A Facilitation Cascade Enhances Local Biodiversity in Seagrass Beds


Division of Marine Science and Conservation, Nicholas School of the Environment, Duke University, Beaufort, NC 28516, USA
*
Author to whom correspondence should be addressed.
Diversity 2019, 11(3), 30; https://doi.org/10.3390/d11030030
Submission received: 17 December 2018
/
Revised: 14 February 2019
/
Accepted: 16 February 2019
/
Published: 26 February 2019
(This article belongs to the Special Issue Diversity of Ecosystem Engineers in the World Coasts and Oceans)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Invertebrate diversity can be a key driver of ecosystem functioning, yet understanding what factors influence local biodiversity remains uncertain. In many marine and terrestrial systems, facilitation cascades where primary foundation and/or autogenic ecosystem engineering species promote the settlement and survival of a secondary foundation/engineering species have been shown to enhance local biodiversity and ecosystem functioning. We experimentally tested if a facilitation cascade occurs among eelgrass (Zostera marina), pen clams (Atrina rigida), and community diversity in temperate seagrass beds in North Carolina, U.S.A., and if this sequence of direct positive interactions created feedbacks that affected various metrics of seagrass ecosystem function and structure. Using a combination of surveys and transplant experiments, we found that pen clam density and survivorship was significantly greater in seagrass beds, indicating that eelgrass facilitates pen clams. Pen clams in turn enhanced local diversity and increased both the abundance and species richness of organisms (specifically, macroalgae and fouling invertebrate fauna)—the effect of which scaled with increasing clam density. However, we failed to detect an impact of pen clams on other seagrass functions and hypothesize that functioning may more likely be enhanced in scenarios where secondary foundation species specifically increase the diversity of key functional groups such as epiphyte grazers and/or when bivalves are infaunal rather than epifaunal. Our findings add to the growing amount of literature that demonstrates that secondary foundation species are important drivers of local biodiversity in marine ecosystems. Further experimentation is needed that directly examines (i) the role of functional versus overall diversity on seagrass functions and (ii) the relative importance of life-history strategy in determining when and where engineering bivalves increase biodiversity and/or functioning of seagrass beds.
1. Introduction
It has long been recognized that cascades of species interactions can indirectly alter ecosystem processes and community organization [1,2]. Tri-trophic cascades are one well-known example that occur when double-negative feeding interactions result in predators indirectly increasing plant populations by suppressing populations of otherwise voracious grazers [3,4,5]. Facilitation cascades generated by sequences of direct positive effects between habitat-forming ecosystem engineers can also have powerful, indirect effects on marine ecosystems [6]. In habitat cascades, primary foundation species, such as mangroves and salt marsh cordgrasses, ameliorate stressful physical conditions and provide habitat for secondary foundation species such as oysters and mussels [6,7,8]. In turn, secondary foundation species further modify ecosystem structure and functioning and enhance community diversity via their own engineering and niche space provisioning [9].
Despite their likely influence on ecosystem functioning and resilience, ecological models often do not include positive interactions [10]. Moreover, the generality of habitat-mediated facilitation cascades and their application in conservation has only gained traction in the last two decades [11]. In salt marshes, facilitation cascades between ecosystem-engineering grasses and mussel aggregations have been found to enhance multiple functions including soil accretion and infiltration rate in addition to plant biomass and local biodiversity [12]. Similarly, facilitation cascades and mutualisms among mangroves, algae, and oysters have been demonstrated to increase growth rates, niche provisioning, and epifaunal diversity and abundance [13,14]. Recent syntheses have further found that positive interactions [15] and facilitation cascades [16] are especially important for enhancing resilience to physical stressors and driving patterns of biodiversity across marine, aquatic, and terrestrial ecosystems.
Studies have only recently begun to examine how biodiversity in seagrass beds may alter ecosystem functioning [17,18], and the role of facilitation cascades in determining seagrass biodiversity has similarly been under-studied. Prior studies of positive interactions between seagrasses and bivalves have focused primarily on the direct effects of bivalves on seagrass productivity [19,20,21,22,23]. These studies also did not first examine the role of seagrasses as primary foundation species prior to testing for bivalve impacts or whether bivalves may have indirect effects on tertiary species via a multi-level facilitation cascade. Secondary foundation species in seagrass beds, such as algae and bivalves, may also independently and directly facilitate fish and invertebrate settlement. [12,16]. Thus, it is possible that habitat cascades between seagrasses and secondary foundation species may also be prevalent drivers of biodiversity patterns in shallow water marine systems [24,25]. However, further examination is required to determine how common and the extent to which facilitation cascades influence seagrass community structure and ecosystem functions.
Bivalves commonly occur in seagrass ecosystems across the globe [20]. Seagrasses have been shown to first facilitate bivalves by attracting bivalve larvae and decreasing the intensity of physical and biological stressors [26,27,28]. Bivalves in turn may positively affect seagrasses by increasing water clarity and beneficially altering sediment nutrient chemistry and soil oxygenation [20,23,24,29]. Lucinid clams, for example, can reduce otherwise toxic sediment sulfide concentrations and enhance seagrass biomass via a mutualism with sulfide-oxidizing bacteria that reside within their gills [20]. Reef-building bivalves such as oysters or mussels may act as both primary or secondary foundation species depending on whether they occur within or adjacent to seagrass meadows. Oyster reefs, for example, may facilitate nearby seagrass growth by ameliorating wave stress [30]. In addition to directly impacting seagrass productivity and functioning, bivalves can also act as ecosystem engineers and create a habitat that supports greater biodiversity [7,12]. Such a habitat cascade could support the formation of tertiary habitats that may only have detectable impacts at high densities of secondary foundation species [16]. These indirect impacts may be density-dependent and increase with density or even disappear [25,31]. For example, secondary foundation species may provide positive effects on biodiversity at low densities but may compete for space or resources beyond a threshold density [8,32].
Clams in the Pinnidae family, collectively known as pen shells or fan clams, are large, habitat-forming bivalves that occur within seagrass meadows and adjacent sand flats in many of the world’s oceans and estuaries. In some regions, such as Baja California in Mexico, they occur in extremely high densities and constitute a critical and profitable fishery [33,34,35]. However, little scientific knowledge exists regarding the ecological role pen clams may play in marine systems. Previous studies have shown that pen clams can provide primary habitat and nest sites for many epifaunal organisms and fish species, particularly post-mortem [25,36]. More recently, Gribben et al. (2017) found that pen clam presence increased local diversity, but the effect was not density-dependent. This contrasts with previous studies that indicate greater facilitation with increasing density [13,37,38,39]. These studies have indicated that pen clams may act as secondary foundation species in seagrass ecosystems; however, whether the initial step in this potential facilitation cascade occurs (i.e., seagrass facilitation of pen clams) has not been tested. Moreover, whether the density-dependent effects of pen clams are general across other species is not known.
In coastal North Carolina (NC), there are two commonly occurring species of pen clam, Atrina rigida and Atrina serrata (i.e., the rigid and the sawtooth pen clam, respectively). Pen clams in NC are functionally both infaunal and epifaunal with only the upper ~1/4 of their shell above the sediment surface. Exposed portions of the shells are frequently colonized by multiple species of invertebrates and algae. Though edible, pen clams in NC are not commonly harvested. The purpose of this study was to first determine the relative distribution and abundance of pen clams in the southern Outer Banks of North Carolina, and second, to ascertain if a facilitation cascade exists between seagrasses—pen clams—and local biodiversity. In particular, we tested if seagrasses enhanced pen shell success (i.e., density and survivorship) and if pen clams in turn impacted community diversity and ecosystem functioning. We further sought to determine if impacts were density-dependent.
2. Materials and Methods
To examine how and if seagrasses and pen clams form the basis for a facilitation cascade, we conducted a survey of seagrass beds in the southern Outer Banks of North Carolina and two field experiments. For all experiments, pen clams were collected from seagrass beds in Back Sound, NC and field manipulations were conducted in Middle Marsh, Beaufort, NC, USA.
