Dactylogyrids (Platyhelminthes: Monogenoidea) Infecting the Gill Lamellae of Flatheads (Scorpaeniformes: Platycephalidae), with Proposal of Platycephalotrema n. gen. and Descriptions of New Species from Australia and Japan


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
- Subclass Polyonchoinea Bychowsky, 1937
- Order Dactylogyridea Bychowsky, 1937
- Dactylogyridae Bychowsky, 1933
3.1. Platycephalotrema n. gen.
3.1.1. Diagnosis
3.1.2. Taxonomic Summary
3.1.3. Remarks
3.2. Platycephalotrema ogawai n. sp.
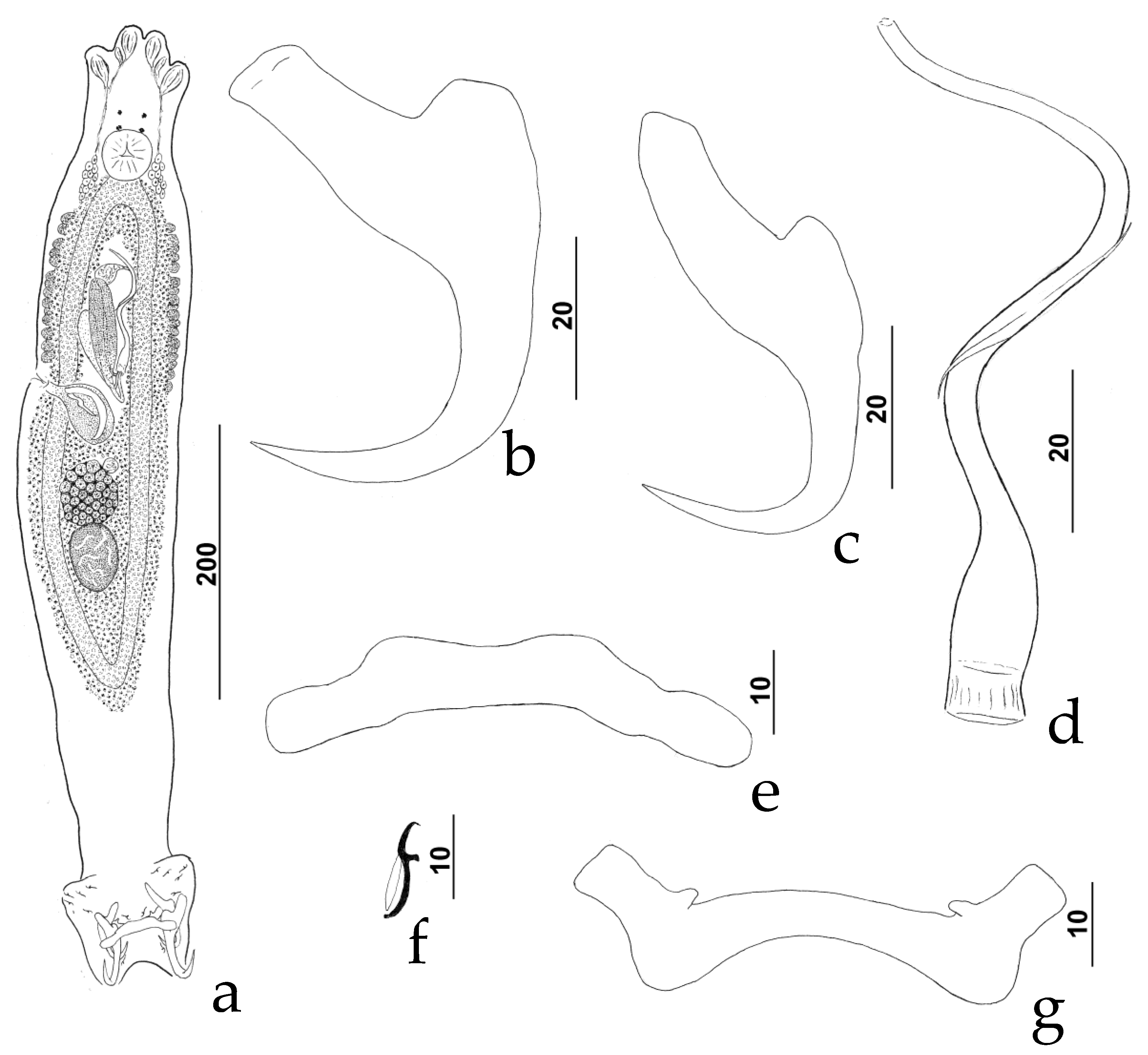
3.2.1. Description (Figure 1)
3.2.2. Measurements
3.2.3. Taxonomic Summary
3.2.4. Remarks
3.3. Platycephalotrema koppa n. sp.
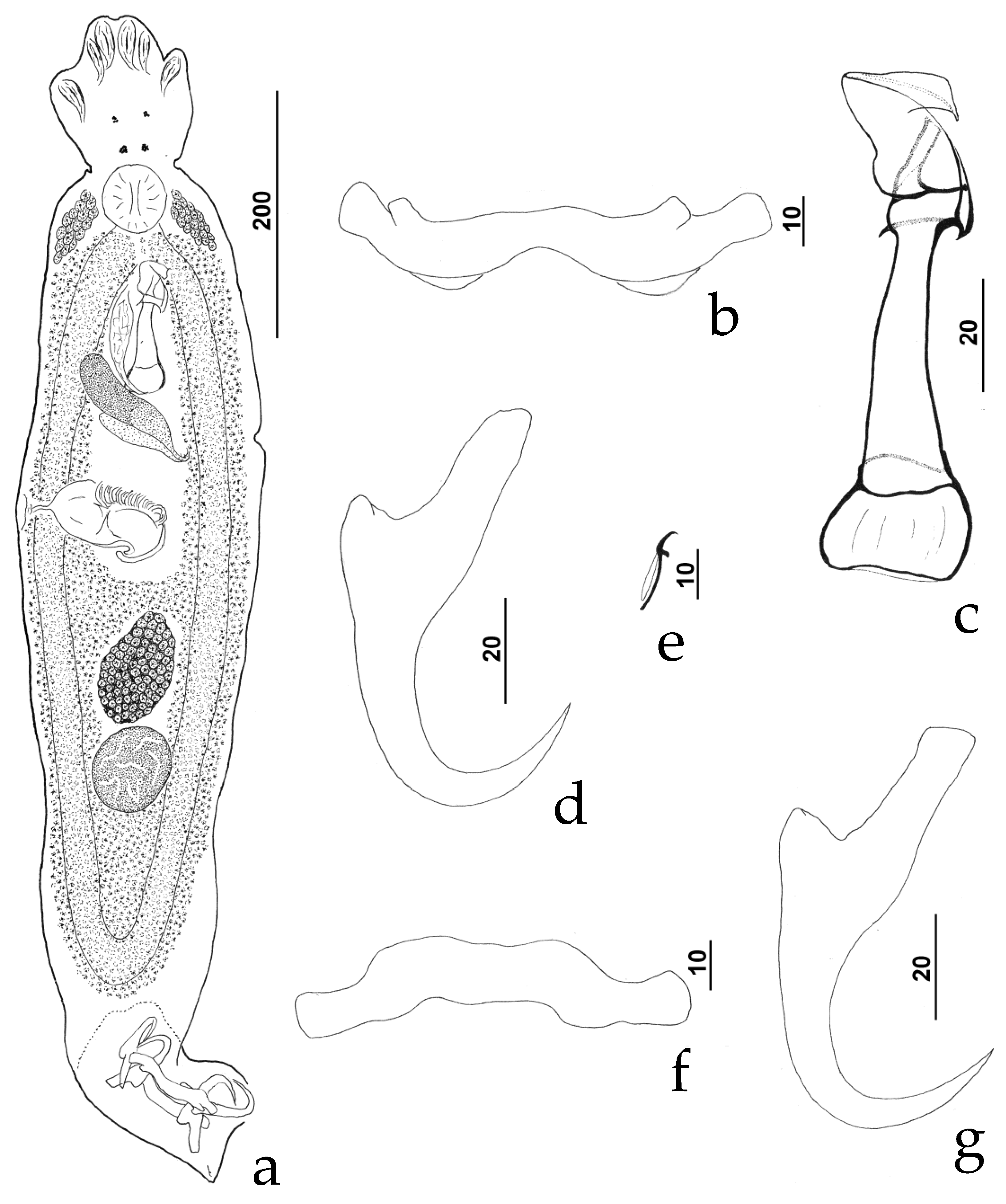
3.3.1. Description (Figure 2)
3.3.2. Measurements
3.3.3. Taxonomic Summary
3.3.4. Remarks
3.4. Platycephalotrema mastix n. sp.
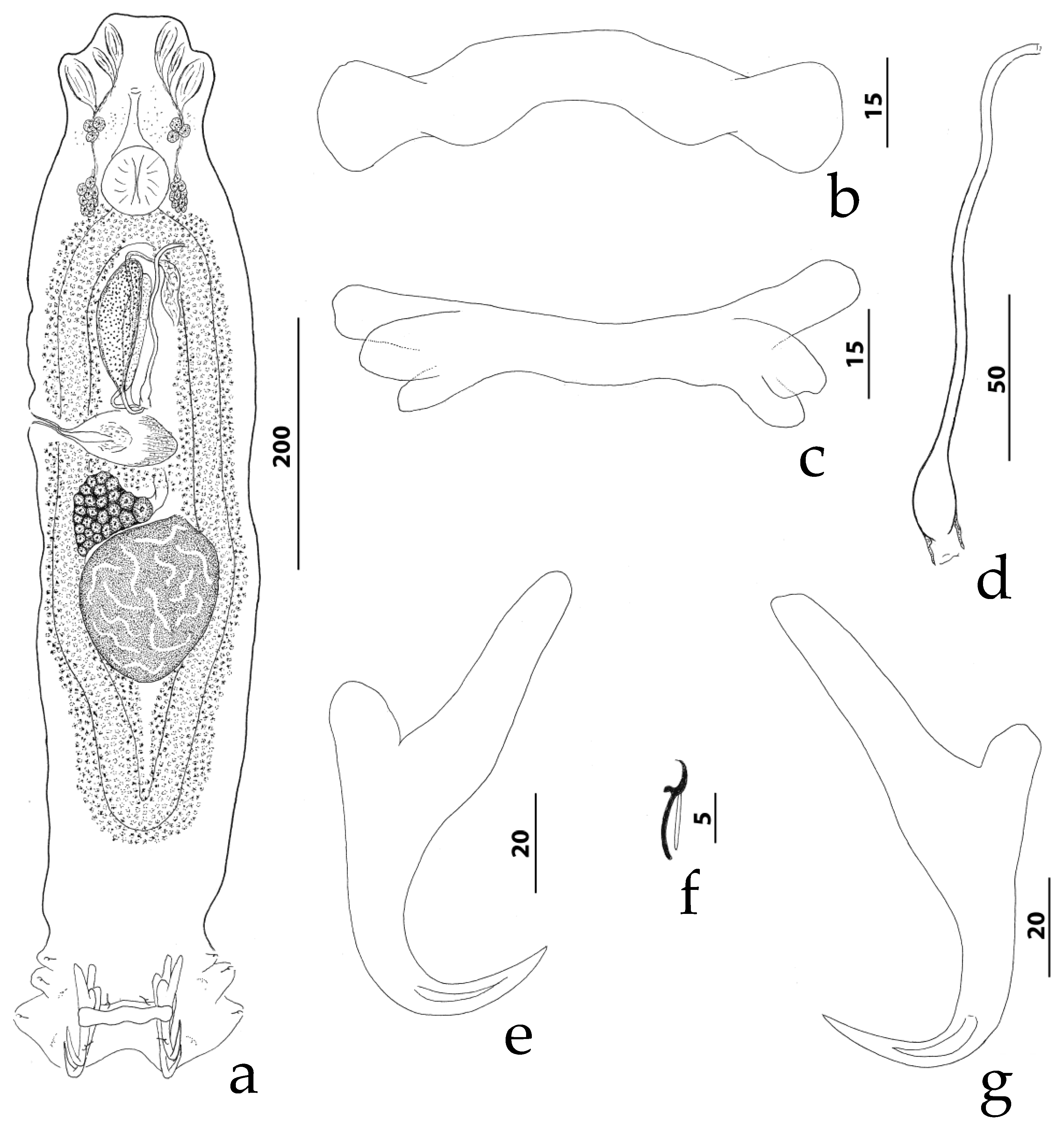
3.4.1. Description (Figure 3)
3.4.2. Measurements
3.4.3. Taxonomic Summary
3.4.4. Remarks
3.5. Platycephalotrema austrinum n. sp.
3.5.1. Description (Figure 4)
3.5.2. Measurements
3.5.3. Taxonomic Summary
3.5.4. Remarks
3.6. Platycephalotrema macassarensis (Yamaguti, 1963) n. comb.
3.6.1. Measurements
3.6.2. Taxonomic Summary
3.6.3. Remarks
3.7. Platycephalotrema thysanophrydis (Yamaguti, 1937) n. comb.
3.7.1. Redescription (Figure 6)
3.7.2. Measurements
3.7.3. Taxonomic Summary
