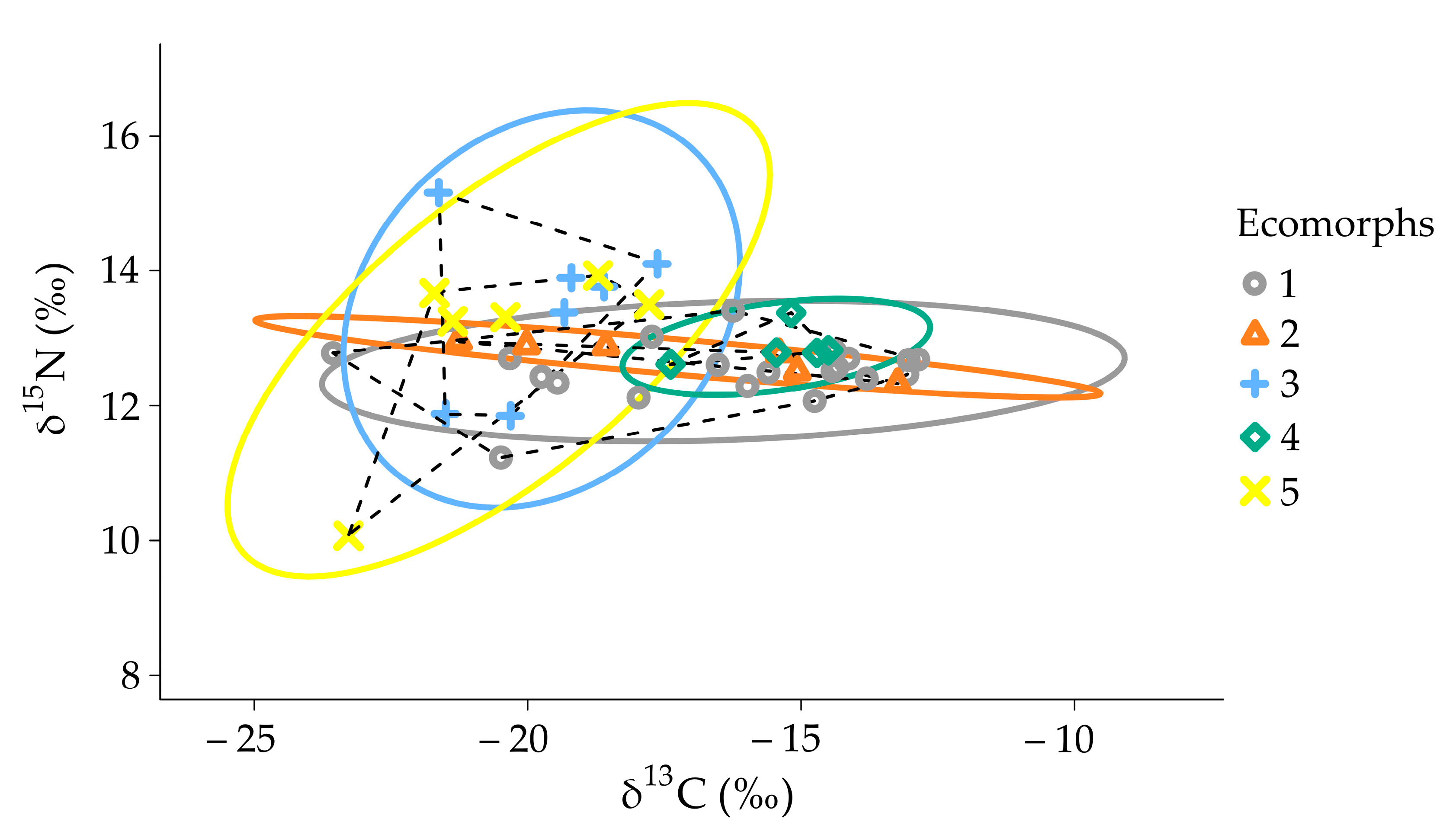
The total area (TA), standard ellipse area (SEA), and corrected standard ellipse area (SEAc) were measured for two datasets for all of the individuals (n = 83) of a broad size range (SL = 39–193; Figure S5, Table S1) and for the reduced dataset (n = 44) with a size range 50–90 mm SL (Figure 7; Table 2). The average niche area, SEAc, significantly overlapped between the ecomorphs when all of the individuals were included, which was supposedly due to ontogeny shift in the dietary preferences. Based on the reduced size range (50–90 mm SL), the largest SEAc values were in ecomorph 5 (8.16) and 3 (6.54), while the smallest SEAc values were in ecomorphs 2 (1.34) and 4 (1.29) (see Table 2). Generally, SEAc was larger in the zoophagous ecomorphs compared to periphyton-feeding ecomorphs (Figure 7, Table 2).
Niche overlap estimates revealed an extensive overlap of the niche region of ecomorph 1 with other periphython-feeding ecomorphs (95.74% in ecomorph 4, and 87.51% in ecomorps 2), while ecomorphs 4 and 2 had lower niche overlapping with ecomorph 1 (30.55% and 29.03% respectively) (Table 3). At the same time, the probability that zoophagous ecomorphs (3 and 5) are included within the niches of periphyton-feeding ecomorphs was significantly lowered (1.22–35.66% in various comparisons—Table 3). The niche overlapping in zoophagous ecomorphs is moderately high (76.33–78.96%).
When all of the size individuals were analyzed (n = 83), niche overlap estimates revealed more extensive overlapping of niches although the tendency of fewer overlapping between periphyton feeding and zoophagous ecomorphs remain (Table S2).