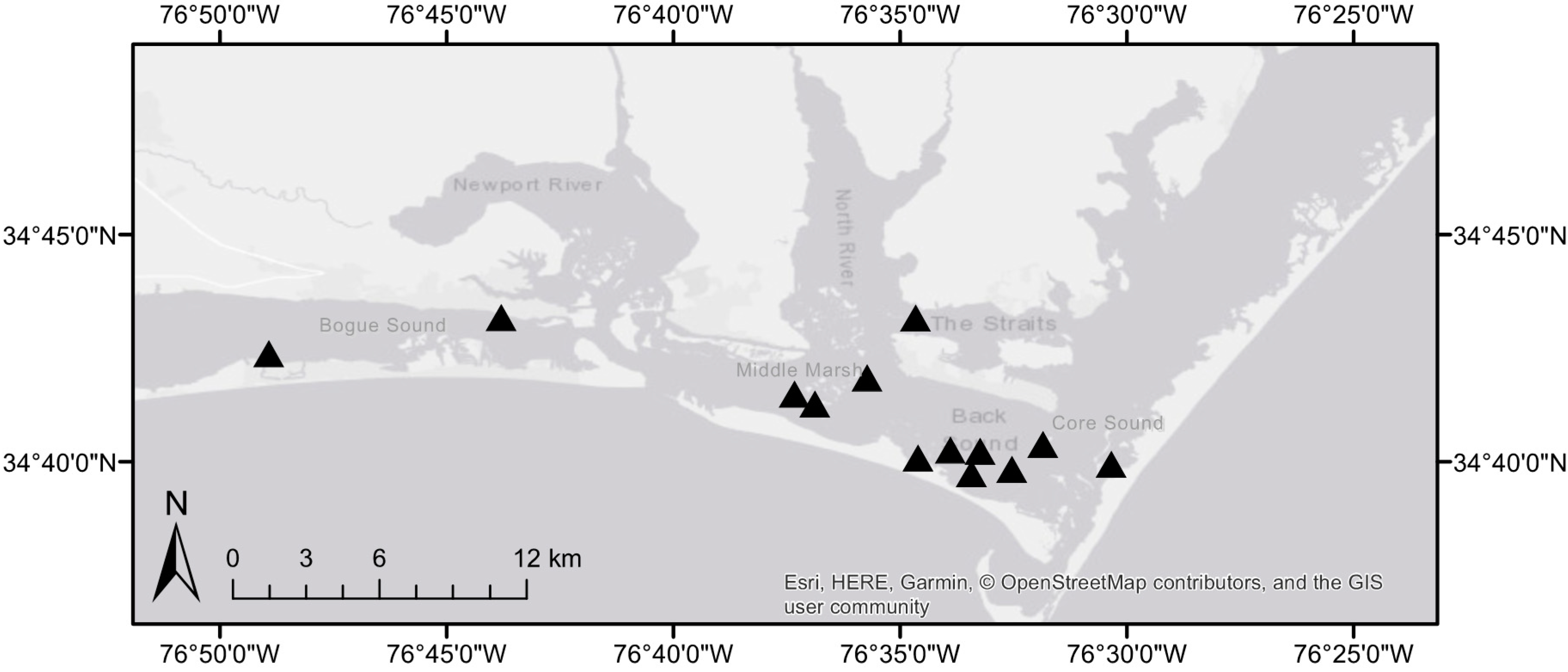
To obtain an estimate of pen clam densities in NC, we surveyed 13 seagrass beds and adjacent sand flats in Back, Bogue, and Core Sounds (Figure 1). Pen clam density was quantified using two survey methods. We first surveyed for clams by haphazardly tossing a 1 m2 quadrat across a seagrass meadow and neighboring sandflat, and recording seagrass species and number of pen clams within the quadrat (n = 25 per site). We also took a more systematic approach and conducted eight 25 m belt transect surveys in both seagrass and sandflat for a total of 16 transect surveys at each of the 13 sites. All pen clams that occurred along the transect were noted, and seagrass species was recorded at 2 m intervals.
To assess if seagrasses could facilitate pen clam survivorship, we conducted a reciprocal transplant experiment. Pen clams (n = 10 per habitat type) were collected from seagrass bed interiors and transplanted into the interior of a seagrass bed or sandflat in June 2017. Mortality, shell condition, and potential cause of death was assessed weekly for the first four weeks and monthly thereafter through October 2018.
In order to test for the presence of a density-dependent facilitation cascade, we manipulated pen clam density within plots to 0, 1, or 4 clams per m2 (n = 5) within a seagrass bed in Middle Marsh, Beaufort, NC to represent densities observed from surveys. Experiments were conducted from November 2015 through August 2017. Seagrass percent cover was measured prior to experiment implementation to ensure similar conditions across plots. Seagrass density, height, growth, and reproductive effort were measured in May 2016 and 2017 to coincide with the period of greatest eelgrass productivity in NC. Seagrass density and reproductive shoots were counted in situ within 0.25 m2 quadrats inside plots within which ten stems were measured for growth. Seagrass growth was measured by marking 10 separate seagrass shoots approximately 1 cm below the sheath. After two weeks, marked shoots were collected and brought to the lab for processing. New and old growth area and biomass were measured to determine proportional growth. We also quantified aboveground seagrass decomposition to determine if pen clams affected seagrass decay rates. Decomposition bags were constructed from 1 × 1 mm mesh screen cloth and packed and massed onshore with 50 g of live, aboveground Zostera marina collected from the same meadow but 30 m away from the experiment area [40]. Bags were deployed within plots in June 2017 for six months, then collected and the remaining material was massed.
To assess community composition within plots, we quantified mobile nekton, seagrass epibiont, macroalgal species and associated fauna, and pen clam epibiont communities. Nekton and macroalgal species present were quantified via in situ snorkeler surveys at mid-tide (~1.5 m water depth) once a month from April to August in 2016 and 2017. Observers floated above plots and enumerated all nekton that entered the plots over a three-minute duration after a two-minute acclimation period [41]. To determine if pen clams/pen clam density affected benthic, free-growing macroalgal biomass, we collected algal samples from plots in May 2016 and 2017. A 25 × 25 cm quadrat was randomly placed within the plot, and all macroalgae within the quadrat were collected and taken back to the lab for processing. Macroalgal samples were rinsed through a 500 µm sieve, dried, and weighed [42,43]. Algal-associated organisms within samples were also enumerated and identified to species. To quantify seagrass epibiont communities, 10 random stems were collected from plots and epibiota identified to family and quantified in June 2016 and 2017 to coincide with documented periods of high epibiont coverage in NC. Clam epibiont communities were assessed from photos taken in the field in August 2016 to minimize damage to plots. In August 2017 (experiment end), clams were collected and epibiont communities on clams were identified to species and enumerated. In instances where species are known to be colonial (e.g., bryozoans), organisms were recorded as present/absent rather than enumerated.
