
Patterns of Herbivory in Neotropical Forest Katydids as Revealed by DNA Barcoding of Digestive Tract Contents

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Katydid Capture and Sample Preparation
2.2. DNA Extraction and Amplification
2.3. Identification of Plant Species
2.4. Growth Height Assignment
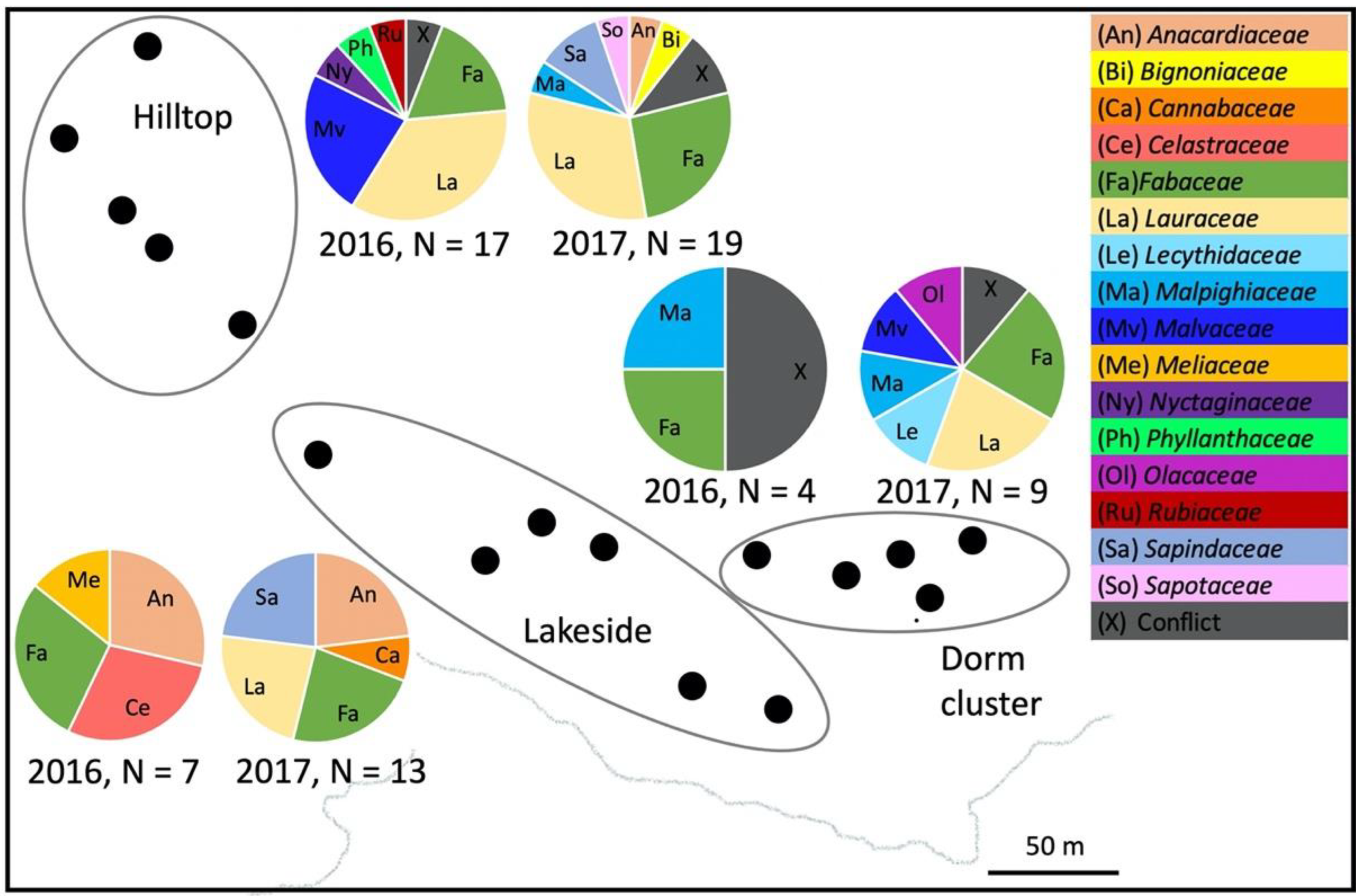
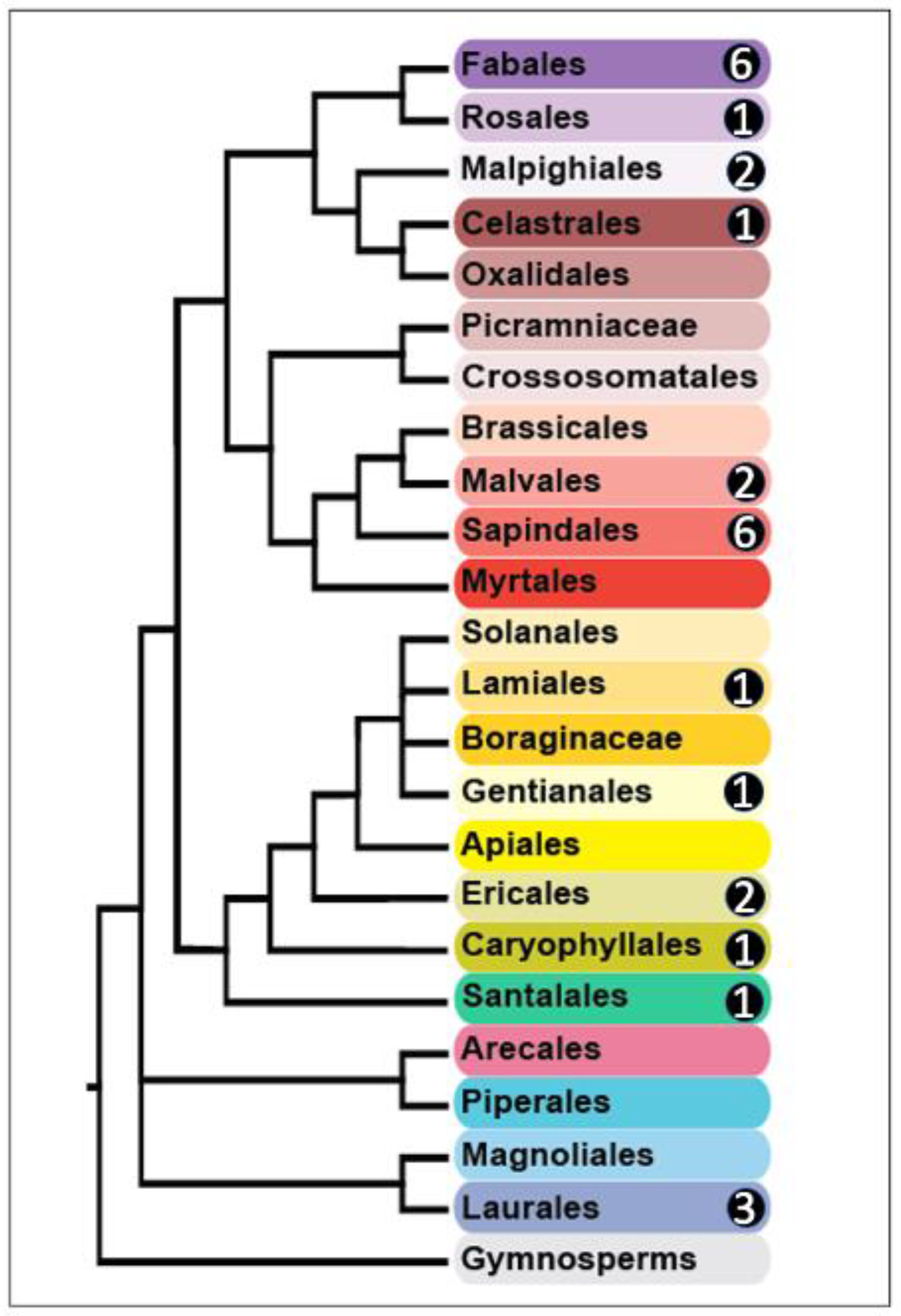
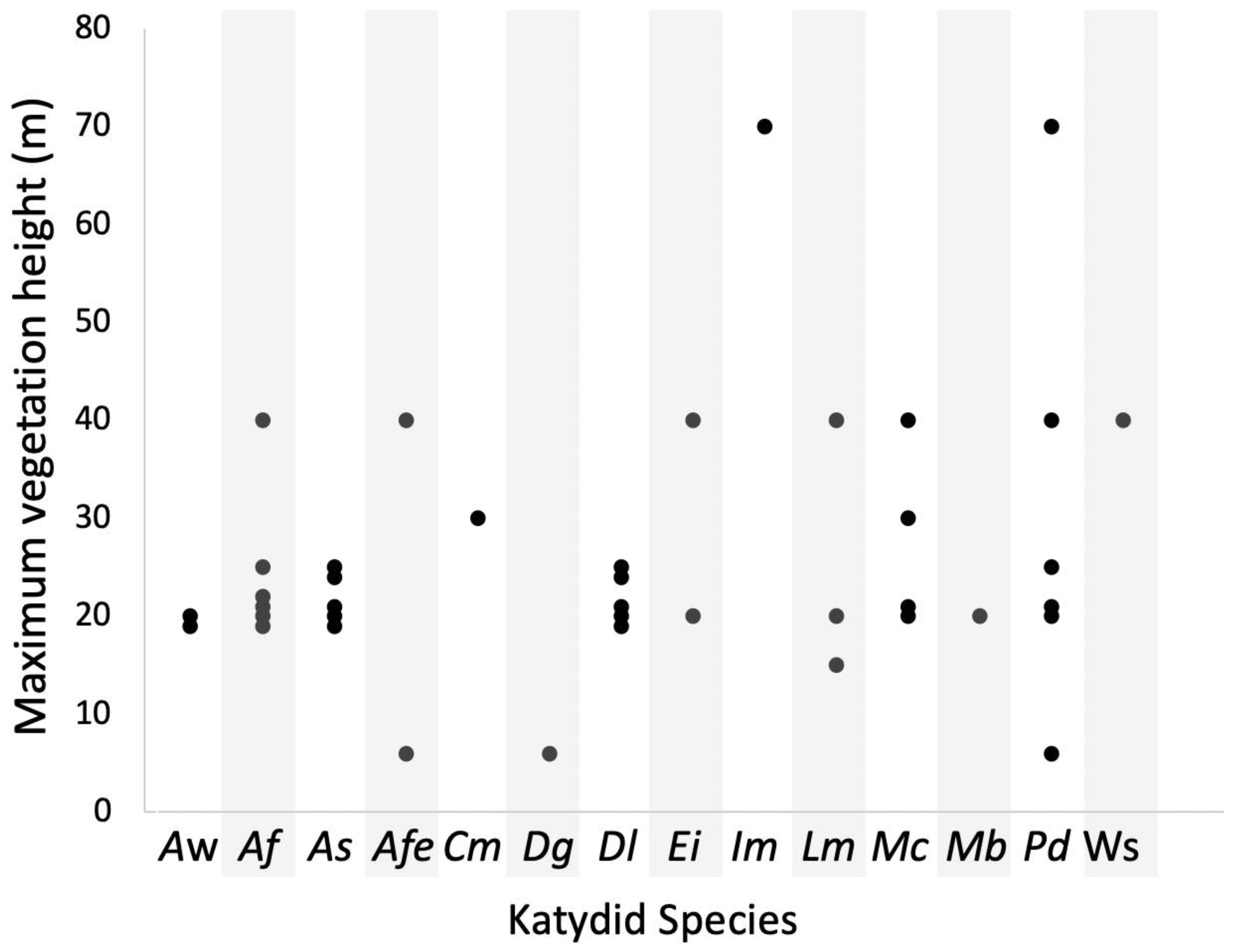
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ryan, M.J.; Tuttle, M.D.; Rand, A.S. Bat Predation and Sexual Advertisement in a Neotropical Anuran. Am. Nat. 1982, 119, 136–139. [Google Scholar] [CrossRef]
- Langerhans, R.B.; Layman, C.A.; DeWitt, T.J. Male Genital Size Reflects a Tradeoff between Attracting Mates and Avoiding Predators in Two Live-Bearing Fish Species. Proc. Natl. Acad. Sci. USA 2005, 102, 7618–7623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halfwerk, W.; Jones, P.L.; Taylor, R.C.; Ryan, M.J.; Page, R.A. Risky Ripples Allow Bats and Frogs to Eavesdrop on a Multisensory Sexual Display. Science 2014, 343, 413–416. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M. Sexual Selection; Princeton University Press: Princeton, NJ, USA, 1994; ISBN 9780691000572. [Google Scholar]
- Zuk, M.; Kolluru, G.R. Exploitation of Sexual Signals by Predators and Parasitoids. Q. Rev. Biol. 1998, 73, 415–438. [Google Scholar] [CrossRef]
- Wing, S.R. Cost of Mating for Female Insects: Risk of Predation in Photinus collustrans (Coleoptera: Lampyridae). Am. Nat. 1988, 131, 139–142. [Google Scholar] [CrossRef]
