Does Thinning Homogenous and Dense Regrowth Benefit Bats? Radio-Tracking, Ultrasonic Detection and Trapping


1
Forest Science Unit, NSW Department of Primary Industries, Parramatta, NSW 2124, Australia
2
Wildlife Research Center, Kyoto University, Kyoto 606-8203, Japan
*
Author to whom correspondence should be addressed.
Diversity 2018, 10(2), 45; https://doi.org/10.3390/d10020045
Submission received: 11 April 2018
/
Revised: 30 May 2018
/
Accepted: 2 June 2018
/
Published: 6 June 2018
(This article belongs to the Special Issue Diversity and Conservation of Bats)
Abstract
:Renewal ecology promotes the creation and enhancement of landscapes that support biodiversity and ecosystem services for humans. Silvicultural thinning of forest regrowth to reduce tree competition represents a form of active management that may also benefit biodiversity, especially where secondary regrowth dominates. However, ecological responses to thinning can be complex, particularly for insectivorous bats whose ecomorphology is often related to vegetation structure. Furthermore, thinning may affect multiple aspects of bat ecology (i.e., roosting and foraging). We assessed this in dense white cypress regrowth in the Pilliga forests of New South Wales, Australia, where recent experimental thinning created thinned stands (4 × 12 ha) surrounded by unthinned regrowth. We contrasted flight activity and roost selection of three narrow-space species with differing conservation statuses (Nyctophilus corbeni, N. gouldi and N. geoffroyi), plus one edge-space species (Vespadelus vulturnus). Radio-tracking over two maternity seasons revealed a preference by all species for roosting in dead trees that were slightly larger than the mean for available dead trees in the vicinity. Although all tagged bats were caught in thinned patches, only 6% of roosts were located there. In contrast, ultrasonic detectors recorded significantly greater activity for V. vulturnus (p = 0.05) in thinned than unthinned patches and no treatment difference for Nyctophilus spp. Systematic trapping using acoustic lures found a higher trap rate for N. gouldi in unthinned than thinned treatments, but no treatment effect for N. corbeni, N. geoffroyi and V. vulturnus. Our results reveal differential use of forest treatments by multiple species, emphasising the value of heterogeneous landscapes supporting thinned and unthinned patches of dense regrowth.
1. Introduction
Renewal ecology is a newly-proposed concept that promotes both the creation and enhancement of landscapes that support biodiversity and provide ecosystem services for human communities [1]. Renewal ecology recognizes the need to harmonize biodiversity with humans, for the benefit of both, and it emphasises active management, using targeted interventions. It differs from restoration ecology that typically aims to return ecosystems to a previous, indigenous state [2]. There are many forms of active management that potentially fit this concept, including certain forestry activities.
Timber production forests throughout the world not only provide a valued timber resource, but are important for maintaining biodiversity and irreplaceable ecosystem services [3,4]. Silvicultural thinning of dense secondary forest regrowth reduces tree competition and encourages faster growth of mature, harvestable trees [5]. This is particularly the case when the secondary regrowth that dominates stands post-harvesting comprises trees that are uniform in age and size, creating a high density of stems with few canopy gaps [6]. In addition to influencing tree growth, thinning affects the physical structure and composition of vegetation and modifies soil properties and microclimates (see [7] for a review). Given that biodiversity responses to thinning are complex, with changes being detrimental to some taxa and beneficial to others [7,8,9,10], it is essential to more fully explore the effects of this practice on a range of taxa.
Insectivorous bats are a diverse group of mammals whose morphological traits, including body size and wing shape, often influence the use of different vegetation structures [11,12,13,14,15,16]. In particular, one ensemble of bats, narrow-space bats [17], that possess specialised traits favouring slow, manoeuvrable flight within vegetation clutter, particularly in forests, is considered especially prone to extinction [18,19]. While these bats can sometimes also forage within open space [20,21], they are often forest-dependent and are predicted to be particularly sensitive to vegetation loss and potentially to altered vegetation structure, for instance due to thinning of dense forests.
Forest thinning reduces clutter levels and so can be beneficial for a range of bat species by increasing echolocation efficiency [5,10,22,23]. Yet, narrow-space species typically have higher activity levels in forests with higher stem densities [24,25,26], highlighting the need for a better understanding of their response to thinning. Despite roosts being a critical part of the bat life cycle [27], it is poorly understood how forest thinning affects roost selection, especially for narrow-space bats. One threatened, narrow-space species (south-eastern long-eared bat Nyctophilus corbeni) prefers roosting where total stem density and the density of dead stems is high and appears to avoid roosting in thinned areas [28]. Both sexes, including maternal females, follow this pattern. In comparison, a less manoeuvrable edge-space species (eastern red bat Lasiurus borealis) roosts in foliage in open conditions created by thinning [29].
Our study aimed to contrast flight activity and roost selection by a small suite of bat species in response to recent experimental thinning in a dry, but dense, forest of white cypress pine Callitris glaucophylla. Callitris glaucophylla is geographically widespread in Australia and has a history of silvicultural thinning in parts of its distribution. The species is slow growing and long-lived, tolerating intense intra-specific competition at high densities [30,31,32], with little evidence of self-thinning [33] for up to 200 years in low rainfall environments [34]. We chose to radio-track multiple bat species to contrast the roost preferences of different narrow-space species that possess differing conservation statuses (Nyctophilus corbeni, Gould’s long-eared bat N. gouldi and lesser long-eared bat N. geoffroyi), with one small, edge-space species (little forest bat Vespadelus vulturnus). We predicted that the three narrow-space species would prefer to roost and be more active in dense unthinned forest [28], but we also expected some partitioning of roost tree attributes between the species. In contrast, the small, edge-space species was predicted to roost and be more active in the thinned forest [26]. We based our assessments on a combination of radio-tracking, ultrasonic detection and systematic trapping. Trapping with an acoustic lure [35] was used to assess the activity of Nyctophilus in the thinned-unthinned mosaic, because it is currently not possible to distinguish echolocation calls of these three Nyctophilus species [36].
2. Materials and Methods
2.1. Study Species
Nyctophilus corbeni is listed nationally as a threatened species in Australia (Vulnerable, Environment Protection and Biodiversity Conservation Act 1999) and is substantially larger than two sympatric conspecifics (Table S1). In contrast, N. gouldi and N. geoffroyi are abundant and widespread [37,38], though all three species are morphologically similar. All three Nyctophilus species belong to the same narrow-space guild with low amplitude, steep, broad band echolocation calls, with lepidopterans (moths) representing a major part of their diets (Table S1). The main threatening processes for N. corbeni are considered to be extensive loss of habitat through clearing for agriculture, grazing and altered fire regimes [39], with different forestry activities having contrasting effects [28]. The fourth study species, V. vulturnus, is the smallest bat in the study area (Table S1) and was selected because individuals were expected to forage within a 1-km radius [40,41,42], thus making the species highly suited, at a local scale, for revealing relative use of thinned and unthinned forest. It is a common species belonging to the edge-space ensemble based on its classic frequency modulated-constant frequency (FM-CF) echolocation call structure, but with relatively slow and manoeuvrable flight due to low wing aspect ratio and loading [43,44]. These ecomorphological traits indicate that it is agile enough to follow the edges provided by well-separated crowns of trees, but less likely to regularly forage in very dense unthinned forest [26,41]. All research was approved by the Department Primary Industries Forestry Corporation NSW Animal Ethics Committee (Authority Numbers 21/15, 21/16 and 21/17).
2.2. Study Area
The study was undertaken in Pilliga West State Forest (−30°44′45′’, 149°6′41′’) in the Brigalow Belt South bioregion of northwest New South Wales (NSW), Australia. The area experiences a hot and dry climate, with the mean annual rainfall for nearby Baradine being 633 mm (1944–1998). The Pilliga area has a long history of European use, and consequently, considerable changes to forest structure have been inferred. It has been suggested that much of the Pilliga area was originally open woodland with a similar plant species composition to today [45,46]. However, the extent of areas with differing tree density and its patchiness generally remain unknown. Increasing tree density is thought to have displaced sheep grazing ventures in the 1800s. Forest management for timber began in 1877, and the first local forestry office opened in 1911 [47]. Natural Callitris regeneration over this time appears to have been strongly influenced by a combination of reduced burning, introduction of rabbits and interactions with drought and flood years.
The vegetation today is classified as a forest that varies in tree height with soil type. A variety of forest types occur throughout the Pilliga with dense stands of C. glaucophylla, black cypress pine C. endlicheri, buloke Allocasuarina luehmannii, Acacia spp. and eucalypts such as narrow-leaved ironbark Eucalyptus crebra scattered throughout the forest [48,49]. Callitris glaucophylla, in particular, was commonly treated silviculturally to maximise tree growth for timber production [47,49]. Locally, our study area was dominated by dense, unthinned stands of regrowth C. glaucophylla. In June–July 2016, experimental thinning was undertaken with small machinery at four 12-ha replicate patches as part of a broader study on the effects of thinning on biodiversity. Thinning at these patches was considered ‘early’ thinning that targeted the suppressed cypress and competing co-dominant cypress trees that were 4–6 m tall and aimed to achieve a remaining tree spacing of approximately 6 m (280 stems ha−1). Each replicate was separated from the nearest thinned or control patch by at least 200 m. Four designated unthinned control patches with similar vegetation (density of stems >5 cm diameter at breast height over bark (DBHOB) was >3600 stems ha−1), each 12 ha in size, were used for comparison (Figure 1).
2.3. Bat Activity
Bat activity was assessed in November (spring) 2016 and March (autumn) 2017 in the thinned-unthinned mosaic using ultrasonic Anabat detectors (Anabat II, SD1 and SD2; Titley Scientific, Brendale, Australia) for two nights simultaneously in each thinned or unthinned replicate. All detectors were calibrated prior to deployment to ensure that microphone sensitivity was similar among all detectors. In the autumn sampling session, two detectors were positioned ~100 m apart within each treatment (~150 m from the unthinned edge), while a single detector was located in the centre of each treatment in the spring sampling session (~200 m from the unthinned edge). Detectors were positioned on the ground but with microphones set at a height of 1 m and oriented at a 45° angle to the ground to face gaps within vegetation at each site. This was done to ensure detectors at each site were sampling a similar volume of airspace. All files were analysed using automated software, AnaScheme [50], in association with a key for bats of the Pilliga (unpublished, B. Law). Only bat passes with more than three valid pulses, a minimum of six data points and model quality of 0.8 were analysed by AnaScheme. Because multiple bat species may call simultaneously, calls were assigned to a species only if >50% of pulses within the sequence were attributed to that species, and only passes with a minimum of three pulses classified to the same species were identified. All passes that could not be assigned to a bat taxon were included in counts of total bat activity, but were labelled as ‘unidentified’. Since it is not possible to distinguish between calls of certain species (e.g., N. corbeni, N. geoffroyi and N. gouldi, or southern free-tailed bat Mormopterus planiceps and inland freetail bat M. petersi, or little broad-nosed bat Scotorepens greyii and central-eastern broad-nosed bat Scotorepens sp.), these calls were assigned to a species group (e.g., Nyctophilus spp., or Mormopterus planiceps/petersi, or Scotorepens greyii/sp.) by AnaScheme. For each detector and each night, the number of bat passes, hereafter “calls”, for each species or species group (nightly activity) and total bat activity (all identified and unidentified bat calls) were tabulated.
In November 2016/2017, we used harp-traps [51] to quantify activity in the thinned-unthinned mosaic for the three species of Nyctophilus, given that their echolocation calls cannot be identified to the species level. To reduce bias from trapping in the open thinned forest and dense, unthinned forest, we used an acoustic lure (Sussex Autobat; [35]) with the aim of luring foraging bats directly into the harp-trap from a distance of <50 m. We systematically trapped the thinned-unthinned mosaic using multiple pairs of traps, one with an acoustic lure and one without. One trap pair was positioned in each of the four thinned and four unthinned replicates, typically for two nights, with the acoustic lure rotated between traps each night. Traps were set near the centre of replicate patches to avoid luring individuals from outside the treated area. Additional trapping was undertaken on any flyways that were present, but these results are not presented here. The number of bats caught per trap-night was summed for each thinned and unthinned replicate.
