Feeding Specialization of Honey Badgers in the Sahara Desert: A Trial of Life in a Hard Environment


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
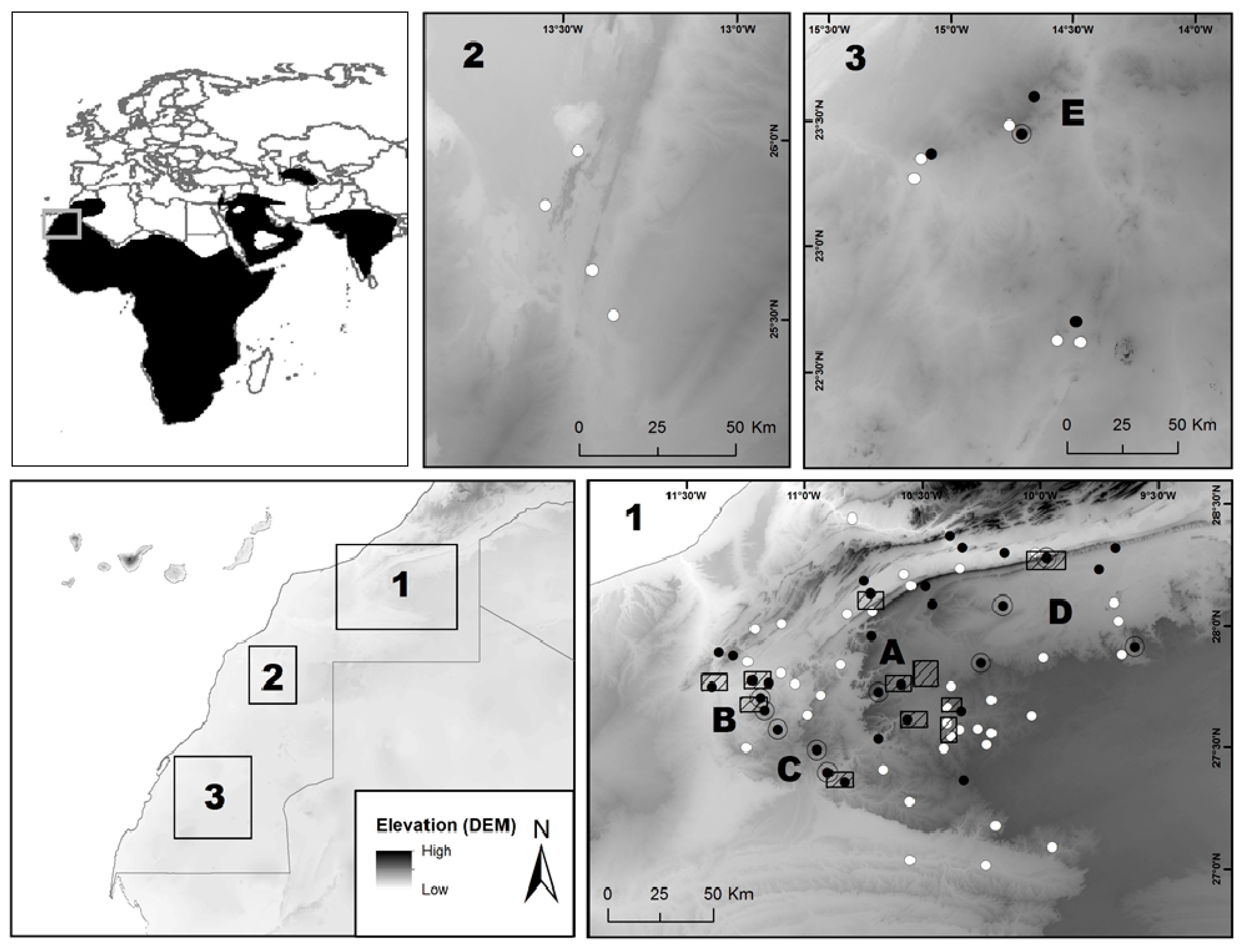
2.1. Study Area
2.2. Field Surveys
2.3. Feces Identification
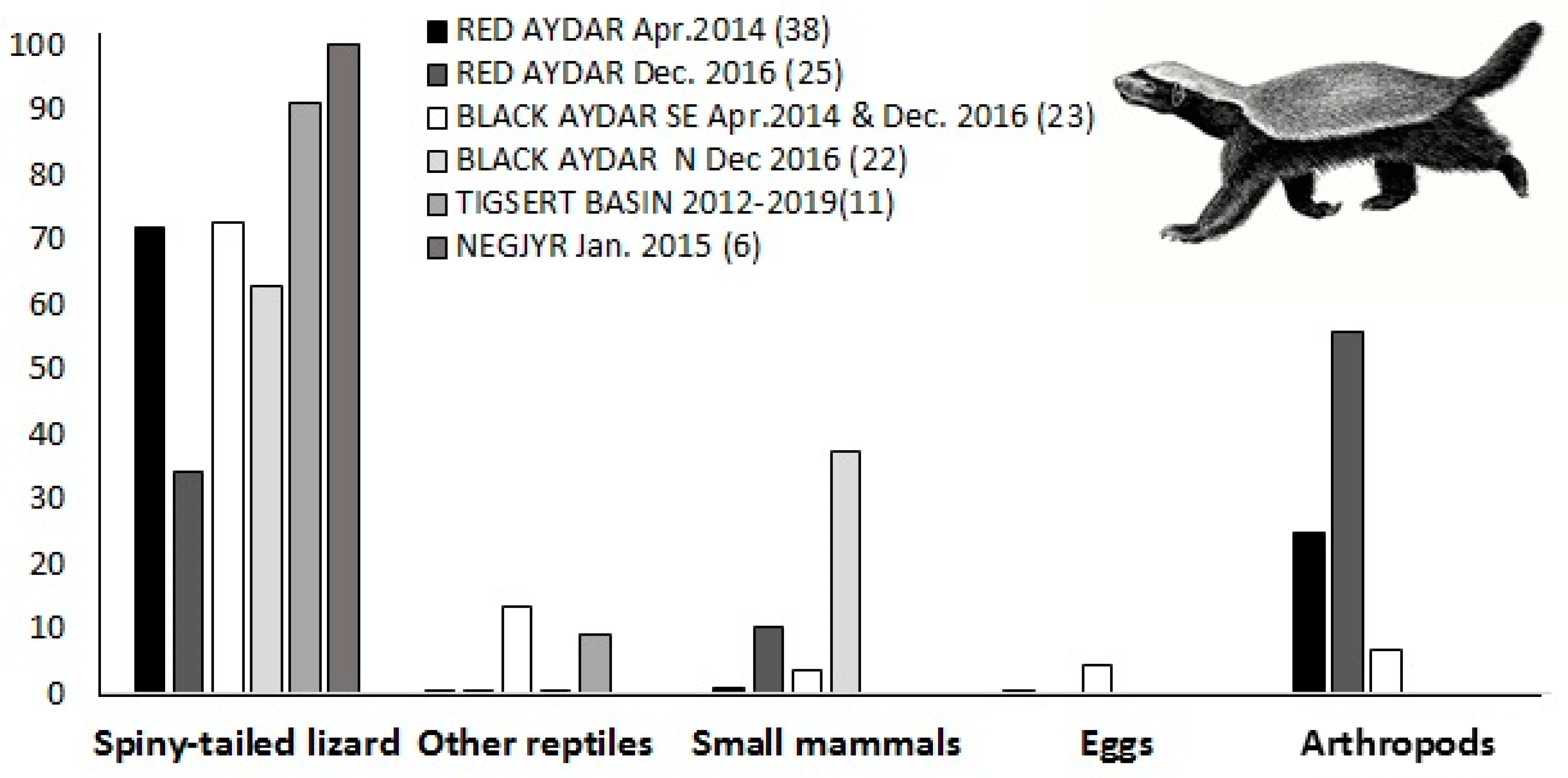
2.4. Faecal Analysis and Diet Composition
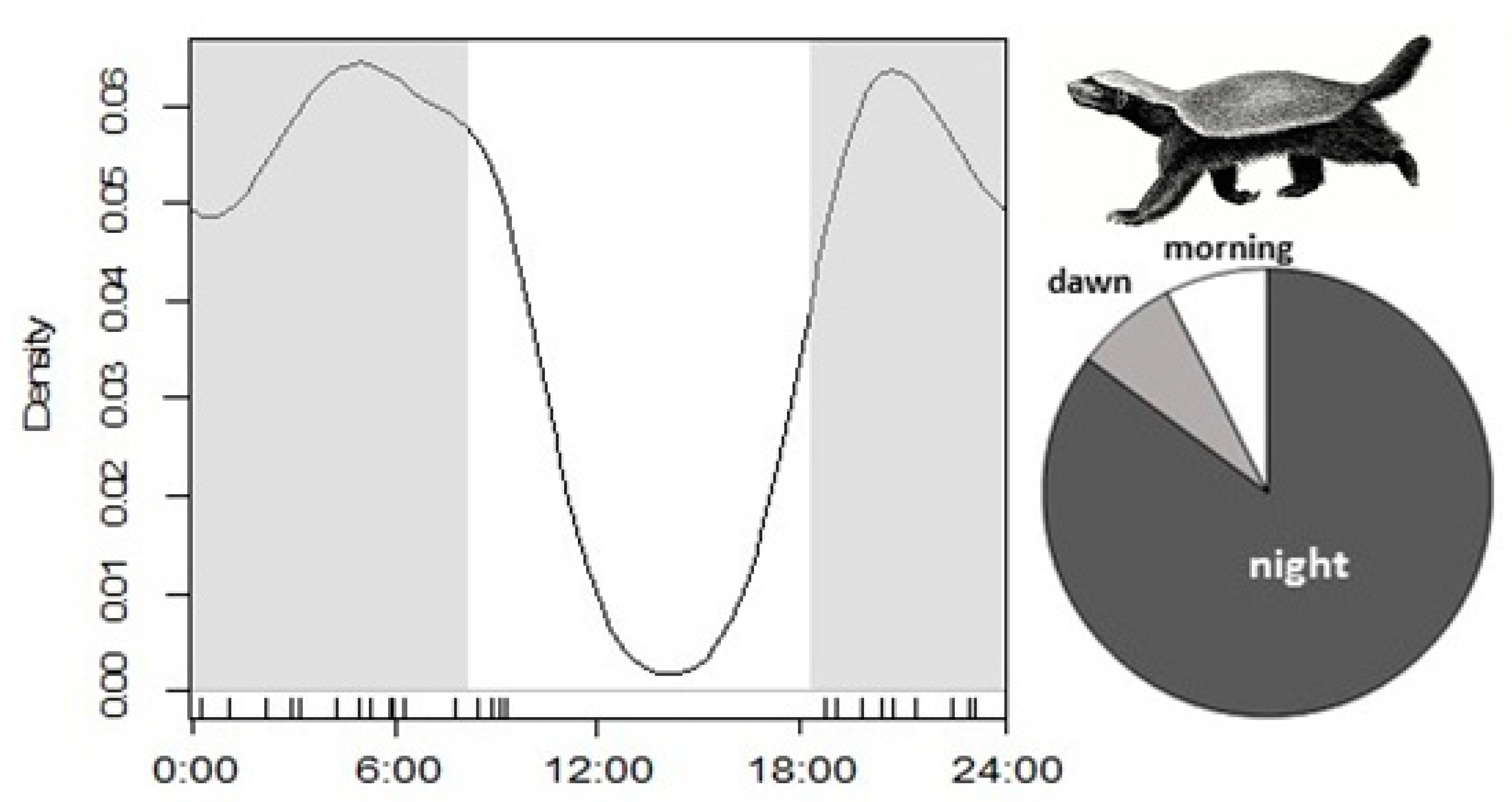
2.5. Predator-Prey Relationships: Abundance and Diel Activity of the Staple prey
2.6. Role of Staple Prey of Honey Badger in the Diet of Other Sympatric Carnivores
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, D.E.; Mittermeier, R.A. (Eds.) Handbook of the Mammals of the World. Vol 1. Carnivores; Lynx Editions: Barcelona, Spain, 2009. [Google Scholar]
- Durant, S.M.; Wacher, T.; Bashir, S.; Woodroffe, P.; de Ornellas, P.; Ransom, C.; Newby, J.; Abáigar, T.; Abdelgadir, M.; El Alqamy, H.; et al. Fidding in biodiersity hotspots while deserts burn? Collapse of the Sahara`s megafauna. Divers. Distrib. 2014, 20, 114–122. [Google Scholar] [CrossRef]
- Brito, J.C.; Acosta, A.L.; Álvares, F.; Cuzin, F. Biogeography and conservation of taxa from remote regions: An application of ecological-niche based models and GIS to North-African Canids. Biol. Conserv. 2009, 142, 3020–3029. [Google Scholar] [CrossRef]
- Belbachir, F.; Pettorelli, N.; Wacher, T.; Belbachir-Bazi, A.; Durant, S.M. Monitoring Rarity: The Critically Endangered Saharan Cheetah as a Flagship Species for a Threatened Ecosystem. PLoS ONE 2015, 10, e0115136. [Google Scholar] [CrossRef] [Green Version]
- Begg, C.; Begg, K.; Kingdon, J. Subfamily Mellivorinae. Ratel. In Mammals of Africa; Kingdon, J., Happold, D., Butynski, T., Hoffmann, M., Happold, M., Kalina, J., Eds.; Bloomsbury Natural History: London, UK, 2013; Volume V: Carnivores, Pangolins, Equids and Rhinoceroses; pp. 119–125. [Google Scholar]
- Kruuk, H. Notes on food and foraging of the honey badger Mellivora capensis in the Kalahari Gemsbok National Park. Koedoe 1983, 26, 153–157. [Google Scholar] [CrossRef]