3. Results
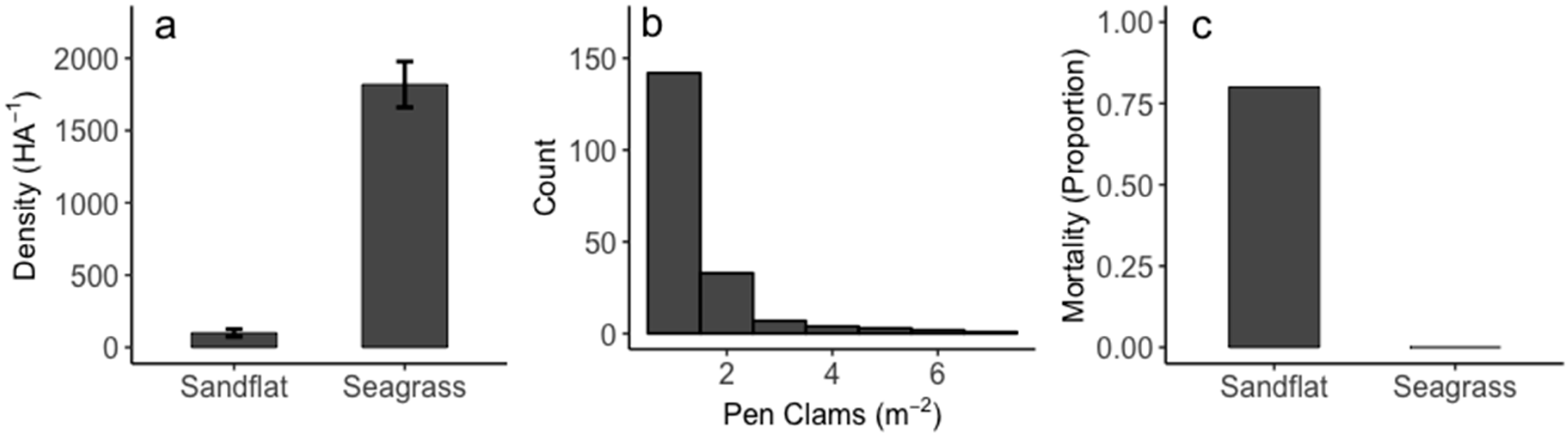
Pen clams were significantly denser and more abundant in seagrass beds compared to adjacent sandflats (Student’s t-test p < 0.001, Figure 2a) at an average of roughly 1818 per hectare in seagrasses, whereas there was on average 100 clams per hectare in adjacent sandflats. When clams were observed (13.2% of all observations in seagrass beds), they were typically solitary, however densities of up to 7 m−2 were observed (Figure 2b). Transplantation into seagrass beds and sandflats found that 16 months post-transplant, there was a significant effect of seagrasses on pen clam survivorship with 80% mortality observed in sand flats and no pen clam mortality observed in seagrass beds (Figure 2c). Assessment of shell condition post-mortem revealed that clams typically remain buried in the sediment with intact shells, potentially indicating death by senescence or due to physical stressors. In our experiments, two clams showed potential signs of predation (shell was crushed), and two were not recovered. When mortality by predation was observed, shells/shell fragments were found within one meter of where clams were originally transplanted.
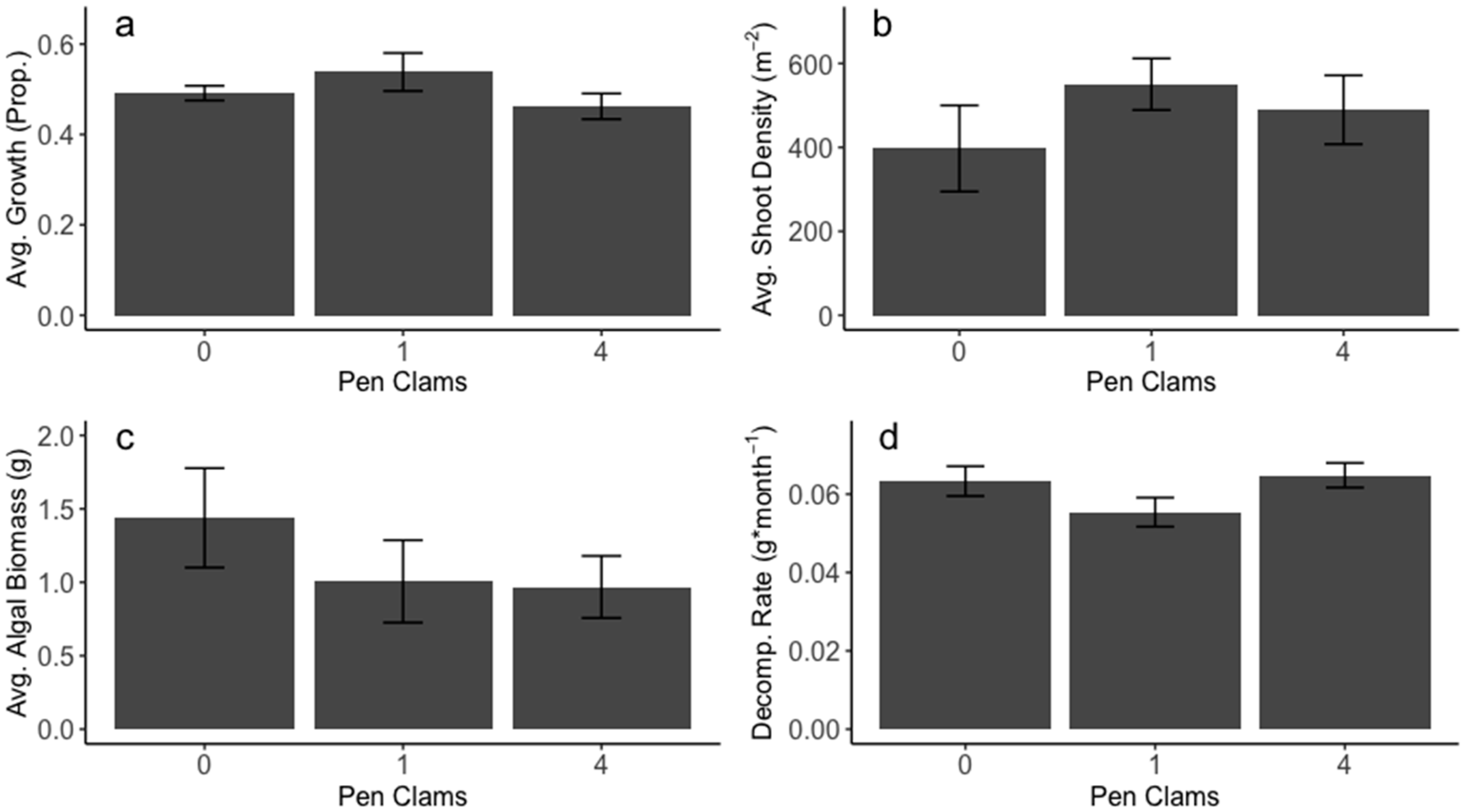
Transplantation experiments at varying densities revealed no significant effects of pen clams on aspects of seagrass ecosystem function and structure (i.e., seagrass growth seagrass density) or decomposition (ANOVA p = 0.642, p = 0.393, and p = 0.814, respectively, Figure 3a–c). We also failed to detect a relationship between algal biomass and pen clam density (ANOVA p = 0.321, Figure 3d). Because no metric of seagrass productivity differed across treatments, we did not further quantify sediment or seagrass nutrient content.
