Community Ecology and Phylogeography of Bats in the Guianan Savannas of Northern South America


1
Royal Ontario Museum, Toronto, ON M5S 2C6, Canada
2
Abilene Christian University, Abilene, TX 79699, USA
*
Author to whom correspondence should be addressed.
Diversity 2018, 10(4), 129; https://doi.org/10.3390/d10040129
Submission received: 20 September 2018
/
Revised: 21 November 2018
/
Accepted: 4 December 2018
/
Published: 13 December 2018
(This article belongs to the Special Issue Diversity and Conservation of Bats)
Abstract
:The Guiana Shield of South America contains savannas within one of the largest contiguous expanses of pristine tropical rainforest remaining in the world, but biodiversity in the grasslands is poorly known. In lowland Neotropical areas, bats typically comprise the most species-rich group of mammals. We compare the bat faunal community and phylogeography in the savanna habitats of the Llanos in Venezuela, Rupununi in Guyana, and Sipaliwini in Suriname. Measures of species diversity and relative abundance from standardized field survey methodology enable comparison among these three grassland regions. Genetic variation is summarized by DNA barcoding to examine biogeographic patterns across larger forest–savanna landscapes. A total of 76 species of bats is documented, of which 18 species are reported from all 3 savannas and 30 species are reported from only 1 of the savannas. Endemism is low with 5 taxa restricted primarily to dry, open habitats. However, 7 other species have divergent phylogeographic lineages associated with savanna populations. Although bat species are usually distributed over wide regions of the Neotropics, the habitat mosaics of the Guiana Shield have different faunal assemblages. Going back into the Miocene, the contractions and expansions of forest–savanna paleoenvironments over time have contributed to speciation and the current high levels of biodiversity in South America.
Keywords:
biogeography; Chiroptera; evolution; Guyana; Llanos; Neotropics; Rupununi; Sipaliwini; Suriname; Venezuela
1. Introduction
In the Guianas of northern South America, there are large areas of primarily undisturbed tropical rainforest with high levels of biodiversity [1]. This region is emerging as one of the last frontiers of relatively pristine natural habitat in the tropics, however, comparatively little is known because of the attention drawn by the larger neighboring Amazon to the south in Brazil. Smaller tracts of savannas are also present, but the biodiversity contained within these grasslands is even more poorly studied. In lowland Neotropical areas, bats typically comprise the most species-rich group of mammals and represent important contributors to forest succession and ecosystem services as seed dispersers and flower pollinators. With increased development such as all-weather roads and cattle ranching, the Guiana Shield savannas are experiencing rapid change. Although these interior habitats have been buffered by remoteness, this situation may be different in the near future.
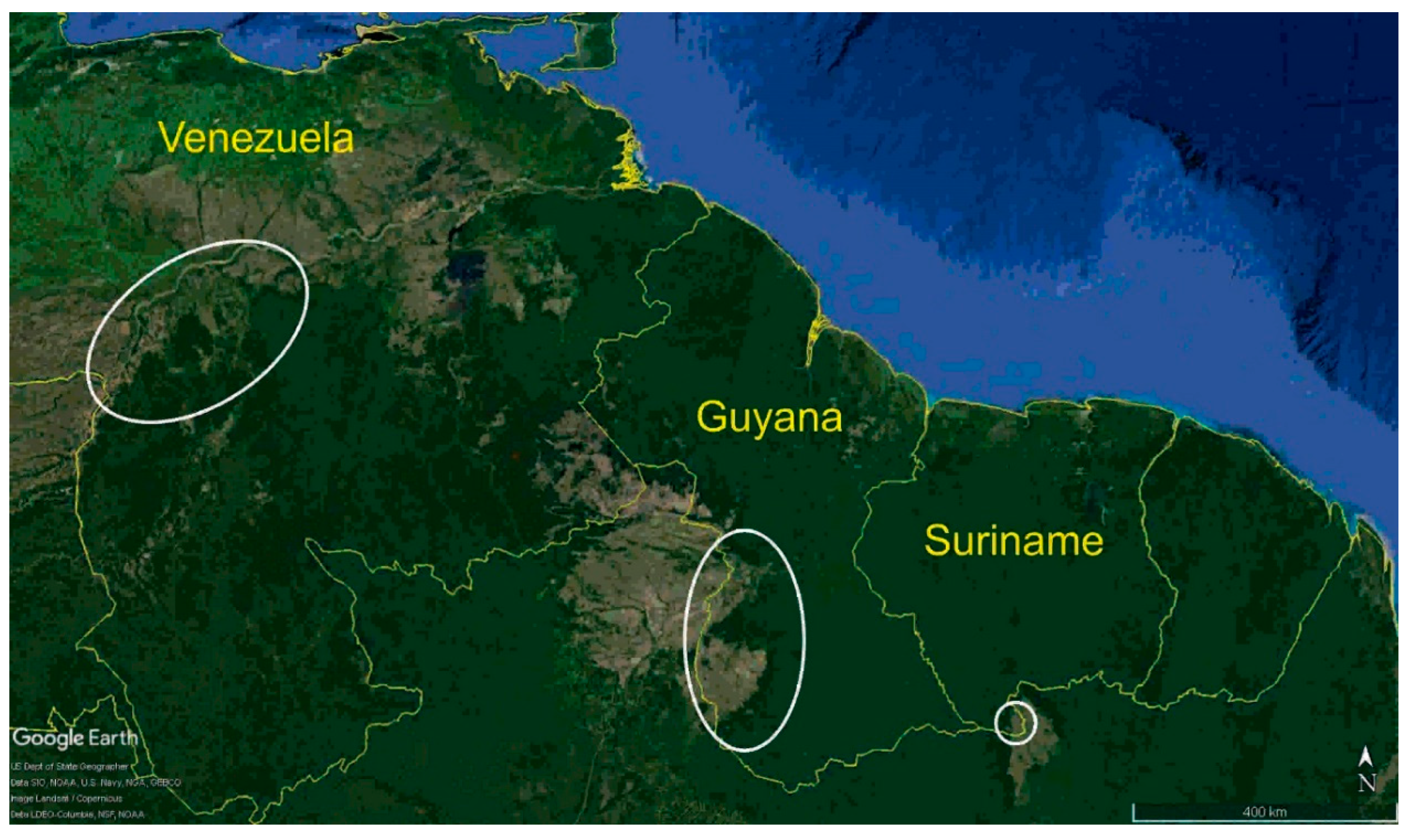
Most research on bat diversity is in forest ecosystems, but a few notable exceptions in savanna habitats are Espíritu in northeastern Bolivia [2], Cerrado in central Brazil [3], and Llanos in eastern Colombia [4]. Until recently, a neglected region of South America was the savannas of the Guianas: Llanos in Venezuela and Colombia; Rupununi–Rio Branco–Gran Sabana in Guyana, Brazil, and Venezuela; and Sipaliwini–Paru in Suriname and Brazil. The Llanos savanna covers approximately 300,000 km2 and is drained by the Orinoco River [5]. The topography is relatively flat rising from sea level to 300 m in elevation. The Llanos is developed with paved highways, towns, and a large cattle industry. The Rupununi savanna encompasses 13,000 km2 in southwestern Guyana and is bisected by the Rupununi River, which is a tributary of the Essequibo River [6]. The savanna is divided into northern and southern regions by the forested Kanuku Mountains that reach almost 1000 m above sea level. There are several indigenous villages with a population of approximately 15,000 people and several cattle ranches. A lateritic road that is prone to flooding during the rainy season connects the paved trans-Amazon highway system in Brazil with the capital city Georgetown on the Atlantic coast. At 600 km2, the Sipaliwini is the smallest of the three sites surveyed [7]. The savanna is drained by the Sipaliwini River, which is a tributary of the Corentyne River, which forms the border of Suriname and Guyana. The grassland is undulating terrain between 275–375 m above sea level with several hills reaching 550 m. There are no cattle or human settlements in the Sipaliwini, but indigenous Trio traverse the savanna to Brazil and there is irregular burning that occurs.
Based on surveys that began in 1990, we compare the species diversity and relative abundance of bat faunal communities in the under-studied grassland habitats of the Llanos in Venezuela, Rupununi in Guyana, and Sipaliwini in Suriname (Figure 1). In addition, genetic data from DNA barcoding are used to examine phylogeographic patterns in the heterogeneous forest and savanna habitats across the Guiana Shield and more broadly throughout the Neotropics. As the only flying mammals, bats are typically hypothesized to be widely distributed. We combine both biodiversity measures and molecular methods to test if there are distinctions in the biogeography and faunal structure of bats among savanna and forest populations and more specifically within grassland communities. An integrative biological approach will give insight into historical and contemporary connections that have driven the high biodiversity seen in South America.
2. Materials and Methods
The Llanos savanna of Venezuela was surveyed for bats during 15 evenings from 19 July to 4 August 1997 at 5 localities [8]. The Rupununi of Guyana was surveyed during 3 field trips: 13 evenings from 20 September to 15 October 1990 at 7 distinct localities; 19 evenings from 28 September to 19 October 2007 at 3 localities; and 11 evenings from 23 October to 4 November 2013 at 2 localities [9]. The Sipaliwini of Suriname was surveyed for 18 evenings from 14 July to 2 August 2009 at 3 localities. Surveying in the Llanos and Sipaliwini was done during similar times in July and August during the end of the rainy season, whereas surveying in the Rupununi was done during the beginning of the dry season from September to November. Although there was habitat variation among and within localities, the Llanos was primarily savanna with rocky outcrops, the Rupununi was typically savanna with gallery forest, and the Sipaliwini was clearly demarcated by savanna on the right bank of the river and forest on the left bank (Figure 2).
