Diversity and Structure of Soil Fungal Communities across Experimental Everglades Tree Islands

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Environmental Data Collection
2.2. Soil Collection
2.3. DNA Sequencing and Data Processing
2.4. Data Analyses
3. Results
3.1. Sequencing and ESV Taxonomic Assignments Statistics
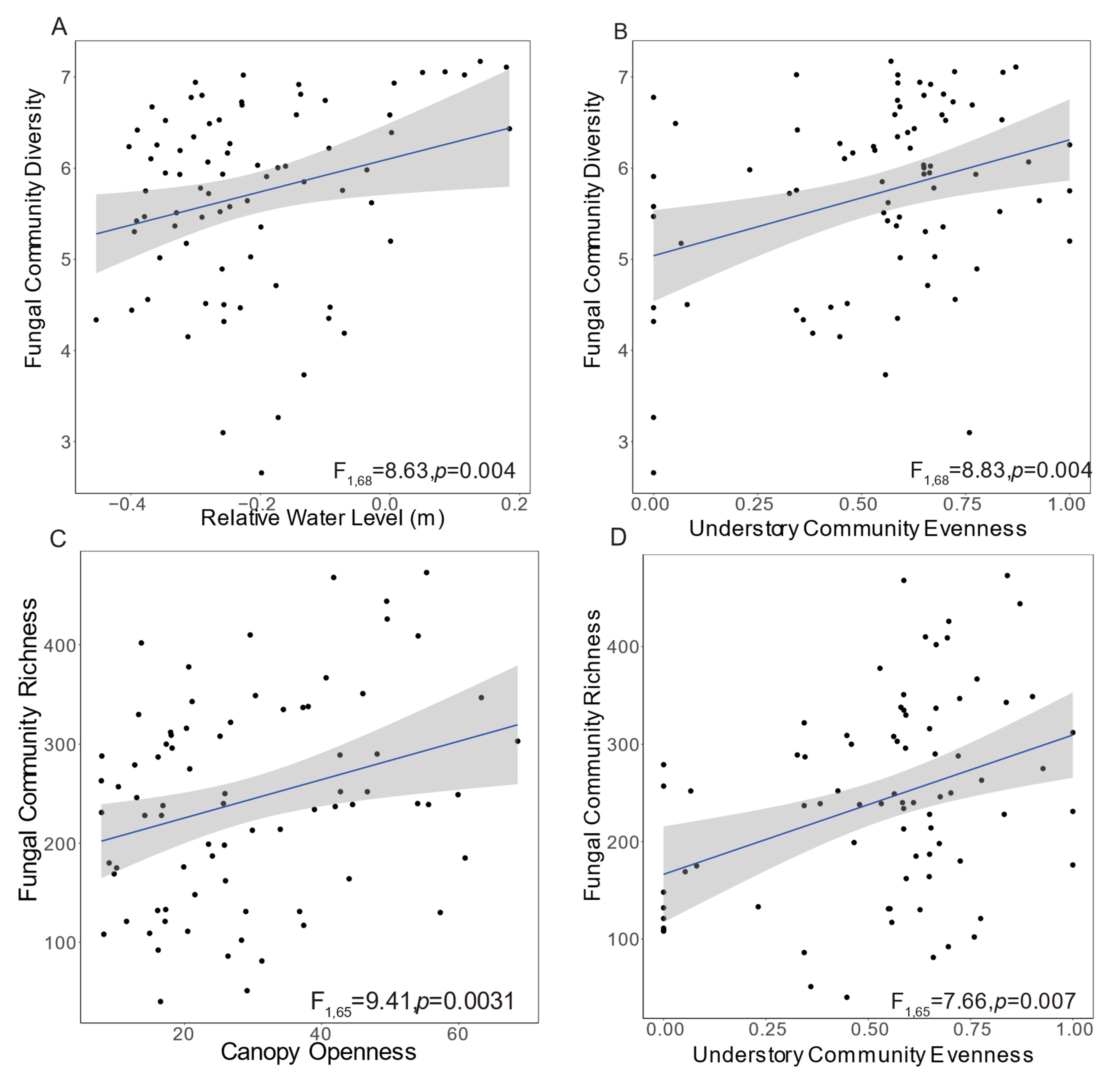
3.2. Fungal Community Diversity and Richness
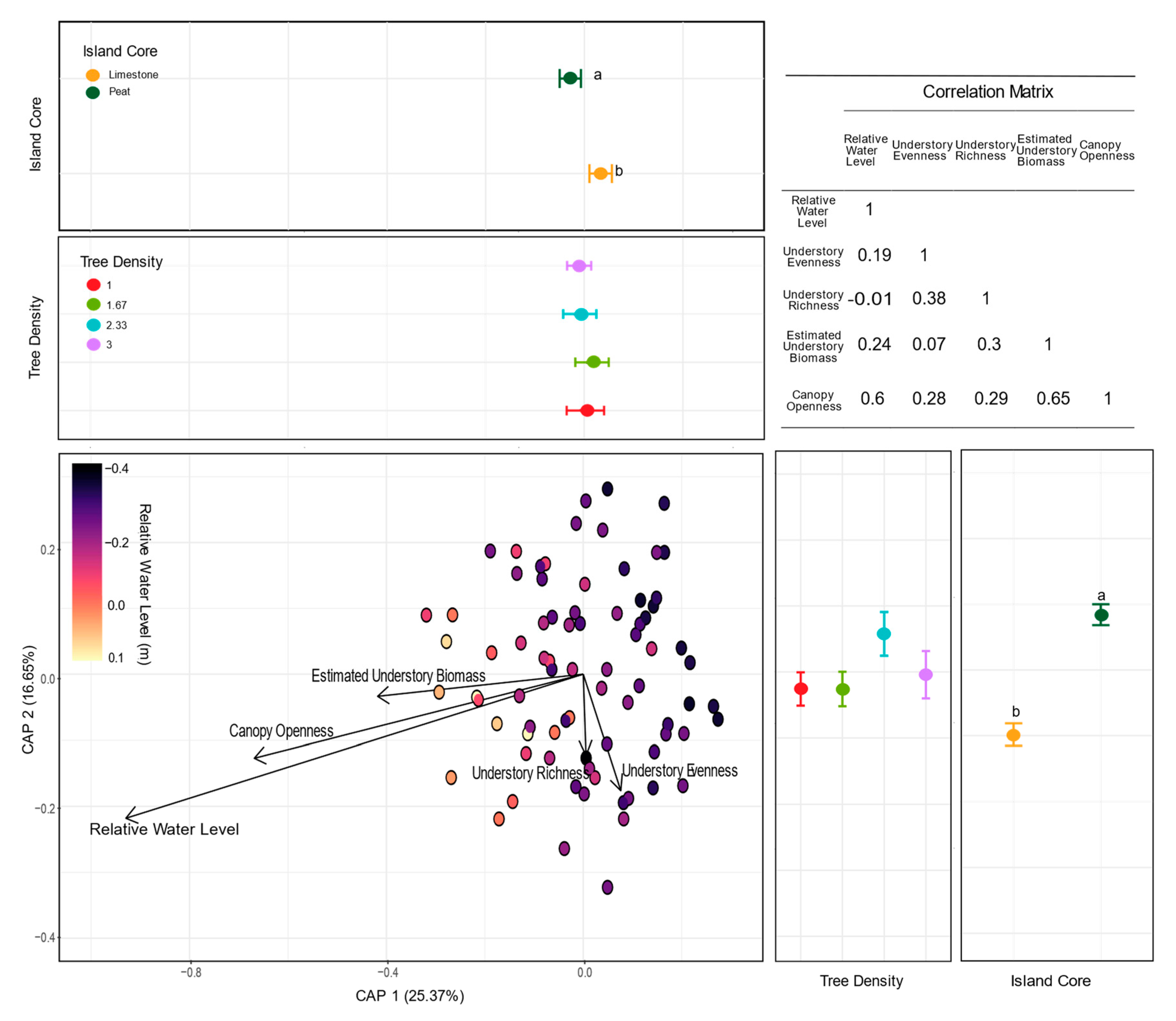
3.3. Fungal Community Composition
3.4. Distribution of Fungal Functional Guilds
4. Discussion
4.1. Abiotic Factors: The Role of Hydrology, Island Core Type, and Light Environment in Fungal Diversity and Composition
4.2. Biotic Factors: The Role of Understory Plant Communities and Tree Density in Fungal Community Diversity and Composition
4.3. Fungal Functional Guilds on Tree Islands
5. Conclusions and Future Work
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Treseder, K.K.; Lennon, J.T. Fungal traits that drive ecosystem dynamics on land. Microbiol. Mol. Biol. Rev. 2015, 79, 243–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dighton, J. Fungi in Ecosystem Processes; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Waring, B.G.; Averill, C.; Hawkes, C.V. Differences in fungal and bacterial physiology alter soil carbon and nitrogen cycling: Insights from meta-analysis and theoretical models. Ecol. Lett. 2013, 16, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Joergensen, R.G.; Emmerling, C. Methods for evaluating human impact on soil microorganisms based on their activity, biomass, and diversity in agricultural soils. J. Plant Nutr. Soil Sci. 2006, 169, 295–309. [Google Scholar] [CrossRef]
- Gougoulias, C.; Clark, J.M.; Shaw, L.J. The role of soil microbes in the global carbon cycle: Tracking the below-ground microbial processing of plant-derived carbon for manipulating carbon dynamics in agricultural systems. J. Sci. Food Agric. 2014, 94, 2362–2371. [Google Scholar] [CrossRef] [PubMed]
- Heilmann-Clausen, J.; Barron, E.S.; Boddy, L.; Dahlberg, A.; Griffith, G.W.; Nordén, J.; Ovaskainen, O.; Perini, C.; Senn-Irlet, B.; Halme, P. A fungal perspective on conservation biology. Conserv. Biol. 2015, 29, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Ritz, K.; Young, I.M. Interactions between soil structure and fungi. Mycologist 2004, 18, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [Green Version]