4. Discussion
Our study revealed trophic diversification of the Garra cf. dembeensis complex, which radiated in the Sore River (White Nile basin) and was composed of six ecomorphs. The data on diet coupled with gut length and stable isotope ratios strongly suggest trophic resource partitioning. According to our results, three feeding modes can be proposed for this Garra complex: (i) periphytonophagy (ecomorphs 1, 2, and 4), (ii) a mixed mode of periphytonophagy and zoophagy (mainly insectivory) in ecomorph 3, and (iii) zoophagy (ecomorph 5). The feeding mode of ‘thick-lipped’ ecomorph (no. 6) was not yet substantiated due to low sample numbers, but combined data on its diet, gut length, stable isotopes, and type of mouth phenotype may point to its more zoophagous habits. The periphyton feeders had a significantly longer gut compared to partially or predominately zoophagous ecomorphs, which confirmed the results of previous studies that specialized algae eaters have elongated guts [49,60,61]. Remarkably, the divergent feeding modes (periphytonophagy and zoophagy) are masked when only the δ15N values compared due to their high level in periphyton feeders. Garra’s periphython feeding ecomorphs had higher δ15N values (12.5–12.9‰) than the sympatric labeobarbs Labeobarus sp. with an omnivorous diet (δ15N mean = 10.5‰; n = 34; our unpublished data). The obtained results look like an anomaly, given the well-accepted results that algae eaters with elongated guts have low trophic position, i.e., characterized by low δ15N values (e.g., Tanganyikan cichlids—[49,62]). Generally, little is known on the trophic position of periphyton feeders. Our results coincide with recently obtained data on the high δ15N values in another periphyton-scraping feeder, Labeobarbus beso, the nitrogen level of which matched such for sympatrically co-occurring piscivorous fish [28]. Periphyton is a community mostly composed of algae (mainly green algae and diatoms) but also contains detritus, bacteria, fungi, protists, zooplankton and other invertebrates hiding within algal mats [63]. Algal periphyton is rather rich in proteins [64,65,66,67], which may explain the remarkably high δ15N values in Garra periphyton feeders. Despite periphyton being protein-rich, it is not easily digested. Apparently, periphyton feeders are adapted for efficient digestion, having a longer gut and a complex of physiological, biochemical and microbiome adaptations (e.g., [68]). Being indistinguishable from zoophagous ecomorphs in δ15N values, periphyton feeders of Garra in the Sore River could be well differentiated from zoophagous sympatric ecomorphs by higher δ13C values that confirm an algal diet, and especially diatoms [62,69]. Generally, the sympatric Garra ecomorphs under study are characterized by a broad range of δ13C values (from –24.06‰ to –12.43‰), which is rather uncommon for sympatrically co-occurring fishes, e.g., [27,70].
Despite the fact that Garra are fishes highly specialized in their foraging strategy, being periphytonophagous [40,42], Garra cf. dembeensis from the Sore River could give rise to a new kind of trophic specialization out of its ancestral feeding mode. We consider the obtained results further in the context of ecological speciation during adaptive radiation, and will try to answer the question of how a phenotypically highly specialized periphyton feeder could radiate out of its specialization.
It is generally accepted that an ancestor radiating into an array of specialized forms had a generalized feature. Indeed, theoretically, the generalists-to-specialists hypothesis is much more plausible considering that populations of generalized species more likely immigrate to a new and distant location with wide dispersal compared to (narrowly) specialized species, the distribution of which may be restricted by a certain type of trophic resource and its restriction from the utilization of other resources. However, related studies did not support the generalists-to-specialists hypothesis (summarized in [1]) assuming that specialization is not an impediment to ecological diversification [71].
In relation to trophic fish ecology, Liem revealed that even species with highly specialized trophic morphologies have dietary flexibility [72]—this phenomenon was later named “Liem’s paradox”. There are many examples corroborating Liem’s paradox, making it a well-established phenomenon among fishes (e.g., [28,49,68,73,74,75,76]). First, this phenomenon reduces the prediction of diet by phenotype, and second, it provides evidence for the unexpectedly greater trophic plasticity of specialists that allows them to more efficiently utilize available resources in some circumstances. This flexibility may result in a relaxation of specialization that, in turn, may facilitate a respecialization.
The diversification of sympatric Garra ecomorphs in the Sore River is particularly interesting because since this diversity evolved in situ [34] and includes two modes of radiation. First, it is a further diversification within ancestral specialization: three ecomorphs were periphytonophagous, having a well-developed gular sucking disc (the ancestral state of character). Two of these, ecomorphs 1 and 2, were similar in all of the studied traits (diet, gut length, stable isotope ratios) but differed strikingly in body shape (see Figure 1), which implies that they prefer different ecotopes. Ecomorph 2, possessing the same shape of the gular disc, has much more slender body and more developed fins (Figure 1), which suggests its adaptation to rapid habitats. Ecomorph 4, also being periphytonophagous, is somewhat divergent from ecomorphs 1 and 2. In particular, it has a modified gular disc and greatly developed labellum (sensu [40]). In addition, it has another pattern of correlation of stable isotopes values with gut length as well as a correlation of δ13C with δ15N values compared to other periphyton feeders (see Figures S6–S8), which that suggests that it might occupy another ecological niche within a periphyton-feeding foraging strategy. As such, this part of Sore’s radiation can be considered a radiation within Garra’s ‘key innovation’—a gular sucking disc accommodated by widened jaws modified into scrapers. This ‘equipment’ corresponds to the algae-scraping mode of feeding.
The remaining three ecomorphs (nos. 3, 5, and 6) manifest modes of feeding other than periphyton consumption, with a transition to zoophagy that means respecialization despite they still retaining jaw-scrapers. Ecomorphs 3 and 6 have significantly modified lower jaw-scrapers that are drastically narrower and shovel-like. A similar shape of lower jaw-scraper is known in the specialized benthivorous cyprinid fishes Sarcocheilichthys lacustris (Dybowski, 1872) from the Eurasian Far East and Exoglossum maxillingua (Le Sueur, 1817) from North America [77,78]. The gular sucking disc of zoophagous ecomorphs is significantly modified or even completely reduced (ecomorph 5). The rare ecomorph 6 has a ‘thick-lipped’ mouth phenotype that was never found before for the whole of the genus Garra, despite its species richness (ca. 190 species) and wide distribution across the Old World. Hypertrophied lips may be considered as a morphological novelty within this genus. In relation to the evolutionary origin of Sore’s Garra, the phylogenomic analysis of Ethiopian Garra [34] showed the following: (i) Sore’s ecomorphs originated from an ancestor with a normally developed gular disc inhabiting the same White Nile system within Ethiopia; (ii) Sore’s ecomorphs are of recent divergence, with a basal position of periphyton feeders on the tree, while the respecialized (zoophagous) ecomorphs are the derived ones; (iii) the ‘thick-lipped’ phenotype has originated due to a past hybridization of other ecomorphs originating in situ. These results strongly suggest the local respecialization of Garra during its adaptive radiation within the periphyton feeder lineage. How is this possible?
The ecological theory of adaptive radiation suggests that adaptive radiation results from divergent natural selection stemming from environment and resource competition [1,79]. It often happens after the dispersal of the species out its ancestral range, i.e., the colonization of a new environment which suggests ecological opportunities. The textbook examples fall to islands and archipelagoes [80,81,82,83], but in relation to fishes this would also be lakes which are fully or half-isolated, as well as other geographic isolates (e.g., riverine segments isolated by waterfalls).
We assume that the ecological opportunities may not be ‘visible’ to specialists even in a new range/environment if it they are not preceded by relaxed selection that permits the increasing/broadening of ecological plasticity and morphological variability, allowing the specialists to relax their diet and jump out of a narrow/canalized way of specialization. The rivers of the Ethiopian Highlands are rich in waterfalls, which are serious barriers for fish dispersal. Some segments of the rivers are characterized by depauperated fauna that suggest lower competition (relaxed selection). When this coincides with other ecological opportunities (e.g., available non-occupied niches, lowered predator’s pressure), it creates a great combination of the conditions necessary for adaptive radiation to unfold. Therefore, the occupying of such species-poor riverine segments by specialists may (under relaxed selection) facilitate phenotypic and ecological variability followed by the respecialization.