- Begg, C.M.; Begg, K.S.; Du Toit, J.T.; Mills, M.G.L. Sexual and seasonal variation in the diet and foraging behaviour of a sexually dimorphic carnivore, the honey badger (Mellivora capensis). J. Zool. 2003, 260, 301–316. [Google Scholar] [CrossRef] [Green Version]
- Valverde, J.A. Aves del Sahara Español (estudio ecológico del desierto); Consejo Superior de Investigaciones Científicas, Instituto de Estudios africanos: Madrid, Spain, 1957. [Google Scholar]
- Dragesco-Joffé, A. La vie sauvage au Sahara; Delachaux et Niestlé: Paris, France, 1993. [Google Scholar]
- Vale, C.G.; Campos, J.C.; Silva, T.L.; Goncalves, D.V.; Sow, A.S.; Martinez-Freiria; Zbyszek Boratynski1, F.Z.; Brito, J.C. Biogeography and conservation of mammals from the West Sahara-Sahel: An application of ecological niche-based models and GIS. Hystrix 2016, 27. [Google Scholar] [CrossRef]
- Aulegnier, S.; Cuzin, F.; Thévenot, M. Mammifères sauvages au Maroc. Peuplement, répartition, ecologie; Société Française pour l’Etude et la Protection des Mammifères: Paris, France, 2017. [Google Scholar]
- Hardin, G. The Competitive Exclusion Principle. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Coyne, J.A.; Orr, H.A. Speciation; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Jdeidi, T.; Masseti, M.; Nader, I.; de Smet, K.; Cuzin, F. Mellivora capensis. The IUCN Red List of Threatened Species 2010. e.T41629A10522349. Available online: https://www.iucnredlist.org/species/41629/45210107 (accessed on 4 November 2019).
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Barea-Azcón, J.M.; Virgós, E.; Ballesteros-Duperón, E.; Moleón, M.; Chirosa, M. Surveying carnivores at large spatial scales: A comparison of four broad-applied methods. Biodivers. Conserv. 2007, 16, 1213–1230. [Google Scholar] [CrossRef]
- Robert, A.L.; MacKay, P.; William, J.Z.; Ray, J.C. Noninvasive survey methods for carnivores; Osiland Press: Washington, DC, USA; Covelo, CA, USA; London, UK, 2008. [Google Scholar]