To survey species diversity and relative abundance, bats were caught using mist nets set in the savanna on low hills, in gallery forest, and at the edge of habitat interfaces [9]. A maximum of 18 nets were typically set in perpendicular pairs of a short (6 m) and long (12 m) net for 4 h from 1800–2200 h. Sampling was done throughout the lunar cycle because cloud coverage obfuscates the effect of moonlight and this practice was consistently done for all surveys. Representative collections were made to document the species diversity and specimens were deposited at the Royal Ontario Museum (ROM), Toronto, Canada; Abilene Christian University Natural History Collection (ACUNHC), Abilene, Texas, USA; Museo de Historia Natural La Salle (MHNLS:), Caracas, Venezuela; National Zoological Collection of Suriname (NZCS), Paramaribo, Suriname; and Centre for the Study of Biological Diversity (CSBD), Georgetown, Guyana. We followed the American Society of Mammalogist guidelines for the ethical use of wild mammals in research [10] and obtained Animal Use Protocols (AUP) from the Royal Ontario Museum and Institutional Animal Care and Use Committee (IACUC) approvals from Abilene Christian University. Research and export permits were obtained from the Environmental Protection Agency in Guyana, Nature Conservation Division in Suriname, and Ministerio del Ambiente y de los Recursos Naturales Renovables in Venezuela.
Biodiversity measures including species richness estimators, diversity indices, accumulation curves, and similarity indices were computed using EstimateS, Version 9 [11]. DNA barcoding of the mitochondrial cytochrome c oxidase subunit 1 (CO1) gene followed molecular procedures outlined by [12,13]. Phylogenetic analyses at the level of genus were done using maximum likelihood search algorithms based on the best-fit model of nucleotide substitutions and default settings including 500 bootstrap replications in MEGA6 [14]. Nodes with >70% bootstrap supports are presented in the phylogenetic trees.
3. Results
Surveying of bats in the Guianan savannas during 76 net nights resulted in 1763 individual captures representing 76 species (Table A1), of which 18 species are reported from all 3 savannas and 30 species are reported from only 1 of the savannas. The most common species was Carollia perspicillata, which was also the most frequently caught bat in all 3 areas. Another fruit-eating bat (Artibeus planirostris) was the next most common species except in the Llanos where it ranked only 8th. The 3rd most abundant species overall was Artibeus cinereus, but it was not present in the Llanos. A nectar-feeding bat (Glossophaga soricina) was the next most captured and it occurred in all 3 savannas.
3.1. Biodiversity Measures
The Rupununi had the highest number of individuals caught and species documented, but also had the most sampling effort, whereas the Llanos had the lowest numbers (Table 1). Average richness estimators indicate that the Sipaliwini is the most completely surveyed (83%) and the Llanos is the least surveyed (76%). In terms of diversity indices, all four measures were highest for the Rupununi and three of the four measures were lowest for the Sipaliwini.
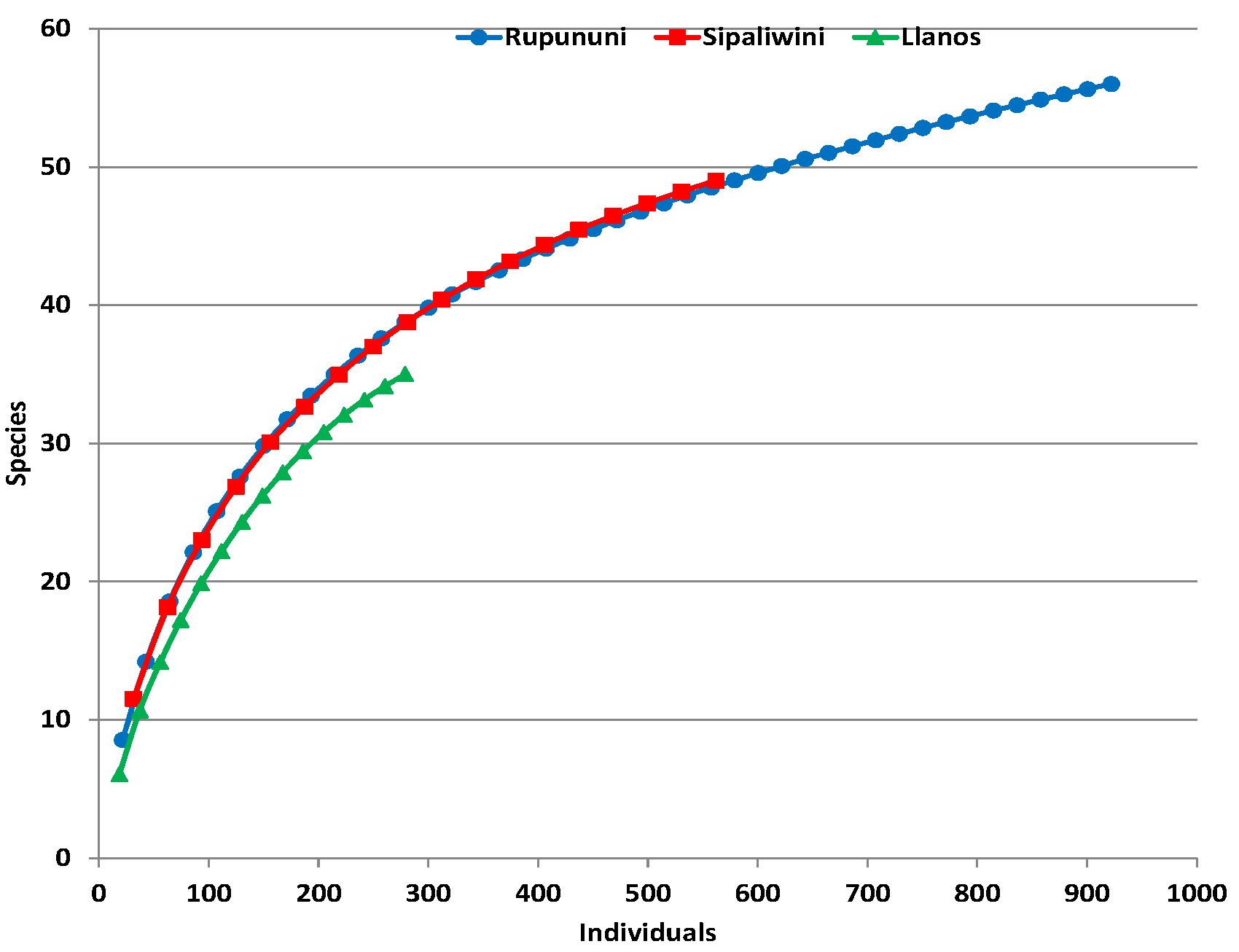
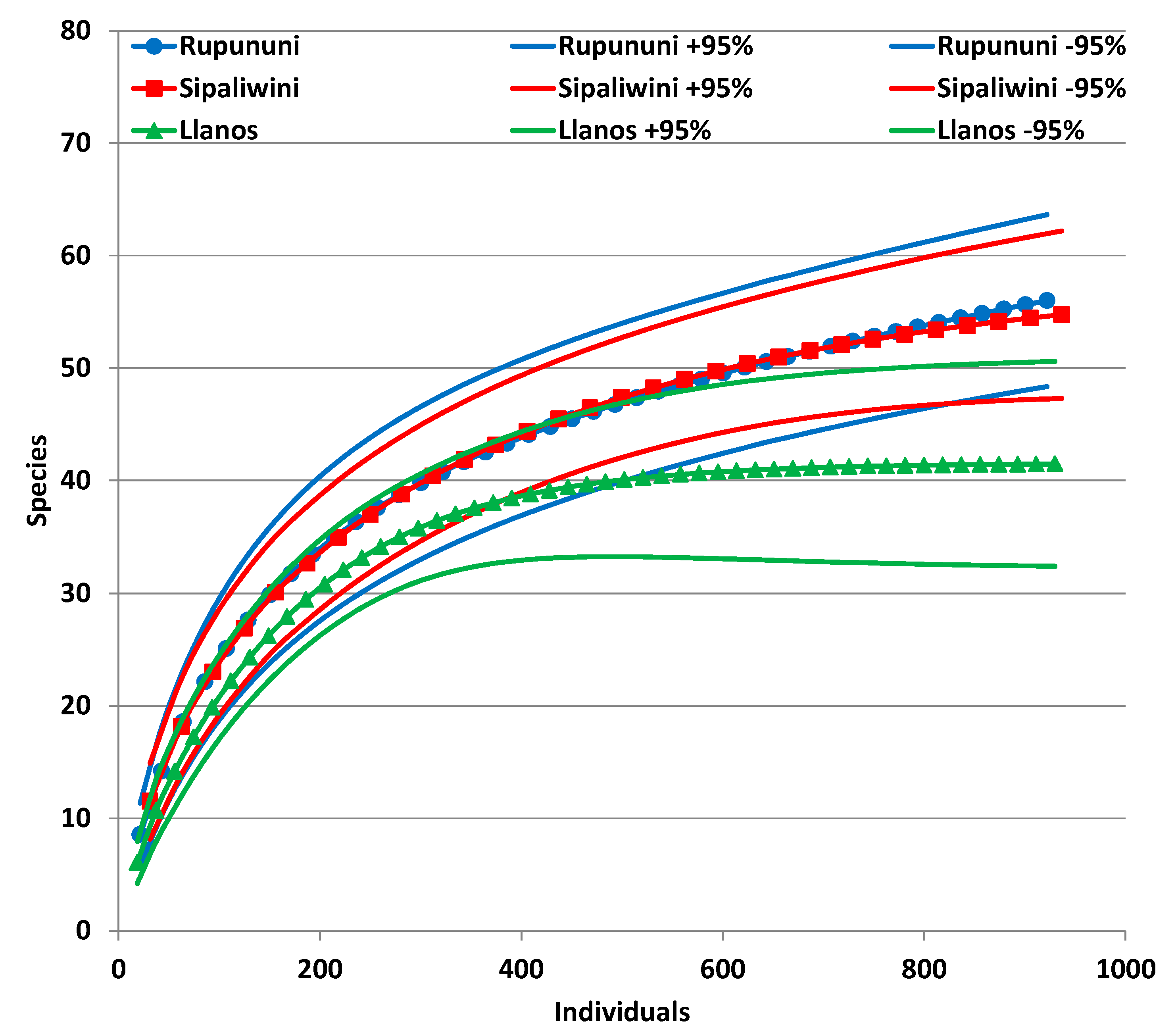
The Rupununi and Sipaliwini had similar species accumulation curve trajectories in spite of different amounts of surveying effort (Figure 3). By contrast, the species accumulation curve for the Llanos is beneath the other two savannas, indicating lower levels of species diversity and relative abundance. When it is extrapolated three times to match the effort of the Rupununi, the Llanos plateaus at 41 species. The 95% upper bound confidence interval of the Llanos is still below the other curves of the Rupununi and Sipaliwini, although there is slight overlap with their 95% lower bound confidence intervals. The curves are not leveling to an asymptote for the Rupununi and Sipaliwini, which suggests that the sampling methods were still discovering additional species at these localities. Likewise, similarity indices are highest for the comparison of Rupununi with Sipaliwini and lowest for shared species in the comparison of Sipaliwini with Llanos (Table A2).