- Latef, A.A.H.A.; Hashem, A.; Rasool, S.; Abd_Allah, E.F.; Alqarawi, A.A.; Egamberdieva, D.; Jan, S.; Anjum, N.A.; Ahmad, P. Arbuscular mycorrhizal symbiosis and abiotic stress in plants: A review. J. Plant Biol. 2016, 59, 407–426. [Google Scholar] [CrossRef]
- Ferrol, N.; Azcón-Aguilar, C.; Pérez-Tienda, J. Review: Arbuscular mycorrhizas as key players in sustainable plant phosphorus acquisition: An overview on the mechanisms involved. Plant Sci. 2019, 280, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal Biodiversity and Their Role in Soil Health. Front. Microbiol. 2018, 9, 707. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.R.; Rillig, M.C. Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol. 2018, 220, 1059–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avis, P.G.; Gaswick, W.C.; Tonkovich, G.S.; Leacock, P.R. Monitoring fungi in ecological restorations of coastal Indiana, U.S.A. Restor. Ecol. 2017, 25, 92–100. [Google Scholar] [CrossRef]
- Barea, J.M.; Palenzuela, J.; Cornejo, P.; Sánchez-Castro, I.; Navarro-Fernández, C.; Lopéz-García, A.; Estrada, B.; Azcón, R.; Ferrol, N.; Azcón-Aguilar, C. Ecological and functional roles of mycorrhizas in semi-arid ecosystems of Southeast Spain. J. Arid Environ. 2011, 75, 1292–1301. [Google Scholar] [CrossRef]
- Asmelash, F.; Bekele, T.; Birhane, E. The Potential Role of Arbuscular Mycorrhizal Fungi in the Restoration of Degraded Lands. Front. Microbiol. 2016, 7, 1095. [Google Scholar] [CrossRef] [Green Version]
- Van der Heijden, M.G.A.; Streitwolf-Engel, R.; Riedl, R.; Siegrist, S.; Neudecker, A.; Ineichen, K.; Boller, T.; Wiemken, A.; Sanders, I.R. The mycorrhizal contribution to plant productivity, plant nutrition and soil structure in experimental grassland. New Phytol. 2006, 172, 739–752. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maltz, M.R.; Treseder, K.K. Sources of inocula influence mycorrhizal colonization of plants in restoration projects: A meta-analysis. Restor. Ecol. 2015, 23, 625–634. [Google Scholar] [CrossRef]