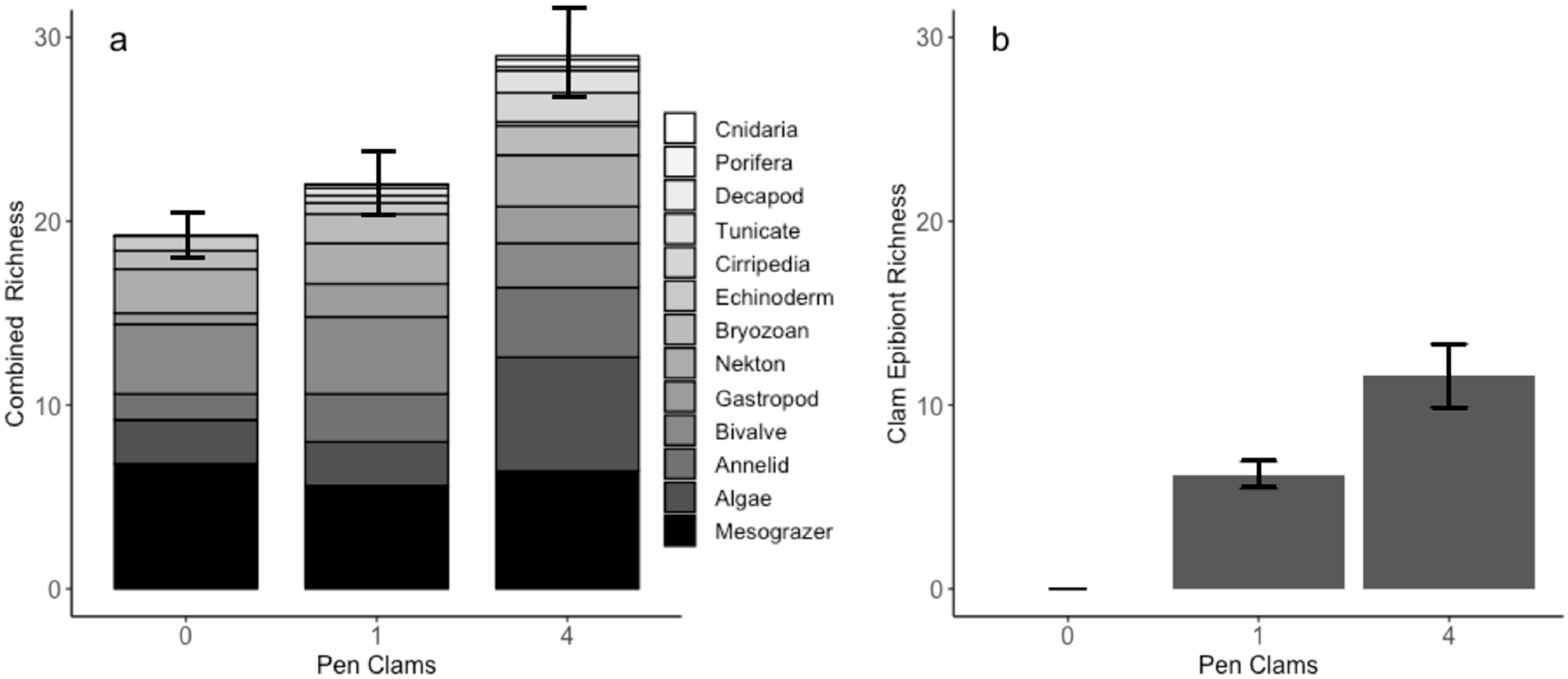
Across all replicates, the number of unique species found increased with pen clam density, with 37 species identified in the clam-absent treatment and 41 and 59 species present in the one- and four-clam treatments, respectively. Moreover, community diversity measured as average species richness increased stepwise with increasing pen clam density and was significantly different between the zero versus four and one versus four pen clam treatments (ANOVA p = 0.0043, Tukey’s Honest Significant Difference 0 vs. 4 p = 0.002, and 1 vs. 4 p = 0.016, Figure 4a). In particular, fouling invertebrates associated with epibiont communities (i.e., corals, anemones, barnacles, tunicates, sessile polychaetes, etc., ANOVA p < 0.00, Figure 4b) and algal species significantly increased in richness with increasing pen clam density (ANOVA p < 0.001). In contrast, species richness for other functional groups or communities assessed (e.g., nekton, seagrass, and algae-associated) did not vary significantly across treatments.
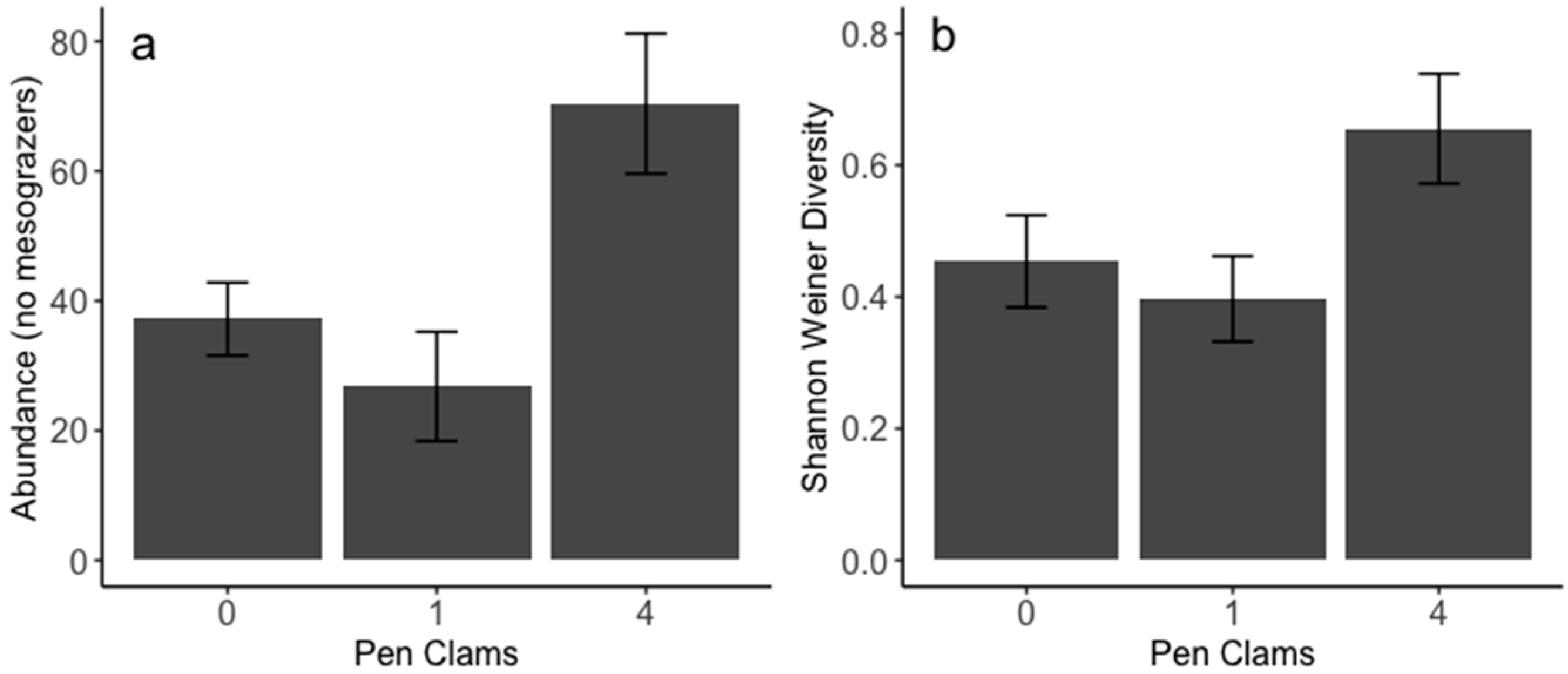
Total organismal abundance was not significantly different across treatments (ANOVA p = 0.359). Because mesograzers (amphipods, isopods, and tanaids) constituted the vast majority of the community, and their abundance did not differ with respect to clam density, we removed mesograzers from the abundance analysis to more closely examine how the rest of the community varied with respect to clam density. In the absence of mesograzers, we found that overall organismal abundance was significantly greater with four pen clams present compared to the no-clam and one-clam treatments (ANOVA p = 0.009, Tukey’s Honest Significant Difference 0 vs. 4 p = 0.044 and 1 vs. 4 p = 0.009, Figure 5a). Across all taxa, Shannon–Wiener diversity was greater at high pen clam density than both absent and one-clam treatments (ANOVA p = 0.037, Figure 5b).
