Water and Muscle Contraction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- Describe the non-ideal behaviour of the myosin filaments suspensions.
- Mimic the behaviour of detached and of attached cross-bridges.
- Reason on the force-length constant.
- Describe the osmotic properties of myosin subfragnent-1.
- Describe the effect of protein osmotic pressure on the stiffness of the attached cross-bridges and on the contractile force.
- Define the power stroke.
- Introduce viscosity, an inseparable partner of muscle contraction.
2. Results
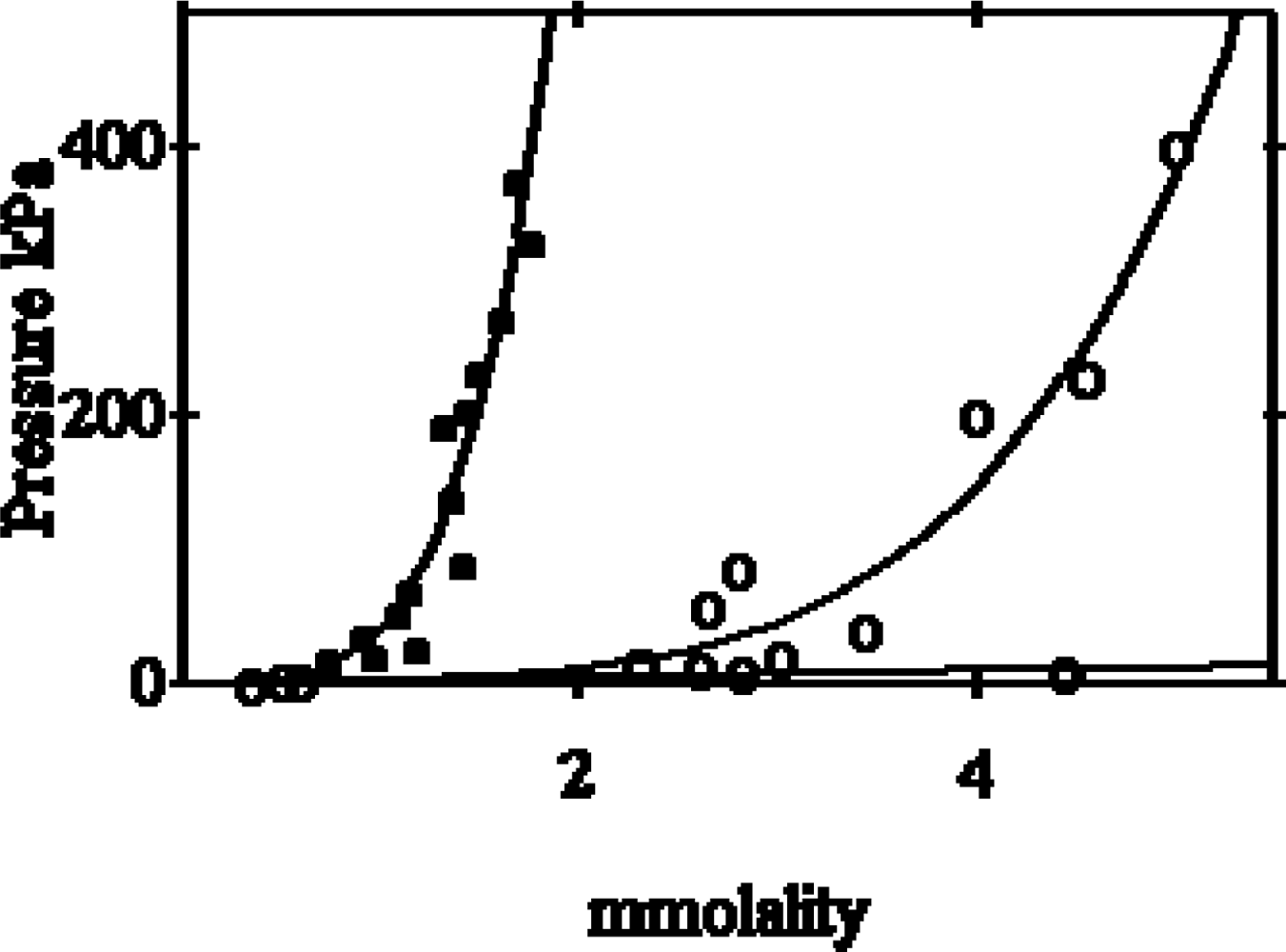
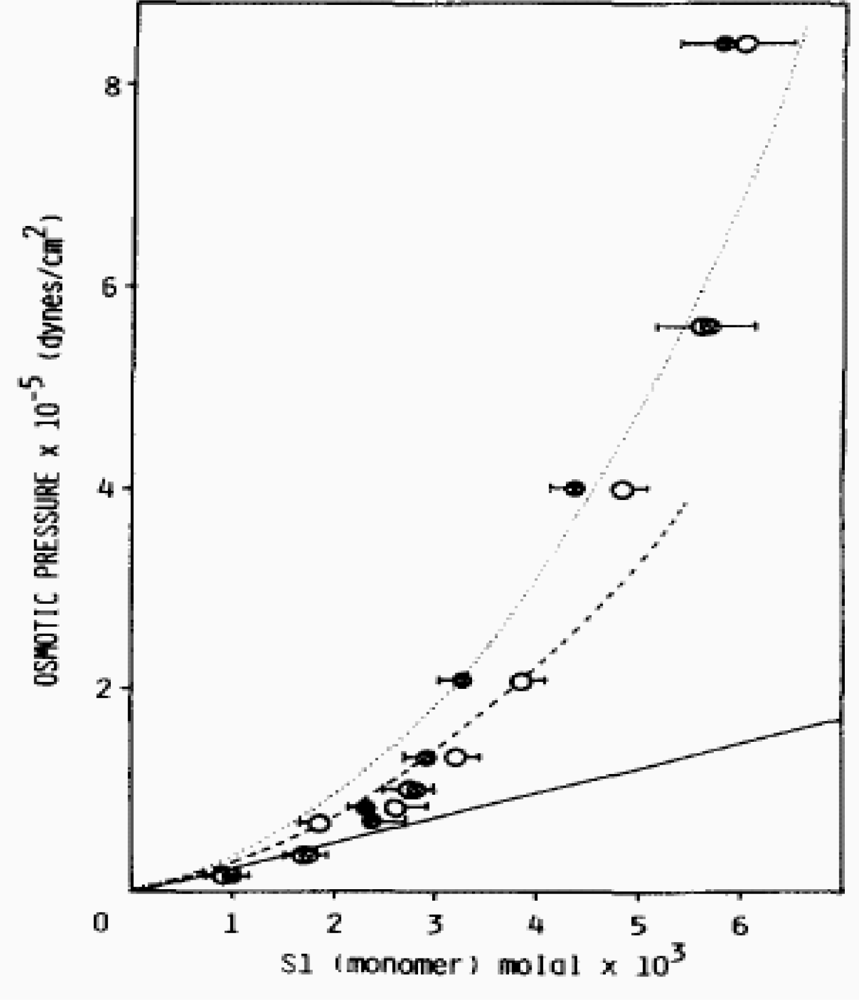
2.1. Non-ideal behaviour of the myosin filaments suspensions
2.2. Mimicking the behaviour of the detached and attached cross-bridges
2.3. The force-length constant
- Beyond a given myosin concentration (or a given protein osmotic pressure), any change of the volume of the solution is accompanied by a change of the volume of the hydrated filament, thus of the radius of the hydrated filament. This sets a relationship between the molality of myosin and the radius of the hydrated filament.
- An equivalence can be set between pressure-volume work and elastic stress and, on the assumption that stress is mostly localized to the cross-bridges, the dependence of cross-bridge distortion on myosin concentration (or protein osmotic pressure) can be calculated.