The Sore River is poor in fish fauna (five indigenous species apart from Garra and one introduced species, Coptodon zillii (Gervais 1848)—[34]), and is characterized by the presence of waterfalls which are unpassable for fish. Isolation by waterfalls is beneficial for fortuitous upstream colonists to occupy new niches, as well as for the prevention of gene flow from downstream populations that may collapse incipient adaptive radiation. In addition, the depauperated fauna of the Sore River do not include predatory fishes or crocodiles (our data), which means lowered predator pressure and favors relaxed selection. This being coupled with rather complex habitats represented by ponds, swift areas, and rapids (ecological opportunities) in this segment of the Sore River suggests: (i) escape from the pressure of stabilizing/canalized selection (relaxed constrains of selection), and (ii) entering into new niches with subsequent trophic diversification (respecialization). Respecialization may also happen in other way due to the gain of phenotypic novelties, as a thick-lipped mouth resulted from a hybridization of ecomorphs of Garra in the Sore River [34]. It is noteworthy that Sore’s ecological opportunities have been confirmed by another diversification of cyprinids detected in the same riverine segment: Labeobarbus’ monophyletic radiation comprising four sympatric ecomorphs with divergent mouth phenotypes (see [33]).
Another example of possible respecialization among Garra in Ethiopia falls to lacustrine species G. tana Getahun and Stiassny, 2007, and G. regressus Getahun and Stiassny, 2007, from Lake Tana. Although the data on their trophic ecology are rather scarce, one may suggest a lacustrine respecialization of these species based on frequent occurrence in a pelagic zone, weakly developed gular disc, zooplankton in the diet, and short guts [84]. Another Garra species, G. quadrimaculata (Rüppell 1835), inhabiting Lake Awassa in the Ethiopian Rift Valley, demonstrates a rather relaxed diet, being omnivorous in fact [85]. The gut of another African Garra species from the Cameroun highlands of Niger basin-G. allostoma Roberts, 1990, contained terrestrial (including winged forms) and aquatic insects that also may indicate respecialization [86].
Apart from Africa, feeding respecialization among Garra has apparently occurred in cave-dwelling species or populations in West Asia. Timmermann et al. [87] assumed that a cave-dwelling population of G. barreimiae (Oman) feed on bat guano, organic matter washed into the cave, and small invertebrates compared to algae feeding in the surface-dwelling population. However, thorough studies on the ecology of the cave populations confirming this are lacking.
Remarkably, Garra is not alone among highly specialized algae scrapers which experienced respecialization. The ecological diversification of Schizopygopsis stolickai Steindachner, 1866, inhabiting mountain Lake Yashilkul (3734 m above sea level) in Pamir resulted in four sympatric ecomorphs consuming different foods [26,88]. The ancestral mode of feeding is algae and macrophyte eating (riverine populations), while piscivorous, detritivorous, benthivorous, and phytophagous ecomorphs were detected based on diet, gut length and stable isotope signatures [26]. Notably, this extremely young diversification happened in a lake formed after an earthquake [89] ca. 800 years ago. One more example of the relaxation of specialization among cyprinid scrapers belongs to Capoeta banarescui Turan, Kottelat, Ekmekçi and Imamoğlu, 2006, inhabiting West Asia, Anatolia and Transcaucasia. This species is nested within a widely distributed monophyletic lineage of algae eaters, genus Capoeta, which is not a basal lineage [61]. In spite of the presence of a horny scraper on its lower jaw, C. banarescui displays great feeding plasticity accompanied by gut length variation [90]. Another scraper, Chondrostoma nasus (Linnaeus 1758), widely distributed in Europe, presents a highly variable diet being greatly relaxed [91]. Hence, the relaxation of diet among specialists is a not-so-rare event that sometimes results in respecialization. Based on the above-mentioned examples of Garra and Schizopygopsis, a trophic respecialization can happen very rapidly.
5. Conclusions
The adaptive radiation of Garra in the Sore River, White Nile basin (East Africa) was accompanied by trophic resource partitioning. Six sympatric ecomorphs partition resources in different manners. First, a further sub-diversification within the ancestral periphyton feeding strategy (driven assumedly by different habitat exploitations) was detected. Second, a novel feeding strategy—zoophagy—with subsequent diversification was gained due to respecialization. Respecialization was possible in faunistically poor segments of the river (lowered competition) with a low pressure of predators. All together, this could lead to relaxed selection constrains that facilitate the phenotypic and ecological (dietary) plasticity of specialists. Relaxed selection here was combined with rather complex habitats presuming vacant niches (ecological opportunities). Thus, we suggest a next hypothetical sequence of process of the trophic respecialization of fishes: (a) relaxed selection; (b) increased phenotypic and/or ecological plasticity due to relaxed selection (relaxed feeding); (c) entering into vacant niches; (d) the actuation of divergent natural selection; and (e) respecialization. Respecialization may go another way at initial stages: via hybridization, as shown for the ‘thick-lipped’ ecomorph [34], for example. The case of the riverine diversification of Garra in the Sore River is seemingly unique among all of the known riverine adaptive radiations, being rapid, rich in ecomorph/species numbers, and characterized by a bright pattern of trophic novelties.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/d14080629/s1. Figure S1: Pearson correlation of body length (SL, mm) and gut length (as % SL) in six ecomorphs of the genus Garra from the Sore River. Figure S2: Biplots of stable isotope compositions (mean δ13C and δ15N values ± SD) in the sympatric ecomorphs of the genus Garra from the Sore River (subset includes all of the individuals, n = 83). Figure S3: Pearson correlation of δ15N values and the gut length in six Garra ecomorphs from the Sore River. Figure S4: Pearson correlation of δ13C values and the gut length in six Garra ecomorphs from the Sore River. Figure S5: Stable isotope Bayesian ellipses showing the trophic niche widths and overlaps in the sympatric ecomorphs of the genus Garra from the Sore River (subset includes all of the individuals, n = 83). Ellipses with 95% credible intervals for the means are based on standard ellipses corrected for small sample sizes (SEAc; isotopic niche metrics; SIBER package). Each mark corresponds to the isotopic values. A broken line connects the extreme points in each sample and delineates the total area (TA). The ecomorphs (1–6) are colored by the same colors as in Figure S2. Table S1: Estimated isotopic niche areas in the sympatric Garra ecomorphs from the Sore River (the subset includes all of the individuals, n = 83); TA, SEA, and SEAc (SIBER package) are the total area of the convex hull, the standard ellipse area, and the corrected standard ellipse with a correction for small sample sizes, respectively. Table S2: Niche overlap estimates (NicheROVER package) showing the posterior probabilities (α = 0.95) that individuals from rows will be found within the niches indicated by the column header. The results (%) are provided for sympatric Garra ecomorphs from the Sore River (the subset includes all of the individuals, n = 83). Figure S6: Pearson correlation of the relative gut length (as % to SL) and δ15N values in six ecomorphs of the genus Garra from the Sore River. Figure S7: Pearson correlation of the relative gut length (as % to SL) and δ13C values in six ecomorphs of the genus Garra from the Sore River. Figure S8: Pearson correlation of δ13C and δ15N values in six ecomorphs of the genus Garra from the Sore River.