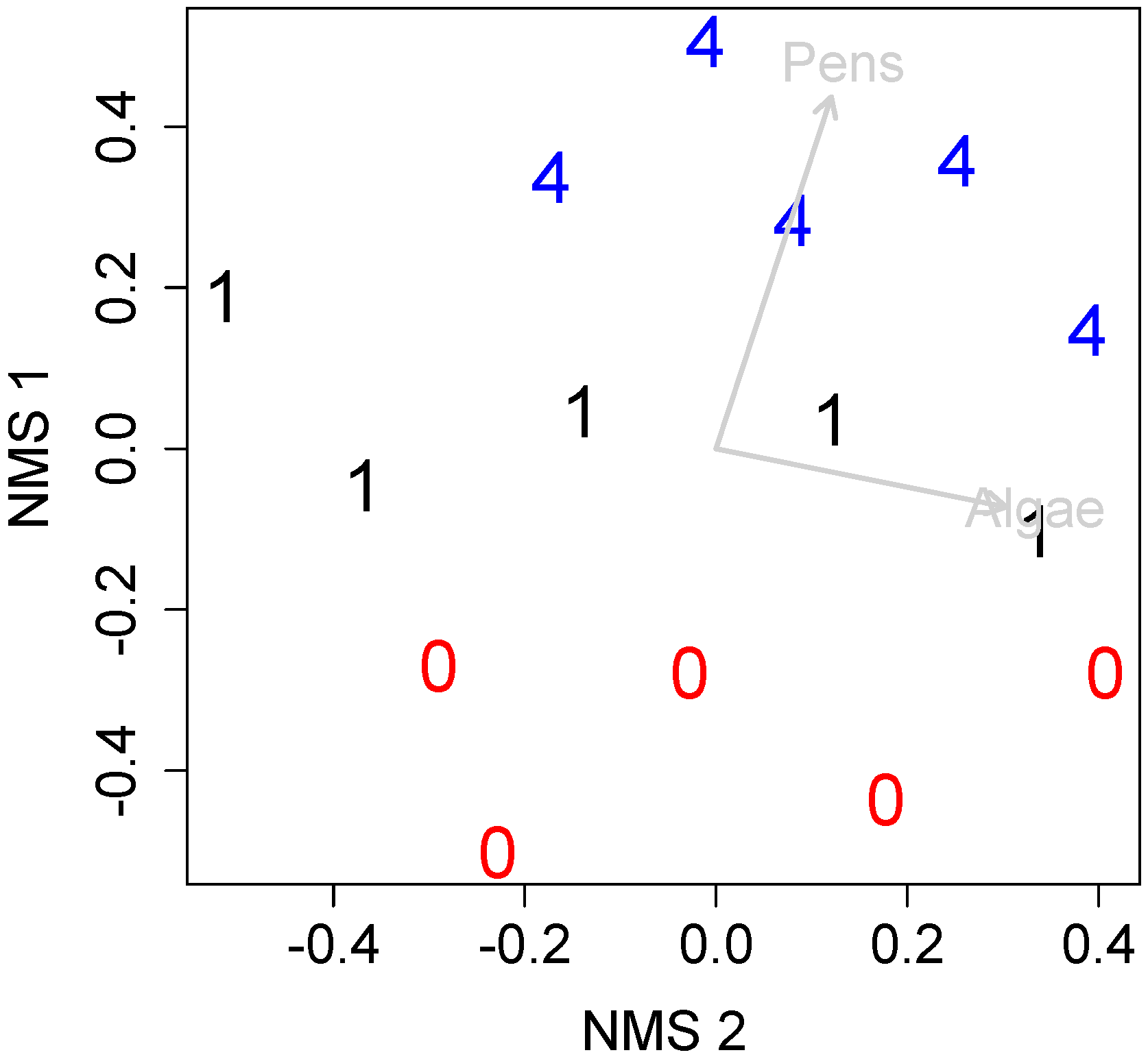
Non-metric multidimensional scaling (NMDS) further indicated that there were differences between the community composition found across treatments (stress = 0.187, Figure 6) with communities sorting most strongly by pen clam density (NMDS 1, r2 = 0.88, p = 0.01) followed by algal biomass (NMDS 2, r2 = 0.61, p = 0.04). Species indicator analysis further identified that a barnacle (Balanus eburneus) and worms, in the family Terebellidae, were strongly associated with high-density clam treatments (p = 0.001, and p = 0.038, respectively) but not low-density treatments.
4. Discussion
4.1. A Facilitation Cascade Enhances Local Biodiversity
Facilitation cascades between ecosystem engineers have been shown in numerous studies and habitats (e.g., salt marsh, temperate and tropical forests, mangroves, among others) to enhance ecosystem functions, biodiversity, and resilience to physiological stressors [6]. Using a combination of observational and experimental studies, we found support for a facilitation cascade among seagrasses and pen clams in coastal North Carolina by showing that (i) seagrasses support higher survivorship and densities of pen clams and (ii) pen clams act as secondary foundation species that enhance local diversity, which scales positively with clam density. Our findings provide further evidence that under certain scenarios, beneficial interactions, such as habitat cascades, can play a structuring role in determining biodiversity in seagrass ecosystems.
Our survey data support the conclusion that seagrasses positively influence pen clam density and survivorship, as pen clams were primarily found within seagrass habitats and not within neighboring sand flats. Our transplantation study further demonstrated that seagrasses promote the survivorship of pen clams, as 8/10 clams in sand flats died while all survived in seagrasses as of October 2018. There are likely multiple mechanisms by which seagrasses facilitated pen clam populations in our study. First, when seagrasses colonize an area, they act as ecosystem engineers and, through their three-dimensional structure, alter the physical environment in ways that can facilitate clams including dampening wave energy, altering turbulence, and changing flow velocity, among others [44,45,46]. Second, studies have shown that settlement of bivalve larvae is higher in seagrass beds compared to unvegetated habitats [26], and bivalves within seagrass beds grow at faster rates than those in nearby sandflats [47]. Finally, structural complexity provided by seagrass roots, rhizomes, and canopies can mediate predator–prey interactions and has been shown in numerous studies to reduce predator efficiency and increase bivalve survivorship, as reviewed in [48,49].
After secondary foundation species have established in seagrass beds, like pen clams, they can further provide microstructure that positively influences settlement of epibiota [26]. Despite only exposing a small portion of their shell, pen clams provide substantial sources of hard substrate when they establish on sand- or mud-dominated bottoms. This biogenic structure can promote the diversity of fouling organisms and macroalgae that require solid substrates for attachment [50]. Our results supported this hypothesis, as plots with a higher density of pen clams had significantly greater species richness than low-density plots. Thus, our findings highlight the often hierarchical structure of community organization, and the value of foundation species overlap in promoting local biodiversity.
4.2. Effects of Functional Diversity versus Diversity per Se
In our study, we found that pen clams increased diversity overall, but this increase occurred primarily in certain functional groups. Specifically, we found that pen clams increased the diversity of fouling organisms, including tunicates, barnacles, and encrusting sponges and bryozoans. There was, however, no difference detected in either amphipods or nekton across pen clam densities. This finding is consistent with the type of habitat that pen shells provide relative to their occurrence in seagrass beds. Hard substrate for fouling organisms to settle upon and colonize is relatively rare in seagrasses, thus pen clams may selectively promote organisms reliant on attachment substrate. The complex, three-dimensional structure that amphipods and fish depend upon for refuge and forage did not vary across treatments as both algal and seagrass biomass remained the same. Had plant structure and abundance differed, we would have expected to observe a greater change in mesograzer and nekton density that may have affected ecosystem functioning.
Although greater overall diversity did not directly translate into enhanced seagrass ecosystem functioning, we cannot definitively conclude that pen clams cannot alter ecosystem functions. Our findings do beg the question of the importance of overall diversity versus functional diversity. Many of the species promoted by pen clams were directly associated with the clam epibiont communities. However, several of these organisms (e.g., barnacles, bryozoans, boring sponges) may not functionally benefit seagrasses. Similarly, we also found an increase in macroalgae richness, but again, these organisms have generally been found to suppress rather than enhance seagrass growth [51].
Though many previous studies have found links between diversity and enhanced seagrass productivity [17,52,53], these studies found that the key force underlying changes in primary productivity was an increase in epiphytic grazer diversity, which was more efficient in consuming epiphytic algae that could otherwise overgrow and suppress seagrass growth. In contrast, we did not find that pen clams increased mesograzer (i.e., amphipods, isopods, tanaids) diversity or abundance [17]. Had we observed an increase in mesograzers in relation to pen clam density, we hypothesize that we would have seen a concomitant increase in ecosystem function [54].
4.3. Bivalves as Key Drivers of Seagrass Structure and Function
Bivalves play unique and foundational roles across multiple marine and aquatic systems [55]. The functional role bivalves play within an ecosystem, however, is context-dependent and strongly related to differences in life-history strategy, that is, whether they are epibenthic or infaunal, solitary or reef-building, intertidal or subtidal, etc. [55]. As filter feeders, they affect benthic-pelagic coupling, alter nutrient cycling, affect food webs, and modify trophic subsidies to nearby habitats [56]. Many reef-building species also act as ecosystem engineers by creating and modifying habitat [57,58,59].
