The Long and Short of Biodiversity: Cumulative Diversity and Its Drivers


Abstract
:1. Introduction
- (1)
- What factors support cumulative diversity?
- (2)
- Do richness and evenness components contribute differently to cumulative diversity in the face of environmental change?
- (3)
- How do multiple paths of causation combine to determine cumulative diversity?
2. Materials and Methods
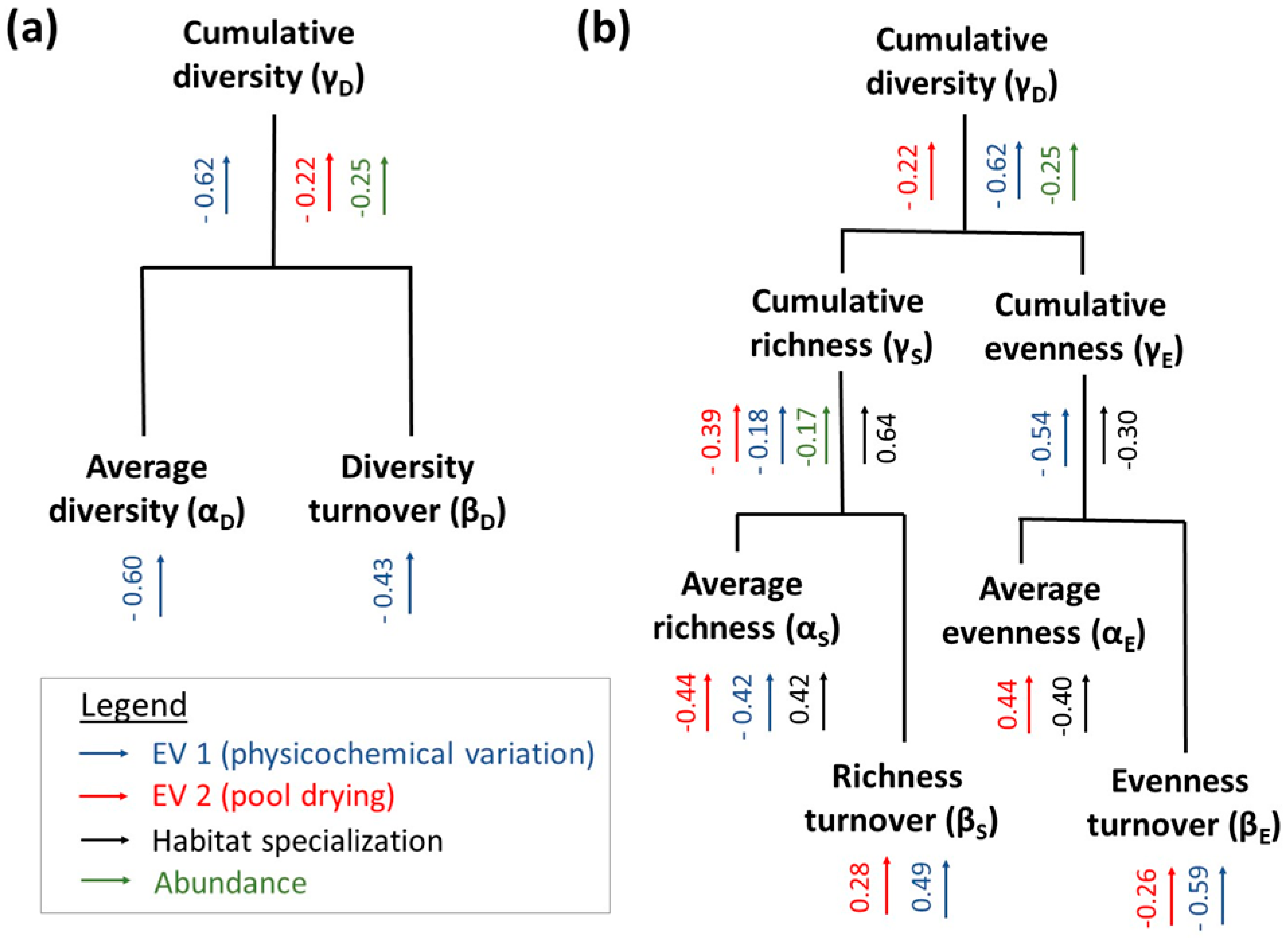
2.1. Hierarchical Partitioning of Cumulative Diversity
2.2. Rock Pool System and Community Sampling
2.3. Environmental and Community Factors
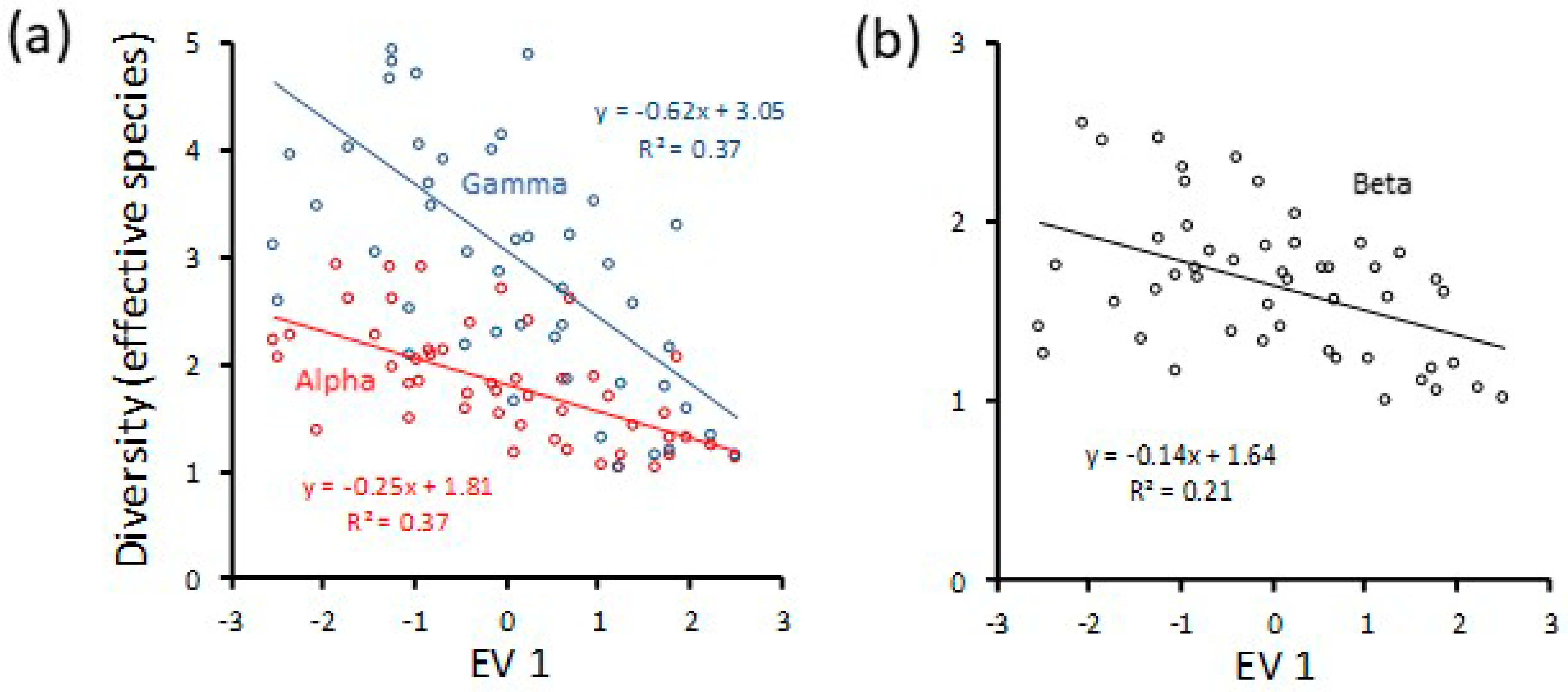
3. Results
4. Discussion
4.1. Environmental Control of Cumulative Diversity
4.2. Differential Pathways to Cumulative Diversity
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moreno, C.E.; Halffter, G. Spatial and temporal analysis of α, β and γ diversities of bats in a fragmented landscape. Biodiversity 2001, 10, 367–382. [Google Scholar]
- Magurran, A.E. Species abundance distributions over time. Ecol. Lett. 2007, 10, 347–354. [Google Scholar] [CrossRef]
- Johnson, C.N.; Balmford, A.; Brook, B.W.; Buettel, J.C.; Galetti, M.; Guangchun, L.; Wilmshurst, J.M. Biodiversity losses and conservation responses in the Anthropocene. Science 2017, 275, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Agoglitta, R.; Moreno, C.; Zunino, M.; Bonsignori, G.; Dellacasa, M. Cumulative annual dung beetle diversity in Mediterranean seasonal environments. Ecol. Res. 2012, 27, 387–395. [Google Scholar] [CrossRef]
- Werner, E.E.; Yurewicz, K.L.; Skelly, D.K.; Relyea, R.A. Turnover in an amphibian metacommunity: The role of local and regional factors. Oikos 2007, 116, 1713–1725. [Google Scholar] [CrossRef]
- Grman, E.; Suding, K.N. Within-year soil legacies contribute to strong priority effects of exotics on native California grassland communities. Restor. Ecol. 2010, 18, 664–670. [Google Scholar] [CrossRef]
- Cuddington, K. Legacy effects: The persistent impact of ecological interactions. Biol. Theory 2011, 6, 203–210. [Google Scholar] [CrossRef]
- Pendergast, T.; Hanlon, S.M.; Long, Z.M.; Royo, A.A.; Carson, W.P. The legacy of deer overabundance: Long-term delays in herbaceous understory recovery. Can. J. For. Res. 2016, 369, 362–369. [Google Scholar] [CrossRef]
- Yang, L.H.; Gratton, C. Insects as drivers of ecosystem processes. Curr. Opin. Insect Sci. 2014, 2, 26–32. [Google Scholar] [CrossRef]