- Ter Hofstede, H.; Voigt-Heucke, S.; Lang, A.; Römer, H.; Page, R.; Faure, P.; Dechmann, D. Revisiting Adaptations of Neotropical Katydids (Orthoptera: Tettigoniidae) to Gleaning Bat Predation. Neotrop. Biodivers. 2017, 3, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Heller, K.-G. Risk Shift between Males and Females in the Pair-Forming Behavior of Bushcrickets. Naturwissenschaften 1992, 79, 89–91. [Google Scholar] [CrossRef]
- Carl Gerhardt, H.; Huber, F. Acoustic Communication in Insects and Anurans: Common Problems and Diverse Solutions; University of Chicago Press: Chicago, IL, USA, 2002; ISBN 9780226288338. [Google Scholar]
- SINA Singing Insects of North America. Available online: https://sina.orthsoc.org/index.htm (accessed on 8 October 2021).
- Symes, L.B.; Ayres, M.P.; Cowdery, C.P.; Costello, R.A. Signal Diversification in Oecanthus Tree Crickets Is Shaped by Energetic, Morphometric, and Acoustic Trade-Offs. Evolution 2015, 69, 1518–1527. [Google Scholar] [CrossRef]
- Prestwich, K.N.; Walker, T.J. Energetics of Singing in Crickets: Effect of Temperature in Three Trilling Species (Orthoptera: Gryllidae). J. Comp. Physiol. B 1981, 143, 199–212. [Google Scholar] [CrossRef]
- Walker, T.J. Experimental Demonstration of a Cat Locating Orthopteran Prey by the Prey’s Calling Song. Fla. Entomol. 1964, 47, 163–165. [Google Scholar] [CrossRef]
- Cade, W. Acoustically Orienting Parasitoids: Fly Phonotaxis to Cricket Song. Science 1975, 190, 1312–1313. [Google Scholar] [CrossRef]
- Falk, J.J.; Ter Hofstede, H.M.; Jones, P.L.; Dixon, M.M.; Faure, P.A.; Kalko, E.K.V.; Page, R.A. Sensory-Based Niche Partitioning in a Multiple Predator–multiple Prey Community. Proc. R. Soc. B 2015, 282, 20150520. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.L.; Ryan, M.J.; Page, R.A. Population and Seasonal Variation in Response to Prey Calls by an Eavesdropping Bat. Behav. Ecol. Sociobiol. 2014, 68, 605–615. [Google Scholar] [CrossRef]
