The Effects of Edaphic and Climatic Factors on Secondary Lichen Chemistry: A Case Study Using Saxicolous Lichens


Abstract
1. Introduction
2. Materials and Methods
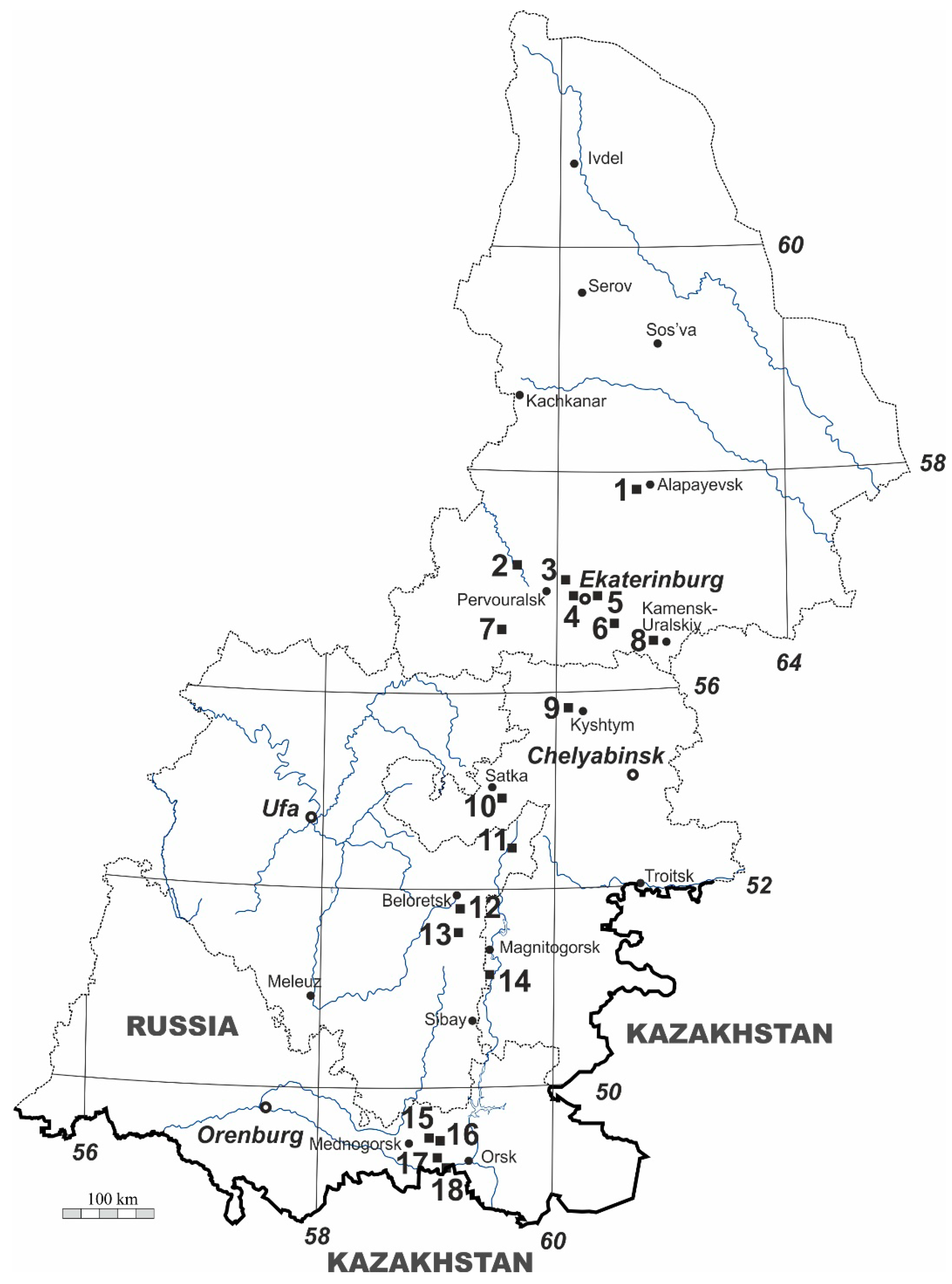
2.1. Study Area
2.2. Field Survey
2.3. Analytical Procedures
2.4. Statistical Analyses
3. Results
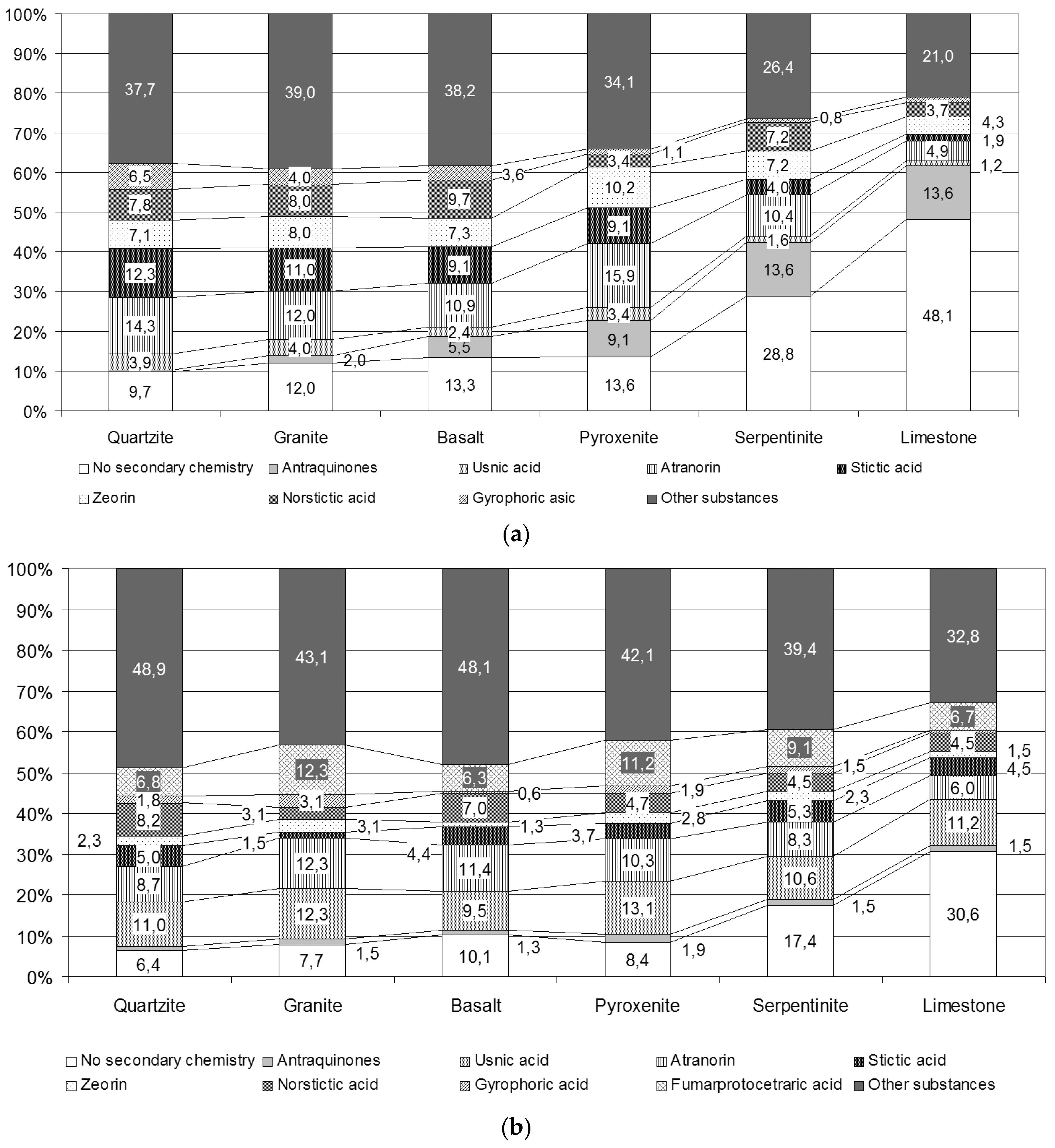
3.1. Species and Secondary Metabolite Diversity
3.2. Chemical Properties of Rock Substrates
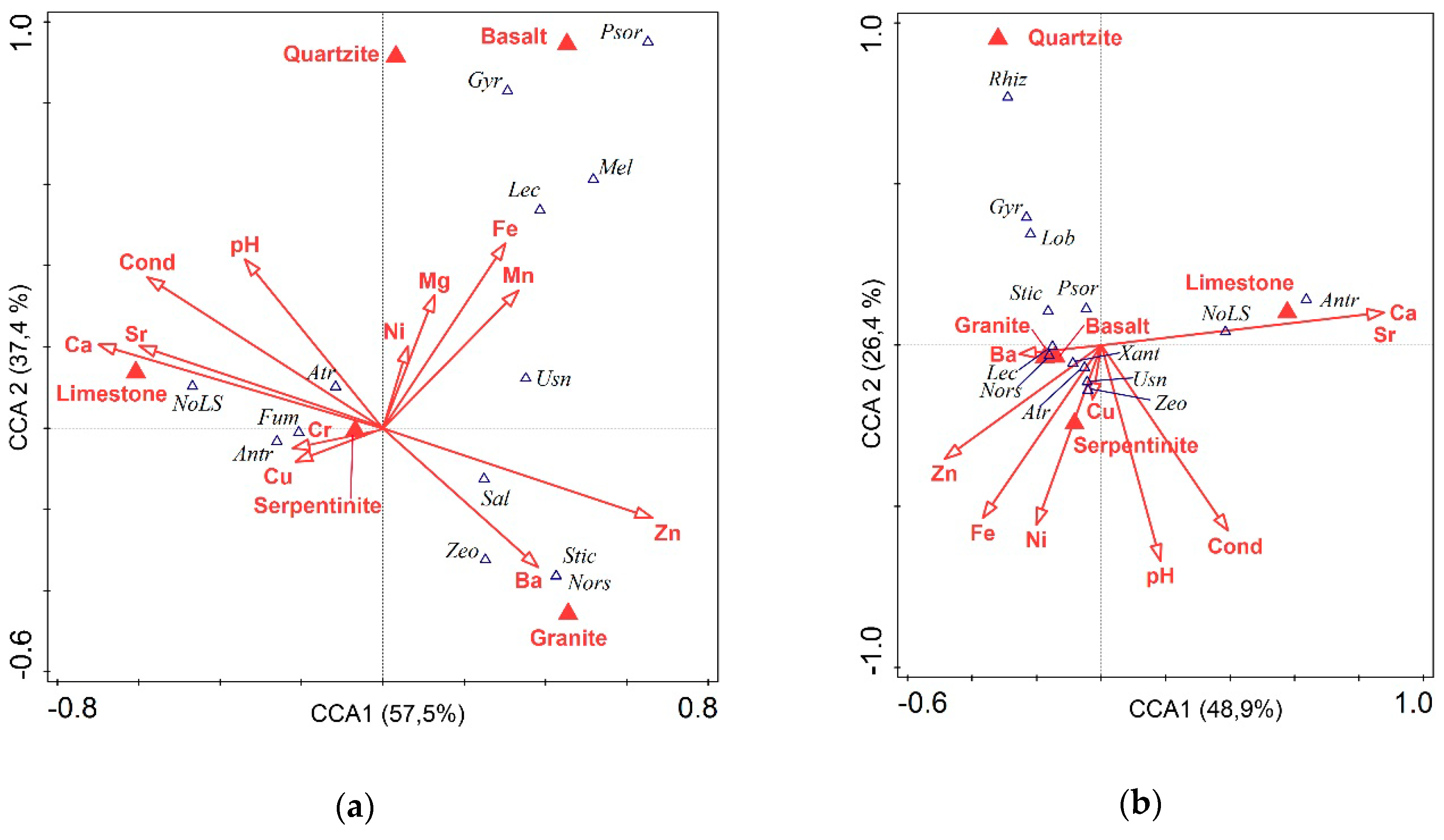
3.3. Secondary Metabolites and Chemistry of Rocks
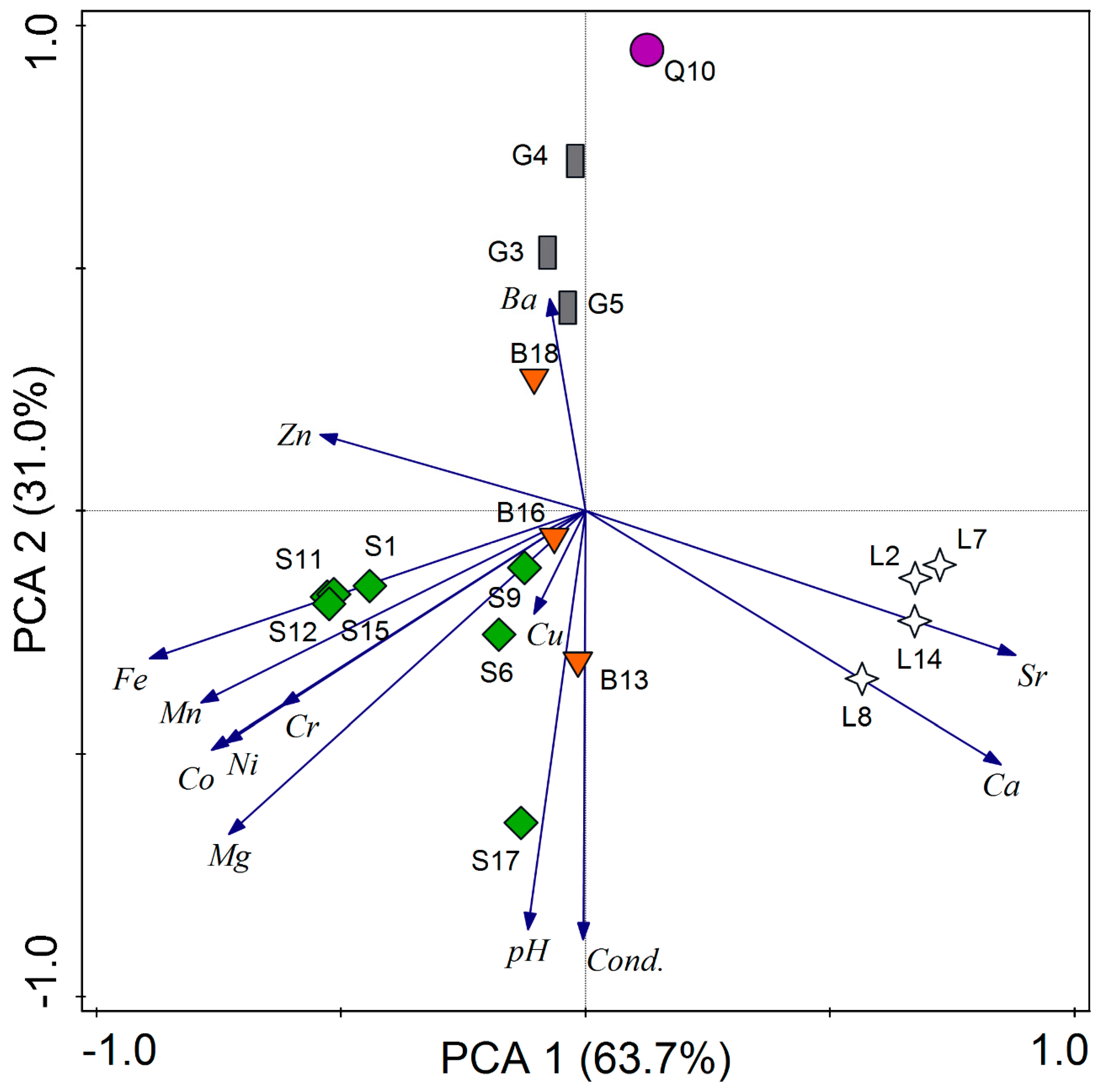
3.4. Secondary Metabolites and Environmental Parameters
4. Discussion
4.1. Secondary Metabolite Diversity on Different Rocks
4.2. Chemical Properties of Rock Substrates
4.3. Secondary Metabolites and Chemistry of Rocks
4.4. Secondary Metabolites and Climatic Factors
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huneck, S.; Yoshimura, I. Identification of Lichen Substances; Springer: Berlin/Heidelberg, Germany, 1996; pp. 1–493. [Google Scholar]
- Elix, J.A. A Catalogue of Standardized Chromatographic Data and Biosynthetic Relationships for Lichen Substances, 3rd ed.; Published by the author: Canberra, Australia, 2014; pp. 1–323. [Google Scholar]
- Rundel, P.W. The ecological role of secondary lichen substances. Biochem. Syst. Ecol. 1978, 6, 157–170. [Google Scholar] [CrossRef]
- Favero-Longo, S.E.; Piervittori, R. Lichen-plant interactions. J. Plant. Interact. 2010, 5, 163–177. [Google Scholar] [CrossRef]
- Lawrey, J.D. Lichen secondary compounds: Evidence for a correspondence between antiherbivore and antimicrobial function. Bryologist 1989, 92, 326–328. [Google Scholar] [CrossRef]
- Gauslaa, Y. Lichen palatability depends on investments in herbivore defence. Oecologia 2005, 143, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Lawrey, J.D. Biological role of lichen substances. Bryologist 1986, 89, 111–122. [Google Scholar] [CrossRef]
- Vainshtein, E.A. The role of lichen substances in symbiosis. Bot. Zhurn. 1991, 76, 793–800. [Google Scholar]
- Solhaug, K.A.; Gauslaa, Y. Parietin, a photoprotective secondary product of the lichen Xanthoria parietina. Oecologia 1996, 108, 412–418. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Solhaug, K.A. Fungal melanins as a sun screen for symbiotic green algae in the lichen Lobaria pulmonaria. Oecologia 2001, 126, 462–471. [Google Scholar] [CrossRef]
- Prokopiev, I.A.; Poryadina, L.N.; Konoreva, L.A.; Chesnokov, S.V.; Shavarda, A.L. Variation in the composition of secondary metabolites in Flavocetraria lichens from Western Siberia. Russ. J. Ecol. 2018, 49, 401–405. [Google Scholar] [CrossRef]
- Chowdhury, D.P.; Solhaug, K.A.; Gauslaa, Y. Ultraviolet radiation reduces lichen growth rates. Symbiosis 2017, 73, 27–34. [Google Scholar] [CrossRef]
- MacGillivray, T.; Helleur, R. Analysis of lichens under environmental stress using TMAH thermochemolysis-gas chromatography. J. Anal. Appl. Pyrolysis 2001, 58–59, 465–480. [Google Scholar] [CrossRef]