3.7.4. Remarks
3.8. Platycephalotrema platycephali (Yin & Sproston, 1948) n. comb.
3.8.1. Taxonomic Summary
3.8.2. Remarks
3.9. Platycephalotrema sinensis (Yamaguti, 1963) n. comb.
3.9.1. Taxonomic summary
3.9.2. Remarks
3.10. Platycephalotrema bassensis (Hughes, 1928) n. comb.
3.10.1. Taxonomic Summary
3.10.2. Remarks
3.11. Dactylogyrids from Scorpaeniform Fishes that Are Potential Members of Platycephalotrema n. gen.
3.12. Ancyrocephalus vesiculosus Murray, 1931
3.12.1. Measurements
3.12.2. Taxonomic Summary
3.12.3. Remarks
3.13. Haliotrema indicum Tripathi, 1957
3.13.1. Taxonomic Summary
3.13.2. Remarks
3.14. Haliotrema swatowensis Yao, Wang, Xia, & Chen, 1998
3.14.1. Taxonomic Summary
3.14.2. Remarks
3.15. Haliotrema pteroisi Paperna, 1972
3.15.1. Taxonomic Summary
3.15.2. Remarks
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethical Approvals
References
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. (accessed on 15 February 2019).
- Puckridge, M.; Andreakis, N.; Appleyard, S.A.; Ward, R.D. Cryptic diversity in flathead fishes (Scorpaeniformes: Platycephalidae) across the Indo-West Pacific uncovered by DNA barcoding. Mole. Ecol. Res. 2013, 13, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Kritsky, D.C. Dactylogyrids (Monogenoidea) infecting the gill lamellae of some beloniform fishes from Moreton Bay, Queensland, Australia, with a redescription of Hareocephalus thaisae Young, 1969 and descriptions of six new species of Hemirhamphiculus Bychowsky & Nagibina, 1969. Syst. Parasitol. 2018, 95, 33–54. [Google Scholar] [PubMed]