Epibenthic and reef-building bivalves, such as oysters and mussels, may act as autogenic ecosystem engineers that create niche space for many small invertebrates thus enhancing local diversity [57,59,60,61]. They may also serve as substrate for algal epibionts which, in turn, may act as secondary engineers or foundation species [62]. At high densities, however, they may compete with other foundation species for space [63]. In contrast, infaunal bivalves more often function as allogenic ecosystem engineers and can modify the physical environment by providing nutrient subsidies via pseudofeces, increasing oxygen penetration into the sediment, and stimulating microbial metabolism via bioturbation [55,64,65,66,67]. Moreover, many infaunal clams can further facilitate seagrasses by reducing sulfides in the sediment via a mutualistic interaction with bacteria that reside in their gills [20].
Our study examined a bivalve that is a mix of both infaunal and epibenthic. Although pen clams in our study region do not occur in high enough densities to form reefs, we found densities of up to 7 m−2 in our surveys, and in other regions of the world, pen clams have been found in aggregations of 14 m−2 [68]. Despite having the majority of their shell buried beneath the sediment surface, bivalves can provide a substantial source of hard substrate for colonization in an otherwise soft bottom [69,70]. Post-mortem they can also serve as refugia for many small fish and crustacean species as their shells can remain intact and partially buried in the sediment for many months before degrading [25,71]. Thus, we predict that the impacts of bivalves on seagrass functions are likely to vary not only with density but also with functional group. Specifically, we hypothesize that, while alive, infaunal bivalves will facilitate seagrass primarily by increasing nutrient availability in sediments and reduce sulfide stress in contrast to epifaunal bivalves that are more likely to promote biodiversity and nursery functions. Post-mortem, however, both infaunal and epifaunal bivalve shells that rise to the sediment surface can promote biodiversity via the provision of refugia and hard substrate [72].
5. Conclusions
A large body of ecological research has focused on how cascades of negative trophic or competitive interactions can structure communities and are mediated by biodiversity [73]. Ecological studies have only begun to elucidate the important role of positive interactions and facilitation cascades in generating habitats and determining the distribution and abundance of biodiversity [7,8,12,16]. Our findings suggest that biodiversity in North Carolina seagrass communities is influenced by a facilitation cascade whereby seagrasses facilitate bivalves and, in turn, those bivalves facilitate fouling and algal diversity.
Our results also have implications for the conservation and restoration of seagrass ecosystems. Restoration of seagrasses that includes the simultaneous planting of both primary and secondary foundation species such as seagrasses and bivalves is likely to yield greater biodiversity. Greater emphasis should be placed on the enhancement of habitat cascade units (secondary foundation species/ecosystem engineers) and whole ecosystems, not just primary foundation species, that promote positive interactions and ecosystem functioning [74]. We suggest that future ecological studies and conservation actions further incorporate facilitation cascades, as they are likely key but underappreciated drivers of spatial and temporal variation of biodiversity and function in seagrass beds.
Author Contributions
Y.S.Z. and B.R.S. conceived and designed the study. Y.S.Z. conducted the investigation, analyzed the data, and acquired funding. Y.S.Z. and B.R.S. wrote and edited the paper.
Funding
This research was funded by a Duke University Wetlands Center Graduate Fellowship and a Joseph P. Ramus Estuarine Research Grant to Y.S. Zhang.
Acknowledgments
We thank D. Barbarich, C. Chen, L. Gaskins, E. Hardgrove, C. Hoyt, S. Huff, B. Hughes, C. Humphreys, J. Fodrie, D. Keller, D. Loh, A. Mercer, J. Morton, S. Ou, N. Roberts, L. Shaver, and W. Wright for field assistance. This work was conducted partially within the Rachel Carson National Research Reserve, for whom we are grateful for supporting our work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hairston, N.G.; Smith, F.E.; Slobodkin, L.B. Community structure, population control, and competition. Am. Nat. 1960, 94, 421–425. [Google Scholar] [CrossRef]
- Paine, R.T. Food web complexity and species diversity. Am. Nat. 1966, 100, 65–75. [Google Scholar] [CrossRef]
- Estes, J.A.; Duggins, D.O. Sea otters and kelp forests in Alaska: Generality and variation in a community ecological paradigm. Ecol. Monogr. 1995, 65, 75–100. [Google Scholar] [CrossRef]
- Terborgh, J.; Lopez, L.; Nuñez, P.; Rao, M.; Shahabuddin, G.; Orihuela, G.; Riveros, M.; Ascanio, R.; Adler, G.H.; Lambert, T.D.; et al. Ecological meltdown in predator-free forest fragments. Science 2001, 294, 1923–1926. [Google Scholar] [CrossRef] [PubMed]
- Silliman, B.R.; Zieman, J.C. Top-down control of Spartina alterniflora production by periwinkle grazing in a Virginia salt marsh. Ecology 2001, 82, 2830–2845. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Wernberg, T.; Altieri, A.; Tuya, F.; Gulbransen, D.; McGlathery, K.J.; Holmer, M.; Silliman, B.R. Habitat cascades: The conceptual context and global relevance of facilitation cascades via habitat formation and modification. Integr. Comp. Biol. 2010, 50, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Altieri, A.H.; Silliman, B.R.; Bertness, M.D. Hierarchical organization via a facilitation cascade in intertidal cordgrass bed communities. Am. Nat. 2007, 169, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Angelini, C.; Altieri, A.H.; Silliman, B.R.; Bertness, M.D. Interactions among foundation species and their consequences for community organization, biodiversity, and conservation. BioScience 2011, 61, 782–789. [Google Scholar] [CrossRef]
- Angelini, C.; Silliman, B.R. Secondary foundation species as drivers of trophic and functional diversity: Evidence from a tree–epiphyte system. Ecology 2014, 95, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Halpern, B.S.; Silliman, B.R.; Olden, J.D.; Bruno, J.P.; Bertness, M.D. Incorporating positive interactions in aquatic restoration and conservation. Front. Ecol. Environ. 2007, 5, 153–160. [Google Scholar] [CrossRef]
- Angelini, C.; van der Heide, T.; Griffin, J.N.; Morton, J.P.; Derksen-Hooijberg, M.; Lamers, L.P.; Smolders, A.J.; Silliman, B.R. Foundation species’ overlap enhances biodiversity and multifunctionality from the patch to landscape scale in southeastern United States salt marshes. Proc. R. Soc. B 2015, 282, 20150421. [Google Scholar] [CrossRef] [PubMed]
- Bishop, M.J.; Byers, J.E.; Marcek, B.J.; Gribben, P.E. Density-dependent facilitation cascades determine epifaunal community structure in temperate Australian mangroves. Ecology 2012, 93, 1388–1401. [Google Scholar] [CrossRef] [PubMed]
- Ellison, A.M.; Farnsworth, E.J.; Twilley, R.R. Facultative mutualism between red mangroves and root-fouling sponges in Belizean mangal. Ecology 1996, 77, 175–188. [Google Scholar] [CrossRef]
- He, Q.; Bertness, M.D.; Altieri, A.H. Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 2013, 16, 695–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, M.S.; Altieri, A.H.; Angelini, C.; Bishop, M.J.; Gribben, P.E.; Lear, G.; He, Q.; Schiel, D.R.; Silliman, B.R.; South, P.M.; et al. Secondary foundation species enhance biodiversity. Nat. Ecol. Evol. 2018, 2, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.E.; Reynolds, P.L.; Boström, C.; Coyer, J.A.; Cusson, M.; Donadi, S.; Douglass, J.G.; Eklöf, J.S.; Engelen, A.H.; Eriksson, B.K. Biodiversity mediates top–down control in eelgrass ecosystems: A global comparative-experimental approach. Ecol. Lett. 2015, 18, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.E.; Richardson, J.P.; Canuel, E.A. Grazer diversity effects on ecosystem functioning in seagrass beds. Ecol. Lett. 2003, 6, 637–645. [Google Scholar] [CrossRef]