- Belwood, J.J. The Influence of Bat Predation on Calling Behavior in Neotropical Forest Katydids (Insecta: Orthoptera: Tettigoniidae). Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 1990. [Google Scholar]
- Symes, L.B.; Robillard, T.; Martinson, S.J.; Dong, J.; Kernan, C.E.; Miller, C.R.; ter Hofstede, H.M. Daily Signaling Rate and the Duration of Sound per Signal Are Negatively Related in Neotropical Forest Katydids. Integr. Comp. Biol. 2021, 61, 877–899. [Google Scholar] [CrossRef]
- Ter Hofstede, H.M.; Symes, L.B.; Martinson, S.J.; Robillard, T.; Faure, P.; Madhusudhana, S.; Page, R.A. Calling Songs of Neotropical Katydids (Orthoptera: Tettigoniidae) from Panama. J. Orthoptera Res. 2020, 29, 137. [Google Scholar] [CrossRef]
- Romer, H.; Lewald, J. High-Frequency Sound Transmission in Natural Habitats: Implications for the Evolution of Insect Acoustic Communication. Behav. Ecol. Sociobiol. 1992, 29, 437–444. [Google Scholar] [CrossRef]
- Symes, L.B.; Martinson, S.J.; Kernan, C.E.; ter Hofstede, H.M. Sheep in Wolves’ Clothing: Prey Rely on Proactive Defences When Predator and Non-Predator Cues Are Similar. Proc. Biol. Sci. 2020, 287, 20201212. [Google Scholar] [CrossRef]
- Lang, A.B.; Römer, H. Roost Site Selection and Site Fidelity in the Neotropical Katydid Docidocercus gigliotosi (Tettigoniidae). Biotropica 2008, 40, 183–189. [Google Scholar] [CrossRef]