2.4. Radio-Tracking
To maximise the likelihood of detecting bats roosting and foraging in thinned patches (which were proportionately small in comparison to the surrounding untreated forest), only bats trapped within thinned forest were radio-tagged. If bats were trapped in the first half of the night, they were processed immediately and released on the same night of capture. All other bats were retrieved the following morning and released at dusk. Since the major aim of this study centred on maternity roost preferences, female bats that were pregnant or lactating were selected for radio-tracking. Radio-transmitters with an aerial length of 12 cm (Holohil, Carp, Canada; Model LB-2X, 0.27–0.42 g–<6.8% of body mass) were attached mid-dorsally (between shoulder blades) to bats using an adhesive (Urobond, Urocare, Pomona, CA, USA). Pregnant females weighed more than lactating females (e.g., an additional 3.75 g for pregnant N. corbeni), but we did not observe difficulty flying or use of unusual roosts with the additional mass of transmitters. After radio-transmitters were attached, bats were placed into calico bags and held for a minimum of 10 min prior to release at the site of capture.
Bats were radio-tracked over 10 days in the maternity season in each of two years (November 2016/2017), though radio-transmitters were typically removed by bats (presumably via grooming) after four days. Roost trees were located by homing in on the signals of radio-tracked bats. Once a roost tree was identified, binoculars were used to identify the roost type (e.g., hollow, fissure, etc.). For each roost tree, diameter at breast height over bark (DBHOB, 1.3 m) was measured while tree height was visually estimated. Each roost tree was also assigned an age-class (senescence score 0–8; following [52]), hollow abundance (0–4; 0 = no apparent hollows, 4 = abundant hollows), and canopy cover was estimated visually or using the ‘Habitapp’ Android application. An emergence count was conducted at selected roost trees at dusk to confirm hollow-type and to count the number of bats that exited the roost. To quantify the amount of time radio-tracked bats spent in thinned plots, two Australis 26K scanning receivers (Titley Scientific, Brendale, Australia) fitted with remote RF data loggers (Titley Scientific) and omni-directional whip antennas (Titley Scientific) were set in the centre of two thinned plots in 2017. Antennas were secured 3 m above the ground and on a tree. Scanning receivers actively scanned through radio frequencies of tagged bats, and RF data loggers logged the presence of a signal (“entry”) every four seconds if a pre-determined signal strength was achieved and a minimum of 3 pulses was detected in that period. If a signal was not detected, this was also logged (“exit”). The time spent by each radio-tracked bat in each thinned plot was tallied and averaged across nights for each bat. Gain settings on receivers were calibrated using an active transmitter to record bats within 200 m of the logger. This was done to minimise the chance of logging bats that were flying outside of thinned patches.
2.5. Roost Tree Availability
We estimated roost tree availability at the local scale and in relation to the surrounding thinned-unthinned mosaic. At the local scale, we estimated tree density around each roost tree by measuring the distance to the nearest tree (>10 cm DBHOB) in each quadrant [53]. The point quarter technique was then used to calculate the local density of trees (including those with a hollow) for all roosts pooled across years. The standard error for the tree density estimate was calculated following Pollard (1971) [53]. At the patch scale, we calculated stem density (plus hollow stem density) from three 20 × 10 m plots located along a 200-m transect in each thinned or unthinned replicate. The number of trees and shrubs within each plot was recorded, along with their DBHOB, which was measured and allocated to one of four categories (<10 cm; >10 cm to <30 cm; >30 to <50 cm; and >50 cm). The number of dead trees within each plot was also recorded, but not allocated to size classes. These data were used to calculate mean tree density for thinned and unthinned treatments.
2.6. Data Analyses
A canonical analysis of principal coordinates (CAP) was used to examine and contrast characteristics of trees used for roosting and those available in the local neighbourhood around roost trees. Prior to analysis, a square-root transformation was applied to continuous variables (tree DBHOB and height), and all data were then normalised. A resemblance matrix (Euclidean distance) was constructed, and a CAP analysis was undertaken. Vectors representing tree characteristics were overlaid onto the CAP output. CAP analyses were conducted using the PERMANOVA+ add-on package in Primer 6 (PRIMER-E Ltd., Plymouth, UK; [54]).
The number of captures per trap-night (to account for the different number of trap-nights in each treatment) of focal species (N. corbeni, N. geoffroyi, N. gouldi, V. vulturnus) in the unthinned-thinned mosaic was compared between thinning treatments using Wilcoxon signed-rank tests.
Bat activity of focal species (Nyctophilus spp. and V. vulturnus) and total bat activity (all species combined) was compared between treatments and season using a generalized linear mixed model (GLMM). Thinning treatment, season and the interaction of treatment by season were fixed effects, while site was used as a random effect. Response variables (bat activity − number of passes detector−1 night−1) were log10-transformed prior to analysis.
Wilcoxon signed-rank tests (used for comparing capture rates in thinned and unthinned treatments) and GLMMs were run using SPSS 19.0 (IBM, Armonk, NY, USA).
3. Results
3.1. Roosting
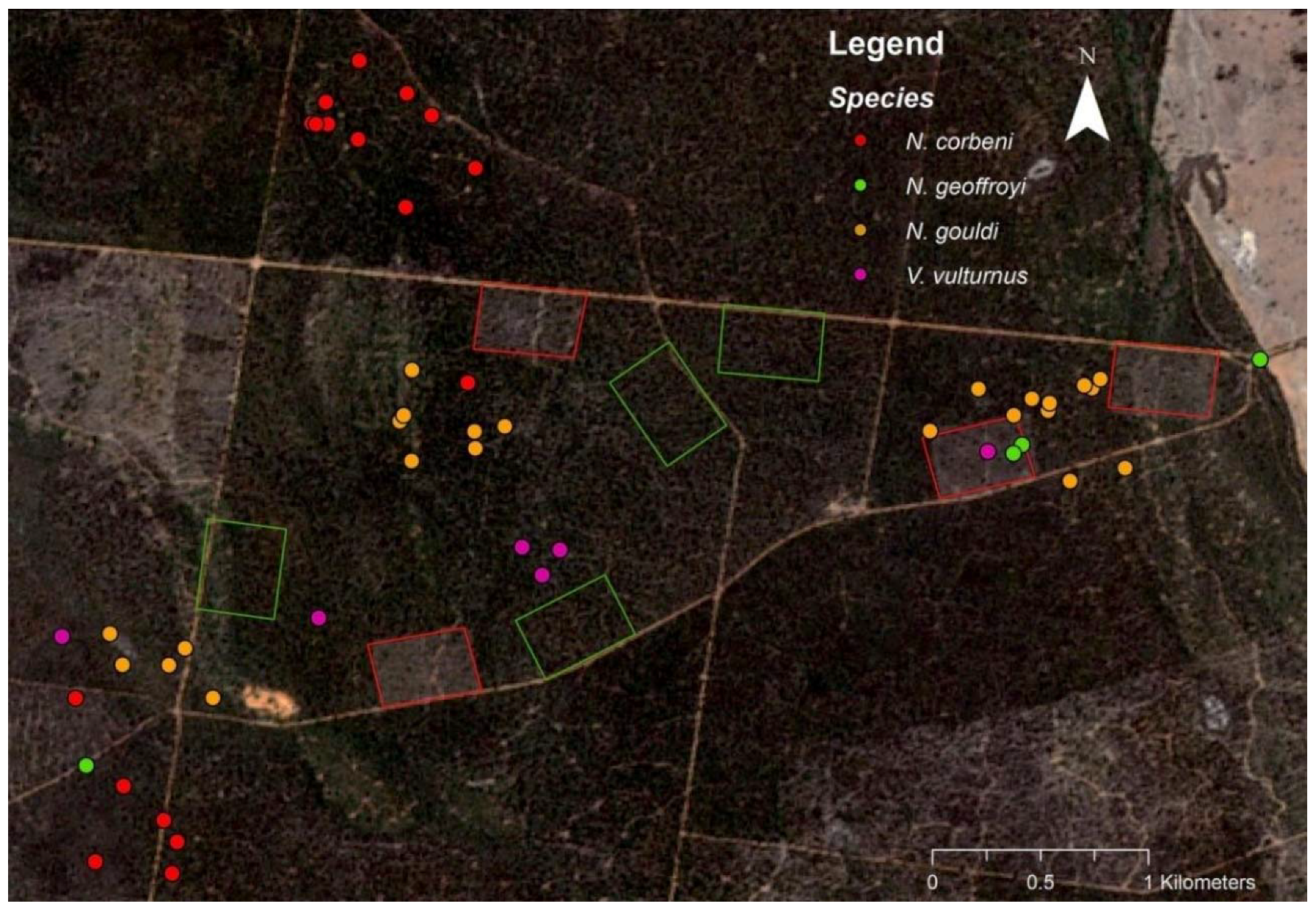
In all, 22 bats representing four species (Nyctophilus corbeni, N. geoffroyi, N. gouldi and Vespadelus vulturnus) were radio-tracked across both years. Of these, 21 were either in the late stages of pregnancy or were lactating, while a single male was also tracked (Table 1). However, no roosts were located for the male, so results represent roosting preferences of maternal females. Although bats likely moved throughout this landscape and potentially between different thinned blocks at night time, roosts were typically located close to the thinned blocks where the individual was trapped (Figure 1).
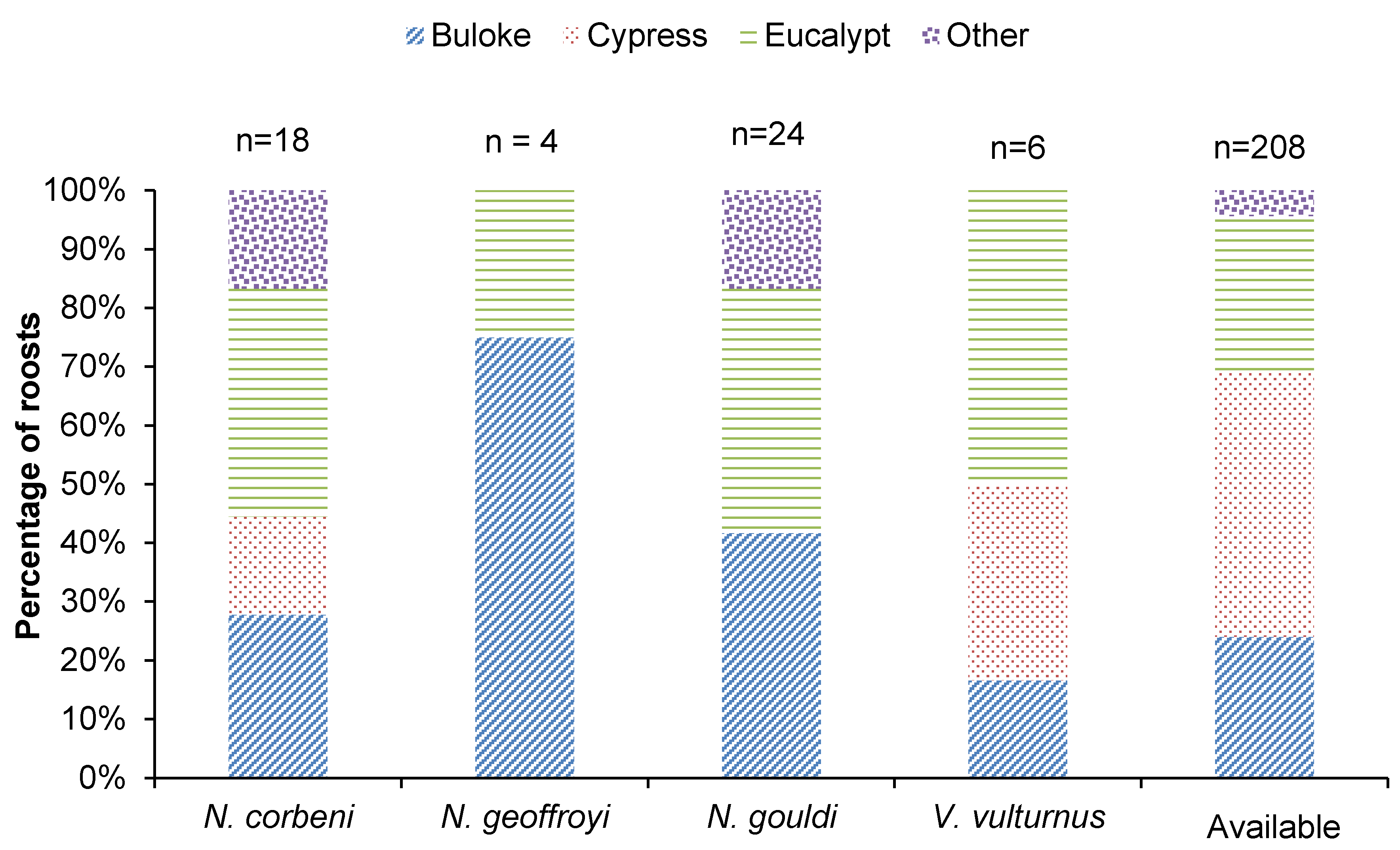
A total of 52 roost trees were located in a range of trees species across both years of radio-tracking (Figure 2). Of these 24, 18, 6 and 4 were used by N. gouldi, N. corbeni, V. vulturnus and N. geoffroyi, respectively. Across both years, >75% of roost trees were dead (Table 1). Typically <10 individuals exited roost trees that were watched at dusk, and roosts were switched almost daily (Table 1).