- O’Connell, A.F.; Nichols, J.D.; Karanth, K.U. Camera Traps in Animal Ecology: Methods and Analyses; Springer: Tokyo, Japan; Dordrecht, The Netherlands; Heildelberg, Germany; London, UK; New York, NY, USA, 2011. [Google Scholar]
- Goldenberger, D.; Perschil, I.; Ritzler, M.; Altwegg, M. A simple “universal” DNA extraction procedure using SDS and proteinase K is compatible with direct PCR amplification. Genome Res. 1995, 4, 368–370. [Google Scholar] [CrossRef] [Green Version]
- Kohn, M.; Knauer, F.; Stoffella, A.; Schröder, W.; Pääbo, S. Conservation genetics of the European brown bear--a study using excremental PCR of nuclear and mitochondrial sequences. Mol. Ecol. 1995, 4, 95–103. [Google Scholar] [CrossRef]
- Leonard, J.A.; Wayne, R.K.; Cooper, A. Population genetics of Ice Age brown bears. Proc. Natl. Acad. Sci. USA 2000, 97, 1651–1654. [Google Scholar] [CrossRef] [Green Version]
- Losey, R.J.; Bazaliiskii, V.I.; Garvie-Lok, S.; Germonpré, M.; Leonard, J.A.; Allen, A.L.; Katzenberg, M.A.; Sablin, M.V. Canids as persons: Early Neolithic dof and wolf burials, Cis-Baikal, Siberia. J. Anthropol. Archaeol. 2011, 30, 174–189. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.; Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold. Spring. Harb. Protoc. 2010, 5, pdb-prot5448. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Putman, R.J. Facts from faeces. Mammal Rev. 1984, 14, 79–97. [Google Scholar] [CrossRef]
- Barea-Azcón, J.M.; Ballesteros-Duperón, E.; Gil-Sánchez, J.M.; Virgós, E. Badger Meles meles feeding ecology in dry Mediterranean environments of the southwest edge of its distribution range. Acta Theriol. 2010, 55, 45–52. [Google Scholar] [CrossRef]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Advanced Distance Sampling: Estimating Abundance of Biological Populations; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Bartlett, R. Spiny-tailed Lizards: Uromastyx and Xenagama; Barron’s Educational Series, Inc.: Hauppauge, NY, USA, 2003. [Google Scholar]