- Peterson, B.J.; Heck, K.L., Jr. An experimental test of the mechanism by which suspension feeding bivalves elevate seagrass productivity. Mar. Ecol. Prog. Ser. 2001, 218, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Van der Heide, T.; Govers, L.L.; de Fouw, J.; Olff, H.; van der Geest, M.; van Katwijk, M.M.; Piersma, T.; van de Koppel, J.; Silliman, B.R.; Smolders, A.J. A three-stage symbiosis forms the foundation of seagrass ecosystems. Science 2012, 336, 1432–1434. [Google Scholar] [CrossRef] [PubMed]
- Castorani, M.C.; Glud, R.N.; Hasler-Sheetal, H.; Holmer, M. Light indirectly mediates bivalve habitat modification and impacts on seagrass. J. Exp. Mar. Biol. Ecol. 2015, 472, 41–53. [Google Scholar] [CrossRef]
- Vinther, H.F.; Laursen, J.S.; Holmer, M. Negative effects of blue mussel (Mytilus edulis) presence in eelgrass (Zostera marina) beds in Flensborg fjord, Denmark. Estuar. Coast. Shelf Sci. 2008, 77, 91–103. [Google Scholar] [CrossRef]
- Reusch, T.B.H.; Chapman, A.R.O.; Groger, J.P. Blue mussels Mytilus edulis do not interfere with eelgrass Zostera marina but fertilize shoot growth through biodeposition. Mar. Ecol. Prog. Ser. 1994, 108, 265–282. [Google Scholar] [CrossRef]
- Wall, C.C.; Peterson, B.J.; Gobler, C.J. Facilitation of seagrass Zostera marina productivity by suspension-feeding bivalves. Mar. Ecol. Prog. Ser. 2008, 357, 165–174. [Google Scholar] [CrossRef]
- Gribben, P.E.; Kimbro, D.L.; Vergés, A.; Gouhier, T.C.; Burrell, S.; Garthwin, R.G.; Cagigas, M.L.; Tordoff, Y.; Poore, A.G.B. Positive and negative interactions control a facilitation cascade. Ecosphere 2017, 8, e02065. [Google Scholar] [CrossRef] [Green Version]
- Bologna, P.A.X.; Heck, K.L. Impacts of seagrass habitat architecture on bivalve settlement. Estuaries 2000, 23, 449–457. [Google Scholar] [CrossRef]
- Williams, S.L.; Heck, K.L. Seagrass Community Ecology. In Marine Community Ecology; Bertness, M.D., Gaines, S.D., Hay, M.E., Eds.; Sinauer Associates: Sunderland, MA, USA, 2001; pp. 317–331. [Google Scholar]
- Eckman, J.E. The role of hydrodynamics in recruitment, growth, and survival of Agropecten irradians (L.) and Anomia simplex (D’Orbigny) within eelgrass meadows. J. Exp. Mar. Biol. Ecol. 1987, 106, 165–191. [Google Scholar] [CrossRef]
- Peterson, B.J.; Heck, K.L., Jr. Positive interactions between suspension-feeding bivalves and seagrass-a facultative mutualism. Mar. Ecol. Prog. Ser. 2001, 213, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Goff, J.; Moody, R.M.; Byron, D.; Heck, K.L.; Powers, S.P.; Ferraro, C.; Cebrian, J. Do restored oyster reefs benefit seagrasses? An experimental study in the Northern Gulf of Mexico. Restor. Ecol. 2016, 24, 306–313. [Google Scholar] [CrossRef]
- Gascoigne, J.C.; Beadman, H.A.; Saurel, C.; Kaiser, M.J. Density dependence, spatial scale and patterning in sessile biota. Oecologia 2005, 145, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Bertness, M.D. Ribbed mussels and Spartina alterniflora production in a new england salt marsh. Ecology 1984, 65, 1794–1807. [Google Scholar] [CrossRef]
- Basurto, X. Commercial diving and the callo de hacha fishery in Seri territory. J. Southwest 2006, 48, 189–209. [Google Scholar]
- Ahumada-Sempoal, M.A.; Serrano-Guzmán, S.J.; Ruiz-García, N. Abundancia, estructura poblacional y crecimiento de Atrina maura (Bivalvia: Pinnidae) en una laguna costera tropical del Pacífico mexicano. Rev. Biol. Trop. 2002, 50, 1091–1100. [Google Scholar] [PubMed]
- García-March, J.R.; García-Carrascosa, A.M.; Peña Cantero, A.L.; Wang, Y.G. Population structure, mortality and growth of Pinna nobilis Linnaeus, 1758 (Mollusca, Bivalvia) at different depths in Moraira bay (Alicante, Western Mediterranean). Mar. Biol. 2006, 150, 861. [Google Scholar] [CrossRef]
- Keough, M.J. Effects of patch size on the abundance of sessile marine invertebrates. Ecology 1984, 65, 423–437. [Google Scholar] [CrossRef]
- Van Hulzen, J.B.; van Soelen, J.; Bouma, T.J. Morphological variation and habitat modification are strongly correlated for the autogenic ecosystem engineer Spartina anglica (common cordgrass). Estuar. Coasts 2007, 30, 3–11. [Google Scholar] [CrossRef]
- Irving, A.D.; Bertness, M.D. Trait-dependent modification of facilitation on cobble beaches. Ecology 2009, 90, 3042–3050. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.D.G.; O’Riley, J.L. Non-linear density-dependent effects of an intertidal ecosystem engineer. Oecologia 2011, 166, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.I.; Pergent, G.; Fazi, S. Chapter 16—Seagrass decomposition. In Global Seagrass Research Methods; Short, F.T., Coles, R.G., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2001; pp. 313–324. [Google Scholar]
- Edgar, G.J.; Mukai, H.; Orth, R.J. Fish, crabs, shrimps and other large mobile epibenthos: Measurement methods for their biomass and abundance in seagrass. In Glogal Seagrass Research Methods; Elsevier: Amesterdam, The Netherlands, 2001; Volume 33, pp. 255–270. [Google Scholar]
- Kendrick, G.A.; Lavery, P.S. Assessing biomass, assemblage structure and productivity of algal epiphytes on seagrasses. In Glogal Seagrass Research Methods; Elsevier: Amesterdam, The Netherlands, 2001; pp. 199–222. [Google Scholar]
- Sidik, B.J.; Bandeira, S.O.; Milchakova, N.A. Chapter 11—Methods to measure macroalgal biomass and abundance in seagrass meadows. In Global Seagrass Research Methods; Short, F.T., Coles, R.G., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2001; pp. 223–235. [Google Scholar]
- Gacia, E.; Duarte, C.M. Sediment retention by a Mediterranean Posidonia oceanica meadow: The balance between deposition and resuspension. Estuar. Coast. Shelf Sci. 2001, 52, 505–514. [Google Scholar] [CrossRef]
- Ward, L.G.; Kemp, W.M.; Boynton, W.R. The influence of waves and seagrass communities on suspended particulates in an estuarine embayment. Mar. Geol. 1984, 59, 85–103. [Google Scholar] [CrossRef]
- Fonseca, M.S.; Fisher, J.S.; Zieman, J.C.; Thayer, G.W. Influence of the seagrass, Zostera marina L. on current flow. Estuar. Coast. Shelf Sci. 1982, 15, 351–364. [Google Scholar] [CrossRef]
- Peterson, C.H.; Summerson, H.C.; Duncan, P.B. The influence of seagrass cover on population structure and individual growth of a suspension-feeding bivalve, Mercenaria mercenaria. J. Mar. Res. 1984, 42, 123–138. [Google Scholar] [CrossRef]
- Heck, K.L.; Orth, R.J. Predation in seagrass beds. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 537–550. [Google Scholar]
- Orth, R.J.; Heck, K.L.; van Montfrans, J. Faunal communities in seagrass beds: A review of the influence of plant structure and prey characteristics on predator-prey relationships. Estuaries 1984, 7, 339–350. [Google Scholar] [CrossRef]
- Keough, M.J. Dynamics of the epifauna of the bivalve Pinna bicolor: Interactions among recruitment, predation, and competition. Ecology 1984, 65, 677–688. [Google Scholar] [CrossRef]
- Hauxwell, J.; Cebrián, J.; Furlong, C.; Valiela, I. Macroalgal canopies contribute to eelgrass (Zostera marina) decline in temperate estuarine ecosystems. Ecology 2001, 82, 1007–1022. [Google Scholar] [CrossRef]
- Hughes, A.R.; Stachowicz, J.J. Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. Proc. Natl. Acad. Sci. USA 2004, 101, 8998–9002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, P.L.; Richardson, J.P.; Duffy, J.E. Field experimental evidence that grazers mediate transition between microalgal and seagrass dominance. Limnol. Oceanogr. 2014, 59, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Palmer, M.A.; Collins, S.L. Species diversity enhances ecosystem functioning through interspecific facilitation. Nature 2002, 415, 426. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, C.C.; Hoellein, T.J. Bivalve Impacts in Freshwater and Marine Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 183–208. [Google Scholar] [CrossRef]
- Dame, R.F. Ecology of Marine Bivalves: An Ecosystem Approach; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Bahr, L.M.; Lanier, W.P. The Ecology of Intertidal Oyster Reefs of the South Atlantic Coast: A Community Profile; 81/15; US Fish and Wildlife Service: Washington, DC, USA, 1981.