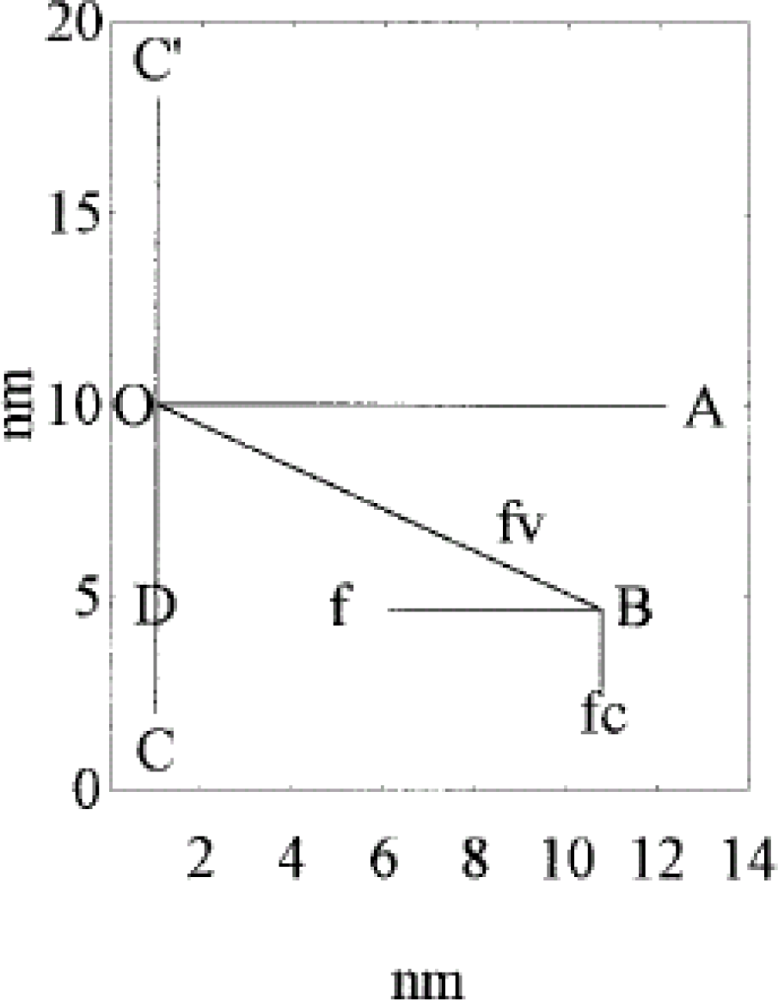
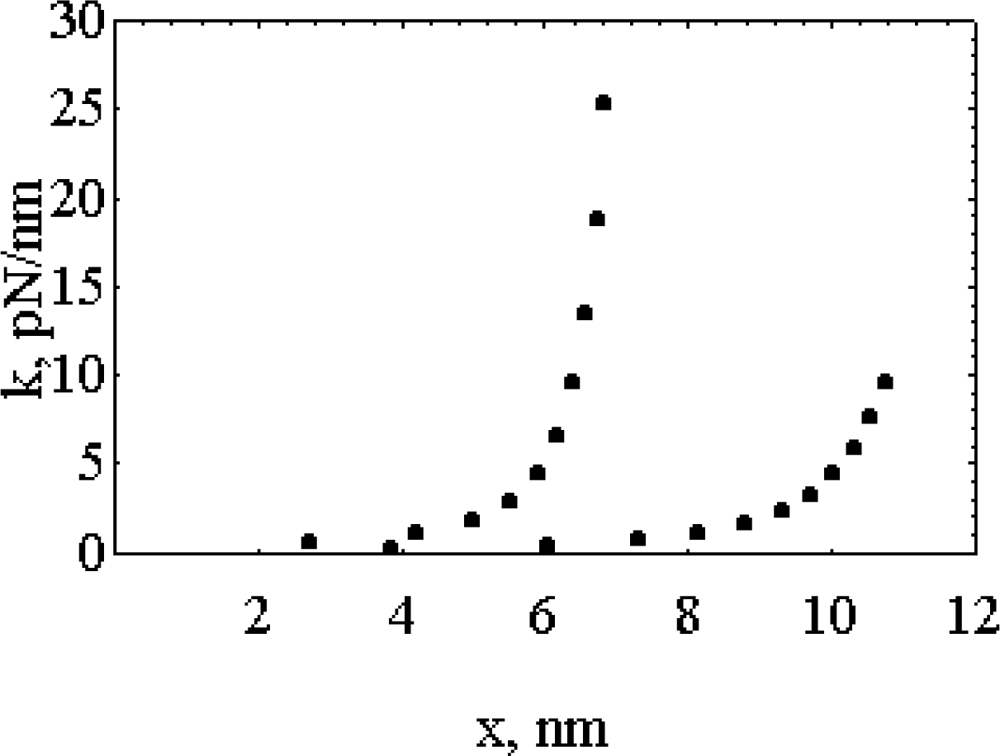
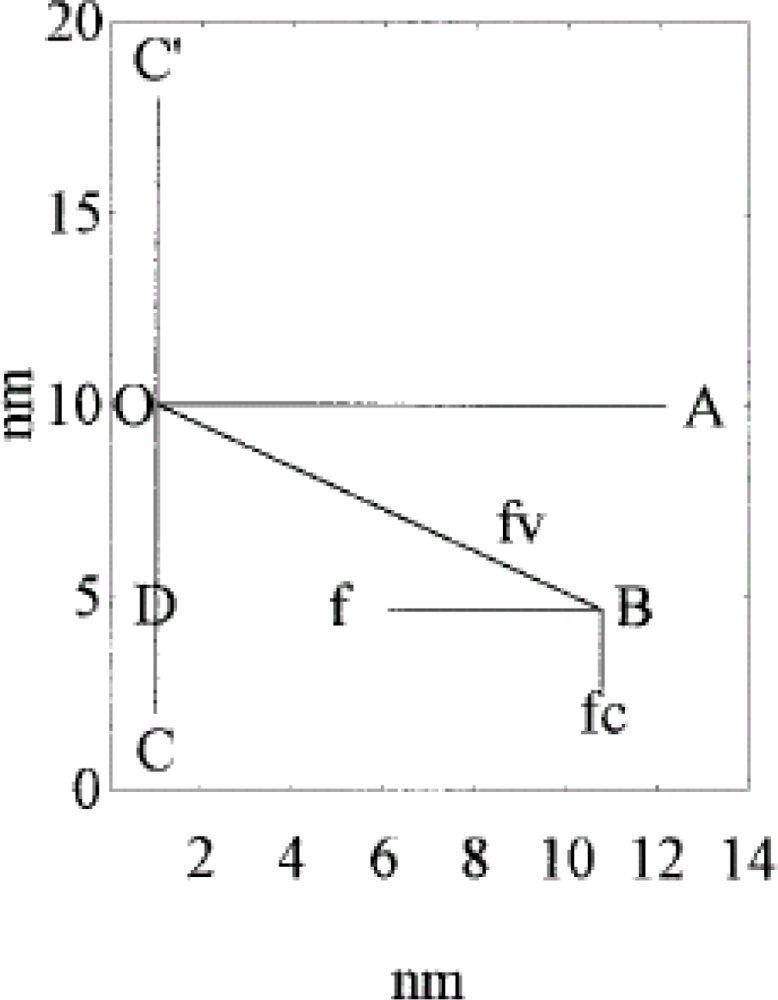
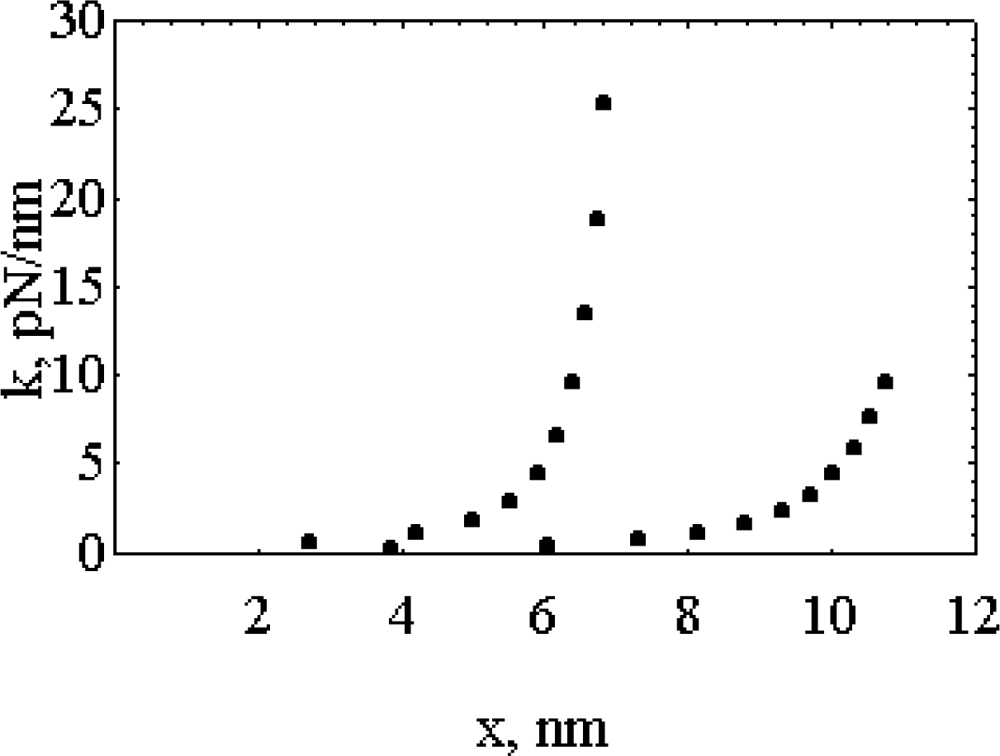
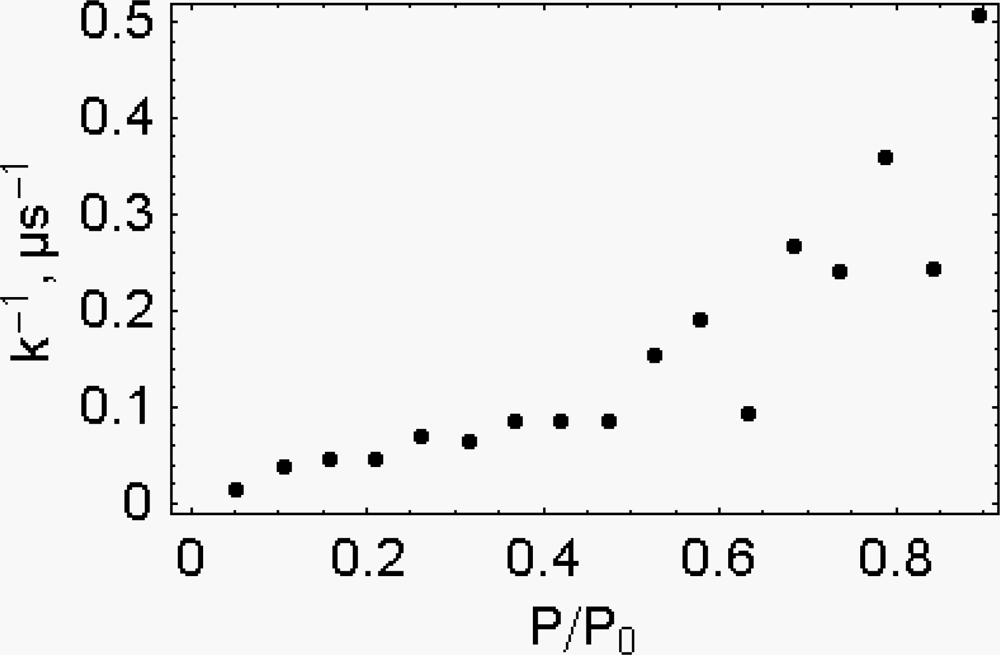
- It is found that, k, the “force length constant” increases significantly with cross-bridge distortion, x, and that the increase depends on the length of the rotating arm. Two models of the rotating arm are selected (Figure 4). The first model assumes that the rotating arm equals r0. In this case r = r0 × sin(a). The second model assumes that the length of the rotating arm is half of the difference between the cross-bridge diameter (30.1 nm) and the shaft diameter (15 nm), thus equals 7.55 nm. In this case sin(a) = (r − b)/(r0− b), where b = r0 − 7.55 nm. By increasing protein osmotic pressure from 18 kPa to 50 kPa, the likely range of protein osmotic pressure in muscle, k, increases from 0.5 to 1.39 pN/nm for model I (rotating arm 15.05 nm) and from 0.79 to 3.48 pN/nm for model II (rotating arm 7.5 nm) (Figure 5).