Author Contributions
Conceptualization, A.S.K., A.S.G. and B.A.L.; methodology, A.S.K., B.A.L.; software, A.S.K.; validation, A.S.K., B.A.L.; formal analysis, A.S.K., B.A.L.; investigation, A.S.K., A.S.G. and B.A.L.; resources, A.S.G., B.A.L.; data curation, A.S.K., B.A.L.; writing—original draft preparation, A.S.K., B.A.L.; writing—review and editing, A.S.K., A.S.G. and B.A.L.; visualization, A.S.K., B.A.L.; supervision, B.A.L.; project administration, A.S.G., B.A.L.; funding acquisition, B.A.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant number 19-14-00218.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, but ethical review and approval were not obtained for this study by the Institutional Ethics Committee of IBIW RAS for the reason that the sampling was performed before the Ethical Committee was established.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Material for this study was collected within the scope of the Joint Ethiopian-Russian Biological Expedition (JERBE). We gratefully acknowledge the JERBE coordinators A.A. Darkov (Severtsov Institute of Ecology and Evolution, Moscow, Russia—IEE) and Simenew Keskes Melaku (Ministry of Innovation and Technology, Addis Ababa, Ethiopia) for administrative assistance. We express our gratitude to S.E. Cherenkov and (IEE), Fekadu Tefera and Genanaw Tesfaye (both from the National Fishery and Other Aquatic Life Research Center of the Ethiopian Institute of Agricultural Research—EIAR, Sebeta) for sharing in the field operations and assistance in collecting the material. We thank S.E. Cherenkov for his help with the photography. We acknowledge A.V. Tiunov and O.L. Rozanova for their valuable help during the stable isotope analysis and discussion of the results.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Meyer, A. Phylogenetic relationships and evolutionary processes in East African cichlid fishes. Trends Ecol. Evol. 1993, 8, 279–284. [Google Scholar] [CrossRef]
- Nagelkerke, L.A.J.; Sibbing, F.A.; van den Boogaart, J.G.M.; Lammens, E.H.R.R.; Osse, J.W.M. The barbs (Barbus spp.) of Lake Tana: A forgotten species flock? Environ. Biol. Fishes 1994, 39, 1–22. [Google Scholar] [CrossRef]
- Mina, M.V.; Mironovsky, A.N.; Dgebuadze, Y.U. Lake Tana large barbs: Phenetics, growth and diversification. J. Fish Biol. 1996, 48, 383–404. [Google Scholar] [CrossRef]
- Seehausen, O.; Wagner, C.E. Speciation in freshwater fishes. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 621–651. [Google Scholar] [CrossRef]
- Alekseyev, S.S.; Bajno, R.; Gordeeva, N.V.; Reist, J.D.; Power, M.; Kirillov, A.F.; Samusenok, V.P.; Matveev, A.N. Phylogeography and sympatric differentiation of the Arctic charr Salvelinus alpinus (L.) complex in Siberia as revealed by mtDNA sequence analysis. J. Fish Biol. 2009, 75, 368–392. [Google Scholar] [CrossRef]
- Østbye, K.; Amundsen, P.-A.; Bernatchez, L.; Klemetsen, A.; Knudsen, R.; Kristoffersen, R.; Naesje, T.F.; Hindar, K. Parallel evolution of ecomorphological traits in the European whitefish Coregonus lavaretus (L.) species complex during postglacial times. Mol. Ecol. 2006, 15, 3983–4001. [Google Scholar] [CrossRef]
- Præbel, K.; Knudsen, R.; Siwertsson, A.; Karhunen, M.; Kahilainen, K.K.; Ovaskainen, O.; Østbye, K.; Peruzzi, S.; Fevolden, S.; Amundsen, P. Ecological speciation in postglacial European whitefish: Rapid adaptive radiations into the littoral, pelagic, and profundal lake habitats. Ecol. Evol. 2013, 3, 4970–4986. [Google Scholar] [CrossRef]
- Jacobs, A.; Carruthers, M.; Yurchenko, A.; Gordeeva, N.V.; Alekseyev, S.S.; Hooker, O.; Leong, J.S.; Minkley, D.R.; Rondeau, E.B.; Koop, B.F.; et al. Parallelism in eco-morphology and gene expression despite variable evolutionary and genomic backgrounds in a Holarctic fish. PLoS Genet. 2020, 16, e1008658. [Google Scholar] [CrossRef]
- Levin, B.; Simonov, E.; Gabrielyan, B.K.; Mayden, R.L.; Rastorguev, S.M.; Roubenyan, H.R.; Sharko, F.S.; Nedoluzhko, A.V. Caucasian treasure: Genomics sheds light on the evolution of half-extinct Sevan trout, Salmo ischchan, species flock. Mol. Phylogenetics Evol. 2022, 167, 107346. [Google Scholar] [CrossRef]
- Masonick, P.; Meyer, A.; Hulsey, C.D. Phylogenomic Analyses Show Repeated Evolution of Hypertrophied Lips Among Lake Malawi Cichlid Fishes. Genome Biol. Evol. 2022, 14, evac051. [Google Scholar] [CrossRef]
- Barluenga, M.; Stölting, K.N.; Salzburger, W.; Muschick, M.; Meyer, A. Sympatric speciation in Nicaraguan crater lake cichlid fish. Nature 2006, 439, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Elmer, K.R.; Fan, S.; Kusche, H.; Luise Spreitzer, M.; Kautt, A.F.; Franchini, P.; Meyer, A. Parallel evolution of Nicaraguan crater lake cichlid fishes via non-parallel routes. Nat. Commun. 2014, 5, 5168. [Google Scholar] [CrossRef] [PubMed]
- Burress, E.D.; Piálek, L.; Casciotta, J.R.; Almirón, A.; Tan, M.; Armbruster, J.W.; Říčan, O. Island-and lake-like parallel adaptive radiations replicated in rivers. Proc. R. Soc. B Biol. Sci. 2018, 285, 20171762. [Google Scholar] [CrossRef] [PubMed]
- Muschick, M.; Nosil, P.; Roesti, M.; Dittmann, M.T.; Harmon, L.; Salzburger, W. Testing the stages model in the adaptive radiation of cichlid fishes in East African Lake Tanganyika. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140605. [Google Scholar] [CrossRef] [PubMed]
- Rüber, L.; Verheyen, E.; Meyer, A. Replicated evolution of trophic specializations in an endemic cichlid fish lineage from Lake Tanganyika. Proc. Nat. Acad. Sci. USA 1999, 96, 10230–10235. [Google Scholar] [CrossRef]
- Alekseyev, S.S.; Samusenok, V.P.; Matveev, A.N.; Pichugin, M.Y. Diversification, Sympatric Speciation, and Trophic Polymorphism of Arctic Charr, Salvelinus Alpinus Complex, in Transbaikalia. Env. Biol. Fish 2002, 64, 97–114. [Google Scholar] [CrossRef]