- Thomas, L.; Buckland, S.T.; Rexstad, E.A.; Laake, J.L.; Strindberg, S.; Hedley, S.L.; Bishop, J.R.; Marques, T.A.; Burnham, K.P. Distance software: Design and analysis of distance sampling surveys for estimating population size. J. App. Ecol. 2010, 47, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Ferreras, P.; Díaz-Ruiz, F.; Monterroso, P. Improving mesocarnivore detectability with lures in camera-trapping studies. Wildl. Res. 2018, 45, 505–517. [Google Scholar] [CrossRef]
- Monterroso, P.; Alves, P.C.; Ferreras, P. Catch me if you can: Diel activity patterns of mammalian prey and predators. Ethology 2013, 119, 1044–1056. [Google Scholar] [CrossRef] [Green Version]
- Ridout, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2009, 14, 322–337. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2018. Available online: http://www.R-project.org/ (accessed on 18 January 2020).
- Martínez del Mármol, G.; Harris, D.J.; Geniez, P.; de Pous, P.; Salvi, D. Amphibians and Reptiles of Morocco; Edition Chimaira: Frankfurt, Germany, 2019. [Google Scholar]
- Roper, T. The European badger Meles meles: Food specialist or generalist? J. Zool. 2009, 234, 437–452. [Google Scholar] [CrossRef]
- Stephens, D.W.; Krebs, J.R. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Schleich, H.H.; Kästel, W.; Kahisch, K. Amphibians and Reptiles of North Africa; Koeltz Sci. Books: Bonn, Germany, 1996. [Google Scholar]
- Kruuk, H. The Social Badger. Ecology and Behaviour of A Group-Living Carnivore (Meles meles); Oxford University Press: Oxford, UK, 1989. [Google Scholar]
- Fedriani, J.M.; Ferreras, P.; Delibes, M. Dietary response of the Eurasian badger, Meles meles, to a decline of its main prey in the Doñana National Park. J. Zool. 1998, 245, 214–218. [Google Scholar]
- Tamar, K.; Metallinou, M.; Wilms, T.; Schmitz, A.; Crochet, P.; Geniez, P.; Carranza, S. Evolutionary history of spiny-tailed lizards (Lizardae: Uromastyx) from the Saharo-Arabian region. Zool. Scr. 2018, 47, 159–173. [Google Scholar] [CrossRef]