- Øvstedal, D.O.; Lewis Smith, R.I. Lichens of Antarctica and South. Georgia: A Guide to Their Identification and Ecology; Cambridge University Press: Cambridge, UK, 2001; pp. 1–411. [Google Scholar]
- Luo, H.; Yamamoto, Y.; Kim, J.A.; Jung, J.S.; Koh, Y.J.; Hur, J.-S. Lecanoric acid, a secondary lichen substance with antioxidant properties from Umbilicaria antarctica in maritime Antarctica (King George Island). Polar Biol. 2009, 32, 1033–1040. [Google Scholar] [CrossRef]
- Larsson, P.; Večeřová, K.; Cempírková, H.; Solhaug, K.A.; Gauslaa, Y. Does UV-B influence biomass growth in lichens deficient in sun-screening pigments? Environ. Exp. Bot. 2009, 67, 215–221. [Google Scholar] [CrossRef]
- Bjerke, J.W.; Joly, D.; Nilsen, L.; Brossard, T. Spatial trends in usnic acid concentrations of the lichen Flavocetraria nivalis along local climatic gradients in the Arctic (Kongsfjorden, Svalbard). Polar Biol. 2004, 27, 409–417. [Google Scholar] [CrossRef]
- Sheard, J.W. The comparative ecology and distribution and within-species variation of the lichenized Ascomycetes Ramalina cuspidata and R. siliquosa in the British Isles. Can. J. Bot. 1978, 56, 939–952. [Google Scholar] [CrossRef]
- LaGreca, S. A Phylogenetic evaluation of the Ramalina americana chemotype complex (Lichenized Ascomycota, Ramalinaceae) based on rDNA ITS sequence data. Bryologist 2006, 102, 602–618. [Google Scholar] [CrossRef]
- Culberson, C.F.; Culberson, W.L.; Arwood, D.A. Physiography and fumarprotocetraric acid production in the Cladonia chlorophaea group in North Carolina. Bryologist 1977, 80, 71–75. [Google Scholar] [CrossRef]
- Renner, B. The presence or absence of secondary metabolites in cephalodia and their possible implications. J. Hattori Bot. Lab. 1982, 52, 367–377. [Google Scholar]
- Renner, B.; Galloway, D.J. Phycosymbiodemes in Pseudocyphellaria in New Zealand. Mycotaxon 1982, 16, 197–231. [Google Scholar]
- Boyle, A.P.; McCarthy, P.M.; Stewart, D. Geochemical control of saxicolous lichen communities on the creggaun gabbro, Letterfrack, Co. Galway, Western Ireland. Lichenologist 1987, 19, 307–317. [Google Scholar] [CrossRef]
- Brodo, I.M. Substrate ecology. In The Lichens; Ahmadjian, V., Hale, M.E., Eds.; Academic Press: New York, NY, USA; London, UK, 1973; pp. 401–441. [Google Scholar]
- Favero-Longo, S.E.; Isocrono, D.; Piervittori, R. Lichens and ultramafic rocks: A review. Lichenologist 2004, 36, 391–404. [Google Scholar] [CrossRef]
- Favero-Longo, S.E.; Isocrono, D.; Piervittori, R. Measuring the biodiversity of saxicolous lichens above timberline with reference to environmental factors: The case of a Natura 2000 site of western Alps. Phytocoenologia 2009, 39, 51–78. [Google Scholar] [CrossRef]
- Favero-Longo, S.E.; Matteucci, E.; Morando, M.; Rolfo, F.; Harris, T.B.; Piervittori, R. Metals and secondary metabolites in saxicolous lichen communities on ultramafic and non-ultramafic rocks of the Western Italian Alps. Aust. J. Bot. 2015, 63, 276–291. [Google Scholar] [CrossRef]