- Kritsky, D.C. Species of Monogenoidea infecting the gill lamellae of the common silver−biddy Gerres oyena (Forsskål) and the common silver belly Gerres subfasciatus Cuvier (Perciformes: Gerreidae) in Moreton Bay, Queensland, Australia. Syst. Parasitol. 2018, 95, 499–525. [Google Scholar] [CrossRef] [PubMed]
- Kritsky, D.C. Euryhaliotrema spp. (Monogenoidea: Dactylogyridae) parasitic on the gill lamellae of perciform fishes in Moreton Bay, Queensland, Australia, with the description of Euryhaliotrema solenophallus sp. nov. from the silver moony Monodactylus argenteus (Linnaeus) (Monodactylidae). Acta Parasitol. 2019, 64, 223–227. [Google Scholar] [PubMed]
- Johnson, J.W. Fishes of the Moreton Bay Marine Park and adjacent continental shelf waters, Queensland, Australia. Mem. Queensland Mus. – Nature 2010, 54, 299–353. [Google Scholar]
- Nakabo, T.; Kai, Y. Platycephalidae. In Fishes of Japan with Pictorial Keys to the Species, 3rd ed.; Nakabo, T., Ed.; Tokai University Press: Hadano, Japan, 2013; pp. 734–740, 1953–1955. (In Japanese) [Google Scholar]