- Wubs, E.R.J.; van der Putten, W.H.; Bosch, M.; Bezemer, T.M. Soil inoculation steers restoration of terrestrial ecosystems. Nat Plants 2016, 2, 16107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emam, T. Local soil, but not commercial AMF inoculum, increases native and non-native grass growth at a mine restoration site. Restor. Ecol. 2016, 24, 35–44. [Google Scholar] [CrossRef]
- Rowe, H.I.; Brown, C.S.; Claassen, V.P. Comparisons of Mycorrhizal Responsiveness with Field Soil and Commercial Inoculum for Six Native Montane Species and Bromus tectorum. Restor. Ecol. 2007, 15, 44–52. [Google Scholar] [CrossRef]
- Brown, M.T.; Cohen, M.J.; Bardi, E.; Ingwersen, W.W. Species diversity in the Florida Everglades, USA: A systems approach to calculating biodiversity. Aquat. Sci. 2006, 68, 254–277. [Google Scholar] [CrossRef]
- Sklar, F.H.; Chimney, M.J.; Newman, S.; McCormick, P.; Gawlik, D.; Miao, S.; McVoy, C.; Said, W.; Newman, J.; Coronado, C.; et al. The ecological–societal underpinnings of Everglades restoration. Front. Ecol. Environ. 2005, 3, 161–169. [Google Scholar]
- Dorcas, M.E.; Willson, J.D.; Reed, R.N.; Snow, R.W.; Rochford, M.R.; Miller, M.A.; Meshaka, W.E., Jr.; Andreadis, P.T.; Mazzotti, F.J.; Romagosa, C.M.; et al. Severe mammal declines coincide with proliferation of invasive Burmese pythons in Everglades National Park. Proc. Natl. Acad. Sci. USA 2012, 109, 2418–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Valk, A.; Volin, J.; Labisky, R. Ecological effects of extreme hydrological events on the Greater Everglades. In Independent Scientific Review Panel Report to RECOVER; Bepress: Berkeley, CA, USA, 2012. [Google Scholar]
- Frohring, P.C.; Voorhees, D.P.; Kushlan, J.A. History of Wading Bird Populations in the Florida Everglades: A Lesson in the Use of Historical Information. Colon. Waterbirds 1988, 11, 328. [Google Scholar] [CrossRef]
- Wetzel, P.R.; van der Valk, A.G.; Newman, S.; Gawlik, D.E.; Gann, T.T.; Coronado-Molina, C.A.; Childers, D.L.; Sklar, F.H. Maintaining tree islands in the Florida Everglades: Nutrient redistribution is the key. Front. Ecol. Environ. 2005, 3, 370–376. [Google Scholar] [CrossRef]
- Meshaka, W.E.; Snow, R.; Bass, O.L.; Robertson, W.B. Occurrence of Wildlife on Tree Islands in the Southern Everglades. In Tree Islands of the Everglades; Springer: Berlin/Heidelberg, Germany, 2002; pp. 391–427. [Google Scholar]