- Purvis, O.W.; Halls, C. A review of lichens in metal-enriched environments. Lichenologist 1996, 28, 571–601. [Google Scholar] [CrossRef]
- Rajakaruna, N.; Knudsen, K.; Fryday, A.M.; O’Dell, R.E.; Pope, N.; Olday, F.C.; Woolhouse, S. Investigation of the importance of rock chemistry for saxicolous lichen communities of the New Idria serpentinite mass, San Benito County, California, USA. Lichenologist 2012, 44, 695–714. [Google Scholar] [CrossRef]
- Wirth, V. Die Silikatflechten-Gemeinschaften im ausseralpinen Zentraleuropa. Dissert. Bot. 1972, 17, 1–306. [Google Scholar]
- Hauck, M.; Jürgens, S.R. Usnic acid controls the acidity tolerance of lichens. Environ. Pollut. 2008, 156, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Paukov, A.G.; Teptina, A.Y.; Pushkarev, E.V. Heavy metal uptake by chemically distinct lichens from Aspicilia spp. growing on ultramafic rocks. Aust. J. Bot. 2015, 63, 111–118. [Google Scholar] [CrossRef]
- Ascaso, C.; Galvan, J.; Rodriguezpascual, C. The weathering of calcareous rocks by lichens. Pedobiologia 1982, 24, 219–229. [Google Scholar]
- Jones, D.; Wilson, M.J.; Tait, J.M. Weathering of a basalt by Pertusaria corallina. Lichenologist 1980, 12, 277–290. [Google Scholar] [CrossRef]
- Wilson, M.J.; Jones, D.; McHardy, W.J. The weathering of serpentinite by Lecanora atra. Lichenologist 1981, 13, 167–176. [Google Scholar] [CrossRef]
- Bačkor, M.; Fahselt, D. Using EDX-microanalysis and X-ray mapping to demonstrate metal uptake by lichens. Biologia (Bratislava) 2004, 59, 39–45. [Google Scholar]
- Favero-Longo, S.E. Lichens on metal-rich substrates. In Plant Ecology and Evolution in Harsh Environments; Rajakaruna, N., Boyd, R.S., Harris, T.B., Eds.; Nova Science Publishers: New York, NY, USA, 2014; pp. 53–76. [Google Scholar]
- Fortuna, L.; Baracchini, E.; Adami, G.; Tretiach, M. Melanization affects the content of selected elements in parmelioid lichens. J. Chem. Ecol. 2017, 43, 1086–1096. [Google Scholar] [CrossRef] [PubMed]
- Hauck, M.; Huneck, S.; Elix, J.A.; Paul, A. Does secondary chemistry enable lichens to grow on iron-rich substrates? Flora Morphol. Distrib. Funct. Ecol. Plants 2007, 202, 471–478. [Google Scholar] [CrossRef]
- Purvis, O.W.; Elix, J.A.; Broomhead, J.A.; Jones, G.C. The occurrence of copper norstictic acid in lichens from cupriferous substrata. Lichenologist 1987, 19, 193–203. [Google Scholar] [CrossRef]
- Purvis, O.W.; Elix, J.A.; Gaul, K.L. The occurrence of copper-psoromic acid in lichens from cupiferous substrata. Lichenologist 1990, 22, 345–354. [Google Scholar] [CrossRef]
- Hauck, M.; Willenbruch, K.; Leuschner, C. Lichen substances prevent lichens from nutrient deficiency. J. Chem. Ecol. 2009, 35, 71–73. [Google Scholar] [CrossRef]