- Kritsky, D.C. Revision of Parancylodiscoides Caballero y C. & Bravo-Hollis, 1961 (Monogenoidea: Dactylogyridae), with a redescription of P. longiphallus (MacCallum, 1915) from the Atlantic spadefish Chaetodipterus faber (Broussonet) (Acanthuroidei: Ephippidae) in the Gulf of Mexico. Syst. Parasitol. 2012, 81, 97–108. [Google Scholar]
- Lim, L.H.S.; Justine, J.-L. Two new species of ancyrocephalid monogeneans from Lethrinus rubrioperculatus Sato (Perciformes: Lethrinidae) off New Caledonia, with the proposal of Lethrinitrema n. g. Syst. Parasitol. 2011, 78, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.-Y.; Sproston, N.G. Studies on the monogenetic trematodes of China: Part 4. Two Tetraonchinae from platycephalid fishes of the Chusan Region, Chekiang. Sinensia 1948, 19, 76–82. [Google Scholar]
- Bychowsky, B.E.; Nagibina, L.F. New and little known species of the genus Haliotrema Johnston et Tiegs, 1922 (Monogenoidea). 2. Zool. Zh. 1971, 50, 25–40. (In Russian) [Google Scholar]
- Young, P.C. The taxonomy of some dactylogyrid Monogenoidea from Australian fishes. Zool. Anz. 1968, 180, 269–279. [Google Scholar]
- Imamura, H. Taxonomic revision of the flathead fish genus Platycephalus Bloch, 1785 (Teleostei: Platycephalidae) from Australia, with description of a new species. Zootaxa 2015, 3904, 151–207. [Google Scholar] [CrossRef] [PubMed]
- Yamaguti, S. Parasitic worms mainly from Celebes. Part 2. Monogenetic trematodes of fishes. Acta Medicinae Okayama 1953, 8, 203–256 + plates I–IX. [Google Scholar]