- Heller, K.-G.; Hemp, C.; Ingrisch, S.; Liu, C. Acoustic Communication in Phaneropterinae (Tettigonioidea)—A Global Review with Some New Data. J. Orthop. Res. 2015, 24, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Villarreal, S.M.; Gilbert, C. Male Scudderia pistillata Katydids Defend Their Acoustic Duet against Eavesdroppers. Behav. Ecol. Sociobiol. 2014, 68, 1669–1675. [Google Scholar] [CrossRef]
- Xu, H.; Turlings, T.C.J. Plant Volatiles as Mate-Finding Cues for Insects. Trends Plant. Sci. 2018, 23, 100–111. [Google Scholar] [CrossRef]
- Cocroft, R.B.; Rodríguez, R.L.; Hunt, R.E. Host Shifts and Signal Divergence: Mating Signals Covary with Host Use in a Complex of Specialized Plant-Feeding Insects. Biol. J. Linn. Soc. 2009, 99, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Kursar, T.A.; Coley, P.D. Convergence in Defense Syndromes of Young Leaves in Tropical Rainforests. Biochem. Syst. Ecol. 2003, 31, 929–949. [Google Scholar] [CrossRef]
- Sedio, B.E.; Rojas Echeverri, J.C.; Boya, P.C.A.; Wright, S.J. Sources of Variation in Foliar Secondary Chemistry in a Tropical Forest Tree Community. Ecology 2017, 98, 616–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coley, P.D. Herbivory and Defensive Characteristics of Tree Species in a Lowland Tropical Forest. Ecol. Monogr. 1983, 53, 209–234. [Google Scholar] [CrossRef]
- Singer, M.S.; Bernays, E.A.; Carrière, Y. The Interplay between Nutrient Balancing and Toxin Dilution in Foraging by a Generalist Insect Herbivore. Anim. Behav. 2002, 64, 629–643. [Google Scholar] [CrossRef]
- Pulliam, H.R. Diet Optimization with Nutrient Constraints. Am. Nat. 1975, 109, 765–768. [Google Scholar] [CrossRef]