- Hauck, M.; Jürgens, S.R.; Leuschner, C. Norstictic acid: Correlations between its physico-chemical characteristics and ecological preferences of lichens producing this depsidone. Environ. Exp. Bot. 2010, 68, 309–313. [Google Scholar] [CrossRef]
- Nakajima, H.; Fujimoto, N.; Yamamoto, Y.; Amemiya, T.; Itoh, K. Response of secondary metabolites to Cu in the Cu-hyperaccumulator lichen Stereocaulon japonicum. Environ. Sci. Pollut. Res. 2019, 26, 905–912. [Google Scholar] [CrossRef]
- Favero-Longo, S.E.; Matteucci, E.; Giordani, P.; Paukov, A.G.; Rajakaruna, N. Diversity and functional traits of lichens in ultramafic areas: A literature-based worldwide analysis integrated by field data at the regional scale. Ecol. Res. 2018, 33, 593–608. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Yemets, O.A.; Asplund, J.; Solhaug, K.A. Carbon based secondary compounds do not provide protection against heavy metal road pollutants in epiphytic macrolichens. Sci. Total Environ. 2016, 541, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Bačkorová, M.; Biľová, I.; Kimáková, T.; Bačkor, M. The presence of usnic acid does not protect the lichen Cladina arbuscula subsp. mitis against the long-term copper excess. S. Afr. J. Bot. 2015, 100, 94–100. [Google Scholar] [CrossRef]
- Ogureeva, G.N.; Leonova, N.B.; Buldakova, E.V.; Kadetov, N.G.; Arkhipova, M.V.; Miklyaeva, I.M.; Bocharnikov, M.V.; Dudov, S.V.; Ignatova, E.A.; Ignatov, M.S.; et al. The Biomes of Russia—Map; WWF-Russia Conservation Strategy, Faculty of Geography Lomonosov Moscow State University: Moscow, Russia, 2018. [Google Scholar]
- Pennino, M.G.; Paradinas, I.; Illian, J.B.; Muñoz, F.; Bellido, J.M.; López-Quílez, A.; Conesa, D. Accounting for preferential sampling in species distribution models. Ecol. Evol. 2019, 9, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Paukov, A.; Teptina, A. New records of lichens from Middle Urals, Russia. Folia Cryptog. Estonica 2012, 49, 39–43. [Google Scholar]
- Paukov, A.G.; Gagarina, L.V.; Frolov, I.V. New and interesting lichen records from the Ural Mountains, Russia. Folia Cryptog. Estonica 2017, 54, 25–30. [Google Scholar] [CrossRef][Green Version]
- Urbanavichene, I.; Urbanavichus, G.; Mežaka, A.; Palice, Z. New records of lichens and lichenicolous fungi from the Southern Ural Mountains, Russia. II. Folia Cryptog. Estonica 2013, 50, 73–80. [Google Scholar] [CrossRef]
- Nordin, A.; Moberg, R.; Tønsberg, T.; Vitikainen, O.; Dalsätt, Å.; Myrdal, M.; Snitting, D.; Ekman, S. Santesson’s Checklist of Fennoscandian Lichen-forming and Lichenicolous Fungi. Ver. 29 April 2011. Available online: http://130.238.83.220/santesson/home.php (accessed on 30 March 2019).
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, H.T.; Liu, J.K.; Maharachchikumbura, S.S.N.; Ekanayaka, A.H.; Tian, Q.; Phookamsak, R. Outline of Ascomycota: 2017. Fungal Divers. 2018. [Google Scholar] [CrossRef]
- Parker, K.C. Environmental relationships and vegetation associates of columnar cacti in the northern Sonoran Desert. Vegetatio 1988, 78, 125–140. [Google Scholar] [CrossRef]