- Wetzel, P.R.; Sah, J.P.; Ross, M.S. Tree islands: The bellwether of Everglades ecosystem function and restoration success. Restor. Ecol. 2017, 25, S71–S85. [Google Scholar] [CrossRef]
- Wetzel, P.R.; van der Valk, A.G.; Newman, S.; Coronado, C.A.; Troxler-Gann, T.G.; Childers, D.L.; Orem, W.H.; Sklar, F.H. Heterogeneity of phosphorus distribution in a patterned landscape, the Florida Everglades. Plant Ecol. 2009, 200, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Sklar, F.H.; van der Valk, A. Tree Islands of the Everglades: An Overview. In Tree Islands of the Everglades; Springer: Berlin/Heidelberg, Germany, 2002; pp. 1–18. [Google Scholar]
- Doren, R.F.; Armentano, T.V.; Whiteaker, L.D.; Jones, R.D. Marsh vegetation patterns and soil phosphorus gradients in the Everglades ecosystem. Aquat. Bot. 1997, 56, 145–163. [Google Scholar] [CrossRef]
- Jones, D.T.; Sah, J.P.; Ross, M.S.; Oberbauer, S.F.; Hwang, B.; Jayachandran, K. Responses of twelve tree species common in Everglades tree islands to simulated hydrologic regimes. Wetlands 2006, 26, 830–844. [Google Scholar] [CrossRef]
- Leifheit, E.F.; Veresoglou, S.D.; Lehmann, A.; Kathryn Morris, E.; Rillig, M.C. Multiple factors influence the role of arbuscular mycorrhizal fungi in soil aggregation—A meta-analysis. Plant Soil 2014, 374, 523–537. [Google Scholar] [CrossRef]
- Grinhut, T.; Hadar, Y.; Chen, Y. Degradation and transformation of humic substances by saprotrophic fungi: Processes and mechanisms. Fungal Biol. Rev. 2007, 21, 179–189. [Google Scholar] [CrossRef]
- Dobson, A.; Crawley, M. Pathogens and the structure of plant communities. Trends Ecol. Evol. 1994, 9, 393–398. [Google Scholar] [CrossRef]
- Gottlieb, A.D.; Richards, J.H.; Gaiser, E.E. Comparative study of periphyton community structure in long and short-hydroperiod Everglades marshes. Hydrobiologia 2006, 569, 195–207. [Google Scholar] [CrossRef]
- Pisani, O.; Gao, M.; Maie, N.; Miyoshi, T.; Childers, D.L.; Jaffé, R. Compositional aspects of herbaceous litter decomposition in the freshwater marshes of the Florida Everglades. Plant Soil 2018, 423, 87–98. [Google Scholar] [CrossRef]
- Wallace, B.; Dickinson, C.H. Peat Microfungi in Three Habitats in the Florida Everglades. Mycologia 1978, 70, 1151. [Google Scholar] [CrossRef]