Buloke and eucalypts together represented more than two-thirds of roost trees for each species (Figure 2), and these were typically dead (>75%). For N. corbeni, living and dead eucalypts represented most roost trees followed by dead buloke, dead cypress and other unknown dead trees. Only a small number (n = 4) of N. geoffroyi roosts were located before transmitters were detached. Three roost trees were located in buloke, whereas one individual roosted in a living Pilliga box Eucalyptus pilligaensis in a riparian zone on the forest-paddock interface <1 km from the capture site. For N. gouldi, roosts were equally spread across dead buloke and living and dead eucalypts, with a small percentage of dead tree roosts unable to be identified to species. Dead eucalypts represented most roost trees of Vespadelus vulturnus, with dead buloke and cypress also used (Figure 2).
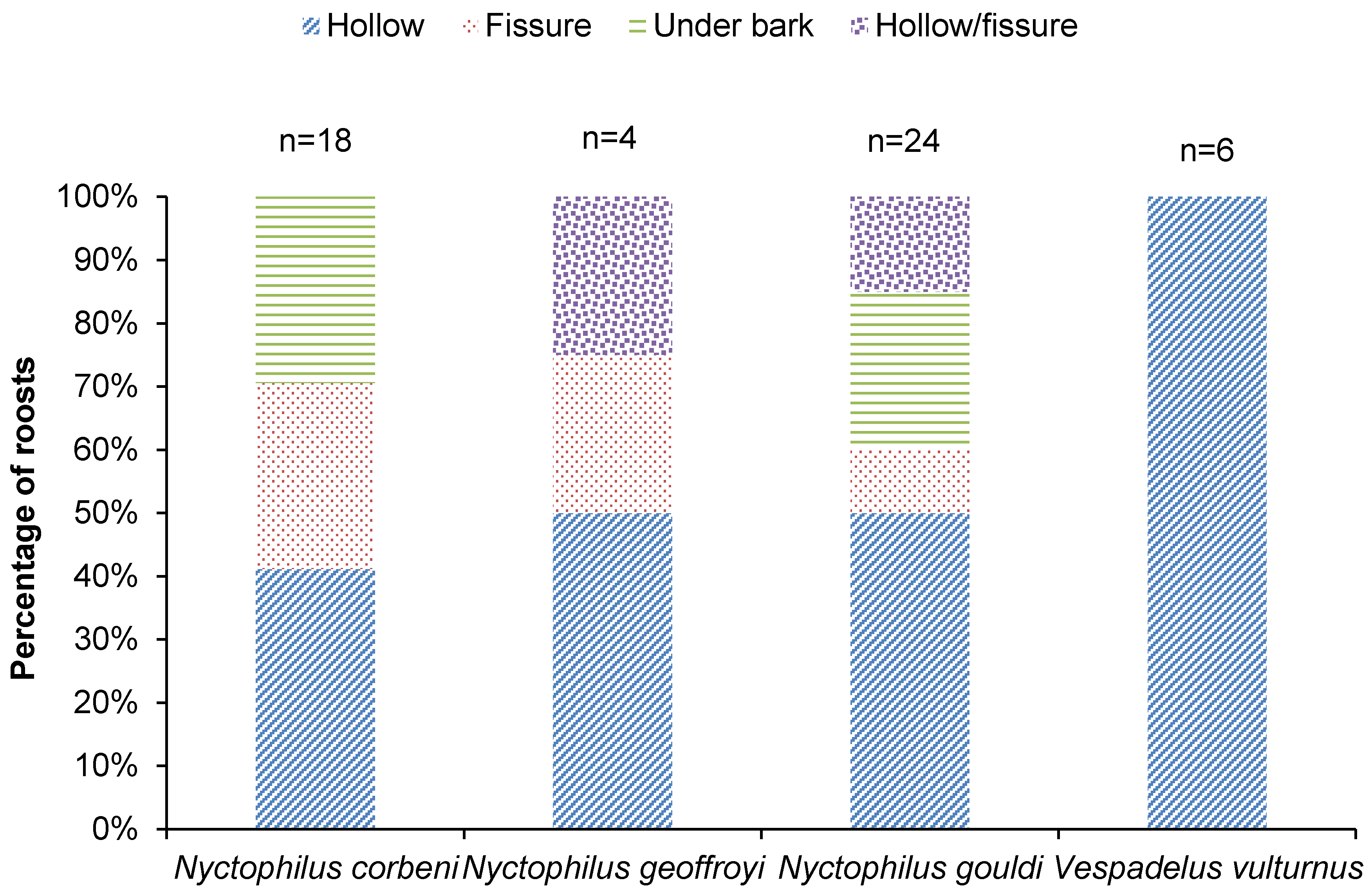
For all species, ≥50% of all roosts were located in a hollow or a fissure (Figure 3). Nyctophilus corbeni and N. gouldi also roosted under bark (Figure 3). Vespadelus vulturnus roosts were exclusively in hollows (Figure 3).
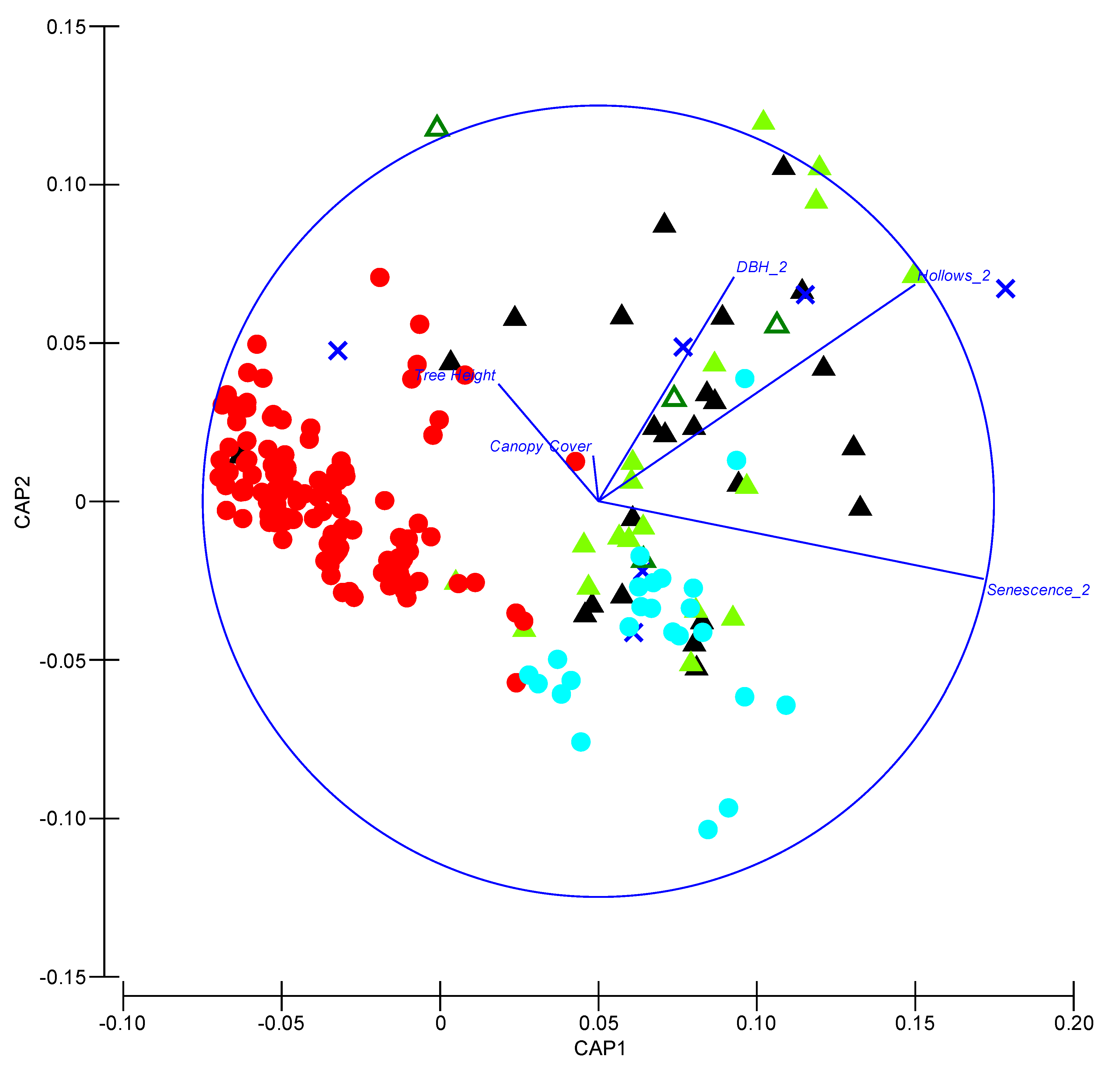
A CAP revealed that roost trees were distinct from other available living trees in the local area (Figure 4). Although available living trees covered a broad range of heights, sizes (DBHOB) and extent of canopy cover, they all generally lacked hollows and received low scores for degree of senescence (Figure 4). Dead available trees were similar to a subset of trees used for roosting by Nyctophilus spp. and V. vulturnus and were characterized by a greater degree of senescence and more hollows than living available trees (Figure 4). A subset of roost trees were larger (>DBHOB) and contained more hollows than available dead trees. Roost trees used by all species were very similar based on measured attributes, indicating considerable overlap between species.