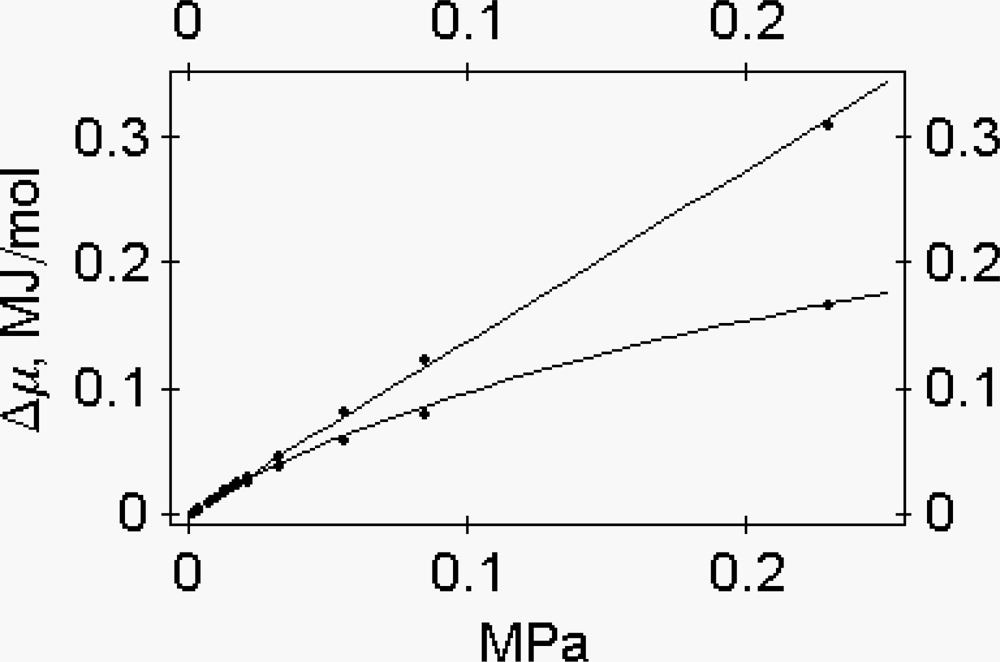
- At constant protein osmotic pressure, if an external, non-osmotic force, parallel to the filament axis, is applied to cross-bridges, these structures are deformed and the water activity coefficient is altered. As a consequence, in muscle, passive and active shortening of the sarcomere is expected to promote the change of the water-water and of the water-protein interactions. We thus depict muscle contraction as a chemo-osmo-elastic transduction, where the analysis of the energy partition during the power stroke requires consideration of the osmotic factor in addition to the chemo-elastic ones [7].