- Cherkaoui, S.I.; Bouajaja, A. Recent records of the elusive Ratel Mellivora capensis (Schreber, 1776) in Morocco and case of human persecution. IUCN/SCC Small Carnivore Conservation Journal 2017, 55, 64–68. [Google Scholar]
- Donadio, E.; Buskirk, S.W. Diet, morphology, and interspecific killing in Carnivora. Am. Naturalist 2006, 167, 524–536. [Google Scholar] [CrossRef]
- Vanak, A.T.; Fortin, D.; Thaker, M.; Ogden, M.; Owen, C.; Greatwood, S.; Slotow, R. Moving to stay in place: Behavioral mechanisms for coexistence of African large carnivores. EcolM MJKgy 2013, 94, 2619–2631. [Google Scholar] [CrossRef]
- Lovari, S.; Pokheral, C.P.; Jnawali, S.R.; Fusani, L.; Ferretti, F. Coexistence of the tiger and the common leopard in a prey-rich area: The role of prey partitioning. J. Zool. 2015, 295, 122–131. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Item | Red Aydar Apr.2014 | Red Aydar Dec. 2016 | Black Aydar SE Apr.2014 & Dec. 2016 | Black Aydar N Dec 2016 | Tigsert Basin 2012–2019 | Negjyr Jan. 2015 |
|---|---|---|---|---|---|---|
| Uromastyx nigriventris | 30 | 9 | 18 | 15 | 10 | 0 |
| U. dispar | 0 | 0 | 0 | 0 | 0 | 6 |
| Agama sp./Trapelus sp. | 1 | 0 | 2 | 1 | 1 | 0 |
| Snake n.i. | 1 | 1 | 1 | 0 | 0 | 0 |
| Lepus sp. | 1 | 0 | 0 | 0 | 0 | 0 |
| Atlantoxerus getullus | 0 | 0 | 0 | 1 | 0 | 0 |
| Gerbillus campestris | 0 | 0 | 0 | 4 | 0 | 0 |
| Psammomys obesus | 0 | 2 | 0 | 4 | 0 | 0 |
| Acomys cahirinus | 0 | 1 | 0 | 0 | 0 | 0 |
| Rodentia n.i. | 0 | 1 | 1 | 1 | 0 | 0 |
| Alectoris barbara eggs | 1 | 0 | 0 | 0 | 0 | 0 |
| Reptile egs | 1 | 0 | 2 | 0 | 0 | 0 |
| Melolonthidae adults | 8 | 1 | 1 | 0 | 0 | 0 |
| Melolonthidae larvae | 18 | 11 | 1 | 0 | 0 | 0 |
| Tenebridae | 0 | 3 | 0 | 0 | 0 | 0 |
| Scorpion | 0 | 1 | 0 | 0 | 0 | 0 |
| Scolopendra | 1 | 0 | 0 | 0 | 0 | 0 |
| Artropod n.i. | 1 | 0 | 0 | 0 | 0 | 0 |
| Lepidoptera pupae | 0 | 0 | 1 | 0 | 0 | 0 |
