Exploring the Bacterial Communities of Infernaccio Waterfalls: A Phenotypic and Molecular Characterization of Acinetobacter and Pseudomonas Strains Living in a Red Epilithic Biofilm

 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Site and Sample Collection
2.2. DNA Extraction, High Throughput Sequencing (HTS) Analysis
2.3. Analysis of 16S rRNA Sequences from Microbial Assemblages
2.4. Red Epilithic Cultivable Bacterial Community Characterization
2.5. Random Amplified Polymorphic DNA (RAPD) Analysis
2.6. Phenotypic Characterization of Acinetobacter and Pseudomonas spp. Bacterial Strains: Resistance to Antibiotics and Heavy Metals, and Statistical Analysis
3. Results
3.1. Chemico-Physical Characterization of Water and Biofilm
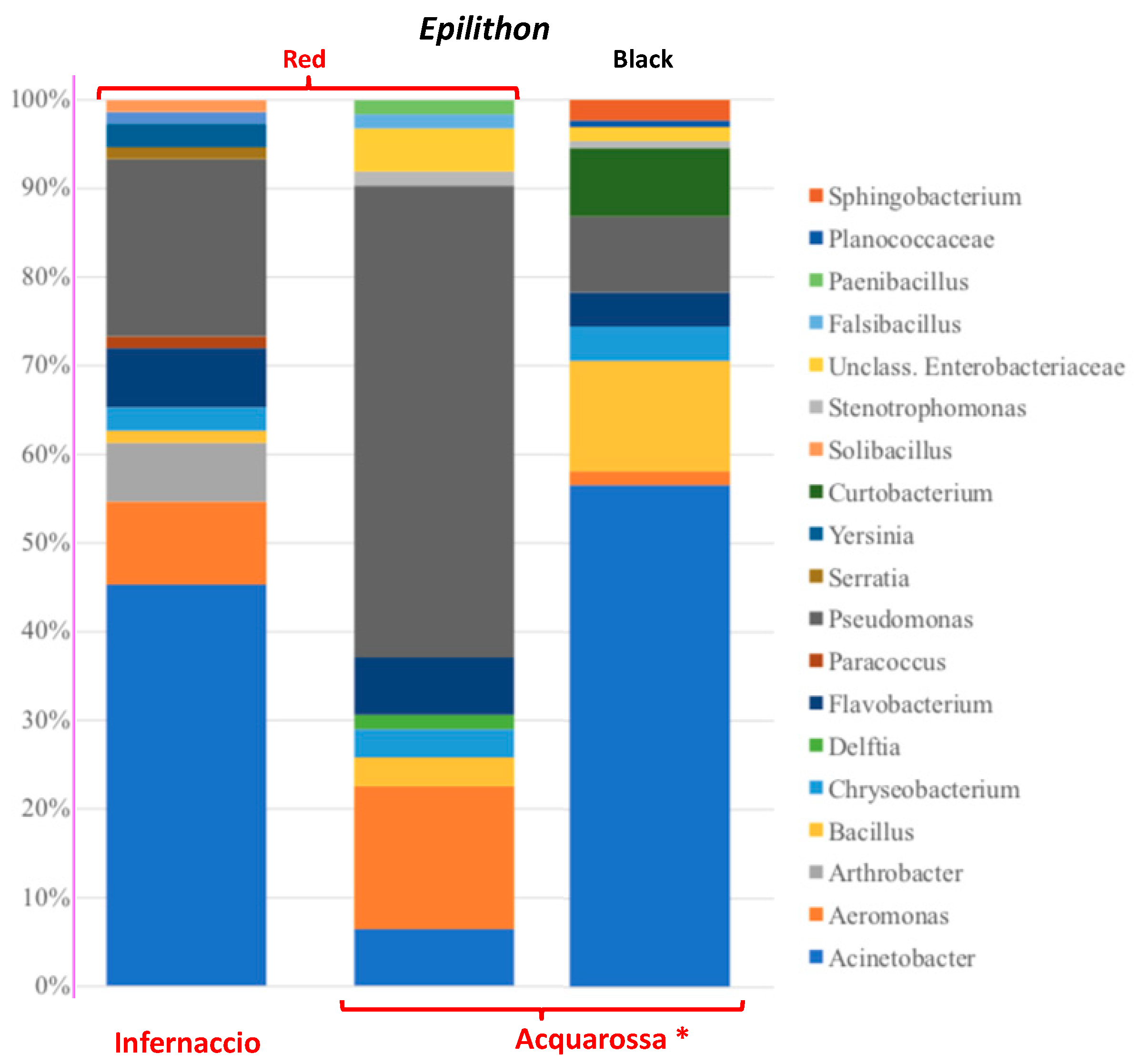
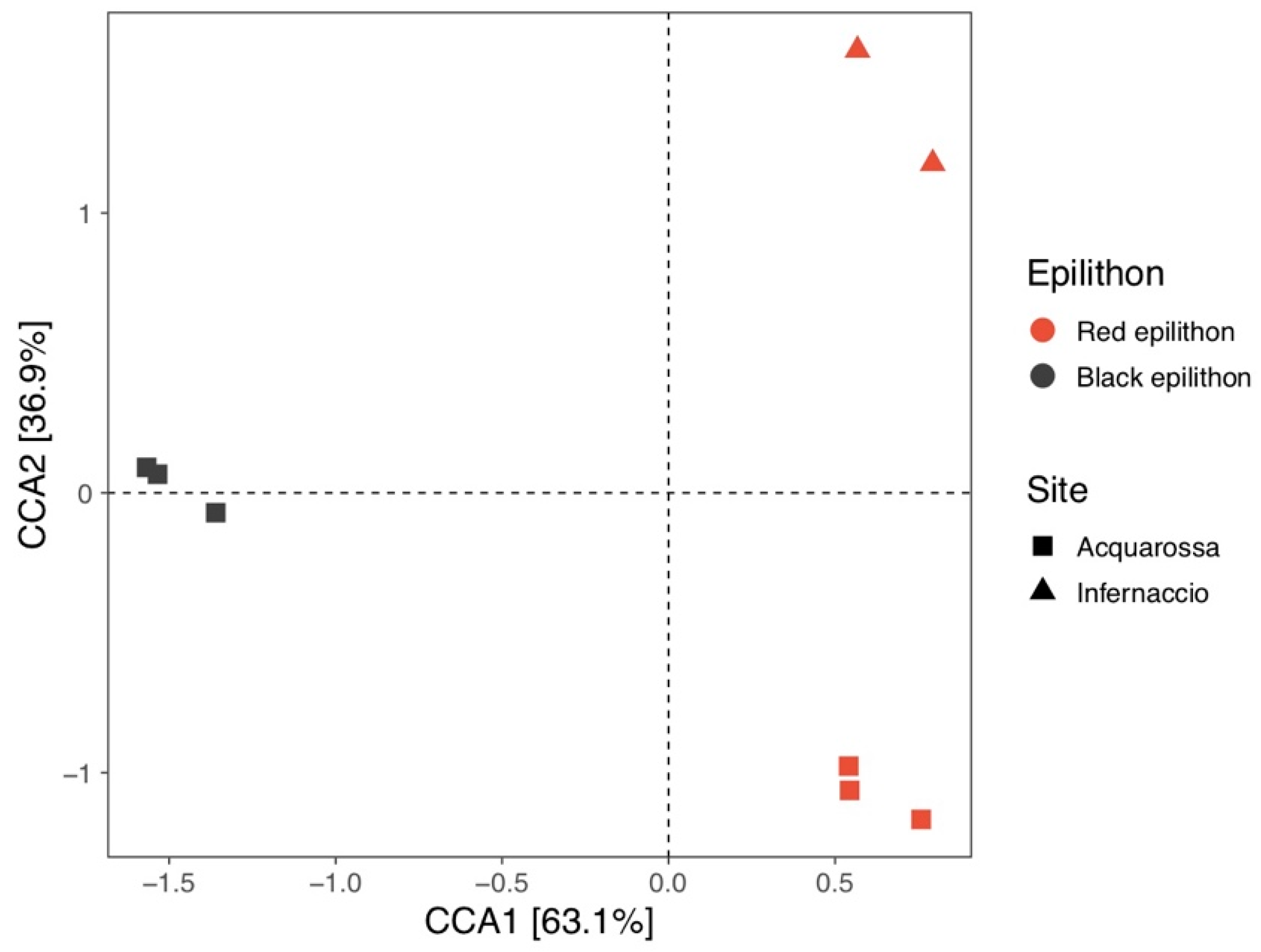
3.2. High Throughput Sequencing (HTS) Analysis
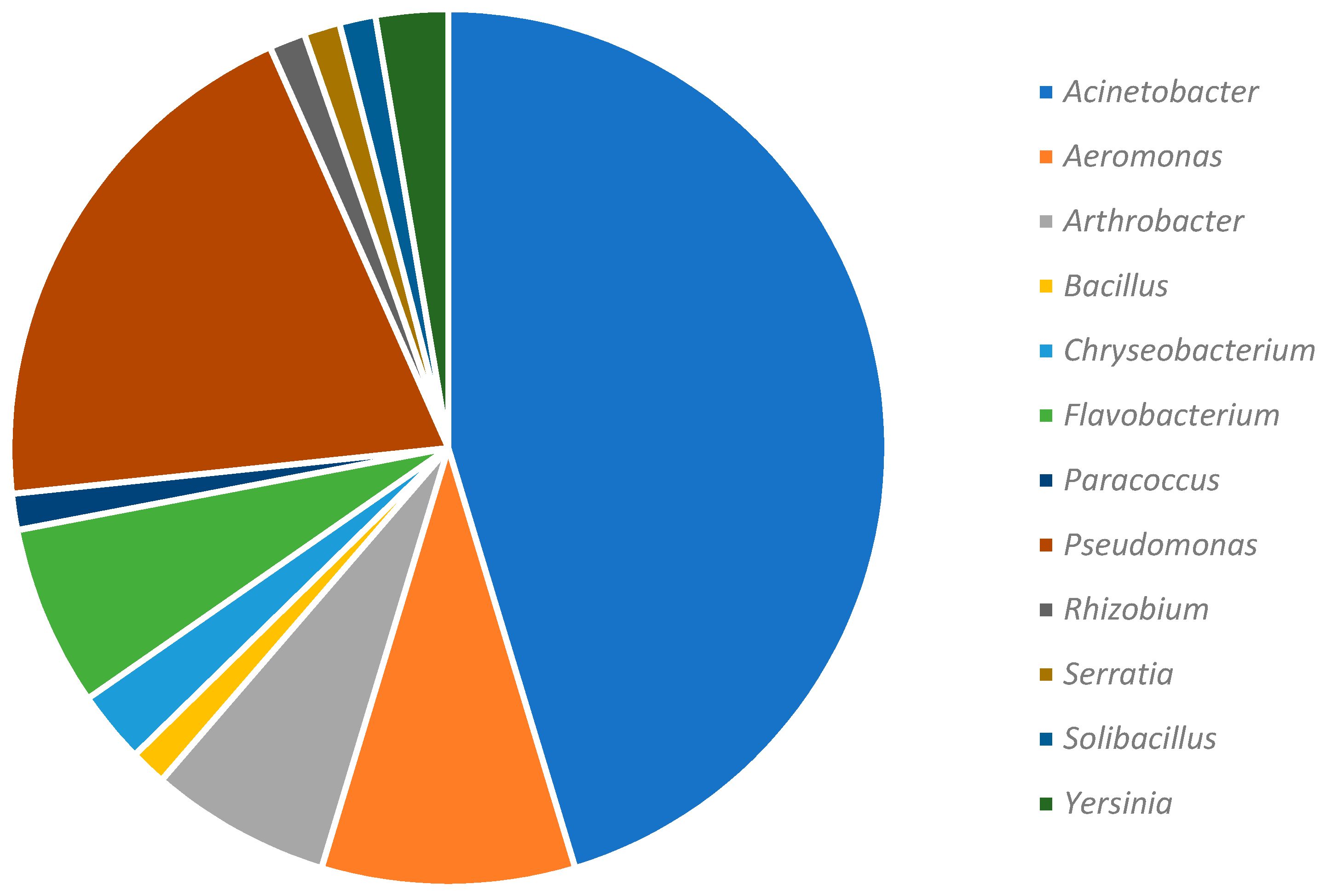
3.3. Composition of Culturable Bacterial Community Isolated from Red Epilithon
3.4. Phenotypic and Molecular Characterization of Pseudomonas and Acinetobacter Isolates
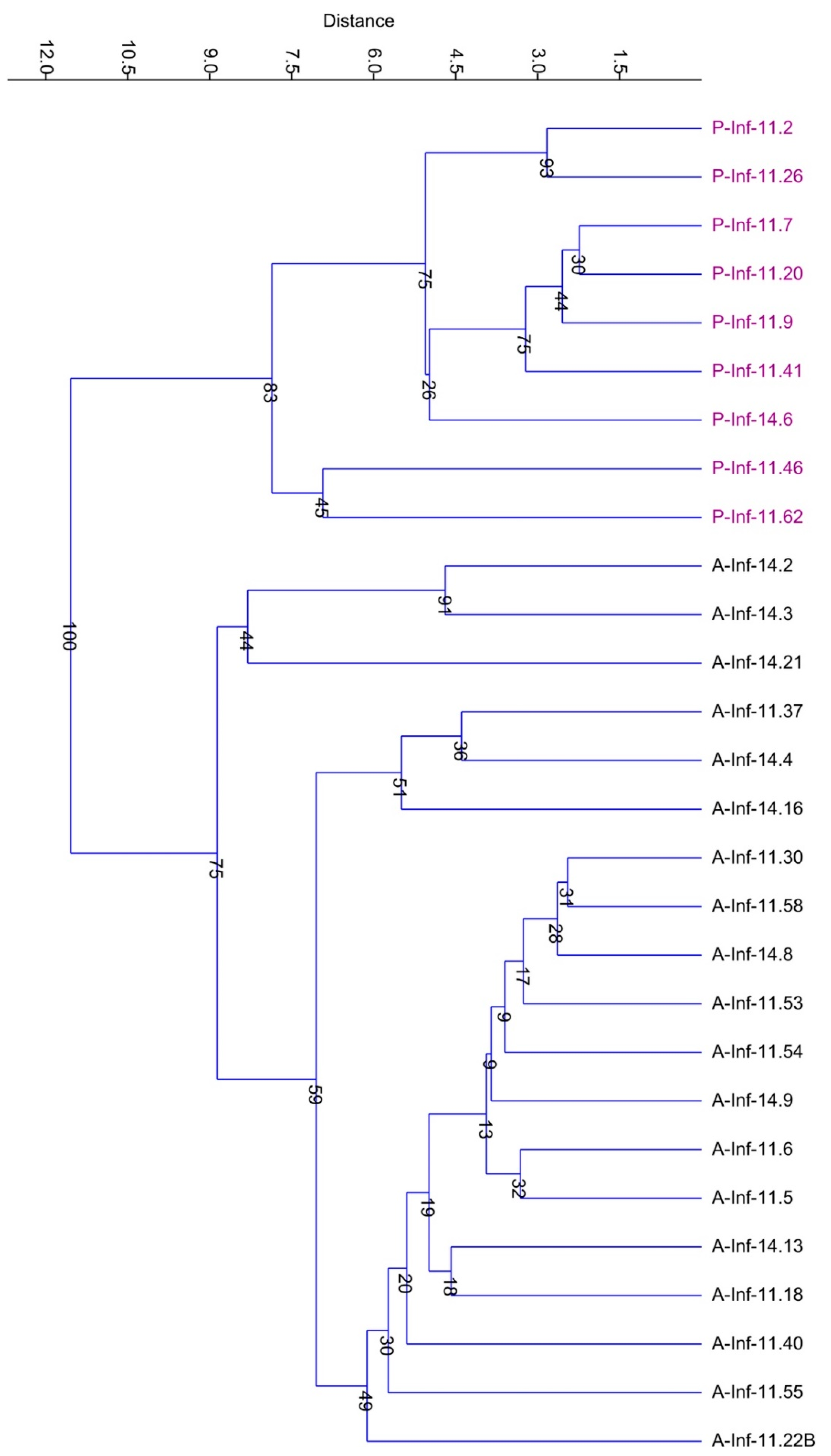