- Birnbaum, S.S.L.; Abbot, P. Insect Adaptations toward Plant Toxins in Milkweed–herbivores Systems—A Review. Entomol. Exp. Appl. 2018, 166, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Dobler, S.; Petschenka, G.; Pankoke, H. Coping with Toxic Plant Compounds—The Insect’s Perspective on Iridoid Glycosides and Cardenolides. Phytochemistry 2011, 72, 1593–1604. [Google Scholar] [CrossRef]
- Unsicker, S.B.; Oswald, A.; Köhler, G.; Weisser, W.W. Complementarity Effects through Dietary Mixing Enhance the Performance of a Generalist Insect Herbivore. Oecologia 2008, 156, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.R.; Capinera, J.L. Host Preferences and Habitat Associations of Some Florida Grasshoppers (Orthoptera: Acrididae). Environ. Entomol. 2005, 34, 210–224. [Google Scholar] [CrossRef] [Green Version]
- Sword, G.A.; Joern, A.; Senior, L.B. Host Plant-Associated Genetic Differentiation in the Snakeweed Grasshopper, Hesperotettix viridis (Orthoptera: Acrididae). Mol. Ecol. 2005, 14, 2197–2205. [Google Scholar] [CrossRef] [PubMed]
- Valtonen, A.; Malinga, G.M.; Junes, P.; Opoke, R.; Lehtovaara, V.J.; Nyeko, P.; Roininen, H. The Edible Katydid Ruspolia differens is a Selective Feeder on the Inflorescences and Leaves of Grass Species. Entomol. Exp. Appl. 2018, 166, 592–602. [Google Scholar] [CrossRef]
- Montealegre, Z.F.; Sarria, F.A.; Pimienta, M.C.; Mason, A.C. Lack of Correlation between Vertical Distribution and Carrier Frequency, and Preference for Open Spaces in Arboreal Katydids That Use Extreme Ultrasound, in Gorgona, Colombia (Orthoptera: Tettigoniidae). Rev. Biol. Trop. 2014, 62, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Kress, W.J. Plant DNA Barcodes: Applications Today and in the Future. J. Syst. Evol. 2017, 55, 291–307. [Google Scholar] [CrossRef] [Green Version]