- Zelený, D.; Chytrý, M. Environmental control of the vegetation pattern in deep river valleys of the Bohemian Massif. Preslia 2007, 79, 205–222. [Google Scholar]
- Diva-Gis. Available online: www.diva-gis.org (accessed on 7 January 2019).
- Culberson, C.F.; Kristinsson, H.D. A standardized method for the identification of lichen products. J. Chromatogr. 1970, 46, 85–93. [Google Scholar] [CrossRef]
- Orange, A.; James, P.W.; White, F.J. Microchemical Methods for the Identification of Lichens; British Lichen Society: London, UK, 2001; pp. 1–101. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeont. Electr. 2001, 4, 1–9. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5; Cambridge University Press: Padstow, UK, 2002; pp. 1–527. [Google Scholar]
- Purvis, O.W. Adaptation and interaction of saxicolous crustose lichens with metals. Bot. Stud. 2014, 55, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Goyal, R.; Seaward, M.R.D. Metal uptake in terricolous lichens. I. Metal localization within the thallus. New Phytol. 1981, 89, 631–645. [Google Scholar] [CrossRef]
- Goyal, R.; Seaward, M.R.D. Metal uptake in terricolous lichens. III. Translocation in the thallus of Peltigera canina. New Phytol. 1982, 90, 85–98. [Google Scholar] [CrossRef]
- Osyczka, P.; Rola, K.; Jankowska, K. Vertical concentration gradients of heavy metals in Cladonia lichens across different parts of thalli. Ecol. Indic. 2016, 61, 766–776. [Google Scholar] [CrossRef]
- Lucas, R.E.; Davis, J.F. Relationships between pH values of organic soils and availabilities of 12 plant nutrients. Soil Sci. 1961, 92, 177–182. [Google Scholar] [CrossRef]
- Teptina, A.; Paukov, A.; Rajakaruna, N. Ultramafic vegetation and soils in the circumboreal region of the Northern Hemisphere. Ecol. Res. 2018, 33, 609–628. [Google Scholar] [CrossRef]
- Paul, A.; Hauck, M.; Leuschner, C. Iron and phosphate uptake explains the calcifuge-calcicole behavior of the terricolous lichens Cladonia furcata subsp. furcata and C. rangiformis. Plant Soil 2009, 319, 49–56. [Google Scholar] [CrossRef]
- Kasama, T.; Murakami, T.; Ohnuki, T. Accumulation mechanisms of uranium, copper and iron by lichen Trapelia involuta. In Biomineralization (BIOM2001): Formation, Diversity, Evolution and Application, Proceedings of the 8th International Symposium on Biomineralization (September 25–28, 2001, Niigata, Japan), 2003; Kobayashi, I., Ozawa, H., Eds.; Tokai University Press: Kanagawa, Japan, 2003; pp. 298–301. [Google Scholar]
- Wedin, M.; Westberg, M.; Crewe, A.T.; Tehler, A. Species delimitation and evolution of metal bioaccumulation in the lichenized Acarospora smaragdula (Ascomycota, Fungi) complex. Cladistics 2009, 25, 161–172. [Google Scholar] [CrossRef]
- Asplund, J.; Siegenthaler, A.; Gauslaa, Y. Simulated global warming increases usnic acid but reduces perlatolic acid in the mat-forming terricolous lichen Cladonia stellaris. Lichenologist 2017, 49, 269–274. [Google Scholar] [CrossRef]