3.2. Stem Density
Although all tagged bats were caught in thinned treatments, only 6% of roosts were located there (n = 52). This was reflected in a relatively high stem density surrounding roost trees. Although there was considerable overlap in roost tree characteristics of Nyctophilus spp. (Figure 4), areas that N. geoffroyi roosted in tended to be more open (thinned patch, riparian zone adjacent to a paddock and along a 4 wheel drive track) than those used for roosting by conspecifics. As a result, stem density in patches around roost sites was calculated for Nyctophilus spp. with and without data for N. geoffroyi roosts. The density of stems (>10 cm DBHOB) around Nyctophilus spp. roosts was 366 ± 4 stems ha−1 (382 ± 4 stems ha−1 excluding N. geoffroyi roosts). The density of stems surrounding roosts was intermediate to stem density in thinned (358 ± 89 stems ha−1) and unthinned (463 ± 76 stems ha−1) sites. However, this ignores stems <5 cm DBHOB, which had a much higher density in unthinned (3654 ± 199 stems ha−1) than thinned (2475 ± 230 stems ha−1) sites (see Figure 5). The density of stems (>10 cm DBHOB) with hollows in patches around roost trees of Nyctophilus spp. was 28 ± 1 stems ha−1, which was less than unthinned (50 ± 13 stems ha−1), but comparable to thinned (25 ± 16 stems ha−1) sites. Hollows were also present in a small number of stumps >1 m in height, but these were excluded from calculations of hollow stems. Density of stems in patches around roost sites could not be calculated for V. vulturnus using the point-quarter technique [53], as there were too few roosts located for the species.
3.3. Night Foraging
Data loggers in the centre of each of two thinned plots provided preliminary data on time spent in these plots by radio-tagged bats. Data logged for two individuals (one lactating N. corbeni and one lactating N. gouldi) over 4–5 nights in 2017 revealed that both individuals spent >25% of the logged period per night (~6.5 h due to battery drainage) in the thinned plot in which they were captured. Each bat was only logged in the thinned plot (37 ± 1% and 26 ± 9% of the logged period for N. gouldi and N. corbeni, respectively) in which it was captured and roosted approximately 0.4 km (N. gouldi) and 1.3 km (N. corbeni) from the centre of their plot. Bats captured in other thinned plots were not recorded on either logger.
3.4. Bat Activity
In all, 8546 bat passes were recorded across all sites and both seasons (1326, spring 2016; 7220, autumn 2017). Of these, 4987 were identified to one of 10 taxa. Vespadelus vulturnus was most frequently recorded (37% of identified calls), followed by Mormopterus planiceps/Mormopterus petersi (20%), Gould’s wattled bat Chalinolobus gouldii (10%), Scotorepens greyii/Scotorepens sp. (9%), yellow-bellied sheathtail bat Saccolaimus flaviventris (8%) and white-striped freetail bat Austronomus australis (6%). Activity from other taxa represented <5% of all identified calls. Total nightly activity (no. of calls night−1) differed significantly between thinning treatments (F11,1 = 6.810, p = 0.024) and seasons (F11,1 = 5.699, p = 0.036), while there was no significant interaction effect of treatment by season (F11,1 = 1.373, p = 0.266). Total nightly activity was almost two-times greater in the thinned treatment (183 ± 43 calls night−1) compared to the unthinned treatment (97 ± 15 calls night−1) and ~64% greater in autumn (167 ± 35 calls night−1) compared to spring (102 ± 29 calls night−1).
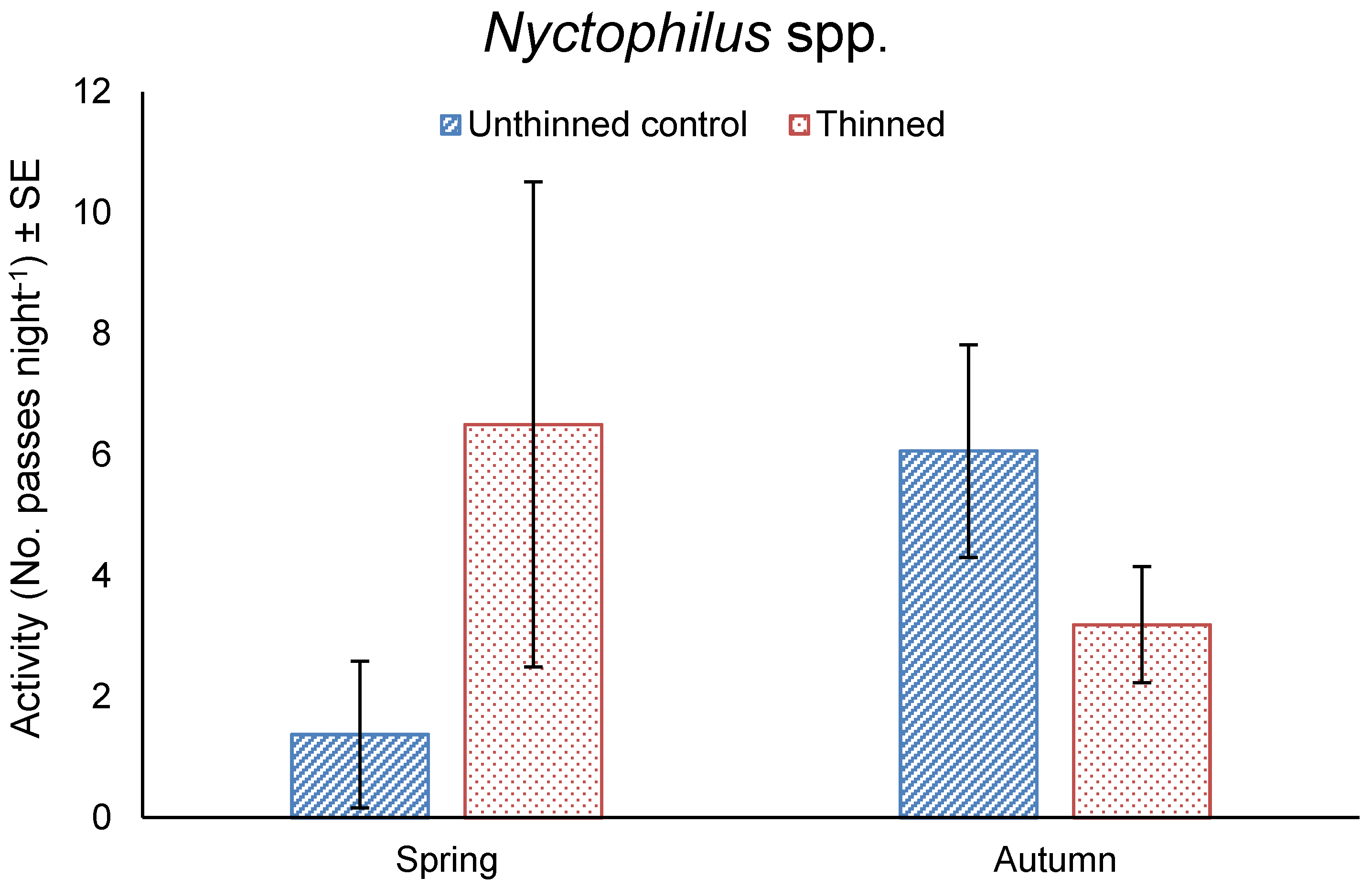
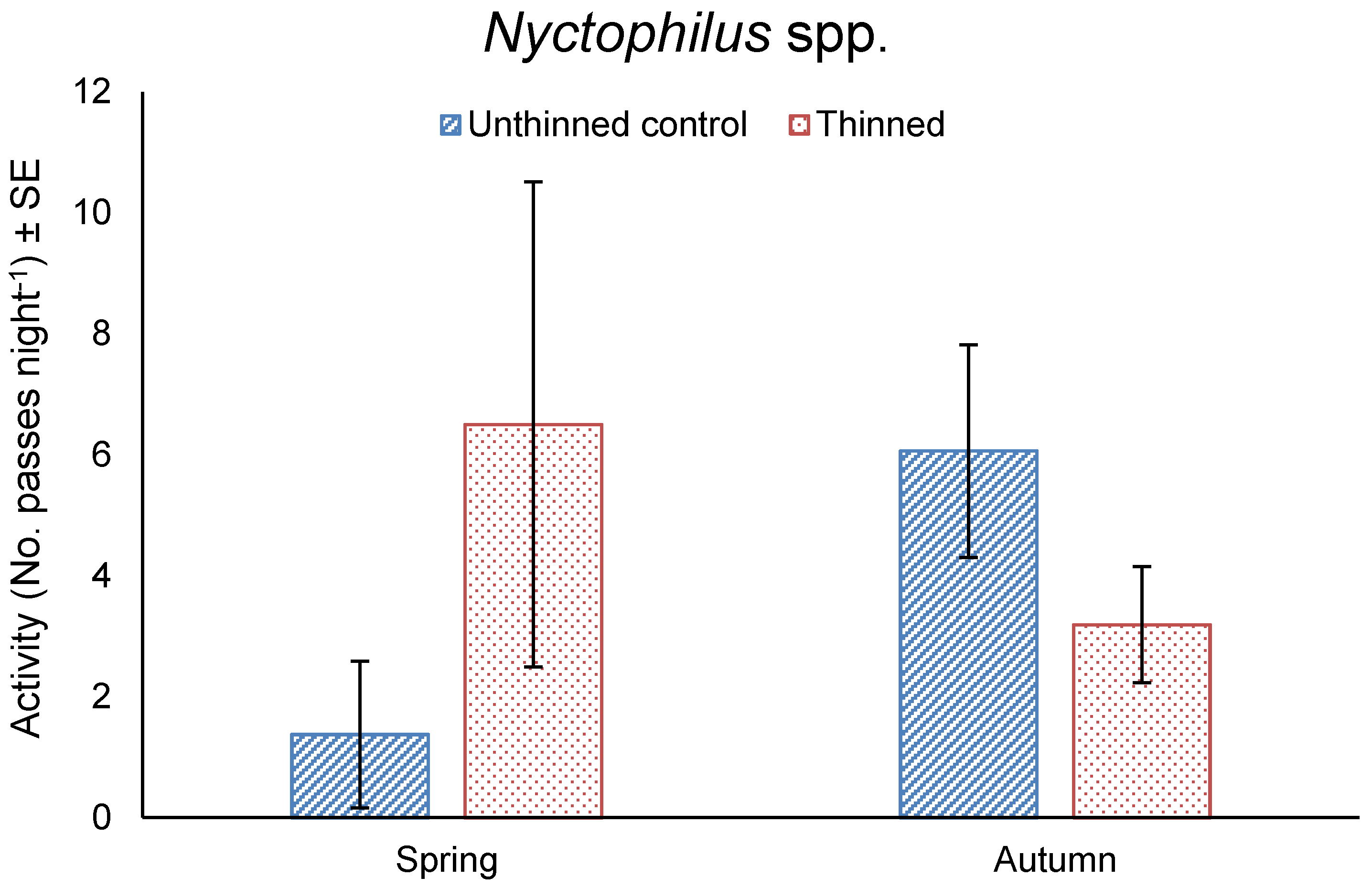
The activity of Nyctophilus spp. did not differ between thinning treatments (F11,1 = 1.095, p = 0.318) or seasons (F11,1 = 1.932, p = 0.192), but was significantly affected by the interaction of treatment by season (F11,1 = 7.055, p = 0.022). Nyctophilus spp. activity was five-times greater in the thinned treatment compared to the unthinned treatment in spring, whereas the activity was two-times greater in the unthinned treatment compared to the thinned treatment in autumn (Figure 6).
The activity of V. vulturnus was ~4 times greater in the thinned treatment compared to the unthinned treatment (F = 4.555, p = 0.056), but did not differ between seasons (F = 0.028, p = 0.871) and was not affected by the interaction of treatment by season (F = 0.003, p = 0.955).
3.5. Harp Trapping Captures
In all, 237 bats were captured in thinned (166) and unthinned (71) sites across both years based on trapping on flyways, traps with Autobat lures and traps without lures (Table 2). The average number of bats captured in the unthinned treatment was 2.0 bats per trap night, whereas 2.6 bats per trap night were captured in the thinned treatment.