| Mud | 1 | 0 | 0 | 0 | 0 | 0 |
| N | 38 scats | 25 scats | 23 scats | 22 scats | 11 scats | 6 scats |
| Spiny-Tailed Lizard | ||||
|---|---|---|---|---|
| Red Aydar Apr. 2016 | Red Aydar Dec. 2014 | Black Aydar SE | Tigsert Basin | |
| Red Aydar Apr. 2014 | - | |||
| Red Aydar Dec. 2016 | n.s. | - | ||
| Black Aydar SE | n.s. | 0.0001 | - | |
| Black Aydar N | n.s. | n.s. | n.s | - |
| Tigsert basin | 0.005 | 0.001 | n.s. | n.s. |
| Melolonthidae | ||||
| Red Aydar Apr. 2014 | - | |||
| Red Aydar Dec. 2014 | n.s. | - | ||
| Black Aydar SE | 0.001 | 0.001 | - | |
| Black Aydar N | 0.0001 | 0.0001 | n.s | - |
| Tigsert basin | 0.006 | 0.009 | n.s. | n.s |
| Small Mammals | ||||
| Red Aydar Apr. 2014 | - | |||
| Red Aydar Dec. 2016 | n.s. | - | ||
| Black Aydar SE | n.s. | n.s. | - | |
| Black Aydar N | 0.0001 | n.s. | 0.001 | - |
| Tigsert basin | n.s. | n.s. | n.s. | n.s. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil-Sánchez, J.M.; Herrera-Sánchez, F.J.; Rodríguez-Siles, J.; Sáez, J.M.; Díaz-Portero, M.Á.; Arredondo, Á.; Álvarez, B.; Cancio, I.; de Lucas, J.; Castillo, S.; et al. Feeding Specialization of Honey Badgers in the Sahara Desert: A Trial of Life in a Hard Environment. Diversity 2020, 12, 59. https://doi.org/10.3390/d12020059
Gil-Sánchez JM, Herrera-Sánchez FJ, Rodríguez-Siles J, Sáez JM, Díaz-Portero MÁ, Arredondo Á, Álvarez B, Cancio I, de Lucas J, Castillo S, et al. Feeding Specialization of Honey Badgers in the Sahara Desert: A Trial of Life in a Hard Environment. Diversity. 2020; 12(2):59. https://doi.org/10.3390/d12020059
Chicago/Turabian StyleGil-Sánchez, Jose María, F. Javier Herrera-Sánchez, Javier Rodríguez-Siles, Juan Manuel Sáez, Miguel Ángel Díaz-Portero, Ángel Arredondo, Begoña Álvarez, Inmaculada Cancio, Jesús de Lucas, Salvador Castillo, and et al. 2020. "Feeding Specialization of Honey Badgers in the Sahara Desert: A Trial of Life in a Hard Environment" Diversity 12, no. 2: 59. https://doi.org/10.3390/d12020059