- Lenihan, H.S.; Peterson, C.H.; Byers, J.E.; Grabowski, J.H.; Thayer, G.W.; Colby, D.R. Cascading of habitat degradation: Oyster reefs invaded by refugee fishes escaping stress. Ecol. Appl. 2001, 11, 764–782. [Google Scholar] [CrossRef]
- Wells, H.W. The fauna of oyster beds, with special reference to the salinity factor. Ecol. Monogr. 1961, 31, 239–266. [Google Scholar] [CrossRef]
- Gedan, K.B.; Kellogg, L.; Breitburg, D.L. Accounting for multiple foundation species in oyster reef restoration benefits. Restor. Ecol. 2014, 22, 517–524. [Google Scholar] [CrossRef]
- Grabowski, J.H.; Peterson, C.H. Restoring oyster reefs to recover ecosystem services. Theor. Ecol. Ser. 2007, 4, 281–298. [Google Scholar]
- Gutiérrez, J.L.; Bagur, M.; Palomo, M.G. Algal Epibionts as Co-Engineers in Mussel Beds: Effects on Abiotic Conditions and Mobile Interstitial Invertebrates. Diversity 2019, 11, 17. [Google Scholar] [CrossRef]
- Menge, B.A. Organization of the new england rocky intertidal community: Role of predation, competition, and environmental heterogeneity. Ecol. Monogr. 1976, 46, 355–393. [Google Scholar] [CrossRef]
- Levinton, J. Bioturbators as ecosystem engineers: Control of the sediment fabric, inter-individual interactions, and material fluxes. In Linking Species & Ecosystems; Springer: Boston, MA, USA, 1995; pp. 29–36. [Google Scholar]
- Vaughn, C.C.; Hakenkamp, C.C. The functional role of burrowing bivalves in freshwater ecosystems. Freshwat. Biol. 2001, 46, 1431–1446. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Rosenberg, R. Ecosystem engineering: The impact of bioturbation on biogeochemical processes in marine and freshwater benthic habitats. Aquat. Sci. 2006, 68, 434–442. [Google Scholar] [CrossRef]
- Kristensen, E.; Penha-Lopes, G.; Delefosse, M.; Valdemarsen, T.; Quintana, C.O.; Banta, G.T. What is bioturbation? The need for a precise definition for fauna in aquatic sciences. Mar. Ecol. Prog. Ser. 2012, 446, 285–302. [Google Scholar] [CrossRef] [Green Version]
- Escamilla-Montes, R.; Diarte-Plata, G.; Luna, A.; Arturo Fierro-Coronado, J.; Esparza-Leal, H.; Granados-Alcantar, S.; Ruiz-Verdugo, C.A. Ecology, Fishery and Aquaculture in Gulf of California, Mexico: Pen Shell Atrina Maura (Sowerby, 1835); IntechOpen: London, UK, 2017. [Google Scholar]
- Beckett, D.C.; Green, B.W.; Thomas, S.A.; Miller, A.C. Epizoic invertebrate communities on upper Mississippi River unionid bivalves. Am. Midl. Nat. 1996, 135, 102–114. [Google Scholar] [CrossRef]
- Gribben, P.E.; Byers, J.E.; Clements, M.; McKenzie, L.A.; Steinberg, P.D.; Wright, J.T. Behavioural interactions between ecosystem engineers control community species richness. Ecol. Lett. 2009, 12, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, M.L. Spatial and temporal patterns in the dynamics and use of pen shells (Atrina rigida) as shelters in St. Joseph Bay, Florida. Bull. Mar. Sci. 1998, 62, 157–179. [Google Scholar]
- Gutiérrez, J.; Iribarne, O. Role of Holocene beds of the stout razor clam Tagelus plebeius in structuring present benthic communities. Mar. Ecol. Prog. Ser. 1999, 185, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Finke, D.L.; Denno, R.F. Predator diversity dampens trophic cascades. Nature 2004, 429, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.S.; Cioffi, W.R.; Cope, R.; Daleo, P.; Heywood, E.; Hoyt, C.; Smith, C.S.; Silliman, B.R. A global synthesis reveals gaps in coastal habitat restoration research. Sustainability 2018, 10, 1040. [Google Scholar] [CrossRef]
Figure 1.
Survey locations (n = 13) of seagrass beds and adjacent sand flats.

Figure 2.
Pen clam (a) distribution and density from surveys of 13 seagrass beds and adjacent sandflats, (b) histogram of observed pen clam occurrences, and (c) observed mortality and location of transplanted pen clams. Error bars represent standard errors.
Figure 2.
Pen clam (a) distribution and density from surveys of 13 seagrass beds and adjacent sandflats, (b) histogram of observed pen clam occurrences, and (c) observed mortality and location of transplanted pen clams. Error bars represent standard errors.

Figure 3.
Ecosystem function responses from density-dependent pen clam transplant experiment. (a) Average proportional growth of seagrasses, (b) average seagrass shoot density per m2, (c) average algal biomass per m2, and (d) decomposition rate (g/month).
Figure 3.
Ecosystem function responses from density-dependent pen clam transplant experiment. (a) Average proportional growth of seagrasses, (b) average seagrass shoot density per m2, (c) average algal biomass per m2, and (d) decomposition rate (g/month).

Figure 4.
Quantified species richness in plots from combined (a) nekton, algal, and shell communities averaged across treatments, gray-toned by taxonomic group, and (b) species richness of clam epibiont communities alone. Error bars represent standard errors.
Figure 4.
Quantified species richness in plots from combined (a) nekton, algal, and shell communities averaged across treatments, gray-toned by taxonomic group, and (b) species richness of clam epibiont communities alone. Error bars represent standard errors.

Figure 5.
Overall (a) organismal abundance without mesograzers and (b) Shannon–Wiener diversity associated with pen clams. Error bars represent standard errors.
Figure 5.
Overall (a) organismal abundance without mesograzers and (b) Shannon–Wiener diversity associated with pen clams. Error bars represent standard errors.

Figure 6.
Non-metric multidimensional scaling plot of pen clam communities. Numbers and colors indicate clam density per plot (treatment). Gray arrows represent correlation vectors where p < 0.1. Stress = 0.187.
Figure 6.
Non-metric multidimensional scaling plot of pen clam communities. Numbers and colors indicate clam density per plot (treatment). Gray arrows represent correlation vectors where p < 0.1. Stress = 0.187.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.S.; Silliman, B.R. A Facilitation Cascade Enhances Local Biodiversity in Seagrass Beds. Diversity 2019, 11, 30. https://doi.org/10.3390/d11030030
AMA Style
Zhang YS, Silliman BR. A Facilitation Cascade Enhances Local Biodiversity in Seagrass Beds. Diversity. 2019; 11(3):30. https://doi.org/10.3390/d11030030
Chicago/Turabian StyleZhang, Y. Stacy, and Brian R. Silliman. 2019. "A Facilitation Cascade Enhances Local Biodiversity in Seagrass Beds" Diversity 11, no. 3: 30. https://doi.org/10.3390/d11030030
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.