- Mouillot, D.; Bellwood, D.R.; Baraloto, C.; Chave, J.; Galzin, R.; Harmelin-Vivien, M.; Kulbicki, M.; Lavergne, S.; Lavorel, S.; Mouquet, N.; et al. Rare species support vulnerable functions in high-diversity ecosystems. PLoS Biol. 2013, 11, e1001569. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hughes, T.P.; Hoey, A.S. Report sleeping functional group drives coral-reef recovery. Curr. Biol. 2006, 16, 2434–2439. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, W. Long-term relationships between ecological stability and biodiversity in Phanerozoic reefs. Nature 2005, 433, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Thibault, K.; White, E.P.; Ernest, S.K.M. Temporal dynamics in the structure and composition of a desert rodent community. Ecology 2004, 85, 2649–2655. [Google Scholar] [CrossRef]
- Magurran, A.E.; Henderson, P.A. Temporal turnover and the maintenance of diversity in ecological assemblages. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3611–3620. [Google Scholar] [CrossRef] [Green Version]
- Adler, P.B.; Lauenroth, W.K. The power of time: Spatiotemporal scaling of species diversity. Ecol. Lett. 2003, 6, 749–756. [Google Scholar] [CrossRef]
- White, E.P.; Adler, P.B.; Lauenroth, W.K.; Gill, R.A.; Greenberg, D.; Kaufman, D.M.; Rassweiler, A.; Rusak, J.A.; Smith, M.D.; Steinbeck, J.R.; et al. A comparison of the species time relationship across ecosystems and taxonomic groups. Oikos 2006, 112, 185–195. [Google Scholar] [CrossRef]
- Preston, F.W. Time and space and the variation of species. Ecology 1960, 41, 612–627. [Google Scholar] [CrossRef]
- Dolan, J.R.; Ritchie, M.E.; Tunin-ley, A. Dynamics of core and occasional species in the marine plankton: Rintinnid ciliates in the north-west Mediterranean Sea. J. Biogeogr. 2009, 36, 887–895. [Google Scholar] [CrossRef]
- Hansen, G.J.A.; Carey, C.C. Fish and phytoplankton exhibit contrasting temporal species abundance patterns in a dynamic north temperate lake. PLoS ONE 2015, 10, 1–19. [Google Scholar] [CrossRef]
- Gascón, S.; Arranz, I.; Argüelles, M.C.; Nebra, A.; Ruhí, A.; Rieradevall, M.; Caiola, N.; Sala, J.; Ibàñez, C.; Quintana, X.D.; et al. Environmental filtering determines metacommunity structure in wetland microcrustaceans. Oecologia 2016, 181, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Tredennick, A.T.; Adler, P.B.; Adler, F. The relationship between species richness and ecosystem variability is shaped by the mechanism of coexistence. Ecol. Lett. 2017, 20, 958–968. [Google Scholar] [CrossRef] [PubMed]
- Kolasa, J.; Li, B. Removing the confounding effect of habitat specialization reveals the stabilizing contribution of diversity to species. Biol. Lett. 2003, 201, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Metz, M.R. Does habitat specialization by seedlings contribute to the high diversity of a lowland rain forest? J. Ecol. 2012, 100, 969–979. [Google Scholar] [CrossRef] [Green Version]