3.2. Phylogeography
A survey of the systematic relationships based on DNA barcoding variation across the Neotropics for the 76 bats species documented from the Guianan savannas identified 11 genera with phylogeographic patterns. Molecular analyses were done on 2401 specimens with CO1 sequences (Table S1), of which 324 are newly generated for 27 species from 12 countries and are deposited in GenBank or BOLD [15].
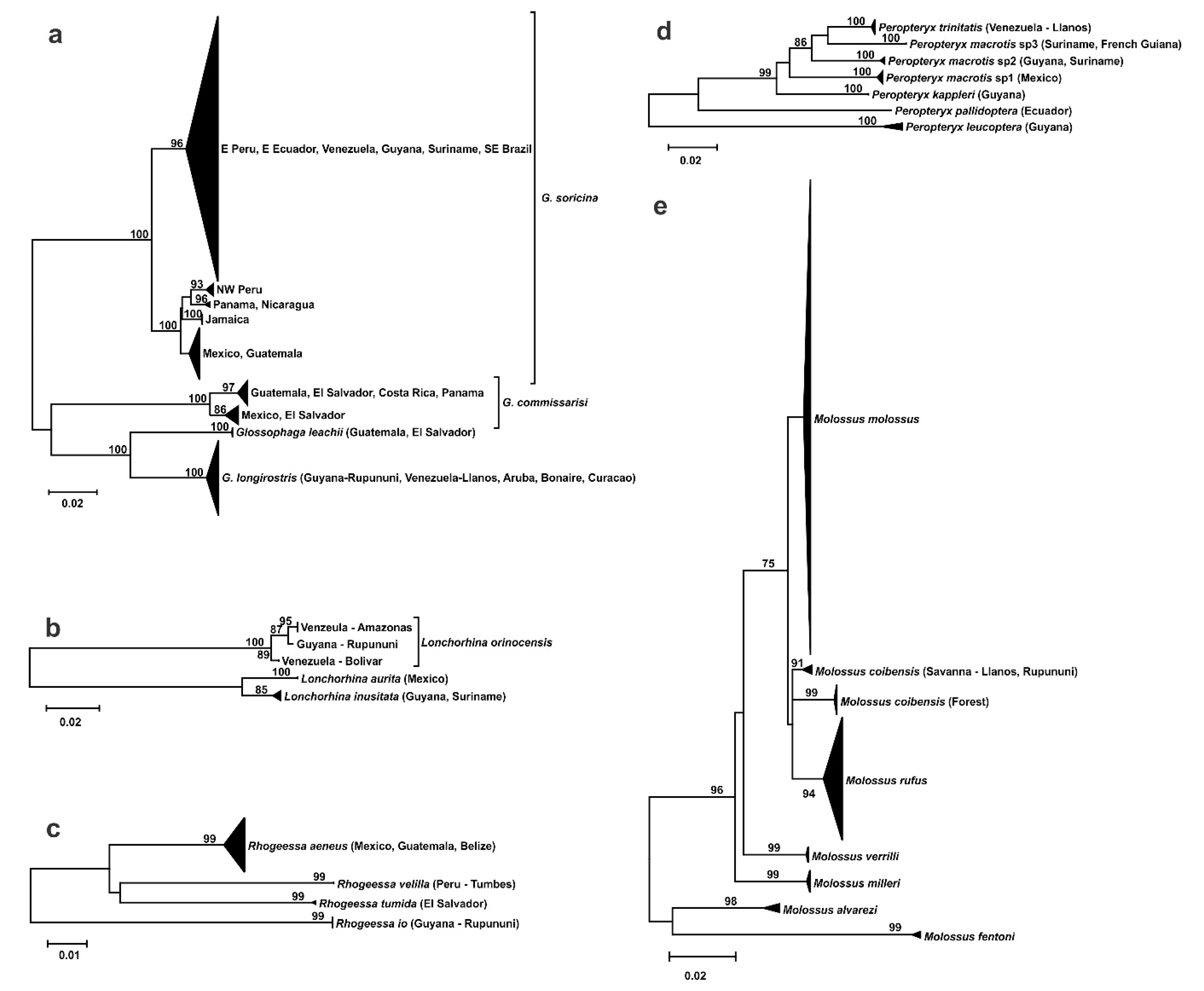
Phylogenetic trees of Neotropical bat genera found five taxa that are endemic to the savannas of the Guiana Shield (Figure 4). Two species of nectar-feeding bats in the genus Glossophaga occur in the Guianas, with Glossophaga soricina widely distributed in savanna and rainforest habitat, whereas Glossophaga longirostris is restricted to the Guyana and Venezuela savannas and the dry, open habitats of Aruba, Bonaire, and Curacao. Lonchorhina orinocensis is found in the Llanos of Venezuela and Rupununi of Guyana, and genetically distinct from two other species in the genus. Similarly, Rhogeessa io occurs in the Rupununi savanna and is divergent from three other species of Rhogeessa. The current taxonomic status within Peropteryx is complex, but it appears that P. trinitatis from the Llanos is genetically different from other taxa in the genus. There is phylogeographic structuring for unresolved, but morphologically and genetically similar populations of Molossus coibensis from Neotropical lowland forest and savannas (Llanos and Rupununi). They form a trichotomy with the larger, distinctive Molossus rufus and are closely related to the common, widespread Molossus molossus.