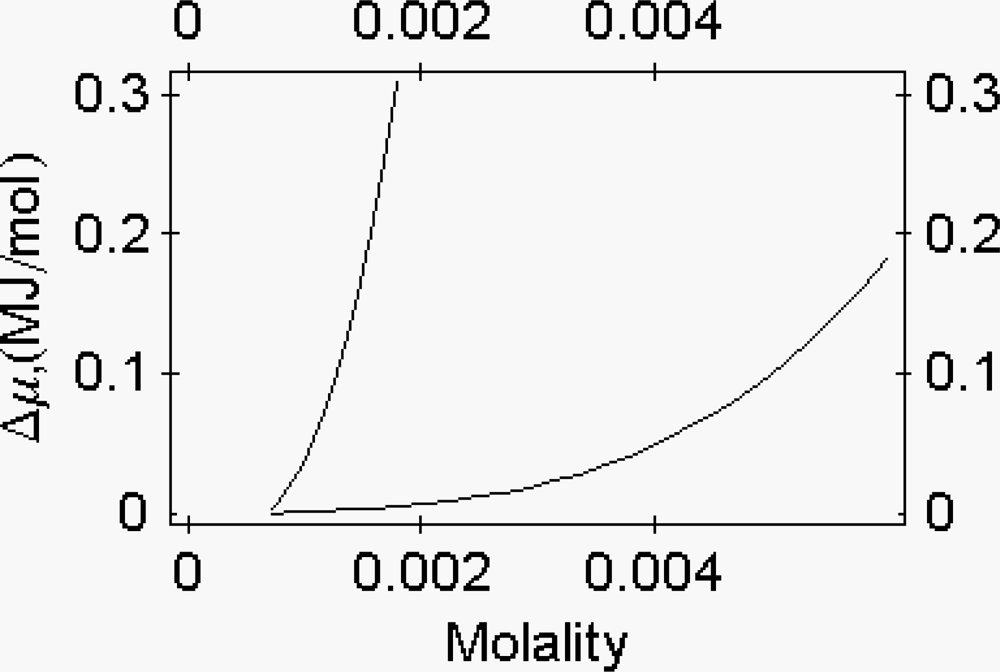
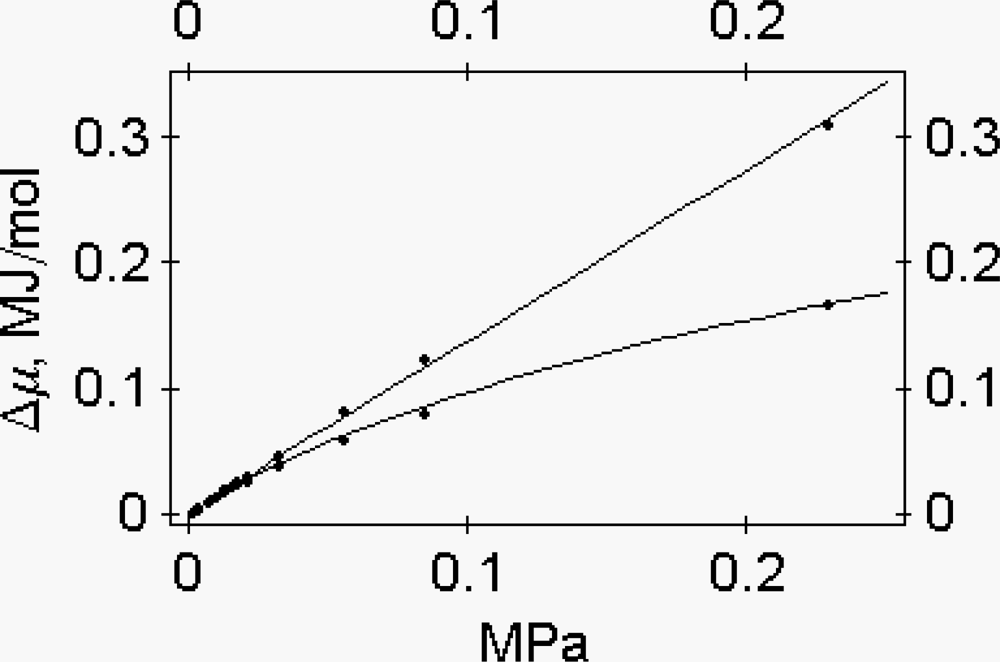
2.4. The osmotic properties of myosin subfragment-1