- Albert, J.; Carvalho, T.; Petry, P.; Holder, M.; Maxime, E.; Espino, J.; Corahua, I.; Quispe, R.; Rengifo, B.; Ortega, H.; et al. Aquatic biodiversity in the Amazon: Habitat specialization and geographic isolation promote species richness. Animals 2011, 1, 205–241. [Google Scholar] [CrossRef] [PubMed]
- Tuomisto, H. A consistent terminology for quantifying species diversity? Yes, it does exist. Oecologia 2010, 164, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Tuomisto, H. An updated consumer’s guide to evenness and related indices. Oikos 2012, 121, 1203–1218. [Google Scholar] [CrossRef]
- Sciullo, L.; Kolasa, J. Linking local community structure to the dispersal of aquatic invertebrate species in a rock pool metacommunity. Community Ecol. 2012, 13, 203–212. [Google Scholar] [CrossRef]
- Pandit, S.N.; Kolasa, J. Opposite effects of environmental variability and species richness on temporal turnover of species in a complex habitat mosaic. Hydrobiologia 2012, 685, 145–154. [Google Scholar] [CrossRef]
- Pandit, S.; Kolasa, J.; Cottenie, K. Contrasts between habitat generalists and specialists: An empirical extension to the basic metacommunity framework. Ecology 2009, 90, 2253–2262. [Google Scholar] [CrossRef]
- Feinsinger, P.; Spears, E.; Poole, R. A simple measure of niche breadth. Ecology 1981, 62, 27–32. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Gaston, K.J. Rarity; Chapman & Hall: London, UK, 1994. [Google Scholar]
- Wu, A.; Liu, J.; He, F.; Wang, Y.; Zhang, X.; Duan, X.; Liu, Y.; Qian, Z. Negative relationship between diversity and productivity under plant invasion. Ecol. Res. 2018, 33, 949–957. [Google Scholar] [CrossRef]
- Mittelbach, G.; Steiner, C.; Scheiner, S.; Gross, K.; Reynolds, H.; Waide, R.; Willig, M.; Dodson, S.; Gough, L. What is the observed relationship between species richness and productivity? Ecology 2001, 82, 2381–2396. [Google Scholar] [CrossRef]
- Algina, J.; Moulder, B.C.; Moser, B.K. Sample size requirements for accurate estimation of squared semi-partial correlation coefficients. Multivar. Behav. Res. 2010, 37, 37–57. [Google Scholar] [CrossRef]
- Mori, A.S.; Isbell, F.; Seidl, R. B-diversity, community assembly, and ecosystem functioning. Trends Ecol. Evol. 2018, 33, 549–564. [Google Scholar] [CrossRef]
- Allan, E.; Weisser, W.; Weigelt, A.; Roscher, C.; Fischer, M.; Hillebrand, H. More diverse plant communities have higher functioning over time due to turnover in complementary dominant species. Proc. Natl. Acad. Sci. USA 2011, 108, 1–6. [Google Scholar] [CrossRef]