- Zhang, J. Ancyrocephalidae Bychowsky & Nagibina, 1978. In Monogeneans of Chinese Marine Fishes; Zhang, J., Yang, T., Liu, L., Eds.; Agriculture Press: Beijing, China, 2001; pp. 79–183. (In Chinese) [Google Scholar]
- International Commission on Zoological Nomenclature. International Code of Zoological Nomenclature; The International Trust for Zoological Nomenclature: London, UK, 1999; p. 306. [Google Scholar]
- Yamaguti, S. Systema Helminthum, Volume IV. Monogenea and Aspidocotylea; Interscience Publishers: New York, NY, USA, 1963; p. 699. [Google Scholar]
- Yamaguti, S. Studies on the Helminth Fauna of Japan. Part. 19. Fourteen New Ectoparasitic Trematodes of Fishes; Kyoto, Japan, 1937; pp. 28 + plates i–vi. Available online: https://www.cabdirect.org/?target=%2fcabdirect%2fabstract%2f19370800159 (accessed on 15 June 2019).
- Ichthyological Society of Japan. Dictionary of Japanese Fish. Names and Their Foreign Equivalents; Sanseido: Tokyo, Japan, 1981; pp. 848 + ii + vii. (In Japanese) [Google Scholar]
- Imamura, H. A new species of the flathead genus Inegocia (Teleostei: Platycephalidae) from East Asia. Bull. Natl. Mus. Nat. Sci. Ser. A Suppl. 4 2010, 4, 21–29. [Google Scholar]
- Froese, R.; Pauly, D. (Eds.) Fishbase. Available online: www.fishbase.org (version 04/2019) (accessed on 15 June 2019).