- Terekhanova, N.V.; Logacheva, M.D.; Penin, A.A.; Neretina, T.V.; Barmintseva, A.E.; Bazykin, G.A.; Kondrashov, A.S.; Mugue, N.S. Fast evolution from precast bricks: Genomics of young freshwater populations of threespine stickleback Gasterosteus aculeatus. PLoS Genet. 2014, 10, e1004696. [Google Scholar] [CrossRef]
- Brodersen, J.; Post, D.M.; Seehausen, O. Upward adaptive radiation cascades: Predator diversification induced by prey diversification. Trends Ecol. Evol. 2018, 33, 59–70. [Google Scholar] [CrossRef]
- Esin, E.V.; Bocharova, E.S.; Borisova, E.A.; Markevich, G.N. Interaction among morphological, trophic and genetic groups in the rapidly radiating Salvelinus fishes from Lake Kronotskoe. Evol. Ecol. 2020, 34, 611–632. [Google Scholar] [CrossRef]
- Skúlason, S. Sympatric morphs, populations and speciation in freshwater fish with emphasis on arctic charr. In Evolution of Biological Diversity; Magurran, A., May, R.M., Eds.; Oxford University Press: New York, NY, USA, 1999; pp. 71–92. [Google Scholar]
- Mina, M.V.; Mironovsky, A.N.; Golubtsov, A.S.; Dgebuadze, Y.Y., II. Morphological diversity of “large barbs”; from Lake Tana and neighbouring areas: Homoplasies or synapomorphies? Ital. J. Zool. 1998, 65, 9–14. [Google Scholar] [CrossRef]
- Dgebuadze, Y.Y.; Mina, M.V.; Alekseyev, S.S.; Golubtsov, A.S. Observations on reproduction of the Lake Tana barbs. J. Fish Biol. 1999, 54, 417–423. [Google Scholar] [CrossRef]
- Dgebuadze, Y.Y.; Mironovsky, A.N.; Mendsaikhan, B.; Slyn’ko, Y.V. Rapid Morphological Diversification of the Cyprinid Fish Oreoleuciscus potanini (Cyprinidae) in the Course of Formation of a Reservoir in a River of the Semiarid Zone. Dokl. Biol. Sci. 2020, 490, 12–15. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, M.; Dejen, E.; Osse, J.W.M.; Sibbing, F.A. Adaptive radiation of Lake Tana’s (Ethiopia) Labeobarbus species flock (Pisces, Cyprinidae). Mar. Freshw. Res. 2008, 59, 391–407. [Google Scholar] [CrossRef]
- Komarova, A.S.; Rozanova, O.L.; Levin, B.A. Trophic resource partitioning by sympatric ecomorphs of Schizopygopsis (Cyprinidae) in a young Pamir Mountain lake: Preliminary results. Ichthyol. Res. 2021, 68, 191–197. [Google Scholar] [CrossRef]
- Levin, B.A.; Casal-López, M.; Simonov, E.; Dgebuadze, Y.Y.; Mugue, N.S.; Tiunov, A.V.; Doadrio, I.; Golubtsov, A.S. Adaptive radiation of barbs of the genus Labeobarbus (Cyprinidae) in an East African river. Freshw. Biol. 2019, 64, 1721–1736. [Google Scholar] [CrossRef]
- Levin, B.A.; Komarova, A.S.; Rozanova, O.L.; Golubtsov, A.S. Unexpected Diversity of Feeding Modes among Chisel-Mouthed Ethiopian Labeobarbus (Cyprinidae). Water 2021, 13, 2345. [Google Scholar] [CrossRef]
- Sibbing, F.A.; Nagelkerke, L.A.; Stet, R.J.; Osse, J.W. Speciation of endemic Lake Tana barbs (Cyprinidae, Ethiopia) driven by trophic resource partitioning; a molecular and ecomorphological approach. Aquat. Ecol. 1998, 32, 217–227. [Google Scholar] [CrossRef]
- Shkil, F.N.; Levin, B.A.; Abdissa, B.; Smirnov, S.V. Variability in the number of tooth rows in the pharyngeal dentition of Barbus intermedius (Teleostei; Cyprinidae): Genetic, hormonal and environmental factors. J. Appl. Ichthyol. 2010, 26, 315–319. [Google Scholar] [CrossRef]
- Shkil, F.N.; Lazebnyi, O.E.; Kapitanova, D.V.; Abdissa, B.; Borisov, V.B.; Smirnov, S.V. Ontogenetic mechanisms of explosive morphological divergence in the Lake Tana (Ethiopia) species flock of large African barbs (Labeobarbus; Cyprinidae; Teleostei). Russ. J. Dev. Biol. 2015, 46, 294–306. [Google Scholar] [CrossRef]
- Dimmick, W.W.; Berendzen, P.B.; Golubtsov, A.S. Genetic comparison of three Barbus (Cyprinidae) morphotypes from the Genale River, Ethiopia. Copeia 2001, 4, 1123–1129. [Google Scholar] [CrossRef]
- Levin, B.A.; Simonov, E.; Dgebuadze, Y.Y.; Levina, M.; Golubtsov, A.S. In the rivers: Multiple adaptive radiations of cyprinid fishes (Labeobarbus) in Ethiopian Highlands. Sci. Rep. 2020, 10, 7192. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.; Simonov, E.; Franchini, P.; Mugue, N.; Golubtsov, A.; Meyer, A. Rapid adaptive radiation in a hillstream cyprinid fish in the East African White Nile River basin. Mol. Ecol. 2021, 30, 5530–5550. [Google Scholar] [CrossRef] [PubMed]
- Melnik, N.O.; Esin, E.V. Skull morphology variation as related to trophic specialization in three forms of Salvelinus malma (Salmonidae) from the Kamchatka River basin. Dokl. Biol. Sci. 2020, 492, 75–78. [Google Scholar] [CrossRef]
- Piálek, L.; Říčan, O.; Casciotta, J.; Almirón, A.; Zrzavý, J. Multilocus phylogeny of Crenicichla (Teleostei: Cichlidae), with biogeography of the C. lacustris group: Species flocks as a model for sympatric speciation in rivers. Mol. Phylogenetics Evol. 2012, 62, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.R.; Khaironizam, M.Z. Trophic polymorphism in the Malaysian fish Neolissochilus soroides and other Old world barbs (Teleostei, Cyprinidae). Nat. Hist. Bull. Siam Soc. 2008, 56, 25–53. [Google Scholar]
- Schwarzer, J.; Misof, B.; Ifuta, S.N.; Schliewen, U.K. Time and origin of cichlid colonization of the lower Congo rapids. PLoS ONE 2011, 6, e22380. [Google Scholar] [CrossRef]
- Golubtsov, A.S.; Cherenkov, S.E.; Tefera, F.T. High morphological diversity of the genus Garra in the Sore River (the White Nile Basin, Ethiopia): One more cyprinid species flock? J. Ichthyol. 2012, 52, 817–820. [Google Scholar] [CrossRef]
- Kottelat, M. Ceratogarra, a genus name for Garra cambodgiensis and G. fasciacauda and comments on the oral and gular soft anatomy in labeonine fishes (Teleostei: Cyprinidae). Raffles Bull. Zool. Suppl. 2020, 35, 156–178. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References. 2022. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 15 February 2022).