- Shurin, J.; Winder, M.; Adrian, R.; Keller, B.; Matthews, B.; Paterson, M.; Pinel-Alloul, B.; Rusak, J.; Yan, N. Environmental stability and lake zooplankton diversity—contrasting effects of chemical and thermal variability. Ecol. Lett. 2010, 13, 453–463. [Google Scholar] [CrossRef]
- Jackson, S.T.; Blois, J.L. Community ecology in a changing environment: Perspectives from the Quaternary. Proc. Natl. Acad. Sci. USA 2014, 12, 1–7. [Google Scholar] [CrossRef]
- Alves-de-souza, C.; Benevides, T.S.; Santos, J.B.O.; Dassow, P.V.O.N. Does environmental heterogeneity explain temporal β diversity of small eukaryotic phytoplankton? Example from a tropical eutrophic coastal lagoon. J. Plankton Res. 2017, 39, 698–714. [Google Scholar] [CrossRef]
- Stevens, G.C. The latitudinal gradient in geographical range: How do so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Ibarra, J.T.; Martin, K. Biotic homogenization: Loss of avian functional richness and habitat specialists in disturbed Andean temperate forests. Biol. Conserv. 2015, 192, 418–427. [Google Scholar] [CrossRef]
- Clavel, J.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2011, 9, 222–228. [Google Scholar] [CrossRef]
- Chu, J.W.F.; Curkan, C.; Tunnicliffe, V. Drivers of temporal beta diversity of a benthic community in a seasonally hypoxic fjord. R. Soc. Open Sci. 2018, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Supp, S.R.; Koons, D.N.; Ernest, S.K.M. Using life history trade-offs to understand core-transient structuring of a small mammal community. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Irz, P.; Lanoiselée, C.; Mouillot, D.; Argillier, C. Evidence that niche specialization explains species—energy relationships in lake fish communities. J. Anim. Ecol. 2008, 77, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Kotiaho, J.S.; Kaitala, V.; Komonen, A.; Paivinen, J. Predicting the risk of extinction from shared ecological characteristics. Proc. Natl. Acad. Sci. USA 2005, 102, 1963–1967. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.E.; Williams, Y.M.; Vanderwal, J.; Isaac, J.L.; Shoo, L.P.; Johnson, C.N. Ecological specialization and population size in a biodiversity hotspot: How rare species avoid extinction. Proc. Natl. Acad. Sci. USA 2009, 106, 19737–19741. [Google Scholar] [CrossRef] [Green Version]
- Altermatt, F.; Pajunen, V.I.; Ebert, D. Desiccation of rock pool habitats and its influence on population persistence in a Daphnia metacommunity. PLoS ONE 2009, 4, e4703. [Google Scholar] [CrossRef]
- Tomasovych, A.; Kidwell, S.M. Predicting the effects of increasing temporal scale on species composition, diversity, and rank-abundance distributions Author. Paleobiology 2010, 36, 672–695. [Google Scholar] [CrossRef]