- Wu, X.-Y.; Zhu, X.-Q.; Xie, M.-Q.; Li, A.-X. The radiation of Haliotrema (Monogenea: Dactylogyridae: Ancyrocephalinae): Molecular evidence and explanation inferred from LSU rDNA sequences. Parasitology 2006, 132, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.D. A comparative study of the biology of five South Queensland flatheads (Platycephalidae). B.S. Honours Thesis, University of Queensland, Brisbane, Australia, February 1971; p. 186. [Google Scholar]
- Boeger, W.A.; Kritsky, D.C. Phylogeny and a revised classification of the Monogenoidea Bychowsky, 1937 (Platyhelminthes). Syst. Parasitol. 1993, 26, 1–32. [Google Scholar] [CrossRef]
- Boeger, W.A.; Kritsky, D.C. Chapter 10. Phylogenetic relationships of the Monogenoidea. In Interrelationships of the Platyhelminthes; Littlewood, D.T.J., Bray, R.A., Eds.; Taylor & Francis: London, UK, 2001; pp. 92–102. [Google Scholar]
- Lebedev, B.I. About some parasitological institutions of Japan. Parazitologiya 1981, 15, 388–391. (In Russian) [Google Scholar]
- Oliver, G. Les Diplectanidae Bychowsky, 1957 (Monogenea, Monopisthocotylea, Dactylogyridea). Systématique. Biologie. Ontogénie. Ecologie. Essai de phylogenèse. Thèse d’état, Université des Sciences et Techniques du Languedoc, Montpellier, France, 1987; p. 433. [Google Scholar]
- Hughes, W.K. Some trematode parasites on the gills of Victorian fishes. Proc. Roy. Soc. Victoria 1928, 41, 45–54 + plates VIII–XI. [Google Scholar]
- Murray, F.V. Gill trematodes from some Australian fishes. Parasitology 1931, 23, 492–506 + plates XX–XXII. [Google Scholar] [CrossRef]
- Hooper, J.N.A. Parasites of estuarine and oceanic flathead fishes (Family Platycephalidae) from northern New South Wales. Aust. J. Zool. (Suppl. Ser.) 1983, 90, 1–69. [Google Scholar] [CrossRef]
- Imamura, H. Synonymy of two species of the genus Platycephalus and validity of Platycephalus westraliae (Teleostei: Platycephalidae). Ichthyol. Res. 2008, 55, 339–406. [Google Scholar] [CrossRef]
- Tripathi, Y.R. Monogenetic trematodes from fishes of India. Indian, J. Helminthol. 1957, 9, 1–149. [Google Scholar]
- Gupta, P.D.; Ghosh, R.K. Catalogue of type specimens present in the Zoological Survey of India. Part 3. Helminths. Rec. Zool. Survey India 1979, 74 (part 3), 243–331. [Google Scholar]
- Yang, T.; Kritsky, D.C.; Sun, Y.; Zhang, J.; Shi, S. Diplectanids infesting the gills of the barramundi Lates calcarifer (Bloch) (Perciformes: Centropomidae), with the proposal of Laticola n. g. (Monogenoidea: Diplectanidae). Syst. Parasitol. 2006, 63, 127–141. [Google Scholar]
- Yao, W.; Wang, W.; Xia, X.; Chen, W. Four new species of Ancyrocephalidae of marine fishes from Nanao Island, Guandong. Acta Hydrobiolog. Sin. 1998, 22 (Suppl.), 120–125, (In Chinese with English abstract). [Google Scholar]
- Paperna, I. Monogenea of Red Sea Fishes. III. Dactylogyridae from littoral and reef fishes. J. Helminthol. 1972, 46, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Colorni, A.; Diamant, A. Hyperparasitism of trichodinid ciliates on monogenean gill flukes of two marine fish. Dis. Aq. Organ. 2005, 65, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, L.J.; Sikkel, P.C.; Cure, K.; Hixon, M.A. Parasite-mediated enemy release and low biotic resistance may facilitate invasion of Atlantic coral reefs by Pacific red lionfish (Pterois volitans). Biol. Invasions 2017, 19, 563–575. [Google Scholar] [CrossRef]
- Gussev, A.V. [Monogenetic trematodes of fishes of the Amur basin.]. Tr. Zool. Inst. AN SSSR 1955, 19, 171–398. (In Russian) [Google Scholar]