- Symes, L.B.; Wershoven, N.L.; Hoeger, L.-O.; Ralston, J.S.; Martinson, S.J.; Ter Hofstede, H.M.; Palmer, C.M. Applying and Refining DNA Analysis to Determine the Identity of Plant Material Extracted from the Digestive Tracts of Katydids. PeerJ 2019, 7, e6808. [Google Scholar] [CrossRef] [PubMed]
- García-Robledo, C.; Erickson, D.L.; Staines, C.L.; Erwin, T.L.; John Kress, W. Tropical Plant–Herbivore Networks: Reconstructing Species Interactions Using DNA Barcodes. PLoS ONE 2013, 8, e52967. [Google Scholar] [CrossRef]
- Kress, W.J.; García-Robledo, C.; Uriarte, M.; Erickson, D.L. DNA Barcodes for Ecology, Evolution, and Conservation. Trends Ecol. Evol. 2015, 30, 25–35. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L.; Jones, F.A.; Swenson, N.G.; Perez, R.; Sanjur, O.; Bermingham, E. Plant DNA Barcodes and a Community Phylogeny of a Tropical Forest Dynamics Plot in Panama. Proc. Natl. Acad. Sci. USA 2009, 106, 18621–18626. [Google Scholar] [CrossRef] [Green Version]
- Jones, F.A.; Erickson, D.L.; Bernal, M.A.; Bermingham, E.; Kress, W.J.; Herre, E.A.; Muller-Landau, H.C.; Turner, B.L. The Roots of Diversity: Below Ground Species Richness and Rooting Distributions in a Tropical Forest Revealed by DNA Barcodes and Inverse Modeling. PLoS ONE 2011, 6, e24506. [Google Scholar] [CrossRef]
- Nickle, D.A. Katydids of Panama (Orthoptera: Tettigoniidae). In Insect Panama Mesoamerica; Oxford University Press: Oxford, UK, 1992; pp. 142–184. [Google Scholar]
- Encyclopedia of Life. Available online: http://eol.org (accessed on 8 January 2022).