- Aich, S.; Ewe, S.M.L.; Gu, B.; Dreschel, T.W. An evaluation of peat loss from an Everglades tree island, Florida, USA. Mires Peat 2014, 14, 1–15. [Google Scholar]
- Stoffella, S.L.; Ross, M.S.; Sah, J.P.; Price, R.M.; Sullivan, P.L.; Cline, E.A.; Scinto, L.J. Survival and growth responses of eight Everglades tree species along an experimental hydrological gradient on two tree island types. Appl. Veg. Sci. 2010, 13, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Sah, P.J.; Richards, J.H.; Ross, M.S.; Stoffella, S.L.; Cline, E.; Dreschel, T.W.; Sklar, F.H. Understory Vegetation Composition and Biomass on the Tree Islands in the Loxahatchee Impoundment Landscape Assessment (LILA) Experimental Site. in preparation.
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Gap Light Analyzer (Gla), Version 2.0: Imaging Software to Extract Canopy Structure and Gap Light Transmission Indices from True-Colour Fisheye Photographs, Users Manual and Program Documentation; Simon Fraser University: Burnaby, BC, Canada; Institute of Ecosystem Studies: New York, NY, USA, 1999; 36p. [Google Scholar]
- Gohl, D.M.; Vangay, P.; Garbe, J.; MacLean, A.; Hauge, A.; Becker, A.; Gould, T.J.; Clayton, J.B.; Johnson, T.J.; Hunter, R.; et al. Systematic improvement of amplicon marker gene methods for increased accuracy in microbiome studies. Nat. Biotechnol. 2016, 34, 942–949. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Glassman, S.I.; Martiny, J.B.H. Broadscale Ecological Patterns Are Robust to Use of Exact Sequence Variants versus Operational Taxonomic Units. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, K. MuMIn: Multi-Model Inference, R Package Version 0.12.0. Available online: http://r-forge.r-project.org/projects/mumin/ (accessed on 29 June 2020).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Wagner, H. Vegan: Community Ecology Package. R Package Version 1.17-4. Available online: http://CRAN.R-project.org/package=vegan (accessed on 29 June 2020).
- De Caceres, M.; Jansen, F.; Dell, N. Indicspecies: Studying the Statistical Relationship between Species and Groups of Sites. R Package Version 1. Available online: https://cran.r-project.org/web/packages/indicspecies/index.html (accessed on 29 June 2020).