To evaluate whether capture rates differed between thinned and unthinned treatments, only trapping data for Autobat traps were analysed. Captures per trap-night for N. corbeni (Wilcoxon W = 16.000, p = 0.554), N. geoffroyi (Wilcoxon W = 13.500, p = 0.163), V. vulturnus (Wilcoxon W = 13.000, p = 0.144) and all bats (Wilcoxon W = 15.000, p = 0.386) were evenly spread across both thinning treatments, whereas capture rates were higher in the unthinned treatment for N. gouldi (Wilcoxon W = 10.000, p = 0.021).
The sex ratio of N. geoffroyi capture rates was biased towards females in thinned regrowth (0.2:0.8; χ2 = 11.1, p = 0.001), but not significantly biased in unthinned regrowth (0.3:0.7, χ2 = 1.3, p = 0.3). Sex ratios for N. gouldi and V. vulturnus showed no significant bias in either thinned (N. gouldi:Ng 0.5:0.5; V. vulturnus (Vv): 0.5:0.5) or unthinned regrowth (Ng 0.5:0.5; Vv 0.6:0.4). Sample sizes of N. corbeni captures were too low for testing (unthinned: 0.7:0.3; thinned: 0.4:0.6).
4. Discussion
Our study of day-roosting and nocturnal activity of multiple bat species in the dry cypress forests of the Pilliga revealed a complex pattern of habitat use across the thinned-unthinned mosaic that was only partially predicted by ecomorphology. Two narrow-space (N. corbeni and N. gouldi) species in the genus Nyctophilus avoided roosting in relatively open, thinned areas, as predicted. Few roosts were found for N. geoffroyi, and these were located in thinned patches or in more open areas (e.g., along a forest road and in a riparian zone), suggesting that this species may be less sensitive to thinning. The edge-space species (V. vulturnus) was found to roost in both unthinned and thinned patches, though sample sizes were low for this species. In contrast, flight activity of Nyctophilus spp. was evenly spread across the forest mosaic, suggesting they actively foraged in both cluttered and uncluttered forest. Systematic trapping allowed this response to be distinguished at a species level with N. gouldi being captured more frequently in unthinned forest, while no difference was found for N. geoffroyi and N. corbeni. The activity, but not capture rate, of V. vulturnus (edge-space species) was higher in thinned than unthinned forest as predicted by ecomorphology. Such varied patterns of habitat use have implications for managing the forest landscape and emphasise the value of heterogeneous landscapes for biodiversity conservation.
Although the experimental nature of this study has its strengths such as including paired thinned and unthinned sites, with consistent thinning practices applied uniformly across replicates, there are also limitations. One of these is the relatively close proximity of thinned and unthinned plots (≥200 m apart). This may mean that thinned and unthinned replicates were not fully independent given the landscape mobility of bats. However, roosts were typically located close to the thinned blocks where bats were caught and tagged, suggesting bats probably focused their activities in this area. Furthermore, limited data from tracking loggers revealed that bats trapped and tagged in a particular thinned block were not logged foraging in other thinned blocks at night. In addition, the relatively low replication of four thinned and four unthinned blocks was a necessary logistical constraint that is characteristic of large-scale field manipulations. This resulted in a low level of power that may have led to type 2 errors, such as not detecting a treatment effect for capture rates for N. geoffroyi when a strong trend towards a higher capture rate in thinned treatments was apparent. However, treatment effects were often detected, suggesting low power was not typically an issue in this study.
4.1. Roost Selection
At the scale of individual trees, small maternity colonies (<10 bats) of all Nyctophilus spp. were typically in trees with a small diameter (<30 cm) and that were usually dead, especially A. luehmannii and Eucalyptus. Maternity colonies were located in hollows and fissures. All three Nyctophilus species overlapped substantially in the attributes of their roost trees. Near identical patterns of roost selection were found for N. corbeni elsewhere in the Pilliga forests [28] and also for N. gouldi in taller, wetter forests in other regions, though larger tree diameters are often preferred [55,56,57,58]. Dead trees are also selected for roosts by N. geoffroyi, though again, larger diameter trees are often used [41,59]. Only a small sample of roosts was found for V. vulturnus, and these were similar to those used by Nyctophilus spp. Roosts of V. vulturnus are also commonly located in dead trees of variable diameter [40,41,60]. Clearly, the retention of dead trees to protect bat roosts is critical during forest thinning [61], yet collateral damage can result due to a lack of knowledge of the importance of smaller dead trees with hollows for roosting bats. This in part resulted in a halving of the hollow tree density in thinned plots compared to unthinned plots. Elsewhere in the Pilliga forests, thinning reduced dead tree density, but had no effect on hollow density or large tree density [61].
A final unexpected observation for roost trees was the frequency with which many had been ring-barked or girdled (removal of cambium layer around a tree with an axe) in the past (26% of roosts). In one portion of our study area (which overlapped a thinned block), ringbarking of small ironbark and buloke was extensive and took place 40 years previously in 1977, leading to an abundance of dead trees with hollows that N. gouldi commonly used. This was the only one of the experimentally-thinned blocks where any roosts were located (two N. geoffroyi and one V. vulturnus), potentially because of the higher dead hollow stem density. Two other roosts located along the northern boundary of this thinned block were used by N. gouldi. Elsewhere in the study area, ringbarked trees were used by N. corbeni. The longevity of ring-barked hardwood trees is in stark contrast to dead conifers in North America, where only 4% of snags used as bat roosts remained standing after 10 years [62]. In that study, snags with large diameter and short height persisted for longer. In general, conifers are likely to decay faster than hardwoods, although white cypress in our study area is termite resistant and so can remain standing for decades and typically does not form hollows.
At the landscape scale, two of the three Nyctophilus species (N. corbeni and N. gouldi) avoided roosting in the open thinned areas, and this has been found previously for N. corbeni in the Pilliga forests [28]. These results are also consistent with N. gouldi avoiding roosting in narrow and open bushland strips in urban areas [57]. Roosts in unthinned areas are typically surrounded by a high density of stems and hollow-bearing trees, which is consistent with previous studies of these species [28,57]. A preference for roosting where stem density (especially dead stems) is high appears to be linked to conservation status. Nyctophilus corbeni is listed as a threatened species in Australia [28]. Although N. gouldi is not listed, genetic studies have revealed that it displays reduced gene flow between populations isolated by larger stretches (>2 km) of agricultural land [63]. It is possible that unsuitable roosting habitat, as well as clearing of foraging habitat could decrease the connectivity and increase the vulnerability of these narrow-space species to habitat fragmentation. For example, small remnants and linear strips of native vegetation are likely to be vulnerable to loss of dead trees for fire wood removal, rendering such habitat unsuitable for roosting. In contrast, N. geoffroyi tended to roost in more open areas within the forest mosaic, including within one thinned block with a scattering of ringbarked dead hollow-bearing trees. Other studies have found the species roosted in large forest blocks, with roosts surrounded by a lower density of stems (176 stems ha−1; [64]) than what has been found for conspecifics (251 stems ha−1 and 400 stems ha−1; [28,57]). Nyctophilus geoffroyi is a common species with a wide distribution in Australia, and genetic studies indicate that it displays no reduced gene flow in fragmented habitats [63]. Vespadelus vulturnus roosts often have reduced canopy cover [40,41,60], but only a single roost was located in thinned areas in our study. This species is widely distributed across a range of habitats, including semi-cleared farmland in south-eastern Australia. Considerably more data are needed on the patterns of roost selection by different bat species in relation to different management treatments [5]. We are aware of just one other study of bat roosts in relation to thinning, which found a less manoeuvrable edge-space species (Lasiurus borealis) roosting in open conditions created by thinning [29]. Based on our results, we predict that roosts of other narrow-space species will be most sensitive to practices that reduce clutter around roost trees.
It is not clear what the adaptive significance of roosting in dense stands is for some narrow-space bats. Dense stands with many hollows provide shelter and options for roost switching on a regular basis. Both of these attributes may be related to predator avoidance [65]. Goannas (Varanidae) are relatively active in the Pilliga forests, and they often climb trees to search hollows for potential prey (authors’ personal observation). However, it is not known whether they are less active in dense stands of trees. Alternatively, roosts in clutter may simply provide roosting options not accessible to other less manoeuvrable species. It also remains unclear as to why a morphologically-similar species (N. geoffroyi) commonly roosts where it is more open.
4.2. Nocturnal Activity
We found that total bat activity was greater in thinned than unthinned areas, and this trend was maintained for V. vulturnus, as measured by the number of echolocation calls, though not capture-rate. Forest thinning is known to be beneficial for a range of bat species due to increased foraging efficiency by echolocation where clutter levels have been reduced [5,10,23,64]. In contrast, flight activity of Nyctophilus spp. was evenly spread across thinned and unthinned forest. Elsewhere in the Pilliga, the activity of Nyctophilus spp. is higher along narrow flyways than clutter in adjacent forest, though wide open flyways were used less [66]. Narrow-space species typically have higher activity levels in forests with higher stem densities [24,25,26], though they are not restricted to clutter [20,67]. For example, N. gouldi has an aspect ratio of 5.77, is slow flying and occasionally uses forest openings [20]. In dense, wet forests, it uses track and off-track sites equally [68]. Interestingly, Brigham et al. (1997) [20] found that habitat use by light tagged bats was indistinguishable for N. gouldi and N. geoffroyi.
Despite very similar morphology and echolocation calls, we found that N. gouldi was captured more than expected in unthinned than thinned regrowth, while N. geoffroyi and Nyctophilus corbeni were captured equally in both treatments. These species-based trapping results (which are not available from ultrasonics due to call similarity) from areas with different amounts of clutter are consistent with other studies of broad habitat use by these species [64]. Nyctophilus geoffroyi can forage among scattered trees in open farmland [41,59] and is less sensitive to urbanisation than N. gouldi [57]. Differences in habitat use have been documented for a number of morphologically-similar species [69,70].
4.3. Management Implications
As an active management practice aiming for an ecological outcome, silvicultural thinning of dense regrowth is a form of ‘renewal ecology’, where goals include both the creation and/or enhancement of landscapes for biodiversity and ecosystem services for humans [1]. Yet, the varied patterns of habitat use by multiple species that we identified emphasise the value of heterogeneous landscapes. In particular, heterogeneity should include a mosaic of thinned areas, but also recognize the value of dense unthinned patches, both to fulfil the requirements of different species [71,72], and to allow for different aspects of a species’ ecology (roosting vs. foraging areas; [73]). The thinned patches in our study (12 ha) were of sufficient size to result in increased flight activity, but the size of unthinned patches required for roosting remains poorly known, though N. corbeni roosts in alleys of unthinned regrowth adjacent to thinned strips [28].
The importance of retaining dead trees as roosts for bats, even those that are relatively small (~20 cm DBH), is a further clear management action emerging from our study (see also [5]). Unexpectedly, we also identified ring-barking as an important contributor to the abundance of dead trees in our study area. This practice was historically used for land-clearing or to remove unwanted trees in forestry areas. Applied as a strictly regulated technique on select species that might occur in high abundance (e.g., buloke), ringbarking may be useful for accelerating hollow development in areas where hollow trees are currently scarce, especially for threatened species such as N. corbeni. This is particularly the case as nest boxes often have limited life spans and effectiveness for threatened species of bats [74,75].
Supplementary Materials
The following are available online at https://www.mdpi.com/1424-2818/10/2/45/s1: Table S1: Morphometric (mass and forearm length) data for study species. Also presented are details about the wing aspect ratio, wing loading, known roost types and diet.
Author Contributions
B.L. and L.G. conceived of and designed the experiments. B.L., L.G., T.B. and D.H. performed the experiments. L.G. analysed the data. B.L. and L.G. wrote the paper.
Funding
This research was funded by the New South Wales government through its Environmental Trust (Grant number 2014/RD/0088) and part-funding by the NSW Office of Environment and Heritage (Murray Ellis).
Acknowledgments
Patrick Tap (Forestry Corporation of NSW) coordinated the thinning experiment in the Pilliga forests that formed the core of our study area and provided comments on a draft of the paper. We thank Sabrina Velasco and Tamara Potter for assistance in the field.