- Lutsak, T.; Fernández-Mendoza, F.; Nadyeina, O.; Şenkardeşler, A.; Printzen, C. Testing the correlation between norstictic acid content and species evolution in the Cetraria aculeata group in Europe. Lichenologist 2017, 49, 39–56. [Google Scholar] [CrossRef]
- Košuthová, A.; Svitková, I.; Pišút, I.; Senko, D.; Valachovič, M.; Zaniewski, P.T.; Hájek, M. Climatic gradients within temperate Europe and small-scale species composition of lichen-rich dry acidophilous Scots pine forests. Fungal Ecol. 2015, 14, 8–23. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
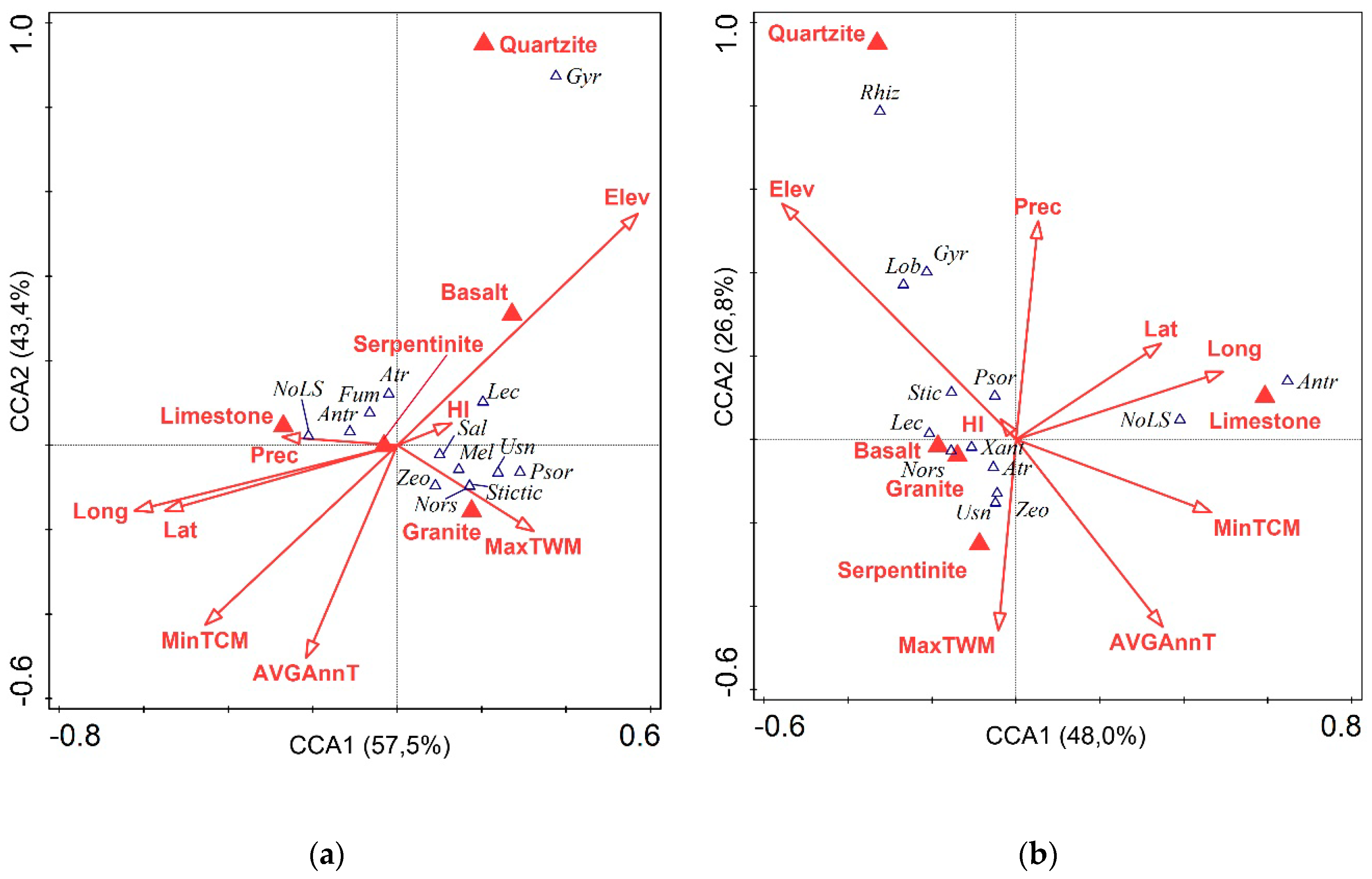{kind=link}
| N | Region | Locality | Latitude, N | Longitude, E | Elevation, m | Rock Type | Number of 1 × 1 m Quadrats | Number of 10 × 10 cm Squares |
|---|---|---|---|---|---|---|---|---|
| 1 | Sverdlovsk | Melkozyorovo village | 57°44′ | 61°28′ | 125 | Serpentinite | 10 | 100 |
| 2 | Sverdlovsk | Kourovka settlement | 57°02′ | 59°38′ | 280 | Limestone | 15 | 150 |
| 3 | Sverdlovsk | Petra Gronskogo rocks | 56°58′ | 60°18′ | 320 | Granite | 20 | 200 |
| 4 | Sverdlovsk | Ekaterinburg, Baran peninsula | 56°50′ | 60°30′ | 280 | Granite | 20 | 200 |
| 5 | Sverdlovsk | Ekaterinburg, Kamennye palatki | 56°50′ | 60°40′ | 280 | Granite | 10 | 100 |
| 6 | Sverdlovsk | Dvurechensk settlement | 56°35′ | 61°03′ | 205 | Serpentinite | 18 | 180 |
| 7 | Sverdlovsk | Bazhukovo settlement ‘Olenyi Ruchyi’ National park | 56°30′ | 59°15′ | 300 | Limestone | 20 | 200 |
| 8 | Sverdlovsk | Kodinka village | 56°25′ | 61°48′ | 120 | Limestone | 16 | 160 |
| 9 | Chelyabinsk | Egoza mountain | 55°45′ | 60°26′ | 570 | Serpentinite | 10 | 100 |
| 10 | Chelyabinsk | Zyuratkul’ mountain | 54°57′ | 59°10′ | 1170 | Quartzite | 20 | 200 |
| 11 | Bashkortostan | Kalkanovo village | 54°25′ | 59°20′ | 670 | Serpentinite | 12 | 120 |
| 12 | Bashkortostan | Shigayevo village | 53°48′ | 58°11′ | 560 | Serpentinite | 12 | 120 |
| 13 | Bashkortostan | Kushai mountain | 53°43′ | 58°37′ | 507-920 | Basalt | 15 | 150 |
| 14 | Chelyabinsk | Yangelskiy settlement | 53°13′ | 58°54′ | 350 | Limestone | 8 | 80 |
| 15 | Orenburg | Mazovo village | 51°30′ | 58°01′ | 340 | Serpentinite | 12 | 120 |
| 16 | Orenburg | Gainulino village | 51°23′ | 58°18′ | 270 | Basalt | 10 | 100 |
| 17 | Orenburg | Akkermanovka settlement | 51°08′ | 58°02′ | 300 | Serpentinite | 10 | 100 |
| 18 | Orenburg | Khabarnoye settlement | 51°06′ | 58°06′ | 220 | Basalt | 10 | 100 |
| Secondary Metabolite | Number of Species Containing the Metabolite | |||
|---|---|---|---|---|
| All Species | Crustose Species | Foliose Species | Fruticose Species | |
| Atranorin | 73 | 36 | 20 | 17 |
| Usnic acid | 42 | 10 | 14 | 18 |
| Gyrophoric acid | 41 | 19 | 22 | 0 |
| Zeorin | 37 | 19 | 15 | 3 |
| Norstictic acid | 36 | 30 | 4 | 2 |
| Anthraquinones | 35 | 32 | 3 | 0 |