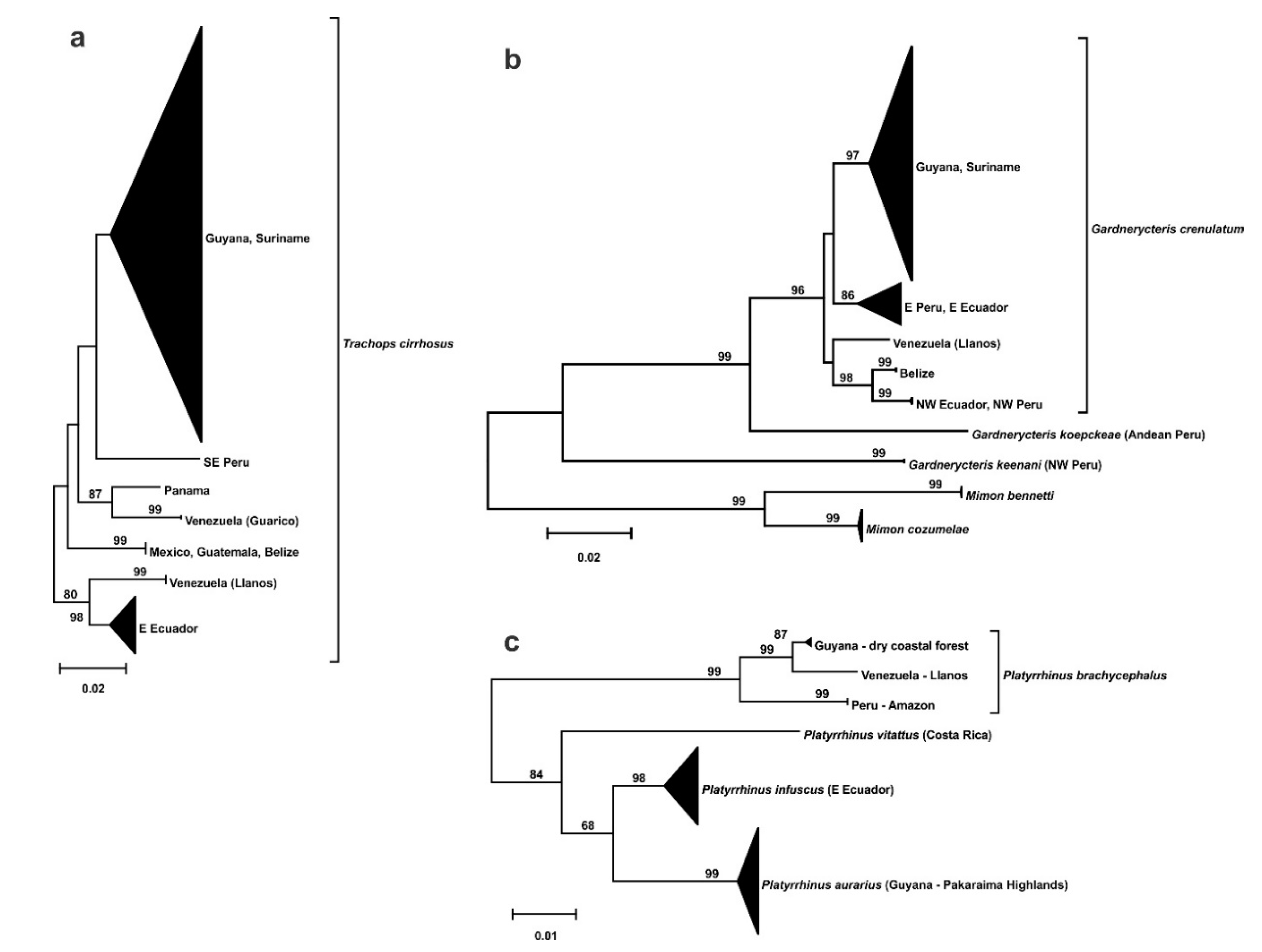
There is similar phylogeographic structuring in three species, with populations from the Llanos of Venezuela as distinct clades (Figure 5). In Trachops cirrhosus, individuals from the Venezuelan states of Amazonas and Bolivar are most closely related to Napo province in eastern Ecuador The Rupununi and Sipaliwini appear with other samples from Guyana and Suriname. Similarly, Gardnerycteris crenulatum has the Llanos population from Amazonas as a divergent lineage from the Rupununi and Sipaliwini savannas that group with others from Guyana and Suriname. Likewise, there is phylogeographic structuring in Platyrrhinus brachycephalus with the Llanos as a distinct lineage from dry coastal forest in Guyana and Amazon rainforest in Peru.
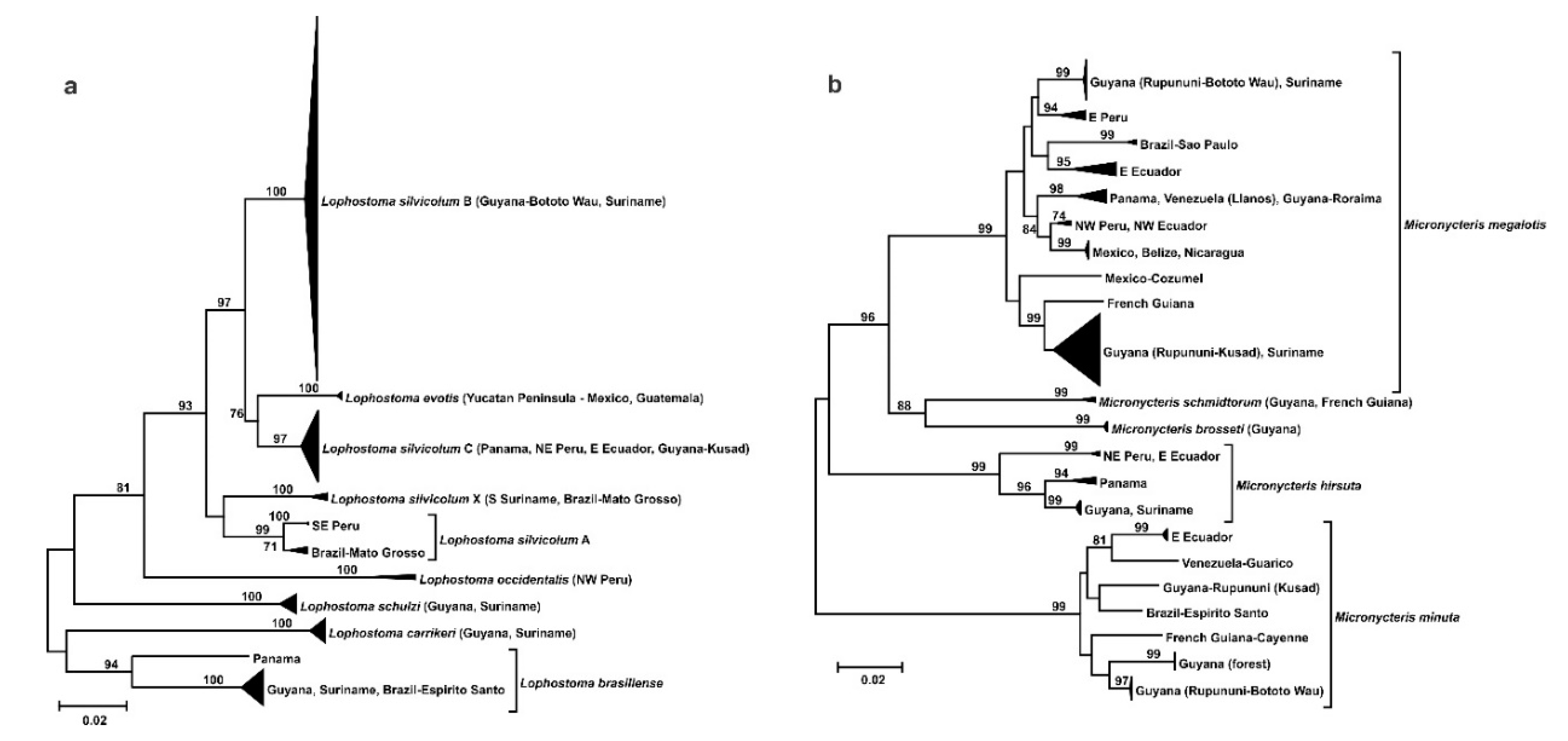
A third phylogeographic pattern has the Rupununi localities of Bototo Wau and Kusad in divergent lineages. For the Lophostoma silvicolum species complex, the Guianan savanna populations are not reciprocally monophyletic. The Guyana Rupununi specimens from Bototo Wau are in the L. silvicolum sp. B clade and not with the Guyana Rupununi specimen from Kusad, which is in the L. silvicolum sp. C clade (Figure 6a). In contrast, populations from Kusad and Bototo Wau in the south Rupununi savannas of Guyana are phylogenetically divergent in two species of Micronycteris (M. megalotis and M. minuta; Figure 6b). Furthermore, there is a third divergent savanna population of M. megalotis from the Llanos of Venezuela.