Conflicts of Interest
The authors declare no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript; nor in the decision to publish the results.
References
- Bowman, D.M.; Garnett, S.T.; Barlow, S.; Bekessy, S.A.; Bellairs, S.M.; Bishop, M.J.; Bradstock, R.A.; Jones, D.N.; Maxwell, S.L.; Pittock, J.; et al. Renewal ecology: Conservation for the Anthropocene. Restor. Ecol. 2017, 25, 674–680. [Google Scholar] [CrossRef]
- Montoya, D.; Rogers, L.; Memmott, J. Emerging perspectives in the restoration of biodiversity-based ecosystem services. TREE 2012, 27, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Beaudry, S.; Duchesne, L.C.; Côté, B. Short-term effects of three forestry practices on carabid assemblages in a jack pine forest. Can. J. For. Res. 1997, 27, 2065–2071. [Google Scholar] [CrossRef]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Pastur, G.M.; Messier, C.; et al. Retention forestry to maintain multifunctional forests: A world perspective. BioScience 2012, 62, 633–645. [Google Scholar] [CrossRef]
- Law, B.S.; Park, K.J.; Lacki, M.J. Insectivorous bats and silviculture: Balancing timber production and bat conservation. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Cham, Switzerland, 2016; pp. 105–150. [Google Scholar]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Verschuyl, J.; Riffell, S.; Miller, D.; Wigley, T.B. Biodiversity response to intensive biomass production from forest thinning in North American forests–a meta-analysis. For. Ecol. Manag. 2011, 261, 221–232. [Google Scholar] [CrossRef]
- Fuller, R.J. Searching for biodiversity gains through woodfuel and forest management. J. Appl. Ecol. 2013, 50, 1295–1300. [Google Scholar] [CrossRef]
- Eyre, T.J.; Ferguson, D.J.; Kennedy, M.; Rowland, J.; Maron, M. Long term thinning and logging in Australian cypress pine forest: Changes in habitat attributes and response of fauna. Biol. Conserv. 2015, 186, 83–96. [Google Scholar] [CrossRef]
- Blakey, R.V.; Law, B.S.; Kingsford, R.T.; Stoklosa, J.; Tap, P.; Williamson, K. Bat communities respond positively to large-scale thinning of forest regrowth. J. Appl. Ecol. 2016, 53, 1694–1703. [Google Scholar] [CrossRef]
- McKenzie, N.L.; Rolfe, J.K. Structure of bat guilds in the Kimberley mangroves, Australia. J. Anim. Ecol. 1986, 55, 401–420. [Google Scholar] [CrossRef]
- Aldridge, H.D.J.N.; Rautenbach, I.L. Morphology, echolocation and resource partitioning in insectivorous bats. J. Anim. Ecol. 1987, 56, 763–778. [Google Scholar] [CrossRef]
- Norberg, U.M.; Rayner, J.M. Ecological morphology and flight in bats (Mammalia; Chiroptera): Wing adaptations, flight performance, foraging strategy and echolocation. Phil. Trans. R. Soc. B 1987, 316, 335–427. [Google Scholar] [CrossRef]
- Crome, F.H.J.; Richards, G.C. Bats and gaps: Microchiropteran community structure in a Queensland rain forest. Ecology 1988, 69, 1960–1969. [Google Scholar] [CrossRef]
- Bullen, R.; McKenzie, N.L. Bat airframe design: Flight performance, stability and control in relation to foraging ecology. Aust. J. Zool. 2001, 49, 235–261. [Google Scholar] [CrossRef]
- Hanspach, J.; Fischer, J.; Ikin, K.; Stott, J.; Law, B.S. Using trait-based filtering as a predictive framework for conservation: A case study of bats on farms in southeastern Australia. J. Appl. Ecol. 2012, 49, 842–850. [Google Scholar] [CrossRef]
- Denzinger, A.; Schnitzler, H.U. Bat guilds, a concept to classify the highly diverse foraging and echolocation behaviors of microchiropteran bats. Front. Physiol. 2013, 4, 164. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Purvis, A.; Gittleman, J.L. Biological correlates of extinction risk in bats. Am. Nat. 2003, 161, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Safi, K.; Kerth, G. A comparative analysis of specialization and extinction risk in temperate-zone bats. Cons. Biol. 2004, 18, 1293–1303. [Google Scholar] [CrossRef] [Green Version]
- Brigham, R.M.; Francis, R.L.; Hamdorf, S. Microhabitat use by two species of Nyctophilus bats: A test of ecomorphology theory. Aust. J. Zool. 1997, 45, 553–560. [Google Scholar] [CrossRef]
- Pavey, C.R.; Grunwald, J.E.; Neuweiler, G. Foraging habitat and echolocation behaviour of Schneider’s leafnosed bat, Hipposideros speoris, in a vegetation mosaic in Sri Lanka. Behav. Ecol. Sociobiol. 2001, 50, 209–218. [Google Scholar] [CrossRef]
- Humes, M.L.; Hayes, J.P.; Collopy, M.W. Bat activity in thinned, unthinned, and old-growth forests in western Oregon. J. Wildl. Manag. 1999, 63, 553–561. [Google Scholar] [CrossRef]
- Patriquin, K.J.; Barclay, R.M. Foraging by bats in cleared, thinned and unharvested boreal forest. J. Appl. Ecol. 2003, 40, 646–657. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Mehr, M.; Bässler, C.; Fenton, M.B.; Hothorn, T.; Pretzsch, H.; Klemmt, H.J.; Brandl, R. Aggregative response in bats: Prey abundance versus habitat. Oecologia 2012, 169, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Montemayor, E.; Goulson, D.; Cavin, L.; Wallace, J.M.; Park, K.J. Fragmented woodlands in agricultural landscapes: The influence of woodland character and landscape context on bats and their insect prey. Agric. Ecosys. Environ. 2013, 172, 6–15. [Google Scholar] [CrossRef]
- Blakey, R.V.; Kingsford, R.T.; Law, B.S.; Stoklosa, J. Floodplain habitat is disproportionately important for bats in a large river basin. Biol. Conserv. 2017, 215, 1–10. [Google Scholar] [CrossRef]
- Kunz, T.H.; Lumsden, L. Ecology of cavity and foliage roosting bats. In Bat Ecology; Kunz, T.H., Fenton, M.B., Eds.; University of Chicago Press: Chicago, IL, USA, 2003; pp. 3–89. [Google Scholar]
- Law, B.; Gonsalves, L.; Chidel, M.; Brassil, T. Subtle use of a disturbance mosaic by the south-eastern long-eared bat (Nyctophilus corbeni): An extinction-prone, narrow-space bat. Wildl. Res. 2016, 43, 153–168. [Google Scholar] [CrossRef]
- Perry, R.W.; Thill, R.E.; Carter, S.A. Sex-specific roost selection by adult red bats in a diverse forested landscape. For. Ecol. Manag. 2007, 253, 48–55. [Google Scholar] [CrossRef]
- Lacey, C.J. Factors Influencing Occurrence of Cypress Pine Regeneration in New South Wales; Technical Paper No. 21; Forestry Commission of New South Wales: Sydney, Australia, 1972.
- Lacey, C.J. Silvicultural Characteristics of White Cypress Pine; Research Note No. 26; Forestry Commision of New South Wales: Taree, Australia, 1973.
- Forestry Commission of New South Wales. Managing the State Forests—The Pilliga Management Area; Forestry Commission of NSW: Sydney, Australia, 1988.
- Thompson, W.A.; Eldridge, D.J. White cypress pine (Callitris glaucophylla): A review of its roles in landscape and ecological processes in eastern Australia. Aust. J. Bot. 2005, 53, 555–570. [Google Scholar] [CrossRef]
- Read, J. Recruitment characteristics of the white cypress pine (Callitris glaucophylla) in arid South Australia. Rangel. J. 1995, 17, 228–240. [Google Scholar] [CrossRef]
- Hill, D.A.; Greenaway, F. Effectiveness of an acoustic lure for surveying bats in British woodlands. Mamm. Rev. 2005, 35, 116–122. [Google Scholar] [CrossRef]
- Reinhold, L.; Law, B.; Ford, G.; Pennay, M. Key to the Bat Calls of South-East Queensland and North-East New South Wales; Forest Ecosystem Research and Assessment Technical Paper 2001-07; Department of Natural Resources and Mines: Queensland, Australia, 2001.
- Turbill, C.; Ellis, M. Distribution and abundance of the south-eastern form of the greater long-eared bat Nyctophilus timoriensis. Aust. Mammal. 2006, 28, 1–6. [Google Scholar] [CrossRef]
- Churchill, S. Australian Bats, 2nd ed.; Allen and Unwin: Sydney, Australia, 2008. [Google Scholar]
- Duncan, A.; Baker, G.B.; Montgomery, N. The Action Plan for Australian Bats; Environment Australia: Canberra, Australia, 1999.
- Campbell, S.; Lumsden, L.F.; Kirkwood, R.; Coulson, G. Day roost selection by female little forest bats (Vespadelus vulturnus) within remnant woodland on Phillip Island, Victoria. Wildl. Res. 2005, 32, 183–191. [Google Scholar] [CrossRef]
- Law, B.S.; Chidel, M.; Penman, T. Do young eucalypt plantations benefit bats in an intensive agricultural landscape? Wildl. Res. 2011, 38, 173–187. [Google Scholar] [CrossRef]
- Gonsalves, L.; Law, B.; Webb, C.; Monamy, V. Foraging ranges of insectivorous bats shift relative to changes in mosquito abundance. PLoS ONE 2013, 8, e64081. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, M.G.; Taylor, R.J. Observations on the flight patterns and foraging behavior of Tasmanian bats. Wildl. Res. 1986, 13, 427–432. [Google Scholar] [CrossRef]
- Rhodes, M.P. Assessment of sources of variance and patterns of overlap in microchiropteran wing morphology in southeast Queensland, Australia. Can. J. Zool. 2002, 80, 450–460. [Google Scholar] [CrossRef]
- Rolls, E. A Million Wild Acres: Two Hundred Years of Man and an Australian Forest; Thomas Nelson: Melbourne, Australia, 1981. [Google Scholar]
- van Kempen, E. A History of the Pilliga Cypress Pine Forests; State Forests of New South Wales: Pennant Hills, Australia, 1997.