- Dufrene, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Prinzing, A.; Durka, W.; Klotz, S.; Brandl, R. Geographic variability of ecological niches of plant species: Are competition and stress relevant? Ecography 2002, 25, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, A.A.; Ackerly, D.D.; Adler, F.; Elizabeth Arnold, A.; Cáceres, C.; Doak, D.F.; Post, E.; Hudson, P.J.; Maron, J.; Mooney, K.A.; et al. Filling key gaps in population and community ecology. Front. Ecol. Environ. 2007, 5, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Brandt, L.A.; Ecker, D.; Rivera, I.G.; Traut, A.; Mazzotti, F.J. Wildlife and vegetation of bayhead islands in the A.R.M. Loxahatchee National Wildlife Refuge. Southeast. Nat. 2003, 2, 179–194. [Google Scholar] [CrossRef]
- Gann, T.T.; Childers, D.L. Relationships between Hydrology and Soils Describe Vegetation Patterns in Seasonally Flooded Tree Islands of the Southern Everglades, Florida. Plant Soil 2006, 279, 271–286. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Y.; Qiu, Q.; Xin, G.; Yang, Z.; Shi, S. Flooding greatly affects the diversity of arbuscular mycorrhizal fungi communities in the roots of wetland plants. PLoS ONE 2011, 6, e24512. [Google Scholar] [CrossRef]
- Yang, H.; Koide, R.T.; Zhang, Q. Short-term waterlogging increases arbuscular mycorrhizal fungal species richness and shifts community composition. Plant Soil 2016, 404, 373–384. [Google Scholar] [CrossRef]
- Dobbs, C.G. On the primary dispersal and isolation of fungal spores. New Phytol. 1942, 41, 63–69. [Google Scholar] [CrossRef]
- Light, S.S.; Dineen, J.W. Water control in the Everglades: A historical perspective. In The Ecosystem and Its Restoration; St. Lucie Press: Boca Raton, FL, USA, 1994; Volume 5, pp. 47–84. [Google Scholar]
- Willard, D.A.; Bernhardt, C.E.; Holmes, C.W.; Landacre, B.; Marot, M. Response of Everglades tree islands to environmental change. Ecol. Monogr. 2006, 76, 565–583. [Google Scholar] [CrossRef]
- Van Nuland, M.E.; Peay, K.G. Symbiotic niche mapping reveals functional specialization by two ectomycorrhizal fungi that expands the host plant niche. Fungal Ecol. 2020, 46, 100960. [Google Scholar] [CrossRef]
- Bougher, N.L.; Matheny, P.B. Two species of Inocybe (fungi) introduced into Western Australia. Nuytsia 2011, 3, 139–148. [Google Scholar]
- Rosling, A.; Cox, F.; Cruz-Martinez, K.; Ihrmark, K.; Grelet, G.-A.; Lindahl, B.D.; Menkis, A.; James, T.Y. Archaeorhizomycetes: Unearthing an ancient class of ubiquitous soil fungi. Science 2011, 333, 876–879. [Google Scholar] [CrossRef]
- Morgenstern, I.; Klopman, S.; Hibbett, D.S. Molecular evolution and diversity of lignin degrading heme peroxidases in the Agaricomycetes. J. Mol. Evol. 2008, 66, 243–257. [Google Scholar] [CrossRef]
- Schadt, C.W.; Martin, A.P.; Lipson, D.A.; Schmidt, S.K. Seasonal dynamics of previously unknown fungal lineages in tundra soils. Science 2003, 301, 1359–1361. [Google Scholar] [CrossRef] [Green Version]
- Morón-Ríos, A.; Gómez-Cornelio, S.; Ortega-Morales, B.O.; De la Rosa-García, S.; Partida-Martínez, L.P.; Quintana, P.; Alayón-Gamboa, J.A.; Cappello-García, S.; González-Gómez, S. Interactions between abundant fungal species influence the fungal community assemblage on limestone. PLoS ONE 2017, 12, e0188443. [Google Scholar] [CrossRef] [Green Version]
- Subedi, S.C.; Ross, M.S.; Scinto, L.J. Nutrient Limitation in Two Everglades Tree Species Planted on Constructed Tree Islands. Wetlands 2012, 32, 1163–1173. [Google Scholar] [CrossRef]