- Condit, R.; Hubbell, S.P.; Lafrankie, J.V.; Sukumar, R.; Manokaran, N.; Foster, R.B.; Ashton, P.S. Species-Area and Species-Individual Relationships for Tropical Trees: A Comparison of Three 50-Ha Plots. J. Ecol. 1996, 84, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat Associations of Trees and Shrubs in a 50-Ha Neotropical Forest Plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Croat, T.B. Flora of Barro Colorado Island; Stanford University Press: Redwood City, CA, USA, 1978; ISBN 9780804709507. [Google Scholar]
- Coley, P.D.; Kursor, T.A. Anti-Herbivore Defenses of Young Tropical Leaves: Physiological Constraints and Ecological Trade-Offs. In Tropical Forest Plant Ecophysiology; Mulkey, S.S., Chazdon, R.L., Smith, A.P., Eds.; Springer: Boston, MA, USA, 1996; pp. 305–336. ISBN 9781461311638. [Google Scholar]
- Aide, T.M. Others Comparison of Herbivory and Plant Defenses in Temperate and Tropical Broad-Leaved Forests. In Plant-Animal Interactions: Evolution Ecology in Tropical and Temperate Regions; John Wiley and Sons: Hoboken, NJ, USA, 1991; pp. 25–49. [Google Scholar]
- Aide, T.M. Patterns of Leaf Development and Herbivory in a Tropical Understory Community. Ecology 1993, 74, 455–466. [Google Scholar] [CrossRef]
- Lieberman, D.; Lieberman, M. The Causes and Consequences of Synchronous Flushing in a Dry Tropical Forest. Biotropica 1984, 16, 193–201. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Knight, A.L.; Byers, J.A.; Judd, G.J.R.; Suckling, D.M. Caterpillar-Induced Plant Volatiles Attract Conspecific Adults in Nature. Sci. Rep. 2016, 6, 37555. [Google Scholar] [CrossRef] [Green Version]
- Szendrei, Z.; Rodriguez-Saona, C. A Meta-Analysis of Insect Pest Behavioral Manipulation with Plant Volatiles. Entomol. Exp. Appl. 2010, 134, 201–210. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Katydid Species | Plant Order | Plant Family | Plant Genus | Plant Species | Growth Habit |
|---|---|---|---|---|---|
| Anaulacomera “wallace” | Laurales (2 *) | Lauraceae (2 *) | Nectandra (2 *) | lineata (2 *) | Understory tree (10–25 m) |
| Anaulacomera furcata (v) | Ericales * | Sapotaceae * | Pouteria * | fossicola * | Understory tree (10–25 m) |
| Fabales (2 + 1 *) | Fabaceae (2 + 1 *) | Inga | sp | ||
| Swartzia * | simplex * | Understory tree (10–25 m) | |||
| Tachigali | versicolor | Canopy tree (>25 m) | |||
| Laurales * | Lauraceae * | Nectandra * | lineata * | Understory tree (10–25 m) | |
| Rosales * | Cannabaceae * | Trema * | micrantha * | Understory tree (10–25 m) | |
| Sapindales | Sapindaceae | Cupania | cinerea | Understory tree (10–25 m) | |
| Anaulacomera spatulata | Ericales | Lecythidaceae | Gustavia | superba | Understory tree (10–25 m) |
| Fabales (1 + 2 *) | Fabaceae (1+2 *) | Inga (2 *) | goldmanii | Understory tree (10–25 m) | |
| punctata | Understory tree (10–25 m) | ||||
| – | – | ||||
| Laurales (3 *) | Lauraceae (3 *) | Nectandra (3 *) | lineata (3 *) | Understory tree (10–25 m) | |
| Sapindales | Sapindaceae | Cupania | rufescens | Understory tree (10–25 m) | |
| Arota festae | Fabales * | Fabaceae * | Inga * | – | |
| Malpighiales * | Malpighiaceae * | Malpighia * | romeroana * | Shrub (<6 m) | |
| Sapindales * | Anacardiaceae * | Anacardium * | excelsum * | Canopy tree (>25 m) | |
| Ceraia mytra | Lamiales | Bignoniaceae | Jacaranda | copaia | Canopy tree (>25 m) |