| Stictic acid | 32 | 27 | 4 | 1 |
| Fumarprotocetraric acid | 21 | 1 | 3 | 17 |
| Lecanoric acid | 14 | 6 | 8 | 0 |
| Lobaric acid | 11 | 4 | 1 | 6 |
| Psoromic acid | 10 | 7 | 3 | 0 |
| Xanthones | 10 | 10 | 0 | 0 |
| Measured Feature | Quartzite | Granite | Basalt | Pyroxenite | Serpentinite | Limestone |
|---|---|---|---|---|---|---|
| Total number of species | 198 | 93 | 193 | 107 | 167 | 233 |
| Number of species within 10 × 10 cm plots | 3.19 ± 0.24 | 2.31 ± 0.14 | 4.41 ± 0.16 | n/a | 3.88 ± 0.18 | 2.81 ± 0.11 |
| Shannon H diversity index | 0.75 ± 0.07 | 0.45 ± 0.04 | 1.01 ± 0.03 | n/a | 0.81 ± 0.04 | 0.71 ± 0.03 |
| Number of secondary metabolites | 63 | 50 | 68 | 49 | 50 | 43 |
| Metabolite to species ratio | 0.32 | 0.54 | 0.35 | 0.46 | 0.30 | 0.18 |
| Average number of metabolites in one species | 1.72 ± 0.09A | 1.56 ± 0.11B | 1.44 ± 0.08B | 1.61 ± 0.11A,B | 1.14 ± 0.09C | 0.74 ± 0.07D |
| Rock/Locality | pH | Conductivity, S*10−3/m | Metals, µg/g (Average ± Standard Error) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ba | Ca | Co | Cr | Cu | Fe | Mg | Mn | Ni | Sr | Zn | |||
| S 1 | 7.36 ± 0.01 | 341 ± 31 | 5 ± 2 | 928 ± 151 | 68 ± 1 | 438 ± 19 | 30 ± 15 | 29,491 ± 14,966 | 124,271 ± 62,183 | 571 ± 4 | 1677 ± 290 | 2 ± 0.0 | 35 ± 6 |
| S 6 | 8.20 ± 0.06 | 294 ± 12 | 40 ± 25 | 9410 ± 5633 | 32 ± 6 | 770 ± 535 | 77 ± 34 | 25,835 ± 4813 | 58,592 ± 18,636 | 446 ± 92 | 526 ± 241 | 7 ± 2 | 31 ± 8 |
| S 9 | 7.88 ± 0.02 | 205 ± 1 | 6 ± 2 | 7544 ± 746 | 16 ± 6 | 413 ± 74 | 8 ± 2 | 13,266 ± 2818 | 43,449 ± 8601 | 219 ± 28 | 569 ± 143 | 1 ± 0.0 | 24 ± 3 |
| S 11 | 8.18 ± 0.04 | 342 ± 43 | 8 ± 3 | 376 ± 62 | 79 ± 8 | 256 ± 31 | 5 ± 0.0 | 39,370 ± 530 | 183,899 ± 6422 | 585 ± 35 | 1550 ± 76 | 2 ± 0.0 | 40 ± 3 |
| S 12 | 8.08 ± 0.02 | 239 ± 9 | 5 ± 0.0 | 613 ± 23 | 70 ± 5 | 576 ± 126 | 3 ± 0.0 | 41,284 ± 1964 | 181,878 ± 13,159 | 567 ± 18 | 1553 ± 95 | 1 ± 0.0 | 17 ± 0.0 |
| S 15 | 8.18 ± 0.08 | 289 ± 12 | 9 ± 3 | 529 ± 89 | 71 ± 7 | 281 ± 52 | 4 ± 2 | 39,499 ± 1707 | 200,647 ± 11,743 | 482 ± 49 | 1768 ± 309 | 3 ± 1 | 17 ± 2 |
| S 17 | 8.08 ± 0.08 | 324 ± 21 | 7 ± 3 | 42,726 ± 28,779 | 61 ± 14 | 184 ± 26 | 3 ± 1 | 37,779 ± 5306 | 166,937 ± 7170 | 534 ± 135 | 1564 ± 438 | 23 ± 10 | 16 ± 3 |
| L 2 | 7.86 ± 0.02 | 318 ± 43 | 11 ± 3 | 305,943 ± 10,157 | nd | 1 ± 1 | 12±1 | 290 ± 169 | 2223 ± 167 | 48 ± 18 | 2 ± 2 | 194 ± 46 | 9 ± 4 |
| L 7 | 8.00 ± 0.01 | 247 ± 44 | 3 ± 2 | 375,458 ± 6176 | nd | nd | 8 ± 0.0 | 74 ± 13 | 1716 ± 164 | 28 ± 10 | 1 ± 0.0 | 128 ± 9 | 2 ± 0.0 |
| L 8 | 7.18 ± 0.04 | 278 ± 29 | 10 ± 3 | 323,865 ± 7008 | nd | 7 ± 1 | 10 ± 0.0 | 347 ± 28 | 13,112 ± 5037 | 52 ± 18 | 4 ± 2 | 136 ± 12 | 5 ± 1 |
| L 14 | 7.88 ± 0.18 | 298 ± 52 | 8 ± 2 | 384,928 ± 17,198 | nd | 7 ± 2 | 7 ± 0.0 | 128 ± 54 | 4263 ± 855 | 61 ± 14 | nd | 118 ± 38 | 6 ± 1 |
| G 3 | 6.48 ± 0.21 | 47 ± 6 | 65 ± 34 | 1076 ± 452 | 4 ± 1 | 9 ± 3 | 14 ± 4 | 7289 ± 2170 | 5090 ± 1884 | 186 ± 13 | 32 ± 23 | 12 ± 7 | 41 ± 8 |