- Sullivan, P.L.; Price, R.M.; Ross, M.S.; Scinto, L.J.; Stoffella, S.L.; Cline, E.; Dreschel, T.W.; Sklar, F.H. Hydrologic processes on tree islands in the Everglades (Florida, USA): Tracking the effects of tree establishment and growth. Hydrogeol. J. 2011, 19, 367–378. [Google Scholar] [CrossRef]
- Estrada-Medina, H.; Canto-Canché, B.B.; De Los Santos-Briones, C.; O’Connor-Sánchez, A. Yucatán in black and red: Linking edaphic analysis and pyrosequencing-based assessment of bacterial and fungal community structures in the two main kinds of soil of Yucatán State. Microbiol. Res. 2016, 188, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Rose, S.L. Mycorrhizal associations of some actinomycete nodulated nitrogen-fixing plants. Can. J. Bot. 1980, 58, 1449–1454. [Google Scholar] [CrossRef]
- Ma, S.; Verheyen, K.; Props, R.; Wasof, S.; Vanhellemont, M.; Boeckx, P.; Boon, N.; De Frenne, P. Plant and soil microbe responses to light, warming and nitrogen addition in a temperate forest. Funct. Ecol. 2018, 32, 1293–1303. [Google Scholar] [CrossRef]
- Bartemucci, P.; Messier, C.; Canham, C.D. Overstory influences on light attenuation patterns and understory plant community diversity and composition in southern boreal forests of Quebec. Can. J. For. Res. 2006, 36, 2065–2079. [Google Scholar] [CrossRef] [Green Version]
- Kulmatiski, A.; Beard, K.H. Long-term plant growth legacies overwhelm short-term plant growth effects on soil microbial community structure. Soil Biol. Biochem. 2011, 43, 823–830. [Google Scholar] [CrossRef]
- McGuire, K.L.; Fierer, N.; Bateman, C.; Treseder, K.K.; Turner, B.L. Fungal community composition in neotropical rain forests: The influence of tree diversity and precipitation. Microb. Ecol. 2012, 63, 804–812. [Google Scholar] [CrossRef] [Green Version]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root exudates regulate soil fungal community composition and diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.H.; Wang, P.; Zhao, H.; Xu, X.H.; Zhu, Y.D. Impact of allelochemical exuded from allelopathic rice on soil microbial community. Soil Biol. Biochem. 2008, 40, 1862–1869. [Google Scholar] [CrossRef]
- Chapman, S.K.; Newman, G.S. Biodiversity at the plant–soil interface: Microbial abundance and community structure respond to litter mixing. Oecologia 2010, 162, 763–769. [Google Scholar] [CrossRef]
- Wang, J.; Chen, C.; Ye, Z.; Li, J.; Feng, Y.; Lu, Q. Relationships Between Fungal and Plant Communities Differ Between Desert and Grassland in a Typical Dryland Region of Northwest China. Front. Microbiol. 2018, 9, 2327. [Google Scholar] [CrossRef]
- Voříšková, J.; Baldrian, P. Fungal community on decomposing leaf litter undergoes rapid successional changes. ISME J. 2013, 7, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Sheldrake, M.; Rosenstock, N.P.; Revillini, D.; Olsson, P.A.; Mangan, S.; Sayer, E.J.; Wallander, H.; Turner, B.L.; Tanner, E.V.J. Arbuscular mycorrhizal fungal community composition is altered by long-term litter removal but not litter addition in a lowland tropical forest. New Phytol. 2017, 214, 455–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittebiere, A.; Vandenkoornhuyse, P.; Maluenda, E.; Gareil, A.; Dheilly, A.; Coudouel, S.; Bahin, M.; Mony, C. Past spatial structure of plant communities determines arbuscular mycorrhizal fungal community assembly. J. Ecol. 2020, 108, 546–560. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.I.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Matulich, K.L.; Martiny, J.B.H. Microbial composition alters the response of litter decomposition to environmental change. Ecology 2015, 96, 154–163. [Google Scholar] [CrossRef]