- Menon, A.G.K. Monograph of the cyprinid fishes of the genus Garra, Hamilton. Mem. Indian. Mus. 1964, 14, 173–260. [Google Scholar]
- Zhang, E. Garra bispinosa, a new species of cyprinid fish (Teleostei: Cypriniformes) from Yunnan, Southwest China. Raffles Bull. Zool. 2005, 13, 9–15. [Google Scholar]
- Hamidan, N.; Jackson, M.C.; Britton, J.R. Diet and trophic niche of the endangered fish Garra ghorensis in three Jordanian populations. Ecol. Freshw. Fish 2016, 25, 455–464. [Google Scholar] [CrossRef]
- Dibaba, A.; Soromessa, T.; Workineh, B. Carbon stock of the various carbon pools in Gerba-Dima moist Afromontane forest, South-western Ethiopia. Carbon Balance Manag. 2019, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Kebede, A.; Diekkrüger, B.; Moges, S.A. Comparative study of a physically based distributed hydrological model versus a conceptual hydrological model for assessment of climate change response in the Upper Nile, Baro-Akobo basin: A case study of the Sore watershed, Ethiopia. Int. J. River Basin Manag. 2014, 12, 299–318. [Google Scholar] [CrossRef]
- Topic Popovic, N.; Strunjak-Perovic, I.; Coz-Rakovac, R.; Barisic, J.; Jadan, M.; Persin Berakovic, A.; Sauerborn Klobucar, R. Tricaine methane-sulfonate (MS-222) application in fish anaesthesia. J. Appl. Ichthyol. 2012, 28, 553–564. [Google Scholar] [CrossRef]
- Google Maps. 2022. Available online: https://www.google.com/maps (accessed on 30 June 2022).
- Wagner, C.E.; McIntyre, P.B.; Buels, K.S.; Gilbert, D.M.; Michel, E. Diet predicts intestine length in Lake Tanganyika’s cichlid fishes. Funct. Ecol. 2009, 23, 1122–1131. [Google Scholar] [CrossRef]
- Ewart-Smith, J.L. The Relationship between Periphyton, Flow and Nutrients in Foothill Rivers of the South-Western Cape, South Africa. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 2012. [Google Scholar]
- Natarajan, A.V.; Jhingran, A.G. Index of preponderance—A method of grading the food elements in the stomach analysis of fishes. Indian J. Fish 1961, 8, 54–59. [Google Scholar]
- Popova, O.A.; Reshetnikov, Y.S. On Complex Indices in Investigation of Fish Feeding. J. Ichthyol. 2011, 51, 686–691. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montana, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2021; Available online: http://www.rstudio.com (accessed on 12 February 2021).
- Comtois, D. Summarytools: Tools to Quickly and Neatly Summarize Data. R Package Version 0.8. 72018. 2018. Available online: https://CRAN.R-project.org/package=summarytools (accessed on 12 February 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Pohlert, T. Package ‘PMCMRplus’. R Package Version 1.9.2.2021. Available online: https://cran.r-project.org/web/packages/PMCMRplus/index.html (accessed on 15 April 2022).
- Jackson, A.L.; Parnell, A.C.; Inger, R.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER0–Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Swanson, H.K.; Lysy, M.; Power, M.; Stasko, A.D.; Johnson, J.D.; Reist, J.D. A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap. Ecology 2015, 96, 318–324. [Google Scholar] [CrossRef]
- Matthes, H.A. Comparative study of the feeding mechanisms of some African Cyprinidae (Pisces, Cypriniformes). Bijdr. Dierk. 1963, 33, 3–35. [Google Scholar] [CrossRef]
- Levin, B.A.; Freyhof, J.; Lajbner, Z.; Perea, S.; Abdoli, A.; Gaffaroğlu, M.; Özuluğ, M.; Rubenyan, H.R.; Salnikov, V.B.; Doadrio, I. Phylogenetic relationships of the algae scraping cyprinid genus Capoeta (Teleostei: Cyprinidae). Mol. Phylogenetics Evol. 2012, 62, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Hata, H.; Shibata, J.; Omori, K.; Kohda, M.; Hori, M. Depth segregation and diet disparity revealed by stable isotope analyses in sympatric herbivorous cichlids in Lake Tanganyika. Zool. Lett. 2015, 1, 1–11. [Google Scholar] [CrossRef]
- Azim, M.E.; Beveridge, M.C.M.; van Dam, A.A.; Verdegem, M.C. Periphyton and aquatic production: An introduction. In Periphyton: Ecology, Exploitation and Management; Azim, M.E., Verdegem, M.C.J., van Dam, A.A., Beveridge, M.C.M., Eds.; Centre for Agriculture and Bioscience International: Wallingford, UK, 2005; pp. 1–13. [Google Scholar]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Saikia, S.K. Review on periphyton as mediator of nutrient transfer in aquatic ecosystems. Ecol. Balk. 2011, 3, 65–78. [Google Scholar]
- Hicks, B.J. Food webs in forest and pasture streams in the Waikato region, New Zealand: A study based on analyses of stable isotopes of carbon and nitrogen, and fish gut contents. N. Z. J. Mar. Freshw. Res. 1997, 31, 651–664. [Google Scholar] [CrossRef]
- Jones, J.I.; Waldron, S. Combined stable isotope and gut contents analysis of food webs in plant-dominated, shallow lakes. Freshw. Biol. 2003, 48, 1396–1407. [Google Scholar] [CrossRef]
- Sturmbauer, C.; Mark, W.; Dallinger, R. Ecophysiology of Aufwuchs-eating cichlids in Lake Tanganyika: Niche separation by trophic specialization. Environ. Biol. Fishes 1992, 35, 283–290. [Google Scholar] [CrossRef]
- Rodelli, M.R.; Gearing, J.N.; Gearing, P.J.; Marshall, N.; Sasekumar, A. Stable isotope ratio as a tracer of mangrove carbon in Malaysian ecosystems. Oecologia 1984, 61, 326–333. [Google Scholar] [CrossRef]