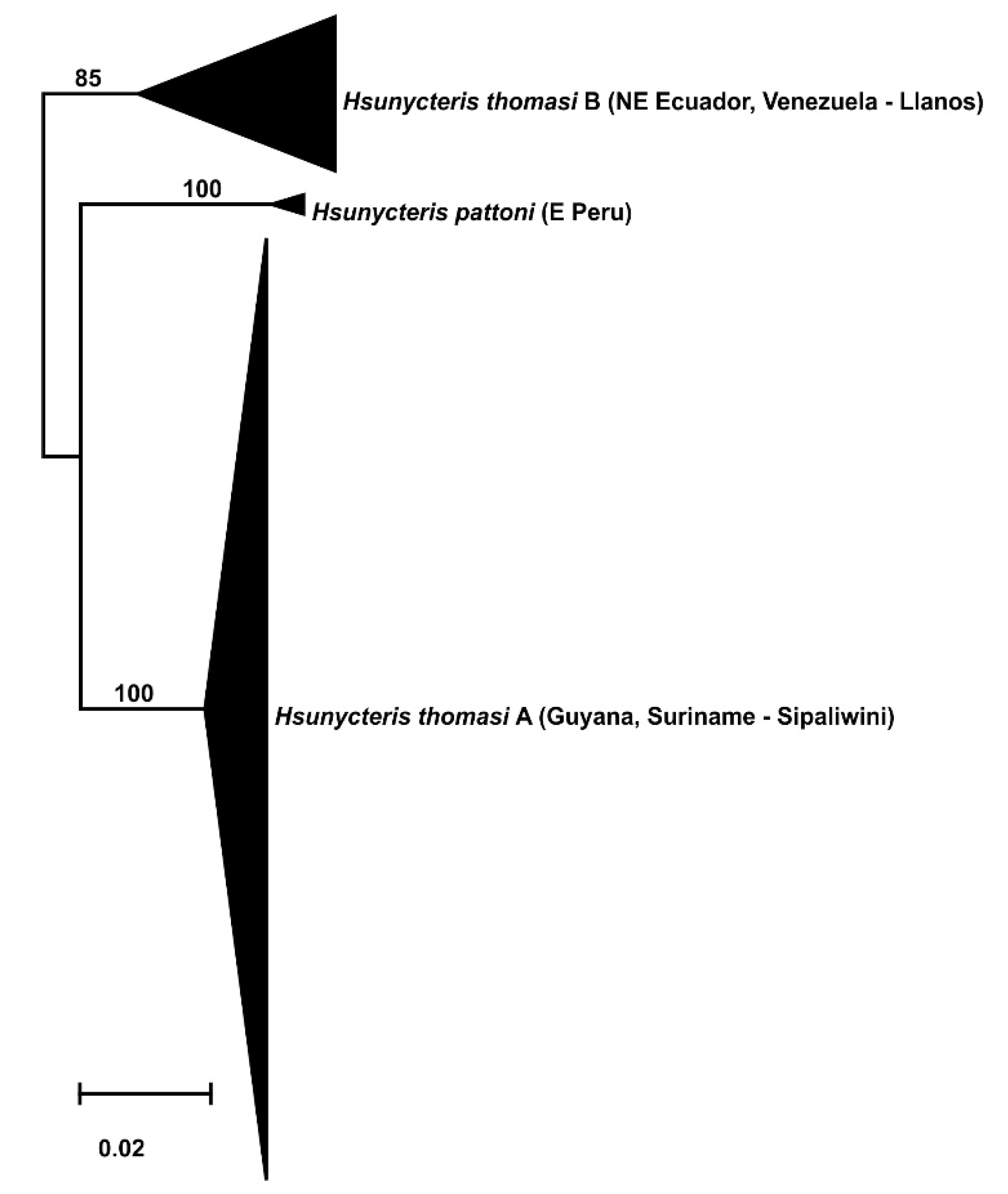
Divergence of lineages from the Llanos and Sipaliwini is another phylogeographic pattern recovered in Neotropical bats. The Hsunycteris species complex has a geographic split with Venezuela west to Ecuador that is separate from Guyana and Suriname to the east (Figure 7). Furthermore, Hsunycteris pattoni renders Hsunycteris thomasi as paraphyletic.
4. Discussion
Although the three Guianan savannas have had differing amounts of sampling effort, all had skewed community structure typical of tropical bat faunas whereby a few species were abundant and many species were rarely caught [16,17,18]. The most common species were fruit-eating bats, which are the usual trophic guild captured in ground-level nets [19,20]. Although triple-high net systems are used now, this specialized equipment was not readily available for our studies to survey the upper canopy or high above the ground. Therefore, high-flying aerial insectivorous bats were not specifically targeted in our survey methodology. In some cases, the short-tailed fruit bat (Carollia perspicillata) has been used as an indicator species of disturbed habitat in lowland tropical rainforest, if its relative abundance is high [21,22]. However, this association is not necessarily applicable to savanna habitats. In the Rupununi and Sipaliwini, C. perspicillata was more than twice as abundant as the next most common species, but based on our observations these habitats were not noticeably disturbed. The short-tailed bat feeds primarily on Piper, which is a common pioneering shrub found in open areas, including both human-induced clearings in rainforest and natural forest islands in savannas [23].
In spite of more nights of surveying, all species diversity indices indicate that the Rupununi was the most diverse of the Guianan savannas. Furthermore, species accumulation curves and analysis of shared species suggest that the Sipaliwini was most similar to the Rupununi and the Llanos was the most different in terms of species diversity and relative abundance. Furthermore, despite sampling at the end of the rainy season for the Llanos and Sipaliwini, Sipaliwini is more similar to the Rupununi, which was sampled at the beginning of the dry season, suggesting that geography and not seasonality had more of an impact on community structure. Geographically, the Rupununi and Sipaliwini are in closer proximity (400 km) to each other and surrounded by lowland rainforest. By contrast, the Llanos is 800 km to the northwest on the periphery of rainforest and the other side of the topological Guiana Highland plateau, which may be acting as a geographic barrier to gene flow. This biogeographic distinction on either side of the highland barrier has not been reported for other organisms, but the Orinoco basin is of relatively recent geological origin in the Pleistocene from Guianan and Andean sediments [24]. The Llanos is also the most developed of the three sites with access by a paved all-season road, whereas the Rupununi has a lateritic road that floods during the rainy season and the Sipaliwini is only accessible by river in terms of ground transportation. Therefore, the landscapes in each of the three savannas may have changed differentially during the three decades of this study, which would benefit from regular monitoring.
Of the 76 species of bats documented in the savanna habitats of the Guianas, 12 species have phylogeography patterns indicating varying levels of genetic divergence. There are five taxa that are essentially endemic to the Guianan savannas or other dry, open habitats in the Neotropics. Lonchorhina orinocensis was previously thought to occur only in the Llanos of Venezuela [25] and Colombia [26], but was recently found in the Rupununi of Guyana [6] (Figure 8). Peropteryx has several undescribed cryptic species [27], which suggests that P. trinitatis may be primarily (75%) restricted to dry, open habitats (Handley, 1976), such as the Llanos savanna, and coastal areas. Similarly, Glossophaga longirostris occurs primarily (92%) in thorn forest and savanna of northern South America and off-shore islands [28,29]. Seven subspecies have been recognized in G. longirostris [30], but our genetic analysis did not recover any phylogeographic structuring with specimens from Aruba, Bonaire, Curacao, Guyana, and Venezuela. Our geographic sampling includes G. l. campestris, G. l. elongata, and G. l. major, which suggests that subspecies designations need to be re-evaluated. In the genus Molossus, populations from the Llanos and Rupununi form a reciprocally monophyletic clade that is genetically most similar to M. coibensis and distinct from the more common, broadly distributed species M. molossus, as recently detailed by [31]. Although Rhogeessa io is considered widely distributed from Nicaragua to Brazil [32], the taxonomic status of this species, especially populations in Central America, and the genus in general are in need of systematic revision [33]. The species boundaries in South America are poorly known and documented by few specimens, but DNA barcoding variation indicates that R. io from the Rupununi savanna is quite divergent from other species in the genus.
There are three species with structuring that identifies the Venezuelan Llanos as a divergent lineage. In Trachops cirrhosis, there are at least seven distinct lineages comprising individuals from the Rupununi and Sipaliwini in a broadly sympatric clade with other samples from Guyana and Suriname that are separate from Venezuela, as reported earlier by [34]. Gardnerycteris crenulatum has a similar pattern with the Venezuelan Llanos in a distinct lineage from the Rupununi and Sipaliwini, which are in a more widely distributed clade of Guyana and Suriname specimens. For Platyrrhinus brachycephalus, there are no samples from the Rupununi or Sipaliwini, but the Llanos is a separate lineage from the dry coastal forest of Guyana and the Amazon rainforest of Peru. Based on geography, the Llanos of Venezuela and Colombia is separated by the Guiana highland plateau from the contiguous lowland forest and savanna in Guyana and Suriname, which suggests a potential barrier to gene flow across northern South America.
A more complex phylogeographic pattern is seen in Micronycteris megalotis, as also reported by [34], with three Guianan savanna populations separately grouping with more broadly distributed clades. The Venezuelan Llanos is in a lineage with Panama and the highlands of Guyana, whereas the Rupununi (Bototo Wau and Kusad) is split into two lineages with other samples from Guyana and Suriname. Divergent populations from Bototo Wau and Kusad are also recovered in Micronycteris minuta and Lophostoma silvicolum. However, the splits in Lophostoma silvicolum correspond to undescribed cryptic species B and C, respectively [35]. In addition, there are other taxonomic issues that need further study, including the status of Lophostoma evotis and another potentially cryptic species Lophostoma silvicolum X. The distinction in the southern Rupununi savanna may be attributed to more recent historical ecological differences during Pleistocene climate fluctuations. Kusad is a forested mountain isolated in savanna, whereas Bototo Wau is a patch of savanna isolated in forest [9].
A final pattern recovered is a split of Hsunycteris thomasi from the Llanos and Sipaliwini in two more broadly distributed clades including Venezuela west to Ecuador and Guyana east to Suriname. One of these lineages was considered an undescribed cryptic species, but the holotype from Ciudad Bolivar in Venezuela could not be unambiguously applied to identify which one corresponds to H. thomasi and which one requires a new taxonomic name [36]. However, our sample (ROM 107906) from Hato La Cerbatana, 20 km S of Guaniamo, Bolivar, Venezuela is in similar lowland Llanos habitat as the type locality, suggesting that the nominal name applies to the population from western Bolivar in Venezuela to Ecuador. If this assumption is correct, the intervening Guiana Highlands is acting as a geographic barrier that separates it from the population of eastern Bolivar in Venezuela, Guyana and Suriname, as previously found in a cytochrome b study by [37].