| Docidocercus gigliotosi | Malpighiales | Malpighiaceae | Malpighia | romeroana | Shrub (<6 m) |
| Dolichocercus latipennis | Celastrales (1 + 1 *) | Celastraceae (1 + 1 *) | Maytenus (1 + 1*) | schippii (1 + 1 *) | Understory tree (10–25 m) |
| Laurales (6 + 1 *) | Lauraceae (6 + 1 *) | Nectandra (5 + 1*) | lineata (5 + 1 *) | Understory tree (10–25 m) | |
| Ocotea | – | Understory tree (10–25 m) | |||
| Malvales | Malvaceae | – | – | ||
| Santalales | Olacaceae | Heisteria | concinna | Understory tree (10–25 m) | |
| Sapindales | Meliaceae | Guarea | – | Understory tree (10–25 m) | |
| Euceraia insignis | Sapindales (2 *) | Anacardiaceae * | Anacardium* | excelsum* | Canopy tree (>25 m) |
| Sapindaceae * | Cupania* | – | Understory tree (10–25 m) | ||
| Hyperphrona irregularis | Fabales * | Fabaceae * | Inga* | – | |
| Idiarthron major | Fabales (2 *) | Fabaceae (2 *) | Dipteryx* | oleifera | Canopy tree (>25 m) |
| Inga* | – | ||||
| Lamprophyllum micans | Gentianales | Rubiaceae | Chimarrhis | parviflora | Understory tree (10–25 m) |
| Laurales (1 + 1 *) | Lauraceae (1 + 1 *) | Nectandra (1 + 1 *) | lineata (1 + 1 *) | Understory tree (10–25 m) | |
| Sapindales (1 + 1 *) | Anacardiaceae (1 + 1 *) | Anacardium (1 + 1 *) | excelsum (1 + 1 *) | Canopy tree (>25 m) | |
| Microcentrum “polka” | Fabales * | Fabaceae * | Inga* | – | |
| Microcentrum championi | Laurales | Lauraceae | Ocotea | puberula | Understory tree (10–25 m) |
| Malvales | Malvaceae | Luehea | seemannii | Canopy tree (>25 m) | |
| Sapindales (1 + 1 *) | Anacardiaceae * | Spondias * | radlkoferi * | Canopy tree (>25 m) | |
| Sapindaceae | Cupania | cinerea | Understory tree (10–25 m) | ||
| Montezumina bradleyi | Fabales (3) | Fabaceae (3) | Inga (3) | – | |
| Laurales (3) | Lauraceae (3) | Nectandra (3) | lineata (3) | Understory tree (10–25 m) | |
| Phylloptera dimidiata | Caryophyllales | Nyctaginaceae | Guapira | standleyana | Canopy tree (>25 m) |
| Fabales * | Fabaceae * | Swartzia * | simplex * | Understory tree (10–25 m) | |
| Malpighiales (1 + 1 *) | Malpighiaceae | Malpighia | romeroana | Shrub (<6 m) | |
| Phyllanthaceae * | Margaritaria * | nobilis * | Understory tree (10–25 m) | ||
| Malvales (2 + 1 *) | Malvaceae (2 + 1 *) | Ceiba (2 + 1 *) | pentandra (2 + 1 *) | Canopy tree (>25 m) | |
| Sapindales * | Sapindaceae * | Cupania * | latifolia * | Understory tree (10–25 m) | |
| “Waxy” sp. | Fabales | Fabaceae | Inga | sp | |
| Sapindales | Anacardiaceae | Anacardium | excelsum | Canopy tree (>25 m) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmer, C.M.; Wershoven, N.L.; Martinson, S.J.; ter Hofstede, H.M.; Kress, W.J.; Symes, L.B. Patterns of Herbivory in Neotropical Forest Katydids as Revealed by DNA Barcoding of Digestive Tract Contents. Diversity 2022, 14, 152. https://doi.org/10.3390/d14020152
Palmer CM, Wershoven NL, Martinson SJ, ter Hofstede HM, Kress WJ, Symes LB. Patterns of Herbivory in Neotropical Forest Katydids as Revealed by DNA Barcoding of Digestive Tract Contents. Diversity. 2022; 14(2):152. https://doi.org/10.3390/d14020152
Chicago/Turabian StylePalmer, Christine M., Nicole L. Wershoven, Sharon J. Martinson, Hannah M. ter Hofstede, W. John Kress, and Laurel B. Symes. 2022. "Patterns of Herbivory in Neotropical Forest Katydids as Revealed by DNA Barcoding of Digestive Tract Contents" Diversity 14, no. 2: 152. https://doi.org/10.3390/d14020152