- Planas, M. Ecological Traits and Trophic Plasticity in The Greater Pipefish Syngnathus acus in the NW Iberian Peninsula. Biology 2022, 11, 712. [Google Scholar] [CrossRef]
- Day, E.H.; Hua, X.; Bromham, L. Is specialization an evolutionary dead end? Testing for differences in speciation, extinction and trait transition rates across diverse phylogenies of specialists and generalists. J. Evol. Biol. 2016, 29, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Liem, K.F. Adaptive Significance of Intra- and Interspecific Differences in the Feeding Repertoires of Cichlid Fishes. Am. Zool. 1980, 20, 295–314. [Google Scholar] [CrossRef]
- Robinson, B.W.; Wilson, D.S. Optimal Foraging, Specialization, and a Solution to Liem’s Paradox. Am. Nat. 1998, 151, 223–235. [Google Scholar] [CrossRef]
- Binning, S.A.; Chapman, L.J.; Cosandey-Godin, A. Specialized morphology for a generalist diet: Evidence for Liem’s Paradox in a cichlid fish. J. Fish Biol. 2009, 75, 1683–1699. [Google Scholar] [CrossRef] [PubMed]
- Golcher-Benavides, J.; Wagner, C.E. Playing out Liem’s Paradox: Opportunistic Piscivory across Lake Tanganyikan Cichlids. Am. Nat. 2019, 194, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Kruppert, S.; Summers, A.P. Fishing out a feeding paradox. Nature 2019, 571, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Nikolsky, G.V. The Ecology of Fishes; Academic Press: New York, NY, USA, 1963; 352p. [Google Scholar]
- Coad, B.W. Carps and Minnows of Iran (Families Cyprinidae and Leuciscidae). Vol. I: General Introduction and Carps (Family Cyprinidae). Available online: http://briancoad.com/Species%20Accounts/Carps%20of%20Iran%2010Sept2opt1.pdf (accessed on 20 December 2021).
- Stroud, J.T.; Losos, J.B. Ecological opportunity and adaptive radiation. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 507–532. [Google Scholar] [CrossRef]
- Losos, J.B. Integrative approaches to evolutionary ecology: Anolis lizards as model systems. Annu. Rev. Ecol. Syst. 1994, 25, 467–493. [Google Scholar] [CrossRef]
- Grant, P.R.; Grant, B.R. Unpredictable evolution in a 30-year study of Darwin’s finches. Science 2002, 296, 707–711. [Google Scholar] [CrossRef]
- Pinto, G.; Mahler, D.; Harmon, L.; Losos, J. Testing the island effect in adaptive radiation: Rates and patterns of morphological diversification in Caribbean and mainland Anolis lizards. Proc. R. Soc. B Biol. Sci. 2008, 275, 2749–2757. [Google Scholar] [CrossRef]
- Richards, E.J.; Martin, C.H. Adaptive introgression from distant Caribbean islands contributed to the diversification of a microendemic adaptive radiation of trophic specialist pupfishes. PLoS Genet. 2017, 13, e1006919. [Google Scholar] [CrossRef] [PubMed]
- Geremew, A. Taxonomic Revision, Relative Abundance, and Aspects of the Biology of Some Species of the Genus Garra, Hamilton 1922 (Pisces: Cyprinidae) in Lake Tana, Ethiopia. Doctoral Dissertation, Addis Ababa University, Addis Ababa, Ethiopia, 2007. [Google Scholar]
- Tekle-Giorgis, Y.; Yilma, H.; Dadebo, E. Feeding habits and trace metal concentrations in the muscle of lapping minnow Garra quadrimaculata (Rüppell, 1835) (Pisces: Cyprinidae) in Lake Hawassa, Ethiopia. Momona Ethiop. J. Sci. 2016, 8, 116–135. [Google Scholar] [CrossRef]
- Roberts, T.S. Garra allostoma, a new species of Cyprinid fish from highlands of the Niger basin in Cameroun. Rev. Hydrobiol. Trop. 1990, 23, 161–169. [Google Scholar]
- Timmermann, M.; Schlupp, I.; Plath, M. Shoaling behaviour in a surface-dwelling and a cave-dwelling population of a barb Garra barreimiae (Cyprinidae, Teleostei). Acta Ethologica 2004, 7, 59–64. [Google Scholar] [CrossRef]
- Savvaitova, K.A.; Shanin, A.Y.; Verigina, I.A. Speciation and species structure of false osman Schizopygopsis stoliczkai in water bodies of Pamir. Vopr. Ikhtiol. 1988, 28, 896–906. [Google Scholar]
- Grishchenko, E.V. Biology, Fishery Importance of False Osman (Schizopygopsis stoliczkai Steind. 1888) and Ways of Increase of Fish Capacity in Pamir Water Bodies. Ph.D. Thesis, Moscow State University, Moscow, Soviet Union, 1984. [Google Scholar]
- Akin, S.; Turan, H.; Kaymak, N. Does diet variation determine the digestive tract length of Capoeta banarescui Turan, Kottelat, Ekmekci and Imamoglu, 2006? J. Appl. Ichthyol. 2016, 32, 883–892. [Google Scholar] [CrossRef]
- Lammens, E.H.R.R.; Hoogenboezem, W. Diets and feeding behaviour. In Cyprinid Fishes; Winfield, I.J., Nelson, J.S., Eds.; Chapman & Hall: London, UK, 1991; pp. 353–376. [Google Scholar]
Figure 1.
Garra ecomorphs from the Sore River: 1, ‘generalized’: 136 mm SL; 2, ‘stream-lined’: 99 mm SL; 3, ‘narrow-mouth’: 100 mm SL; 4, ‘wide-mouth’: 100 mm SL; 5, ‘predator’: 193 mm SL; 6, ‘thick-lipped’: 128 mm SL.
Figure 1.
Garra ecomorphs from the Sore River: 1, ‘generalized’: 136 mm SL; 2, ‘stream-lined’: 99 mm SL; 3, ‘narrow-mouth’: 100 mm SL; 4, ‘wide-mouth’: 100 mm SL; 5, ‘predator’: 193 mm SL; 6, ‘thick-lipped’: 128 mm SL.

Figure 2.
Map with sampling sites of Garra in the Sore River, White Nile basin, East Africa (left); satellite images of the riverine segments with certain sampling points (right). The map was created using QGIS v.3.16.4-Hannover.
Figure 2.