On an older geological time-scale beyond species-level phylogeography, a biogeographic study of New World sheath-tailed bats [38] coupled with a fossil-calibrated molecular phylogenetic dating analysis [39] hypothesized that most genera in the tribe Diclidurini evolved during the Early Miocene in the northern Amazon with the Guiana Shield acting as a stable core area of biotic diversification. At this time approximately 19 million years ago, the paleoenvironments were drier and more open habitats than today with the faunal community dominated by now-extinct large mammalian herbivores [40]. Tectonic activity combined with temperature fluctuations and sea level changes continued to influence the ecosystems into the Late Miocene and Pliocene when most of the contemporary species differentiated during repeated episodes of range expansion and contractions in the heterogeneous rainforest and savanna habitats of South America. However, none of the emballonurid bat species exhibited geographic structuring in the Guianas.
From a different perspective of relative abundance for the three species of small-sized Artibeus occurring in the Guianas (A. bogotensis, A. cinereus, and A. gnomus), an earlier study found that A. cinereus was overwhelmingly the most common in the Rupununi savanna of Guyana [41]. With our expanded study, the pattern is not as prominent in Suriname, however, this may be attributable to the abrupt transition from savanna to rainforest along the Sipaliwini River as opposed to the more typical gallery forest in Guyana. We did not catch A. cinereus in the Llanos, but this is near the western extent of its distribution [28,42]. Although not as obvious as the presence of endemic species to savannas, such as Lonchorhina orinocensis and Glossophaga longirostris, Artibeus cinereus is a more subtle indicator species of drier, open habitats in the Neotropics.
5. Conclusions
The Rupununi of Guyana is the most diverse for bats in the savannas of the Guianas and is more similar to the Sipaliwini of Suriname than it is to the Llanos of Venezuela, which is congruent with geography and proximity. In terms of genetic divergence and other than widely distributed across the Guianas, there are three general patterns of phylogeography detected for bats in the Guianan savannas in comparison to throughout the Neotropics:
- Endemism in the savannas of the Guianas at the species or subspecies level.
- Divergent population from the Llanos of Venezuela.
- Split within the southern Rupununi of Guyana between Kusad and Bototo Wau.
Biogeographically, phylogenetic signatures suggest that the evolution of bats in the Guianas began in the Early Miocene and continued into the Pleistocene with differing degrees of lineage divergence. Although the levels of endemism are low because of the dispersal abilities of bats, the Guiana highland plateau has acted as a geographic barrier to gene flow for some species, as has the fluctuating heterogeneous paleoenvironments of savanna and rainforest habitats. Future research should focus on including the savannas in Amapa state, Brazil, and to increase the surveying in the Llanos and Sipaliwini. On a broader scale, more comprehensive taxonomic and geographic sampling throughout South America will result in better resolved species-level phylogenies, which will allow for more robust interpretations of the biogeography and evolution of bats in the Neotropics.
Supplementary Materials
The following is available online at https://www.mdpi.com/1424-2818/10/4/129/s1, Table S1: Specimens used in the DNA analysis of the mitochondrial gene cytochrome c oxidase subunit 1 for species of bats that occur in the Guiana savannas of northern South America and their close relatives.
Author Contributions
B.K.L. conceived, designed, and performed the experiments; B.K.L. analyzed the data; B.K.L. and T.E.L.J. contributed reagents/materials/analysis tools; and B.K.L. and T.E.L.J. wrote the paper.
Funding
Field work was funded by grants from the Abilene Christian University Math/Science Research Council to T.E.L. and the Royal Ontario Museum Governors, Youth Challenge International, World Wildlife Fund, and Global Wildlife Conservation to B.K.L.
Acknowledgments
We thank John Hanson for field assistance in Venezuela, Lina Arcila and Mark Engstrom for field assistance in Suriname, and Diana Singh, Regis James, Zacharias Norman, Chetwynd Osborne, Abraham Ignace, Mark Engstrom, and Fiona Ried for field assistance in Guyana. Sequencing was done by the Canadian Centre for DNA Barcoding at the University of Guelph and the Laboratory of Molecular Systematics at the Royal Ontario Museum.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Bat species documented in the Guianan savannas of northern South America and their relative abundance in the Llanos of Venezuela, Rupununi of Guyana, and Sipaliwini of Suriname.
Table A1.
Bat species documented in the Guianan savannas of northern South America and their relative abundance in the Llanos of Venezuela, Rupununi of Guyana, and Sipaliwini of Suriname.
| Species | Llanos | Rupununi | Sipaliwini | Total |
|---|---|---|---|---|
| Carollia perspicillata | 60 | 239 | 201 | 500 |
| Artibeus planirostris | 7 | 103 | 77 | 187 |
| Artibeus cinereus | 0 | 94 | 9 | 103 |
| Glossophaga soricina | 6 | 51 | 22 | 79 |
| Artibeus lituratus | 0 | 30 | 46 | 76 |
| Glossophaga longirostris | 36 | 34 | 0 | 70 |
| Trachops cirrhosus | 13 | 32 | 1 | 46 |
| Peropteryx trinitatis | 46 | 0 | 0 | 46 |
| Desmodus rotundus | 5 | 36 | 2 | 43 |
| Sturnira lilium | 3 | 31 | 8 | 42 |
| Artibeus obscurus | 5 | 7 | 20 | 32 |
| Phyllostomus discolor | 2 | 19 | 9 | 30 |
| Ametrida centurio | 1 | 10 | 18 | 29 |
| Artibeus bogotensis | 2 | 17 | 8 | 27 |
| Phyllostomus elongatus | 3 | 17 | 6 | 26 |
| Pteronotus fuscus | 26 | 0 | 0 | 26 |
| Platyrrhinus lineatus | 0 | 0 | 22 | 22 |
| Lonchorhina orinocensis | 20 | 1 | 0 | 21 |
| Molossus molossus | 0 | 12 | 8 | 20 |
| Noctilio albiventris | 0 | 20 | 0 | 20 |
| Rhinophylla pumilio | 0 | 3 | 15 | 18 |
| Saccopteryx bilineata | 1 | 14 | 1 | 16 |
| Artibeus concolor | 1 | 8 | 7 | 16 |
| Lophostoma silvicolum | 0 | 12 | 4 | 16 |
| Phyllostomus hastatus | 10 | 6 | 0 | 16 |
| Myotis nigricans | 0 | 14 | 0 | 14 |
| Pteronotus rubiginosus | 0 | 13 | 1 | 14 |
| Myotis albescens | 0 | 10 | 3 | 13 |
| Rhynchonycteris naso | 2 | 10 | 0 | 12 |
| Gardnerycteris crenulatum | 2 | 7 | 2 | 11 |
| Micronycteris megalotis | 5 | 5 | 0 | 10 |
| Hsunycteris thomasi | 2 | 0 | 8 | 10 |
| Phylloderma stenops | 1 | 3 | 5 | 9 |
| Rhogeessa io | 1 | 8 | 0 | 9 |
| Chiroderma villosum | 1 | 2 | 5 | 8 |
| Carollia brevicauda | 0 | 5 | 2 | 7 |
| Molossus coibensis | 6 | 1 | 0 | 7 |
| Mesophylla macconnelli | 0 | 5 | 2 | 7 |
| Artibeus gnomus | 1 | 1 | 5 | 7 |
| Micronycteris minuta | 1 | 5 | 0 | 6 |
| Eumops auripendulus | 0 | 6 | 0 | 6 |
| Eptesicus furinalis | 0 | 3 | 2 | 5 |
| Trinycteris nicefori | 1 | 3 | 1 | 5 |
| Vampyriscus bidens | 1 | 0 | 4 | 5 |
| Uroderma bilobatum | 0 | 4 | 0 | 4 |
| Platyrrhinus guianensis | 0 | 2 | 2 | 4 |
| Platyrrhinus incarum | 0 | 1 | 3 | 4 |
| Vampyriscus brocki | 0 | 0 | 4 | 4 |
| Pteronotus alitonos | 0 | 0 | 4 | 4 |
| Sturnira tildae | 0 | 0 | 4 | 4 |
| Saccopteryx canescens | 2 | 1 | 0 | 3 |
| Saccopteryx leptura | 0 | 2 | 1 | 3 |
| Lophostoma brasiliense | 0 | 2 | 1 | 3 |
| Choeroniscus minor | 0 | 2 | 1 | 3 |
| Chrotopterus auritus | 0 | 0 | 3 | 3 |
| Chiroderma trinitatum | 0 | 0 | 3 | 3 |
| Anoura geoffroyi | 1 | 0 | 2 | 3 |
| Pteronotus personatus | 0 | 2 | 0 | 2 |
| Cormura brevirostris | 0 | 1 | 1 | 2 |
| Lionycteris spurrelli | 0 | 0 | 2 | 2 |
| Myotis riparius | 0 | 0 | 2 | 2 |
| Platyrrhinus fusciventris | 0 | 0 | 2 | 2 |
| Artibeus amplus | 2 | 0 | 0 | 2 |
| Platyrrhinus brachycephalus | 2 | 0 | 0 | 2 |
| Cynomops planirostris | 0 | 1 | 0 | 1 |
| Lasiurus blossevillii | 0 | 1 | 0 | 1 |
| Noctilio leporinus | 0 | 1 | 0 | 1 |
| Micronycteris schmidtorum | 0 | 1 | 0 | 1 |
| Natalus tumidirostris | 0 | 1 | 0 | 1 |
| Glyphonycteris sylvestris | 0 | 1 | 0 | 1 |
| Uroderma magnirostrum | 0 | 1 | 0 | 1 |
| Molossus fentoni | 0 | 1 | 0 | 1 |
| Lophostoma carrikeri | 0 | 0 | 1 | 1 |
| Diaemus youngi | 0 | 0 | 1 | 1 |
| Glyphonycteris daviesi | 0 | 0 | 1 | 1 |
| Pteronotus gymnonotus | 1 | 0 | 0 | 1 |
| Total | 279 | 922 | 562 | 1763 |
Table A2.