3.4.1. Structure of the Pseudomonas and Acinetobacter Communities
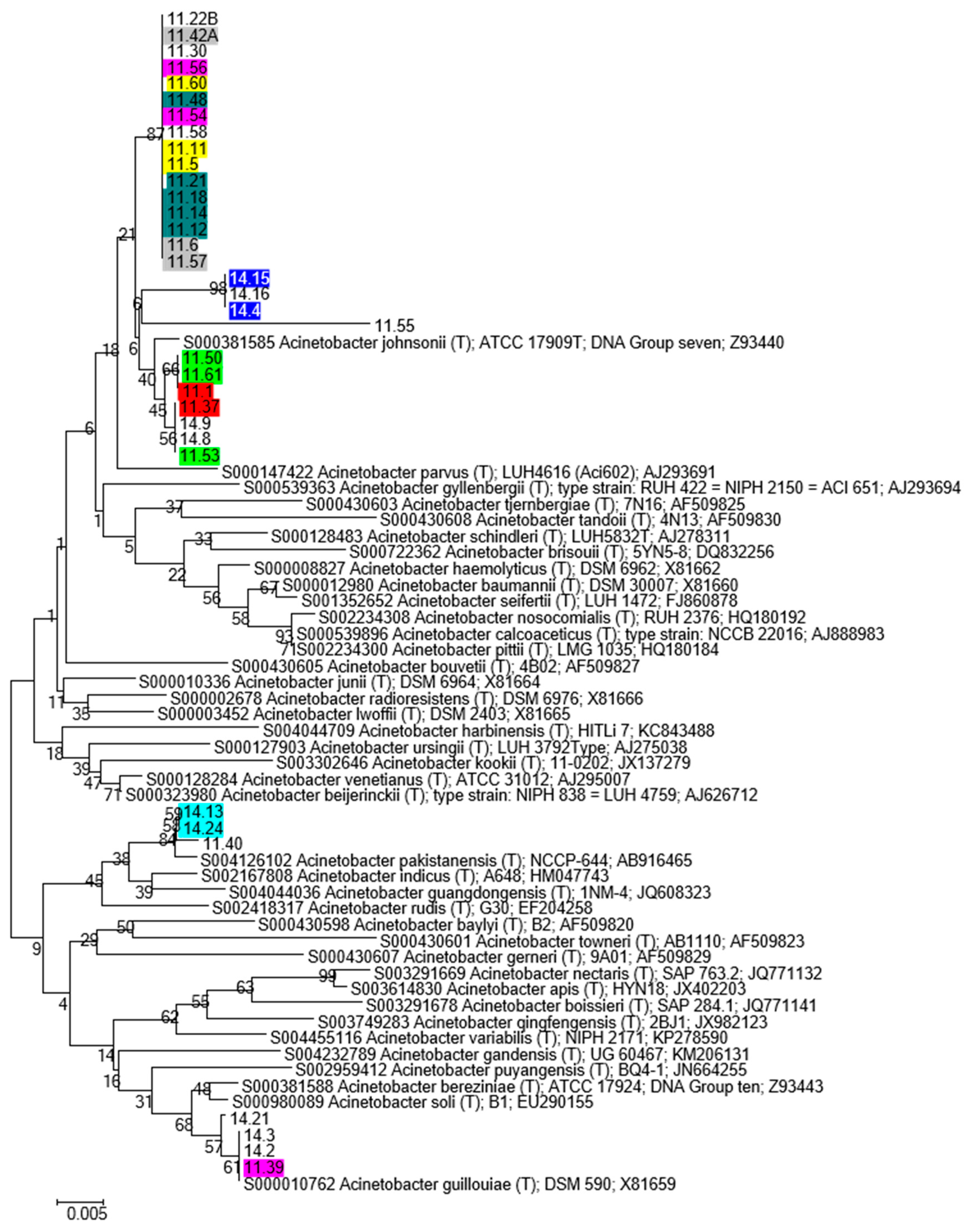
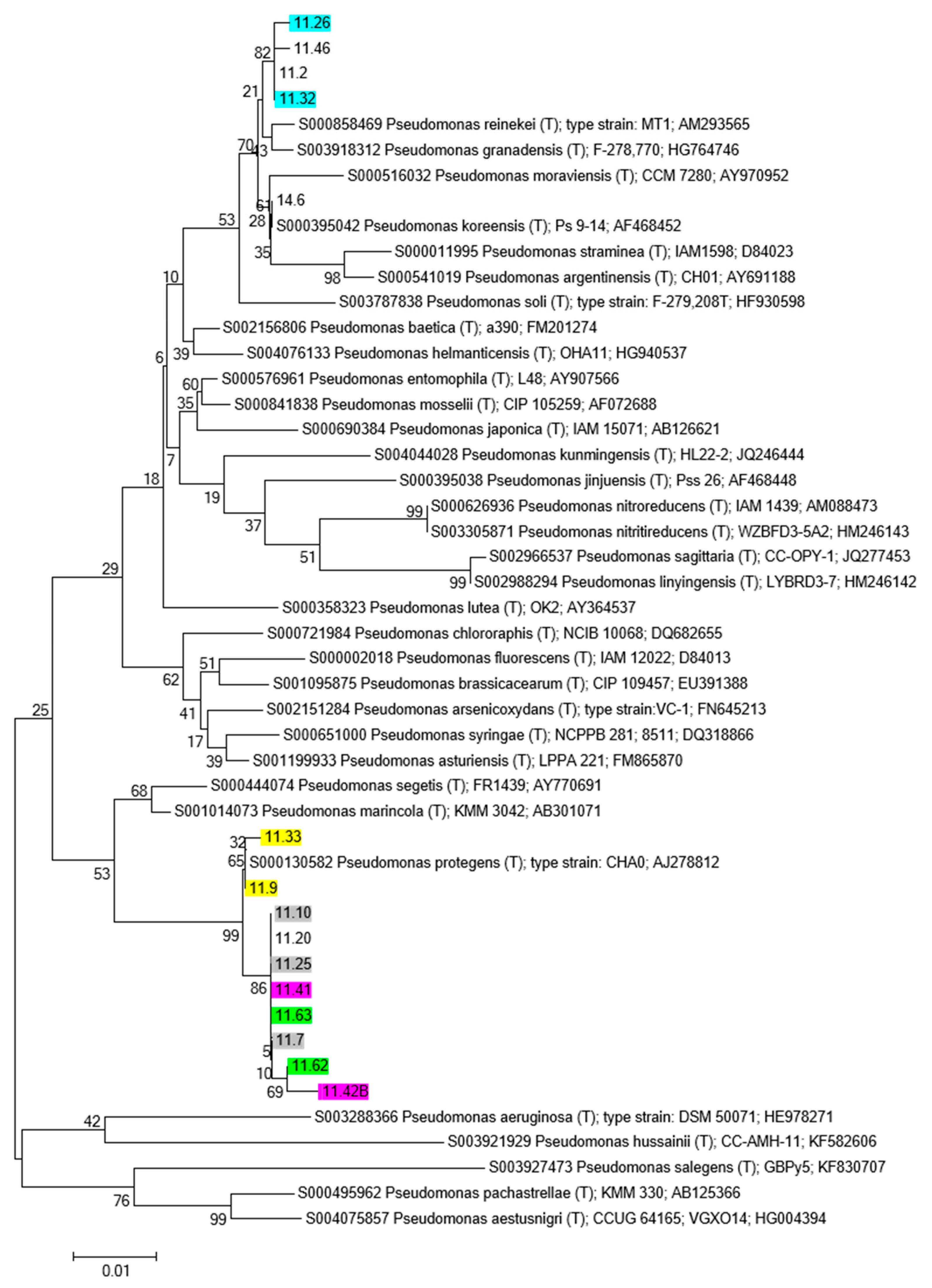
3.4.2. Phylogenetic Analysis
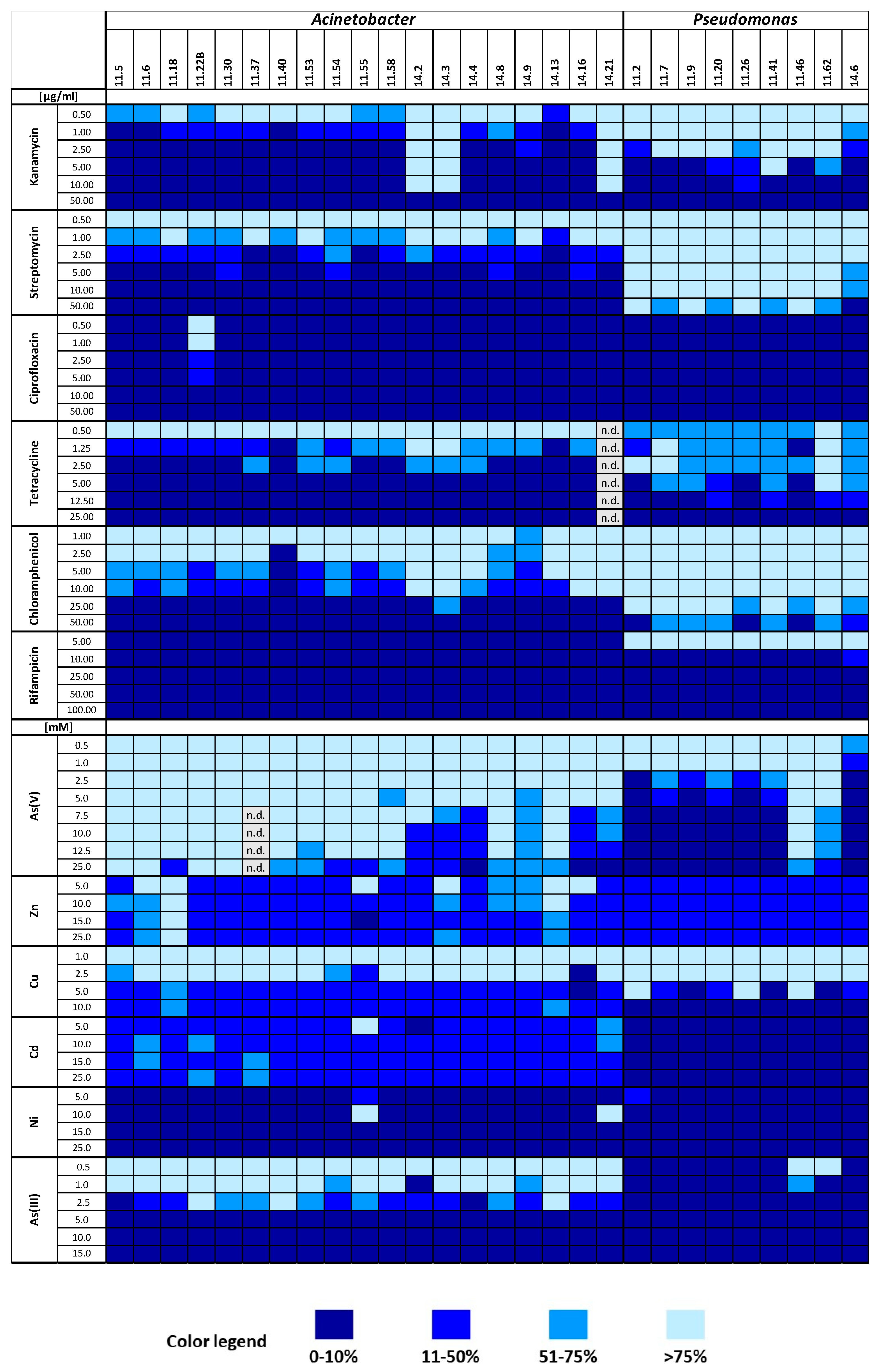
3.4.3. Phenotypic Characterization of Bacterial Strains: Antibiotic and Heavy Metal Resistance Patterns
4. Discussion
- (i)
- The major fraction of Acinetobacter isolates was located on the same branch of the phylogenetic tree (very close to A. johnsonii and A. parvus.). In this branch the isolates coming from Infernaccio and Acquarossa were intermixed.
- (ii)
- Some Acinetobacter isolates coming from Infernaccio clustered together joining different branches of the tree and were separated from the Acquarossa isolates.