- Maron, J.L.; Marler, M.; Klironomos, J.N.; Cleveland, C.C. Soil fungal pathogens and the relationship between plant diversity and productivity. Ecol. Lett. 2011, 14, 36–41. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Boller, T.; Wiemken, A.; Sanders, I.R. Different arbsucular mycorrhizal fungal species are potential determinants of plant community structure. Ecology 1998, 79, 2082–2091. [Google Scholar] [CrossRef]
- Bartelt-Ryser, J.; Joshi, J.; Schmid, B.; Brandl, H.; Balser, T. Soil feedbacks of plant diversity on soil microbial communities and subsequent plant growth. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 27–49. [Google Scholar] [CrossRef]
- Arnold, A.E.; Elizabeth Arnold, A. Understanding the diversity of foliar endophytic fungi: Progress, challenges, and frontiers. Fungal Biol. Rev. 2007, 21, 51–66. [Google Scholar] [CrossRef]
- Crowther, T.W.; Boddy, L.; Hefin Jones, T. Functional and ecological consequences of saprotrophic fungus–grazer interactions. ISME J. 2012, 6, 1992–2001. [Google Scholar] [CrossRef] [Green Version]
- Dreschel, T.W.; Cline, E.A.; Hill, S.D. Everglades tree island restoration: Testing a simple tree planting technique patterned after a natural process. Restor. Ecol. 2017, 25, 696–704. [Google Scholar] [CrossRef]
- Amend, A.S.; Seifert, K.A.; Bruns, T.D. Quantifying microbial communities with 454 pyrosequencing: Does read abundance count? Mol. Ecol. 2010, 19, 5555–5565. [Google Scholar] [CrossRef] [PubMed]
- Troskie, A.M.; Vlok, N.M.; Rautenbach, M. A novel 96-well gel-based assay for determining antifungal activity against filamentous fungi. J. Microbiol. Methods 2012, 91, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-J.; van Nostrand, J.D.; Tu, Q.; Lu, Z.; Cheng, L.; Yuan, T.; Deng, Y.; Carter, M.Q.; He, Z.; Wu, L.; et al. The PathoChip, a functional gene array for assessing pathogenic properties of diverse microbial communities. ISME J. 2013, 7, 1974–1984. [Google Scholar] [CrossRef] [Green Version]
- Carvalhais, L.C.; Dennis, P.G.; Tyson, G.W.; Schenk, P.M. Application of metatranscriptomics to soil environments. J. Microbiol. Methods 2012, 91, 246–251. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Island Factors | Fungal Diversity | Fungal Richness |
|---|---|---|
| Relative Water Level | F1,68 = 8.63, p = 0.0045 | F1,65 = 2.40, p = 0.1255 |
| Island Core | -- | F1,6 = 1.18, p = 0.3188 |
| Canopy Openness | -- | F1,65 = 9.41, p = 0.0031 |
| Tree Density | -- | F1,65 = 1.99, p = 0.1626 |
| Understory Evenness | F1,68 = 8.83, p = 0.0041 | F1,65 = 7.66, p = 0.0073 |
| Understory Richness | -- | F1,65 = 1.78, p = 0.1864 |
| Estimated Understory Biomass | -- | -- |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, B.K.; Ross, M.S.; Stoffella, S.L.; Sah, J.P.; Cline, E.; Sklar, F.; Afkhami, M.E. Diversity and Structure of Soil Fungal Communities across Experimental Everglades Tree Islands. Diversity 2020, 12, 324. https://doi.org/10.3390/d12090324
Almeida BK, Ross MS, Stoffella SL, Sah JP, Cline E, Sklar F, Afkhami ME. Diversity and Structure of Soil Fungal Communities across Experimental Everglades Tree Islands. Diversity. 2020; 12(9):324. https://doi.org/10.3390/d12090324
Chicago/Turabian StyleAlmeida, Brianna K., Michael S. Ross, Susana L. Stoffella, Jay P. Sah, Eric Cline, Fred Sklar, and Michelle E. Afkhami. 2020. "Diversity and Structure of Soil Fungal Communities across Experimental Everglades Tree Islands" Diversity 12, no. 9: 324. https://doi.org/10.3390/d12090324