Shared species between the savannas of the Rupununi of Guyana, Sipaliwini or Suriname, and Llanos of Venezuela. Highest values are in bold, which indicated that the Rupununi and Sipaliwini are more similar to each other than to the Llanos.
Table A2.
Shared species between the savannas of the Rupununi of Guyana, Sipaliwini or Suriname, and Llanos of Venezuela. Highest values are in bold, which indicated that the Rupununi and Sipaliwini are more similar to each other than to the Llanos.
| First Sample | Second Sample | Shared Species | Jaccard Classic | Sorensen Classic | Chao-Jaccard-Raw | Chao-Jaccard-Est | Chao-Sorensen-Raw | Chao-Sorensen-Est | Morisita-Horn | Bray-Curtis |
|---|---|---|---|---|---|---|---|---|---|---|
| Rupununi | Sipaliwini | 34 | 0.479 | 0.648 | 0.779 | 0.824 | 0.876 | 0.903 | 0.886 | 0.592 |
| Rupununi | Llanos | 27 | 0.422 | 0.593 | 0.561 | 0.729 | 0.719 | 0.843 | 0.642 | 0.278 |
| Sipaliwini | Llanos | 21 | 0.333 | 0.5 | 0.369 | 0.415 | 0.539 | 0.586 | 0.608 | 0.247 |
References
- Mittermeier, R.A.; Myers, N.; Thomsen, J.B.; da Fonseca, G.A.B.; Olivieri, S. Biodiversity hotspots and major tropical wilderness areas: Approaches to setting conservation priorities. Conserv. Biol. 1998, 12, 516–520. [Google Scholar] [CrossRef]
- Aguirre, L.F. Structure of a Neotropical savanna bat community. J. Mammal. 2002, 83, 775–784. [Google Scholar] [CrossRef]
- Zortéa, M.; Alho, C.J.R. Bat diversity of a Cerrado habitat in central Brazil. Biodivers. Conserv. 2008, 17, 791–805. [Google Scholar] [CrossRef]
- Morales-Martínez, D.M.; Rodríguez-Posada, M.E.; Fernández-Rodríguez, C.; Calderón-Capote, M.C.; Gutiérrez-Sanabria, D.R. Spatial variation of bat diversity between three floodplain-savanna ecosystems of the Colombian Llanos. Therya 2018, 9, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento, G.; Monasterio, M. Studies on the savanna vegetation of the Venezuelan Llanos: I. The use of association analysis. J. Ecol. 1969, 57, 579–598. [Google Scholar] [CrossRef]
- Jansen-Jacobs, M.; ter Steege, H. Southwest Guyana: A complex mosaic of savannahs and forest. In Plant Diversity in Guyana: With Recommendations for a Protected Areas Strategy; Ter Steege, H., Ed.; Tropenbos Series; Tropenbos International: Wageningen, The Netherlands, 2000; Volume 18, pp. 147–157. [Google Scholar]
- Van Donselaar, J. Observations on savanna vegetation-types in the Guianas. Plant Ecol. 1969, 17, 271–312. [Google Scholar] [CrossRef]
- Lee, T.E., Jr.; Lim, B.K.; Hanson, J.D. Noteworthy records of mammals from the Orinoco River drainage of Venezuela. Tex. J. Sci. 2000, 52, 264–266. [Google Scholar]
- Lim, B.K.; Osborne, C.; Ignace, A. Small mammals of the South Rupununi region, Guyana. In Biodiversity Assessment Survey of the South Rupununi Savannah, Guyana; Alonso, L.E., Persaud, J., Williams, A., Eds.; BAT Survey Report No. 1; WWF-Guianas: Georgetown, Guyana, 2016; pp. 103–118. [Google Scholar]
- Sikes, R.S.; Animal Care and Use Committee of the American Society of Mammalogists. 2016 guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples, Version 9. 2013. Available online: http://purl.oclc.org/estimates (accessed on 1 December 2017).
- Clare, E.L.; Lim, B.K.; Engstrom, M.D.; Eger, J.L.; Hebert, P.D.N. DNA barcoding of Neotropical bats: Species identification and discovery within Guyana. Mol. Ecol. Notes 2007, 7, 184–190. [Google Scholar] [CrossRef]
- Lim, B.K. Review of genetic diversification of bats in the Caribbean and biogeographic relationships to Neotropical species based on DNA barcodes. Genome 2017, 60, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed]
- LaVal, R.K.; Fitch, H.S. Structure, movements and reproduction in three Costa Rican bat communities. Occas. Pap. 1977, 69, 1–28. [Google Scholar]
- Lim, B.K.; Engstrom, M.D. Bat community structure at Iwokrama Forest, Guyana. J. Trop. Ecol. 2001, 17, 647–665. [Google Scholar] [CrossRef]
- Wilson, D.E. Mammals of La Selva, Costa Rica. In Four Neotropical Rainforests; Gentry, A.H., Ed.; Yale University Press: New Haven, CT, USA, 1990; pp. 273–286. ISBN 9780300054484. [Google Scholar]
- Lim, B.K.; Engstrom, M.D. Mammals of Iwokrama Forest. Proc. Acad. Nat. Sci. Phila. 2005, 154, 71–108. [Google Scholar] [CrossRef]
- Rex, K.; Kelm, D.H.; Wiesner, K.; Kunz, T.H.; Voigt, C.C. Species richness and structure of three Neotropical bat assemblages. Biol. J. Linn. Soc. 2008, 94, 617–629. [Google Scholar] [CrossRef] [Green Version]
- Lim, B.K. Environmental assessment at the Bakhuis bauxite concession: Small-sized mammal diversity and abundance in the lowland humid forests of Suriname. Open Biol. J. 2009, 2, 42–53. [Google Scholar] [CrossRef]
- Wilson, D.E.; Ascorra, C.F.; Solari, S. Bats as indicators of habitat disturbance. In Manu: The Biodiversity of Southeastern Peru; Wilson, D.E., Sandoval, A., Eds.; Smithsonian Institution and Editorial Horizonte: Washington, DC, USA; Lima, Peru, 1996; pp. 613–625. ISBN 1560987103. [Google Scholar]
- Arteaga, L.L.; Aguirre, L.F.; Moya, M.I. Seed rain produced by bats and birds in forest islands in a Neotropical savanna. Biotropica 2006, 38, 718–724. [Google Scholar] [CrossRef]
- Rivero-Blanco, C.; Dixon, J.R. Origin and distribution of the herpetofauna of the dry lowland regions of northern South America. In The South America Herpetofauna: Its Origin, Evolution, and Dispersal; Duellman, W.E., Ed.; Monograph of the Museum of Kansas: Lawrence, Kansas, 1979; Volume 7, pp. 281–298. ISBN 0-89338-008-3. [Google Scholar]
- Linares, O.J.; Ojasti, J. Una nueva especie de murcielago del genero Lonchorhina (Chiroptera: Phyllstomidae) del sur de Venezuela. Serie Zoológica 1971, 36, 1–8. [Google Scholar]
- Hernández-Camacho, J.; Cadena-G., A. Notas para la revision del genero Lonchorhina (Chiroptera, Phyllstomidae). Caldasia 1978, 12, 199–251. [Google Scholar]
- Lim, B.K.; Engstrom, M.D.; Bickham, J.W.; Patton, J.C. Molecular phylogeny of New World emballonurid bats (Tribe Diclidurini) based on loci from the four genetic transmission systems in mammals. Biol. J. Linn. Soc. 2008, 93, 189–209. [Google Scholar] [CrossRef]
- Handley, C.O., Jr. Mammals of the Smithsonian Venezuelan Project. Brigh. Young Univ. Sci. Bull. Biol. Ser. 1976, 20, 1–89. [Google Scholar] [CrossRef]
- Webster, W.D.; Handley, C.O., Jr. Systematics of Miller’s Long-Tongued Bat, Glossophaga Longirostris, with Description of Two New Subspecies; Occasional Papers, No. 100; The Museum, Texas Tech University: Lubbock, TX, USA, 1986; pp. 1–22. [Google Scholar]
- Griffiths, T.A.; Gardner, A.L. Subfamily Glossophaginae Bonaparte, 1845. In Mammals of South America: Volume 1: Marsupials, Xenarthrans, Shrews, and Bats; Gardner, A.L., Ed.; The University of Chicago Press: Chicago, IL, USA; London, UK, 2008; pp. 224–255. ISBN 978-0-226-28240-4. [Google Scholar]
- Loureiro, L.O.; Lim, B.K.; Engstrom, M.D. A new species of mastiff bat (Chiroptera, Molossidae, Molossus) from Guyana and Ecuador. Mammal. Biol. 2018, 90, 10–21. [Google Scholar] [CrossRef]
- Bickham, J.W.; Ruedas, L.A. Genus Rhogeessa H. Allen, 1866. In Mammals of South America: Volume 1: Marsupials, Xenarthrans, Shrews, and Bats; Gardner, A.L., Ed.; The University of Chicago Press: Chicago, IL, USA; London, UK, 2008; pp. 481–484. ISBN 978-0-226-28240-4. [Google Scholar]