- (iii)
- A similar scenario was disclosed for the Pseudomonas tree.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Harrison, A.P.; Cattani, I.; Turfa, J.M. Metallurgy, environmental pollution and the decline of Etruscan civilisation. Environ. Sci. Pollut. Res. Int. 2010, 17, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Hook, D. The composition and technology of selected Bronze Age and Early Iron Age Copper alloy artefacts from Italy. In Prehistoric Metal Artefacts from Italy (3500–720 BC) in the British Museum; Bietti Sestieri, A.M., Macnamara, E., Eds.; The British Museum: London, UK, 2007; pp. 308–323. [Google Scholar]
- Chiellini, C.; Miceli, E.; Bacci, G.; Fagorzi, C.; Coppini, E.; Fibbi, D.; Bianconi, G.; Mengoni, A.; Canganella, F.; Fani, R. Spatial structuring of bacterial communities in epilithic biofilms in the Acquarossa river (Italy). FEMS Microbiol. Ecol. 2018, 94, fiy181. [Google Scholar] [CrossRef] [PubMed]
- Papaleo, M.C.; Fondi, M.; Maida, I.; Perrin, E.; Lo Giudice, A.; Michaud, L.; Mangano, S.; Bartolucci, G.; Romoli, R.; Fani, R. Sponge-associated microbial Antarctic communities exhibiting antimicrobial activity against Burkholderia cepacia complex bacteria. Biotechnol. Adv. 2012, 30, 272–293. [Google Scholar] [CrossRef] [PubMed]
- Papaleo, M.C.; Romoli, R.; Bartolucci, G.; Maida, I.; Perrin, E.; Fondi, M.; Orlandini, V.; Mengoni, A.; Emiliani, G.; Tutino, M.L.; et al. Bioactive volatile organic compounds from Antarctic (sponges) bacteria. N. Biotechnol. 2013, 30, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Orlandini, V.; Maida, I.; Fondi, M.; Perrin, E.; Papaleo, M.C.; Bosi, E.; de Pascale, D.; Tutino, M.L.; Michaud, L.; Lo Giudice, A.; et al. Genomic analysis of three sponge-associated Arthrobacter Antarctic strains, inhibiting the growth of Burkholderia cepacia complex bacteria by synthesizing volatile organic compounds. Microbiol. Res. 2014, 169, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Maida, I.; Fondi, M.; Papaleo, M.C.; Perrin, E.; Orlandini, V.; Emiliani, G.; de Pascale, D.; Parrilli, E.; Tutino, M.L.; Michaud, L.; et al. Phenotypic and genomic characterization of the Antarctic bacterium Gillisia sp. CAL575, a producer of antimicrobial compounds. Extremophiles 2014, 18, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Fondi, M.; Orlandini, V.; Maida, I.; Perrin, E.; Papaleo, M.C.; Emiliani, G.; de Pascale, D.; Parrilli, E.; Tutino, M.L.; Michaud, L.; et al. Draft genome sequence of the volatile organic compound-producing Antarctic bacterium Arthrobacter sp. strain TB23, able to inhibit cystic fibrosis pathogens belonging to the Burkholderia cepacia complex. J. Bacteriol. 2012, 194, 6334–6335. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Muhammad, S.A.; Ahmad, S.; Hameed, A. Report: Antibiotic production by thermophilic Bacillus specie SAT-4. Pak. J. Pharm. Sci. 2009, 22, 339–345. [Google Scholar]
- Dobretsov, S.; Abed, R.M.M.; Al Maskari, S.M.S.; Al Sabahi, J.N.; Victor, R. Cyanobacterial mats from hot springs produce antimicrobial compounds and quorum-sensing inhibitors under natural conditions. J. Appl. Phycol. 2011, 23, 983–993. [Google Scholar] [CrossRef]
- Ballav, S.; Kerkar, S.; Thomas, S.; Augustine, N. Halophilic and halotolerant actinomycetes from a marine saltern of Goa, India producing anti-bacterial metabolites. J. Biosci. Bioeng. 2015, 119, 323–330. [Google Scholar] [CrossRef]
- Staccioli, R.A. Considerazioni sui complessi monumentali di Murlo e di Acquarossa. In L’Italie Préromaine Et La Rome Républicaine; École Française de Rome: Rome, Italy, 1976; Volume 27, pp. 961–972. [Google Scholar]
- Meyers, G.E. Etrusco-Italic Monumental Architectural Space from the Iron Age to the Archaic Period: An Examination of Approach and Access. Ph.D. Thesis, Faculty of the Graduate School of the University of Texas at Austin, The University of Texas at Austin, Austin, TX, USA, December 2003. [Google Scholar]
- Meyers, G.E. Approaching monumental architecture: Mechanics and movement in Archaic Etruscan palaces. Pap. Br. Sch. Rome 2013, 81, 39–66. [Google Scholar] [CrossRef]
- Chiellini, C.; Lombardo, K.; Mocali, S.; Miceli, E.; Fani, R. Pseudomonas strains isolated from different environmental niches exhibit different antagonistic ability. Ethol. Ecol. Evol. 2019. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Illumina Support Center. Available online: https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 29 July 2019).
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Bacci, G.; Bazzicalupo, M.; Benedetti, A.; Mengoni, A. StreamingTrim 1.0: A Java software for dynamic trimming of 16S rRNA sequence data from metagenetic studies. Mol. Ecol. Resour. 2014, 14, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Beckers, B.; Op De Beeck, M.; Thijs, S.; Truyens, S.; Weyens, N.; Boerjan, W.; Vangronsveld, J. Performance of 16s rDNA primer pairs in the study of rhizosphere and endosphere bacterial microbiomes in metabarcoding studies. Front. Microbiol. 2016, 7, 650. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Di Cello, F.; Fani, R. A molecular strategy for the study of natural bacterial communities by PCR-based techniques. Minerva Biotecnol. 1996, 8, 126–134. [Google Scholar]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/ NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [PubMed]