Map with sampling sites of Garra in the Sore River, White Nile basin, East Africa (left); satellite images of the riverine segments with certain sampling points (right). The map was created using QGIS v.3.16.4-Hannover.

Figure 3.
Violin plots of the relative gut length of six sympatric ecomorphs, Garra, from the Sore River. Min–max values (whiskers), first and third quartiles (white vertical bars), median values (black horizontal bars), and outliers (black points) are indicated. The lowercase letters above the violin plots indicate significant differences between ecomorphs (p < 0.05, Kruskal–Wallis test with Dunn’s post hoc test).
Figure 3.
Violin plots of the relative gut length of six sympatric ecomorphs, Garra, from the Sore River. Min–max values (whiskers), first and third quartiles (white vertical bars), median values (black horizontal bars), and outliers (black points) are indicated. The lowercase letters above the violin plots indicate significant differences between ecomorphs (p < 0.05, Kruskal–Wallis test with Dunn’s post hoc test).

Figure 4.
Food spectra (IR: the index of relative importance) of the six Garra ecomorphs from the Sore River.
Figure 4.
Food spectra (IR: the index of relative importance) of the six Garra ecomorphs from the Sore River.

Figure 5.
Biplots of stable isotope composition (means of δ15N and δ13C values ± SD) of the sympatric ecomorphs of the genus Garra (length of fish individuals: 50–90 mm, n = 44). The ellipses delineate zoophagous and periphython-feeding ecomorphs.
Figure 5.
Biplots of stable isotope composition (means of δ15N and δ13C values ± SD) of the sympatric ecomorphs of the genus Garra (length of fish individuals: 50–90 mm, n = 44). The ellipses delineate zoophagous and periphython-feeding ecomorphs.

Figure 6.
(a) Pearson correlation of the δ15N values with body length in six Garra ecomorphs from the Sore River. (b) Pearson correlation of the δ13C values with body length in six Garra ecomorphs from the Sore River.
Figure 6.
(a) Pearson correlation of the δ15N values with body length in six Garra ecomorphs from the Sore River. (b) Pearson correlation of the δ13C values with body length in six Garra ecomorphs from the Sore River.


Figure 7.
Stable isotope Bayesian ellipses showing the trophic niche widths and overlaps in sympatric ecomorphs of the genus Garra from the Sore River (subset of individuals of 50–90 mm SL, n = 44). Ellipses with 95% credible intervals for the means are based on standard ellipses corrected for small sample sizes (SEAc; isotopic niche metrics; SIBER package). Each mark corresponds to the isotopic values. A broken line connects the extreme points in each sample and delineates the total area (TA). The ecomorphs (1–5) are colored by the same colors as in Figure 5.
Figure 7.
Stable isotope Bayesian ellipses showing the trophic niche widths and overlaps in sympatric ecomorphs of the genus Garra from the Sore River (subset of individuals of 50–90 mm SL, n = 44). Ellipses with 95% credible intervals for the means are based on standard ellipses corrected for small sample sizes (SEAc; isotopic niche metrics; SIBER package). Each mark corresponds to the isotopic values. A broken line connects the extreme points in each sample and delineates the total area (TA). The ecomorphs (1–5) are colored by the same colors as in Figure 5.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sample size of the studied ecomorphs of Garra in the Sore River.
| Ecomorphs | SL Range, mm | Gut Length (n) | Diet (n) | Stable Isotopes * (n) |
|---|---|---|---|---|
| ‘generalized’ (1) | 40–162 | 80 | 80 | 30 |
| ‘stream-lined’ (2) | 71–112 | 23 | 23 | 15 |
| ‘narrow-mouth’ (3) | 47–107 | 22 | 22 | 11 |
| ‘wide-mouth’ (4) | 51–96 | 32 | 29 | 9 |
| ‘predator’ (5) | 39–193 | 26 | 26 | 14 |
| ‘thick-lipped’ (6) | 39–41 | 2 | 2 | 4 |
| Total | 185 | 182 | 83 |
* Sampling was performed in 2014 only.
Table 2.
Estimated isotopic niche areas in the sympatric Garra ecomorphs from the Sore River (subset of 50–90 mm SL, n = 44); TA, SEA, and SEAc (SIBER package) are the total area of the convex hull, the standard ellipse area, and the corrected standard ellipse with a correction for small sample sizes, respectively.
Table 2.
Estimated isotopic niche areas in the sympatric Garra ecomorphs from the Sore River (subset of 50–90 mm SL, n = 44); TA, SEA, and SEAc (SIBER package) are the total area of the convex hull, the standard ellipse area, and the corrected standard ellipse with a correction for small sample sizes, respectively.
| Ecomorphs | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| TA | 12.97 | 1.15 | 7.89 | 0.90 | 7.90 |
| SEA | 3.95 | 1.07 | 5.45 | 0.97 | 6.53 |
| SEAc | 4.17 | 1.34 | 6.54 | 1.29 | 8.16 |
Table 3.
Niche overlap estimates (NicheROVER package) showing the posterior probabilities (α = 0.95) that individuals from rows will be found within the niches indicated by the column header. Results (%) are provided for sympatric Garra ecomorphs from the Sore River (subset of 50–90 mm SL, n = 44).
Table 3.
Niche overlap estimates (NicheROVER package) showing the posterior probabilities (α = 0.95) that individuals from rows will be found within the niches indicated by the column header. Results (%) are provided for sympatric Garra ecomorphs from the Sore River (subset of 50–90 mm SL, n = 44).
| Ecomorphs | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| 1 | NA | 29.03 | 37.85 | 30.55 | 29.33 |
| 2 | 87.51 | NA | 48.93 | 36.50 | 40.87 |
| 3 | 35.66 | 11.54 | NA | 2.36 | 78.96 |
| 4 | 95.74 | 34.23 | 16.20 | NA | 8.48 |
| 5 | 34.06 | 9.94 | 71.31 | 1.22 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Komarova, A.S.; Golubtsov, A.S.; Levin, B.A. Trophic Diversification Out of Ancestral Specialization: An Example from a Radiating African Cyprinid Fish (Genus Garra). Diversity 2022, 14, 629. https://doi.org/10.3390/d14080629
AMA Style
Komarova AS, Golubtsov AS, Levin BA. Trophic Diversification Out of Ancestral Specialization: An Example from a Radiating African Cyprinid Fish (Genus Garra). Diversity. 2022; 14(8):629. https://doi.org/10.3390/d14080629
Chicago/Turabian StyleKomarova, Aleksandra S., Alexander S. Golubtsov, and Boris A. Levin. 2022. "Trophic Diversification Out of Ancestral Specialization: An Example from a Radiating African Cyprinid Fish (Genus Garra)" Diversity 14, no. 8: 629. https://doi.org/10.3390/d14080629
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.