- Baird, A.B.; Hillis, D.M.; Patton, J.C.; Bickham, J.W. Evolutionary history of the genus Rhogeessa (Chiroptera: Vespertilionidae) as revealed by mitochondrial DNA sequences. J. Mammal. 2008, 89, 744–754. [Google Scholar] [CrossRef]
- Clare, E.L. Cryptic species: Patterns of maternal and paternal gene flow in eight Neotropical bats. PLoS ONE 2011, 6, e21460. [Google Scholar] [CrossRef] [PubMed]
- Velazco, P.M.; Cadenillas, R. On the identify of Lophostoma silvicolum occidentalis (Davis & Carter, 1978) (Chiroptera: Phyllostomidae). Zootaxa 2011, 2962, 1–20, ISSN 1175-5326. [Google Scholar]
- Velazco, P.M.; Soto-Centeno, J.A.; Fleck, D.W.; Voss, R.S.; Simmons, N.B. A new species of nectar-feeding bat of the genus Hsunycteris (Phyllostomidae: Lonchophyllinae) from northeastern Peru. Am. Mus. Novit. 2017, 3881, 1–26, ISSN 0003-0082. [Google Scholar] [CrossRef]
- Parlos, J.A.; Timm, R.M.; Swier, V.J.; Zeballos, H.; Baker, R.J. Evaluation of Paraphyletic Assemblages within Lonchophyllinae, with Description of a New Tribe and Genus; Occasional Papers; Museum of Texas Tech University: Lubbock, TX, USA, 2014; Volume 320, pp. 1–23, ISSN 0149-175X. [Google Scholar]
- Lim, B.K. Historical biogeography of New World emballonurid bats (Tribe Diclidurini): Taxon pulse diversification. J. Biogeogr. 2008, 35, 1385–1401. [Google Scholar] [CrossRef]
- Lim, B.K. Divergence times and origin of Neotropical sheath-tailed bats (Tribe Diclidurini) in South America. Mol. Phylogenet. Evol. 2007, 45, 777–791. [Google Scholar] [CrossRef]
- Wyss, A.R.; Flynn, J.J.; Norell, M.A.; Swisher, C.C.; Charrier, R.; Novacek, M.J.; McKenna, M.C. South America’s earliest rodent and recognition of a new interval of mammalian evolution. Nature 1993, 365, 434–437. [Google Scholar] [CrossRef]
- Lim, B.K.; Engstrom, M.D.; Patton, J.C.; Bickham, J.W. Systematic review of small fruit-eating bats (Artibeus) from the Guianas, and a re-evaluation of A. glaucus bogotensis. Acta Chiropterol. 2008, 10, 243–256. [Google Scholar] [CrossRef]
- Marques-Aguiar, S.A. Genus Artibeus Leach, 1821. In Mammals of South America: Volume 1: Marsupials, Xenarthrans, Shrews, and Bats; Gardner, A.L., Ed.; The University of Chicago Press: Chicago, IL, USA; London, UK, 2008; pp. 301–321. ISBN 978-0-226-28240-4. [Google Scholar]
Figure 1.
Map of the Guianas in northern South America with white outlines identifying the savanna study sites in Venezuela (Llanos), Guyana (Rupununi), and Suriname (Sipaliwini) that were surveyed for bats ©Burton K. Lim.
Figure 1.
Map of the Guianas in northern South America with white outlines identifying the savanna study sites in Venezuela (Llanos), Guyana (Rupununi), and Suriname (Sipaliwini) that were surveyed for bats ©Burton K. Lim.

Figure 2.
Savannas from left to right of the Llanos in Venezuela with rocky outcrops, Rupununi in Guyana characterized by gallery forest, and Sipaliwini in Suriname with forest on the opposite bank of the river.
Figure 2.
Savannas from left to right of the Llanos in Venezuela with rocky outcrops, Rupununi in Guyana characterized by gallery forest, and Sipaliwini in Suriname with forest on the opposite bank of the river.

Figure 3.
Species accumulation curves with 95% confidence intervals of bats surveyed in three savanna habitats in the Guianas with the Llanos extrapolated from 279 individuals and Sipaliwini extrapolated from 562 individuals to match more closely the 922 individuals of the Rupununi.
Figure 3.
Species accumulation curves with 95% confidence intervals of bats surveyed in three savanna habitats in the Guianas with the Llanos extrapolated from 279 individuals and Sipaliwini extrapolated from 562 individuals to match more closely the 922 individuals of the Rupununi.

Figure 4.
DNA barcoding trees of bat taxa with endemism in the Guianan savannas: (a) Glossophaga, (b) Lonchorhina, (c) Rhogeessa, (d) Peropteryx, and (e) Molossus.
Figure 4.
DNA barcoding trees of bat taxa with endemism in the Guianan savannas: (a) Glossophaga, (b) Lonchorhina, (c) Rhogeessa, (d) Peropteryx, and (e) Molossus.

Figure 5.
DNA barcoding trees of bat species with phylogeographic structuring of distinct lineages for populations from the Venezuelan Llanos in the states of Amazonas and Bolivar: (a) Trachops, (b) Gardnerycteris, and (c) Platyrrhinus.
Figure 5.
DNA barcoding trees of bat species with phylogeographic structuring of distinct lineages for populations from the Venezuelan Llanos in the states of Amazonas and Bolivar: (a) Trachops, (b) Gardnerycteris, and (c) Platyrrhinus.

Figure 6.
DNA barcoding trees of bat species with complex phylogeographic structuring involving the Guianan savannas: (a) Lophostoma and (b) Micronycteris.
Figure 6.
DNA barcoding trees of bat species with complex phylogeographic structuring involving the Guianan savannas: (a) Lophostoma and (b) Micronycteris.

Figure 7.
DNA barcoding tree of Hsunycteris with a divergent split between lineages that include savanna populations from the Sipaliwini in Suriname and the Llanos in Venezuela.
Figure 7.
DNA barcoding tree of Hsunycteris with a divergent split between lineages that include savanna populations from the Sipaliwini in Suriname and the Llanos in Venezuela.

Figure 8.
The first record of the Orinoco sword-nosed bat (Lonchorhina orinocensis), known previously from the Llanos of Colombia and Venezuela, is now documented in the Rupununi savanna of Guyana.
Figure 8.
The first record of the Orinoco sword-nosed bat (Lonchorhina orinocensis), known previously from the Llanos of Colombia and Venezuela, is now documented in the Rupununi savanna of Guyana.

Table 1.
Summary of bat survey data and biodiversity measures in the Guianan savannas. (ACE: Abundance-based Coverage Estimators ICE: Incidence-based Coverage Estimators).
Table 1.
Summary of bat survey data and biodiversity measures in the Guianan savannas. (ACE: Abundance-based Coverage Estimators ICE: Incidence-based Coverage Estimators).
| Variable | Llanos | Rupununi | Sipaliwini | Total |
|---|---|---|---|---|
| Observed data: | ||||
| Individuals | 279 | 922 | 562 | 1763 |
| Species | 35 | 56 | 49 | 76 |
| Trap-nights | 15 | 43 | 18 | 76 |
| Richness estimators: | ||||
| ACE | 48.75 | 69.14 | 56.77 | 83.31 |
| ICE | 51.04 | 69.3 | 61.4 | 86.52 |
| Chao 1 | 42.31 | 68.99 | 53.99 | 84.25 |
| Chao 2 | 40.6 | 72.74 | 56.81 | 92.92 |
| Jack 1 | 47.13 | 71.63 | 62.22 | 91.79 |
| Jack 2 | 48.74 | 81.3 | 66.3 | 101.6 |
| Bootstrap | 41.17 | 62.81 | 55.5 | 83.16 |
| Average richness | 45.68 | 70.84 | 59 | 89.7 |
| Diversity indices: | ||||
| Alpha | 10.57 | 13.13 | 12.91 | 16.17 |
| Shannon | 2.66 | 2.95 | 2.64 | 3.11 |
| Shannon Exponential | 14.31 | 19.16 | 14.02 | 22.52 |
| Simpson | 9.04 | 9.72 | 6.18 | 9.42 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lim, B.K.; Lee, T.E., Jr. Community Ecology and Phylogeography of Bats in the Guianan Savannas of Northern South America. Diversity 2018, 10, 129. https://doi.org/10.3390/d10040129
AMA Style
Lim BK, Lee TE Jr. Community Ecology and Phylogeography of Bats in the Guianan Savannas of Northern South America. Diversity. 2018; 10(4):129. https://doi.org/10.3390/d10040129
Chicago/Turabian StyleLim, Burton K., and Thomas E. Lee, Jr. 2018. "Community Ecology and Phylogeography of Bats in the Guianan Savannas of Northern South America" Diversity 10, no. 4: 129. https://doi.org/10.3390/d10040129
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.