- Mori, E.; Liò, P.; Daly, S.; Damiani, G.; Perito, B.; Fani, R. Molecular nature of RAPD markers from Haemophilus influenzae Rd genome. Res. Microbiol. 1999, 150, 83–93. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Jorgensen, J.H. Antimicrobial susceptibility testing of bacteria that grow aerobically. Infect. Dis. Clin. N. Am. 1993, 7, 393–409. [Google Scholar]
- Mengoni, A.; Maida, I.; Chiellini, C.; Emiliani, G.; Mocali, S.; Fabiani, A.; Fondi, M.; Firenzuoli, F.; Fani, R. Antibiotic resistance differentiates Echinacea purpurea endophytic bacterial communities with respect to plant organs. Res. Microbiol. 2014, 165, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Stoodley, P. Developmental regulation of microbial biofilms. Curr. Opin. Biotechnol. 2002, 13, 228–233. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Infernaccio Waterfalls | Acquarossa River | |||
|---|---|---|---|---|---|---|
| Water | Red Epilithon | Water | Red Epilithon | Black Epilithon | ||
| Conductivity | µs/CM 20 °C | 598 | 843 * | |||
| COD | mg/L O2 | <10 | <10 * | |||
| P (tot) | mg/L - mg/kg | 0.2 | 2965 | 0.3 | 120 | 300 |
| Ca | mg/L - mg/kg | 46 | 112 | |||
| Al | mg/L - mg/kg | 0.29 | 1170 | 0.29 | 340 | 3813 |
| Cr (tot) | mg/L - mg/kg | <0.05 | <5 | <0.05 | <5 | <5 |
| Cr(VI) | mg/L - mg/kg | <0.005 | 0.2 | <0.005 | 0.2 | 0.2 |
| Pb | mg/L - mg/kg | <0.05 | 5 | <0.05 | <5 | 17 |
| Cd | mg/L - mg/kg | <0.01 | <1 | <0.01 * | <1 * | <1 * |
| Ni | mg/L - mg/kg | <0.05 | 8 | <0.05 * | <5 * | 54 * |
| Cu | mg/L - mg/kg | <0.05 | 10 | <0.05 * | <5 * | 17 * |
| As | mg/L - mg/kg | <0.02 | 48 | <0.02 * | 7 * | 17 * |
| Hg | mg/L - mg/kg | <0.0005 | <1 | <0.0005 | <1 | <1 |
| Tl | mg/L - mg/kg | <0.001 | <1 | <0.001 | <1 | 70 |
| Sb | mg/L - mg/kg | <0.02 | <2 | <0.02 | <2 | <2 |
| Fe tot | mg/L - mg/kg | 4.04 | 110,255 | 1.788 * | 54,314 * | 11,668 * |
| Fe2+ | mg/L | 2.17 | 0.733 * | |||
| Fe3+ | mg/L | 2.19 | 1.05 * | |||
| Mn | mg/L - mg/kg | 0.36 | 964 | 0.58 | 395 | 72,851 |
| Be | mg/L - mg/kg | <0.01 | 17 | <0.01 | 11 | 13 |
| Co | mg/L - mg/kg | <0.02 | 6 | <0.02 | <2 | 6 |
| Mo | mg/L - mg/kg | <0.01 | <2 | <0.01 | <2 | 17 |
| Se | mg/L - mg/kg | <0.04 | <1 | <0.04 | <1 | 38 |
| Sn | mg/L - mg/kg | <0.01 | <1 | <0.02 | <1 | <1 |
| V | mg/L - mg/kg | <0.05 | 15 | <0.05 | 8 | 65 |
| Ba | mg/L - mg/kg | 0.1 | 302 | <0.1 | 56 | 290 |
| B | mg/L - mg/kg | 0.24 | <5 | 0.27 | <5 | <5 |
| Zn | mg/L - mg/kg | 0.01 | 22 | <0.01 * | 14 * | 108 * |
| NH4+ | mg/L | <0.1 | 0.1 * | |||
| NO2− | mg/L | 0.7 | < 0.05 * | |||
| NO3− | mg/L | <0.02 | <1 * | |||
| SO42− | mg/L | 42 | <40 * | |||
| S2− | mg/L | <0.02 | <0.1 * | |||
| pH | 6.3 | 6.50 * | ||||
| Strain ID | Accession Number | Taxonomic Affiliation (RDP) | No of Strains | Reference |
|---|---|---|---|---|
| 11.1 | MK909965 | Acinetobacter sp. | 34 | This work |
| 11.6 | MK909969 | Acinetobacter sp. | This work | |
| 11.37 | MK909970 | Acinetobacter sp. | This work | |
| 11.18 | MK909971 | Acinetobacter sp. | This work | |
| 11.14 | MK909974 | Acinetobacter sp. | This work | |
| 11.12 | MK909976 | Acinetobacter sp. | This work | |
| 14.3 | MK909978 | Acinetobacter sp. | This work | |
| 14.21 | MK909979 | Acinetobacter sp. | This work | |
| 14.13 | MK909980 | Acinetobacter sp. | This work | |
| 14.8 | MK909983 | Acinetobacter sp. | This work | |
| 14.9 | MK909984 | Acinetobacter sp. | This work | |
| 14.4 | MK909986 | Acinetobacter sp. | This work | |
| 11.5 | MK909987 | Acinetobacter sp. | This work | |
| 14.2 | MK909989 | Acinetobacter sp. | This work | |
| 14.16 | MK909991 | Acinetobacter sp. | This work | |
| 11.21 | MK909992 | Acinetobacter sp. | This work | |
| 14.24 | MK909993 | Acinetobacter sp. | This work | |
| 14.15 | MK909994 | Acinetobacter sp. | This work | |
| 11.56 | MK909998 | Acinetobacter sp. | This work | |
| 11.60 | MK909999 | Acinetobacter sp. | This work | |
| 11.48 | MK910000 | Acinetobacter sp. | This work | |
| 11.39 | MK910001 | Acinetobacter sp. | This work | |
| 11.54 | MK910003 | Acinetobacter sp. | This work | |
| 11.58 | MK910004 | Acinetobacter sp. | This work | |
| 11.11 | MK910009 | Acinetobacter sp. | This work | |
| 11.40 | MK910010 | Acinetobacter sp. | This work | |
| 11.22b | MK910012 | Acinetobacter sp. | This work | |
| 11.53 | MK910013 | Acinetobacter sp. | This work | |
| 11.30 | MK910014 | Acinetobacter sp. | This work | |
| 11.61 | MK910019 | Acinetobacter sp. | This work | |
| 11.50 | MK910020 | Acinetobacter sp. | This work | |