- Kidwell, S.M.; Tomasovych, A. Implications of time-averaged Death Assemblages for ecology and conservation biology. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 539–563. [Google Scholar] [CrossRef]
- Korhonen, J.; Soininen, J.; Hillebrand, H. A quantitative analysis of temporal turnover in aquatic species assemblages across ecosystems. Ecology 2010, 91, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.J.; Gove, J.M.; Eynaud, Y.; Zgliczynski, B.J.; Sandin, S.A. Local human impacts decouple natural biophysical relationships on Pacific coral reefs. Ecography 2015, 38, 751–761. [Google Scholar] [CrossRef]
- Rooney, N.; Mccann, K.; Gellner, G.; Moore, J.C. Structural asymmetry and the stability of diverse food webs. Nature 2006, 442, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Seabloom, E.W. Compensation and the stability of restored grassland communities. Ecol. Appl 2007, 17, 1876–1885. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D. The ecological consequences of changes in biodiversity: A search for general principles. Ecology 1999, 80, 1455–1474. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species Type | Mean % | Standard Deviation | Extinction/Colonization Pattern |
|---|---|---|---|
| Permanent | 3.3 | 4.8 | Present in all annual samples |
| Cyclic | 53.0 | 13.0 | Recurs intermittently at least once |
| Transient | 34.5 | 10.7 | Not initially present, colonizes and goes extinct, does not recur |
| Lost from site | 3.2 | 5.0 | Initially present, goes extinct, does not recur |
| New addition | 6.1 | 7.3 | Not initially present, recurs every year after colonization |
| Variable | EV 1 (28.1%) | EV 2 (29.1%) |
|---|---|---|
| Temperature variability | 0.72 | 0.09 |
| Salinity variability | −0.44 | 0.71 |
| Dissolved O2 variability | −0.15 | −0.61 |
| pH variability | 0.66 | 0.29 |
| Chlorophyll a variability | 0.67 | −0.46 |
| Number of sampling days dry | 0.37 | 0.72 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hammond, M.; Kolasa, J. The Long and Short of Biodiversity: Cumulative Diversity and Its Drivers. Diversity 2019, 11, 41. https://doi.org/10.3390/d11030041
Hammond M, Kolasa J. The Long and Short of Biodiversity: Cumulative Diversity and Its Drivers. Diversity. 2019; 11(3):41. https://doi.org/10.3390/d11030041
Chicago/Turabian StyleHammond, Matthew, and Jurek Kolasa. 2019. "The Long and Short of Biodiversity: Cumulative Diversity and Its Drivers" Diversity 11, no. 3: 41. https://doi.org/10.3390/d11030041
APA StyleHammond, M., & Kolasa, J. (2019). The Long and Short of Biodiversity: Cumulative Diversity and Its Drivers. Diversity, 11(3), 41. https://doi.org/10.3390/d11030041