| G 4 | 6.68 ± 0.15 | 35 ± 9 | 38 ± 4 | 640 ± 157 | nd | 2 ± 0.0 | 9 ± 2 | 5277 ± 719 | 1688 ± 491 | 223 ± 49 | 2 ± 0.0 | 5 ± 2 | 44±6 |
| G 5 | 6.64 ± 0.08 | 206 ± 24 | 257 ± 89 | 1558 ± 55 | nd | 6 ± 1 | 16 ± 2 | 7620 ± 1225 | 3555 ± 554 | 199 ± 30 | 6 ± 0.0 | 10 ± 1 | 36 ± 5 |
| B 13 | 7.74 ± 0.04 | 206 ± 38 | 47 ± 13 | 34,996 ± 7749 | 22 ± 1 | 33 ± 5 | 57 ± 4 | 46,306 ± 372 | 26,663 ± 2538 | 767 ± 55 | 33 ± 8 | 32 ± 7 | 58 ± 3 |
| B 16 | 7.82 ± 0.03 | 352 ± 41 | 19 ± 2 | 7881 ± 414 | 14 ± 2 | 23 ± 8 | 8±3 | 15,687 ± 2573 | 19,252 ± 3465 | 228 ± 31 | 95 ± 37 | 7 ± 0.0 | 22 ± 4 |
| B 18 | 7.35 ± 0.15 | 87 ± 19 | 171 ± 48 | 3387 ± 924 | 2 ± 0.0 | 9 ± 8 | 13±4 | 24,667 ± 1895 | 6920 ± 471 | 480 ± 124 | 6 ± 3 | 15 ± 9 | 36 ± 9 |
| Q 10 | 6.16 ± 0.06 | 39 ± 9 | 28 ± 8 | 183 ± 19 | nd | 1 ± 0.0 | 1 ± 0.0 | 244 ± 93 | 316 ± 199 | 6±1 | 3 ± 2 | 2 ± 0.0 | 3 ± 1 |
| Edaphic Feature | Conductivity | Ba | Ca | Co | Cr | Cu | Fe | Mg | Mn | Ni | Sr | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | 0.67 | −0.41 | 0.13 | 0.57 | 0.38 | 0.12 | 0.53 | 0.58 | 0.45 | 0.52 | 0.10 | −0.11 |
| Conductivity | −0.31 | 0.24 | 0.38 | 0.30 | 0.08 | 0.16 | 0.35 | 0.16 | 0.44 | 0.25 | −0.23 | |
| Ba | −0.21 | −0.34 | −0.24 | 0.09 | −0.12 | −0.33 | −0.03 | −0.33 | −0.13 | 0.37 | ||
| Ca | −0.39 | −0.26 | −0.10 | −0.46 | −0.34 | −0.48 | −0.35 | 0.91 | −0.55 | |||
| Co | 0.40 | −0.02 | 0.83 | 0.93 | 0.74 | 0.85 | −0.40 | 0.18 | ||||
| Cr | 0.48 | 0.22 | 0.39 | 0.19 | 0.51 | −0.27 | −0.03 | |||||
| Cu | 0.18 | −0.11 | 0.24 | −0.13 | −0.06 | 0.42 | ||||||
| Fe | 0.81 | 0.90 | 0.58 | −0.43 | 0.45 | |||||||
| Mg | 0.66 | 0.87 | −0.36 | 0.04 | ||||||||
| Mn | 0.47 | −0.43 | 0.60 | |||||||||
| Ni | −0.36 | −0.05 | ||||||||||
| Sr | −0.42 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paukov, A.; Teptina, A.; Morozova, M.; Kruglova, E.; Favero-Longo, S.E.; Bishop, C.; Rajakaruna, N. The Effects of Edaphic and Climatic Factors on Secondary Lichen Chemistry: A Case Study Using Saxicolous Lichens. Diversity 2019, 11, 94. https://doi.org/10.3390/d11060094
Paukov A, Teptina A, Morozova M, Kruglova E, Favero-Longo SE, Bishop C, Rajakaruna N. The Effects of Edaphic and Climatic Factors on Secondary Lichen Chemistry: A Case Study Using Saxicolous Lichens. Diversity. 2019; 11(6):94. https://doi.org/10.3390/d11060094
Chicago/Turabian StylePaukov, Alexander, Anzhelika Teptina, Maria Morozova, Ekaterina Kruglova, Sergio E. Favero-Longo, Cora Bishop, and Nishanta Rajakaruna. 2019. "The Effects of Edaphic and Climatic Factors on Secondary Lichen Chemistry: A Case Study Using Saxicolous Lichens" Diversity 11, no. 6: 94. https://doi.org/10.3390/d11060094
APA StylePaukov, A., Teptina, A., Morozova, M., Kruglova, E., Favero-Longo, S. E., Bishop, C., & Rajakaruna, N. (2019). The Effects of Edaphic and Climatic Factors on Secondary Lichen Chemistry: A Case Study Using Saxicolous Lichens. Diversity, 11(6), 94. https://doi.org/10.3390/d11060094