| 11.57 | MK910024 | Acinetobacter sp. | This work | |
| 11.42a | MK910026 | Acinetobacter sp. | This work | |
| 11.55 | MK910027 | Acinetobacter sp. | This work | |
| 11.4 | MK909964 | Aeromonas sp. | 7 | This work |
| 11.15 | MK909973 | Aeromonas sp. | This work | |
| 14.17 | MK909990 | Aeromonas sp. | This work | |
| 11.47 | MK910005 | Aeromonas sp. | This work | |
| 11.52 | MK910016 | Aeromonas sp. | This work | |
| 11.43 | MK910017 | Aeromonas sp. | This work | |
| 11.45 | MK910023 | Aeromonas sp. | This work | |
| 14.14 | MK909985 | Arthrobacter sp. | 5 | This work |
| 11.23 | MK910007 | Arthrobacter sp. | This work | |
| 11.27 | MK910021 | Arthrobacter sp. | This work | |
| 11.49 | MK910022 | Arthrobacter sp. | This work | |
| 11.38 | MK910025 | Arthrobacter sp. | This work | |
| 14.22 | MK909995 | Bacillus sp. | 1 | This work |
| 14.11 | MK909982 | Chryseobacterium sp. | 2 | This work |
| 14.18 | MK909988 | Chryseobacterium sp. | This work | |
| 11.19 | MK909966 | Flavobacterium sp. | 5 | This work |
| 11.31 | MK909967 | Flavobacterium sp. | This work | |
| 11.51 | MK909968 | Flavobacterium sp. | This work | |
| 14.19 | MK909996 | Flavobacterium sp. | This work | |
| 11.22a | MK910015 | Flavobacterium sp. | This work | |
| 14.10 | MK910029 | Paracoccus sp. | 1 | This work |
| 11.10 | MK909977 | Pseudomonas sp. | 15 | This work |
| 11.63 | MK909997 | Pseudomonas sp. | This work | |
| 11.25 | MK910002 | Pseudomonas sp. | This work | |
| 11.33 | MK910006 | Pseudomonas sp. | This work | |
| 11.32 | MK910011 | Pseudomonas sp. | This work | |
| 11.42b | MK910028 | Pseudomonas sp. | This work | |
| 11.2 | MK272750 | Pseudomonas sp. | [3] | |
| 11.7 | MK272749 | Pseudomonas sp. | [3] | |
| 11.9 | MK272751 | Pseudomonas sp. | [3] | |
| 11.20 | MK272752 | Pseudomonas sp. | [3] | |
| 11.26 | MK272754 | Pseudomonas sp. | [3] | |
| 11.41 | MK272756 | Pseudomonas sp. | [3] | |
| 11.46 | MK272757 | Pseudomonas sp. | [3] | |
| 11.62 | MK272755 | Pseudomonas sp. | [3] | |
| 14.6 | MK272753 | Pseudomonas sp. | [3] | |
| 14.1 | MK909981 | Rhizobium sp. | 1 | This work |
| 11.13 | MK909975 | Serratia sp. | 1 | This work |
| 14.20 | MK910008 | Solibacillus sp. | 1 | This work |
| 11.17 | MK909972 | Yersinia sp. | 2 | This work |
| 11.29 | MK910018 | Yersinia sp. | This work | |
| Total | 75 | |||
| ID Haplotype | Name | Total Number of Isolates |
|---|---|---|
| 19 | 11.60; 11.11; 11.5; | 3 |
| 20 | 11.61; 11.53; 11.50 | 3 |
| 21 | 14.13; 14.24 | 2 |
| 22 | 11.22B | 1 |
| 23 | 11.30 | 1 |
| 24 | 11.55 | 1 |
| 25 | 11.40 | 1 |
| 26 | 11.39; 11.54, 11.56 | 3 |
| 29 | 14.16 | 1 |
| 30 | 14.2 | 1 |
| 31 | 14.21 | 1 |
| 37 | 11.1; 11.37 | 2 |
| 39 | 11.42A; 11.57; 11.6 | 3 |
| 40 | 11.21; 11.48; 11.14; 11.18; 11.12 | 5 |
| 41 | 11.58 | 1 |
| 42 | 14.4; 14.15 | 2 |
| 44 | 14.3 | 1 |
| 45 | 14.9 | 1 |
| 46 | 14.8 | 1 |
| Total | 34 | |
| ID Haplotype | Name | Total Number of Isolates |
|---|---|---|
| 2P | 11.41 11.42B | 2 |
| 3P | 11.9 11.33 | 2 |
| 4P | 11.62 11.63 | 2 |
| 5P | 11.26 11.32 | 2 |
| 6P | 11.7 11.10 11.25 | 3 |
| 7P | 11.2 | 1 |
| 8P | 11.20 | 1 |
| 9P | 11.46 | 1 |
| 10P | 14.6 | 1 |
| Total | 15 | |
| Index | Acinetobacter | Pseudomonas |
|---|---|---|
| Shannon H | 2.78 | 2.119 |
| Evenness | 0.848 | 0.9244 |
| Chao-1 | 32.75 | 10.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiellini, C.; Chioccioli, S.; Vassallo, A.; Mocali, S.; Miceli, E.; Fagorzi, C.; Bacci, G.; Coppini, E.; Fibbi, D.; Bianconi, G.; et al. Exploring the Bacterial Communities of Infernaccio Waterfalls: A Phenotypic and Molecular Characterization of Acinetobacter and Pseudomonas Strains Living in a Red Epilithic Biofilm. Diversity 2019, 11, 175. https://doi.org/10.3390/d11100175
Chiellini C, Chioccioli S, Vassallo A, Mocali S, Miceli E, Fagorzi C, Bacci G, Coppini E, Fibbi D, Bianconi G, et al. Exploring the Bacterial Communities of Infernaccio Waterfalls: A Phenotypic and Molecular Characterization of Acinetobacter and Pseudomonas Strains Living in a Red Epilithic Biofilm. Diversity. 2019; 11(10):175. https://doi.org/10.3390/d11100175
Chicago/Turabian StyleChiellini, Carolina, Sofia Chioccioli, Alberto Vassallo, Stefano Mocali, Elisangela Miceli, Camilla Fagorzi, Giovanni Bacci, Ester Coppini, Donatella Fibbi, Giovanna Bianconi, and et al. 2019. "Exploring the Bacterial Communities of Infernaccio Waterfalls: A Phenotypic and Molecular Characterization of Acinetobacter and Pseudomonas Strains Living in a Red Epilithic Biofilm" Diversity 11, no. 10: 175. https://doi.org/10.3390/d11100175
APA StyleChiellini, C., Chioccioli, S., Vassallo, A., Mocali, S., Miceli, E., Fagorzi, C., Bacci, G., Coppini, E., Fibbi, D., Bianconi, G., Canganella, F., & Fani, R. (2019). Exploring the Bacterial Communities of Infernaccio Waterfalls: A Phenotypic and Molecular Characterization of Acinetobacter and Pseudomonas Strains Living in a Red Epilithic Biofilm. Diversity, 11(10), 175. https://doi.org/10.3390/d11100175