- Forestry Commission of NSW. Management Plan for the Pilliga Management Area; Forestry Commission of New South Wales: Sydney, Australia, 1986. [Google Scholar]
- Binns, D.; Beckers, D. Floristic patterns in the Pilliga. In Perfumed Pineries: Environmental History of Australia’s Callitris Forests; Dargavel, J., Hart, D., Libbis, B., Eds.; Australian National University: Canberra, Australia, 2001; pp. 104–110. [Google Scholar]
- Whipp, R.K.; Lunt, I.D.; Spooner, P.G.; Bradstock, R.A. Changes in forest structure over 60 years: Tree densities continue to increase in the Pilliga forests, New South Wales, Australia. Aust. J. Bot. 2012, 60, 1–8. [Google Scholar] [CrossRef]
- Adams, M.D.; Law, B.S.; Gibson, M.S. Reliable automation of bat call identification for eastern New South Wales, Australia, using classification trees and AnaScheme software. Acta Chiropterol. 2010, 12, 231–245. [Google Scholar] [CrossRef]
- Tidemann, C.R.; Woodside, D.P. A collapsible bat-trap and a comparison of results obtained with the trap and with mist-nets. Wildl. Res. 1978, 5, 355–362. [Google Scholar] [CrossRef]
- Gibbons, P.; Lindenmayer, D.B.; Barry, S.C.; Tanton, M.T. Hollow formation in eucalypts from temperate forests in southeastern Australia. Pac. Conserv. Biol. 2000, 6, 218–228. [Google Scholar] [CrossRef]
- Pollard, J.H. On distance estimators of density in randomly distributed forests. Biometrics 1971, 27, 991–1002. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v6; PRIMER-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- Lunney, D.; Barker, J.; Priddel, D.; Oconnell, M. Roost Selection by Goulds Long-Eared Bat, Nyctophilus gouldi Tomes (Chiroptera, Vespertilionidae), in Logged Forest on the South Coast of New South-Wales. Wildl. Res. 1988, 15, 375–384. [Google Scholar] [CrossRef]
- Webala, P.W.; Craig, M.D.; Law, B.S.; Wayne, A.F.; Bradley, J.S. Roost site selection by southern forest bat Vespadelus regulus and Gould’s long-eared bat Nyctophilus gouldi in logged jarrah forests; south-western Australia. For. Ecol. Manag. 2010, 260, 1780–1790. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Law, B.; Banks, P.B. Roost selection in suburban bushland by the urban sensitive bat Nyctophilus gouldi. J. Mamm. 2013, 94, 307–319. [Google Scholar] [CrossRef]
- Burgar, J.M.; Craig, M.D.; Stokes, V.L. The importance of mature forest as bat roosting habitat within a production landscape. For. Ecol. Manag. 2015, 356, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Lumsden, L.F.; Bennett, A.F.; Silins, J.E. Selection of roost sites by the lesser long-eared bat (Nyctophilus geoffroyi) and Gould’s wattled bat (Chalinolobus gouldii) in south-eastern Australia. J. Zool. 2002, 257, 207–218. [Google Scholar] [CrossRef]
- Rueegger, N.; Law, B.; Goldingay, R. Interspecific differences and commonalities in maternity roosting by tree cavity-roosting bats over a maternity season in a timber production landscape. PLoS ONE 2018, 13, e0194429. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Gonsalves, L.; Law, B.; Melville, G.; Toole, I.; Brassil, T.; Tap, P. The effect of thinning on structural attributes of a low rainfall forest in eastern Australia. For. Ecol. Manag. 2018, 409, 571–583. [Google Scholar] [CrossRef]
- Lacki, M.J.; Baker, M.D.; Johnson, J.S. Temporal dynamics of roost snags of long-legged myotis in the Pacific Northwest, USA. J. Wildl. Manag. 2012, 76, 1310–1316. [Google Scholar] [CrossRef]
- Fuller, N.C. Landscape Genetics and Sociobiology of Gould’s Long-Eared (Nyctophilus gouldi) and the Lesser Long-Eared Bat (N. geoffroyi) in Fragmented Populations of South-Eastern Australia. Ph.D Thesis, University of Adelaide, Adelaide, Australia, 2013. [Google Scholar]
- Lumsden, L.F.; Bennett, A.F.; Silins, J.E. Location of roosts of the lesser long-eared bat Nyctophilus geoffroyi and Gould’s wattled bat Chalinolobus gouldii in a fragmented landscape in south-eastern Australia. Biol. Conserv. 2002, 106, 237–249. [Google Scholar] [CrossRef]
- Lewis, S.E. Roost fidelity of bats: A review. J. Mammal. 1995, 76, 481–496. [Google Scholar] [CrossRef]
- Law, B.; Tap, P.; Chidel, M. Bat activity in ephemeral stream-beds in the Pilliga forests: Clarifying the importance of flyways and buffer widths in open forest and woodland. In The Biology and Conservation of Australasian Bats; Law, B., Eby, P., Lunney, D., Lumsden, L., Eds.; Royal Zoological Society of New South Wales: Sydney, Australia, 2011; pp. 308–321. [Google Scholar]
- Fenton, M.B. The foraging behaviour and ecology of animal-eating bats. Can. J. Zool. 1990, 68, 411–422. [Google Scholar] [CrossRef]
- Law, B.S.; Chidel, M. Tracks and riparian zones facilitate the use of Australian regrowth forest by insectivorous bats. J. Appl. Ecol. 2002, 39, 605–617. [Google Scholar] [CrossRef] [Green Version]
- Arlettaz, R. Habitat selection as a major resource partitioning mechanism between the two sympatric sibling bat species Myotis myotis and Myotis blythii. J. Anim. Ecol. 1999, 68, 460–471. [Google Scholar] [CrossRef]
- McConville, A.; Law, B.; Penman, T.; Mahony, M. Contrasting habitat use of morphologically similar bat species with differing conservation status in south-eastern Australia. Aust. Ecol. 2013, 39, 83–94. [Google Scholar] [CrossRef]
- Gonsalves, L.; Law, B.; Brassil, T.; Waters, C.; Toole, I.; Tap, P. Ecological outcomes for multiple taxa from silvicultural thinning of regrowth forest. For. Ecol. Manag. 2018, 425, 177–188. [Google Scholar] [CrossRef]
- Gonsalves, L.; Law, B.; Blakey, R. Experimental evaluation of the initial effects of large-scale thinning on structure and biodiversity of river red gum (Eucalyptus camaldulensis) forests. Wildl. Res. 2018, in press. [Google Scholar]
- Law, B.S.; Dickman, C.R. The use of habitat mosaics by terrestrial vertebrate fauna: Implications for conservation and management. Biodivers. Conserv. 1998, 7, 323–333. [Google Scholar] [CrossRef]
- Kavanagh, R.; Law, B.; Lemckert, F.; Stanton, M.; Chidel, M.; Brassil, T.; Towerton, A.; Penman, T. Conservation Value of Eucalypt Plantations Established for Wood Production and Multiple Environmental Benefits in Agricultural Landscapes; Final Report for NAP/NHT2 Eucalypt Plantations project. SLA 0013, R3 NAP; Industry & Investment NSW, Forest Science Centre: West Pennant Hills, Australia, 2010.
- Rueegger, N. Bat boxes—A review of their use and application, past, present and future. Acta Chiropterol. 2016, 18, 279–299. [Google Scholar] [CrossRef]
Figure 1.
Satellite image (Sentinel-2) showing the location of roost trees (filled circles) and four 12-ha thinned blocks (red border) where radio-tagged bats were trapped. Four unthinned controls (green border) are also shown. Larger areas of commercial timber harvesting are also apparent in the southeast and northwest.
Figure 1.
Satellite image (Sentinel-2) showing the location of roost trees (filled circles) and four 12-ha thinned blocks (red border) where radio-tagged bats were trapped. Four unthinned controls (green border) are also shown. Larger areas of commercial timber harvesting are also apparent in the southeast and northwest.
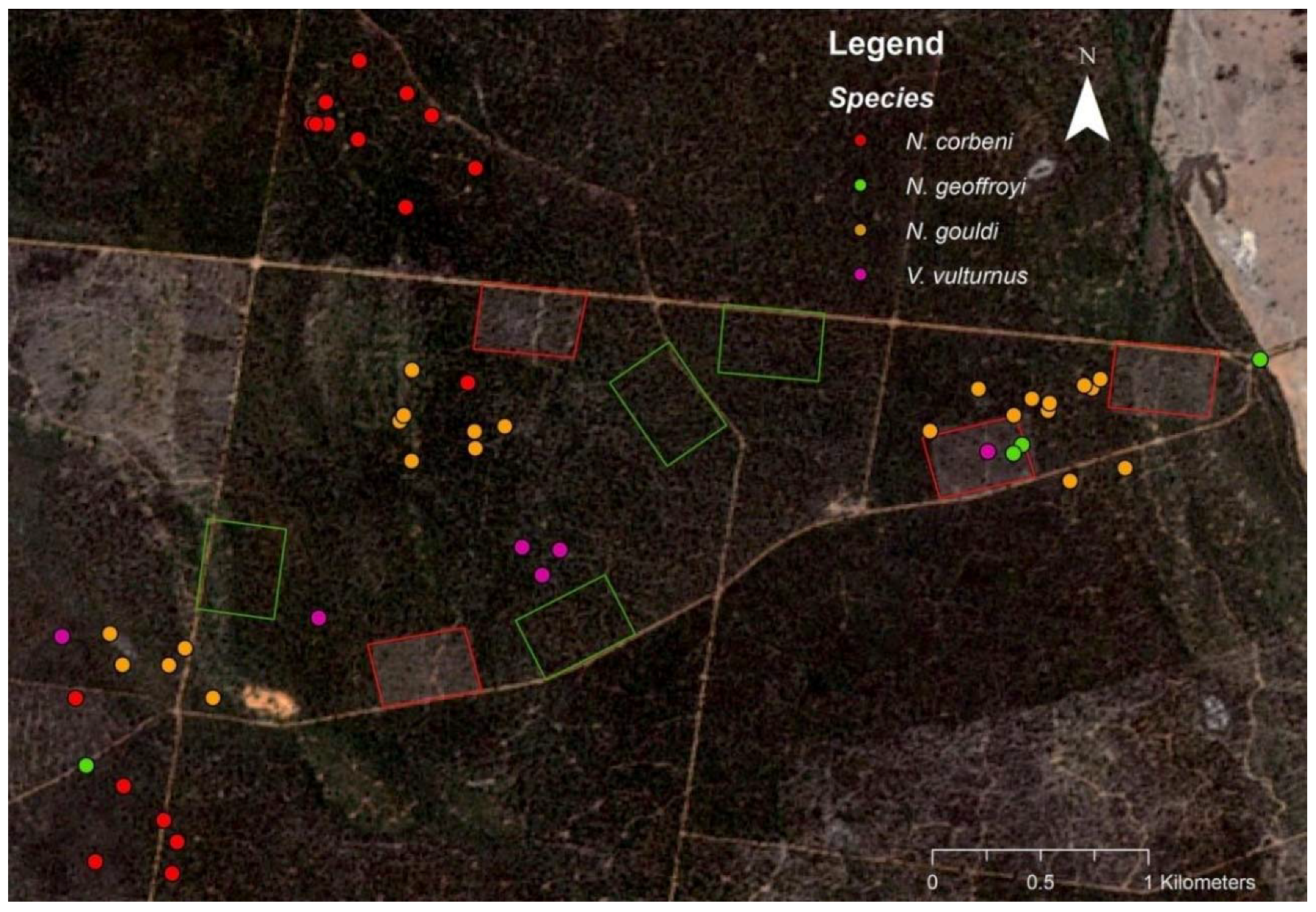
Figure 2.
Percentage of roost trees of each species that were represented by buloke, cypress, eucalypts and other trees (2016 and 2017) plus their local availability in the proximity of roost trees.
Figure 2.
Percentage of roost trees of each species that were represented by buloke, cypress, eucalypts and other trees (2016 and 2017) plus their local availability in the proximity of roost trees.

Figure 3.
Percentage of roosts of each species that were located in a hollow, fissure, under bark and hollow/fissure (classification uncertain) (2016 and 2017).
Figure 3.
Percentage of roosts of each species that were located in a hollow, fissure, under bark and hollow/fissure (classification uncertain) (2016 and 2017).