- Bychowsky, B.E.; Nagibina, L.F. [Contributions to the revision of the genus Ancyrocephalus Creplin, 1839 (Dactylogyridae, Ancyrocephalinae).]. Parazitologiya 1970, 4, 193–200. [Google Scholar]
- Kritsky, D.C.; Stephens, F. Haliotrema abaddon n. sp. (Monogenoidea: Dactylogyridae) from the gills of wild and maricultured West Australian dhufish Glaucosoma hebraicum (Teleostei: Glaucosomatidae), in Australia. J. Parasitol. 2001, 87, 749–754. [Google Scholar]
- Klassen, G.J. Phylogeny of Haliotrema species (Monogenea: Ancyrocephalidae) from boxfishes (Tetraodontiformes: Ostraciidae): Are Haliotrema species from boxfishes monophyletic? J. Parasitol. 1994, 80, 596–610. [Google Scholar] [CrossRef] [PubMed]
- Euzet, L.; Suriano, D.M. Ligophorus n. g. (Monogenea, Ancyrocephalidae) parasite des Mugilidae (Téléostéens) en Méditerranée. Bull. Mus. Nat. d’Hist. Naturelle 1977, 329, 799–822. [Google Scholar]
- Kritsky, D.C.; Boeger, W.A. Neotropical Monogenoidea. 41: New and previously described species of Dactylogyridae (Platyhelminthes) from the gills of marine and freshwater perciform fishes (Teleostei) with proposal of a new genus and a hypothesis on phylogeny. Zoosystema 2002, 24, 7–40. [Google Scholar]
- Plaisance, L.; Kritsky, D.C. Dactylogyrids (Platyhelminthes: Monogenoidea) parasitizing butterfly fishes (Teleostei: Chaetodontidae) from the coral reefs of Palau, Moorea, Wallis, New Caledonia, and Australia: Species of Euryhaliotrematoides n. gen. and Aliatrema n. gen. J. Parasitol. 2004, 90, 328–341. [Google Scholar] [PubMed]
- Kritsky, D.C.; Yang, T.; Sun, Y. Dactylogyrids (Monogenoidea, Polyonchoinea) parasitizing the gills of snappers (Perciformes, Lutjanidae): Proposal of Haliotrematoides n. gen. and descriptions of new and previously described species from marine fishes of the Red Sea, the eastern and Indo-west Pacific Ocean, Gulf of Mexico and Caribbean Sea. Zootaxa 2009, 1970, 1–51. [Google Scholar]
- Kritsky, D.C. Dactylogyrids (Monogenoidea: Polyonchoinea) parasitizing the gills of snappers (Perciformes: Lutjanidae): Revision of Euryhaliotrema with new and previously described species from the Red Sea, Persian Gulf, the eastern and Indo-west Pacific Ocean, and the Gulf of Mexico. Zoologia 2012, 29, 227–276. [Google Scholar]
- Řehulková, E.; Justine, J.-L.; Gelnar, M. Five new monogenean species from the gills of Mulloidichthys vanicolensis (Perciformes: Mullidae) off New Caledonia, with the proposal of Volsellituba n. g. and Pennulituba n. g. (Monogenea: Dactylogyridae). Syst. Parasitol. 2010, 75, 125–145. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kritsky, D.C.; Nitta, M. Dactylogyrids (Platyhelminthes: Monogenoidea) Infecting the Gill Lamellae of Flatheads (Scorpaeniformes: Platycephalidae), with Proposal of Platycephalotrema n. gen. and Descriptions of New Species from Australia and Japan. Diversity 2019, 11, 132. https://doi.org/10.3390/d11080132
Kritsky DC, Nitta M. Dactylogyrids (Platyhelminthes: Monogenoidea) Infecting the Gill Lamellae of Flatheads (Scorpaeniformes: Platycephalidae), with Proposal of Platycephalotrema n. gen. and Descriptions of New Species from Australia and Japan. Diversity. 2019; 11(8):132. https://doi.org/10.3390/d11080132
Chicago/Turabian StyleKritsky, Delane C., and Masato Nitta. 2019. "Dactylogyrids (Platyhelminthes: Monogenoidea) Infecting the Gill Lamellae of Flatheads (Scorpaeniformes: Platycephalidae), with Proposal of Platycephalotrema n. gen. and Descriptions of New Species from Australia and Japan" Diversity 11, no. 8: 132. https://doi.org/10.3390/d11080132