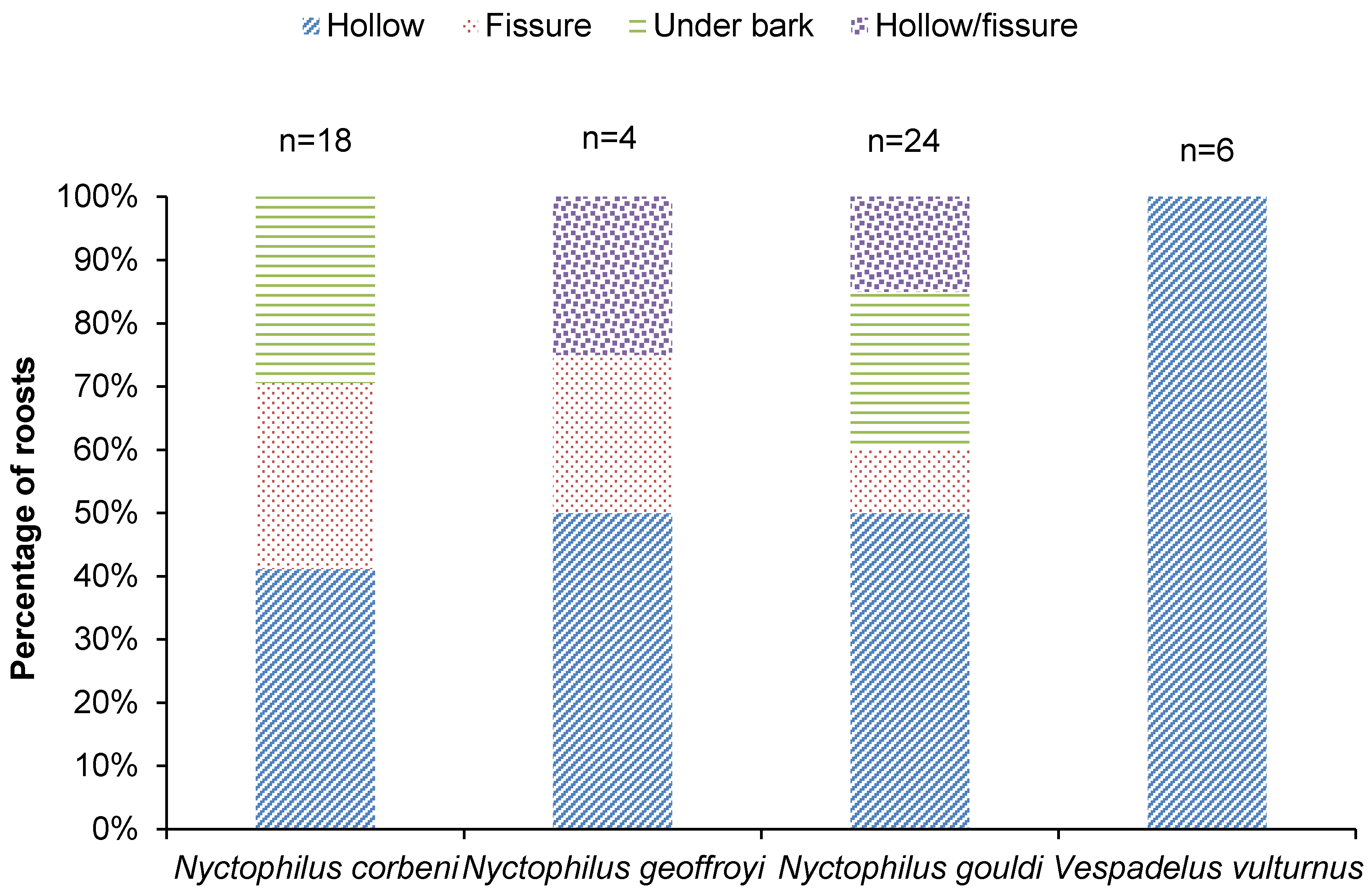
Figure 4.
Canonical analysis of principal (CAP) coordinates illustrating characteristics of trees used for roosting and local available trees. Solid red and cyan circles indicate living and dead available trees, respectively. Blue crosses indicate trees used for roosting by V. vulturnus. Solid light green, open dark green and solid black triangles indicate trees used for roosting by N. corbeni, N. geoffroyi and N. gouldi, respectively.
Figure 4.
Canonical analysis of principal (CAP) coordinates illustrating characteristics of trees used for roosting and local available trees. Solid red and cyan circles indicate living and dead available trees, respectively. Blue crosses indicate trees used for roosting by V. vulturnus. Solid light green, open dark green and solid black triangles indicate trees used for roosting by N. corbeni, N. geoffroyi and N. gouldi, respectively.
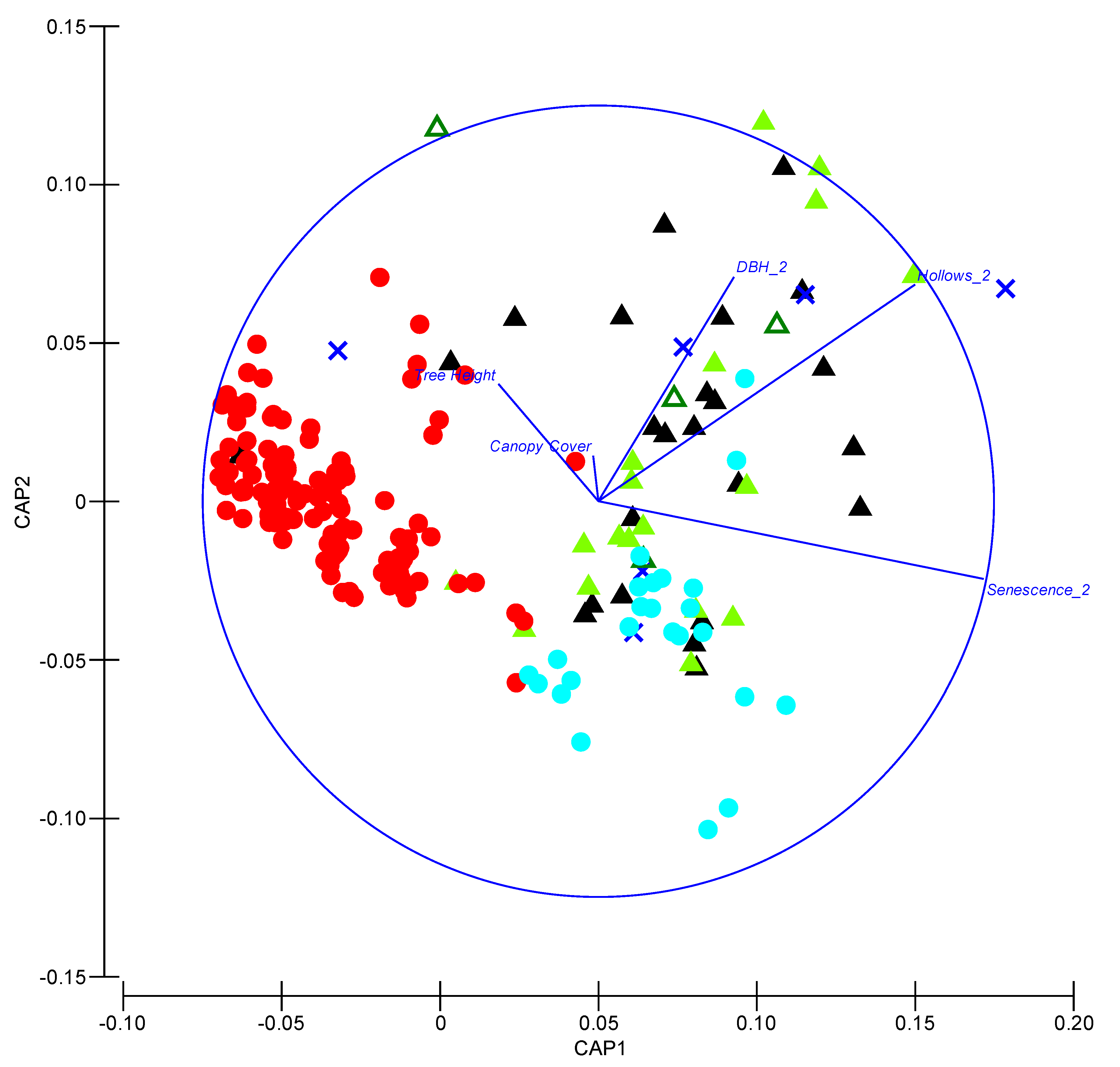
Figure 5.
Photographs illustrating the density of small stems (<10 cm DBHOB) in (a) unthinned control sites, (b) thinned sites, (c) areas around roost trees used by N. corbeni and N. gouldi and (d) areas around roost trees used by N. geoffroyi and V. vulturnus located in thinned sites. Pink flagging tape (c) and the arrow (d) indicate the roost tree.
Figure 5.
Photographs illustrating the density of small stems (<10 cm DBHOB) in (a) unthinned control sites, (b) thinned sites, (c) areas around roost trees used by N. corbeni and N. gouldi and (d) areas around roost trees used by N. geoffroyi and V. vulturnus located in thinned sites. Pink flagging tape (c) and the arrow (d) indicate the roost tree.


Figure 6.
Mean total nightly activity ± SE of Nyctophilus spp. in unthinned and thinned treatments in spring 2016 and autumn 2017.
Figure 6.
Mean total nightly activity ± SE of Nyctophilus spp. in unthinned and thinned treatments in spring 2016 and autumn 2017.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of trapping and roost-tree data collected for four species during radio-tracking over two maternity seasons (2016 and 2017). DBHOB, diameter at breast height over bark.
Table 1.
Summary of trapping and roost-tree data collected for four species during radio-tracking over two maternity seasons (2016 and 2017). DBHOB, diameter at breast height over bark.
| Trapping/Roost Data | 2016 | 2017 |
|---|---|---|
| No. of bats trapped | 141 | 94 |
| No. of bats tracked | 12 | 9 |
| Species tracked | Nc (3), Nge (2), Ngo (4), Vv (3) | Nc (3), Nge (2), Ngo (2), Vv (2) |
| No. of roost trees | 21 | 31 |
| Roost tree DBHOB (cm) | 27.3 ± 1.9 | 31.6 ± 4.4 |
| Roost tree height (m) | 15.1 ± 1.4 | 11.9 ± 1.2 |
| Dead roost trees (% of all roost trees) | 76.2 | 93.1 |
| Average length of stay at roosts (days) | 1.3 ± 0.2 | 1.3 ± 0.2 |
Nc = Nyctophilus corbeni, Nge = Nyctophilus geoffroyi, Ngo = Nyctophilus gouldi, Vv = Vespadelus vulturnus. Trapping data include trap sites that were located in experimental thinned and unthinned treatments, including traps on flyways that were not paired for comparison with traps set with an acoustic lure.
Table 2.
Harp trapping data for experimental unthinned and thinned treatments over two years of trapping (2016 and 2017). Grey shading indicates species that were radio-tracked.
Table 2.
Harp trapping data for experimental unthinned and thinned treatments over two years of trapping (2016 and 2017). Grey shading indicates species that were radio-tracked.
| Species | Unthinned (n = 35 Trap Nights) | Thinned (n = 65 Trap Nights) |
|---|---|---|
| Chalinolobus gouldii | 7 | 12 |
| Chalinolobus picatus | 0 | 3 |
| Mormopterus petersi | 0 | 1 |
| M. planiceps | 1 | 2 |
| Nyctophilus corbeni | 3 | 8 |
| N. geoffroyi | 7 | 36 |
| N. gouldi | 43 | 23 |
| Saccolaimus flaviventris | 0 | 1 |
| Scotorepens balstoni | 0 | 5 |
| S. greyii | 3 | 27 |
| Scotorepens sp. | 0 | 6 |
| Vespadelus vulturnus | 7 | 42 |
| All bats | 71 | 166 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Law, B.; Gonsalves, L.; Brassil, T.; Hill, D. Does Thinning Homogenous and Dense Regrowth Benefit Bats? Radio-Tracking, Ultrasonic Detection and Trapping. Diversity 2018, 10, 45. https://doi.org/10.3390/d10020045
AMA Style
Law B, Gonsalves L, Brassil T, Hill D. Does Thinning Homogenous and Dense Regrowth Benefit Bats? Radio-Tracking, Ultrasonic Detection and Trapping. Diversity. 2018; 10(2):45. https://doi.org/10.3390/d10020045
Chicago/Turabian StyleLaw, Bradley, Leroy Gonsalves, Traecey Brassil, and David Hill. 2018. "Does Thinning Homogenous and Dense Regrowth Benefit Bats? Radio-Tracking, Ultrasonic Detection and Trapping" Diversity 10, no. 2: 45. https://doi.org/10.3390/d10020045
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.