2.5. Protein osmotic pressure, stiffness of the attached cross-bridges and contractile force
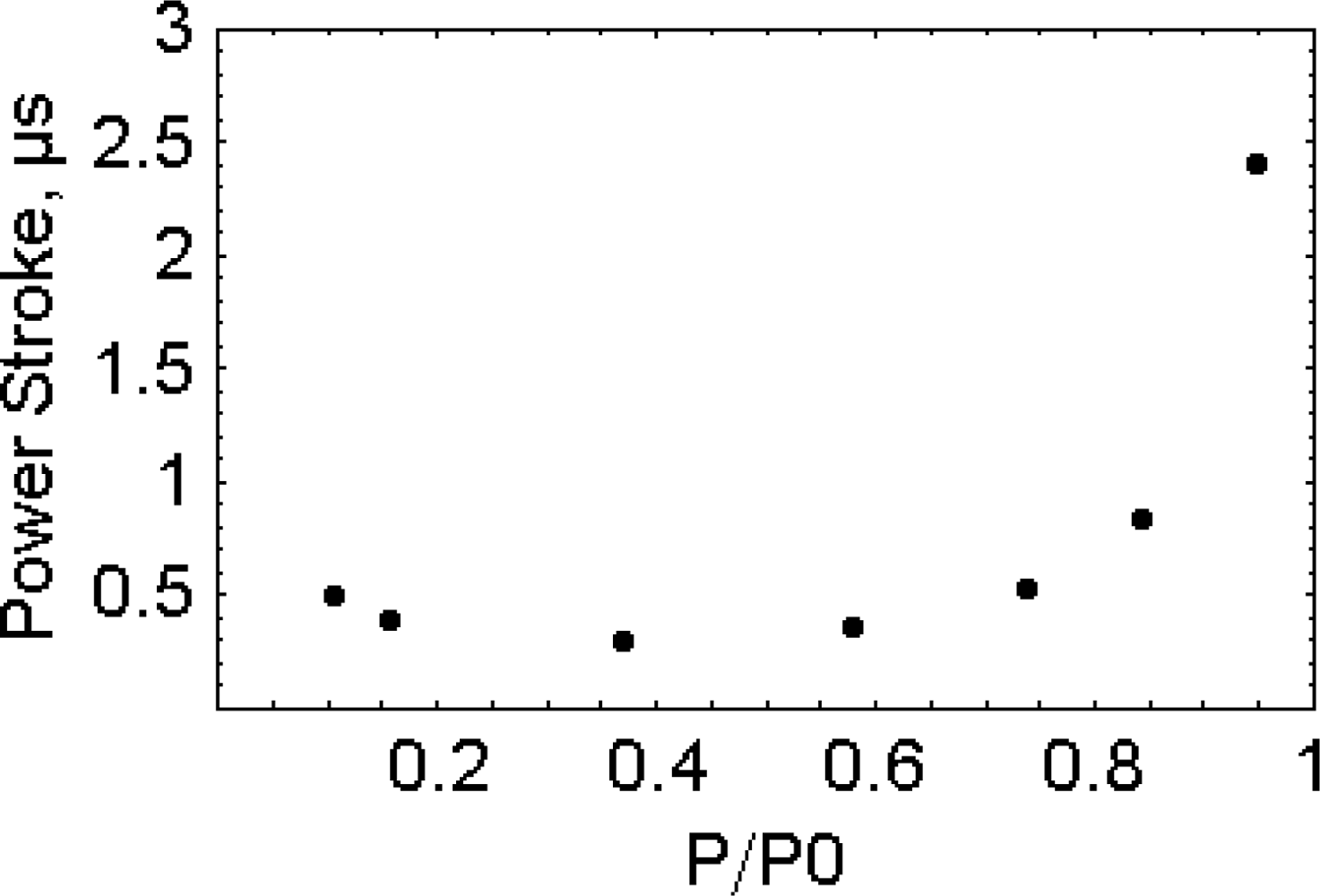
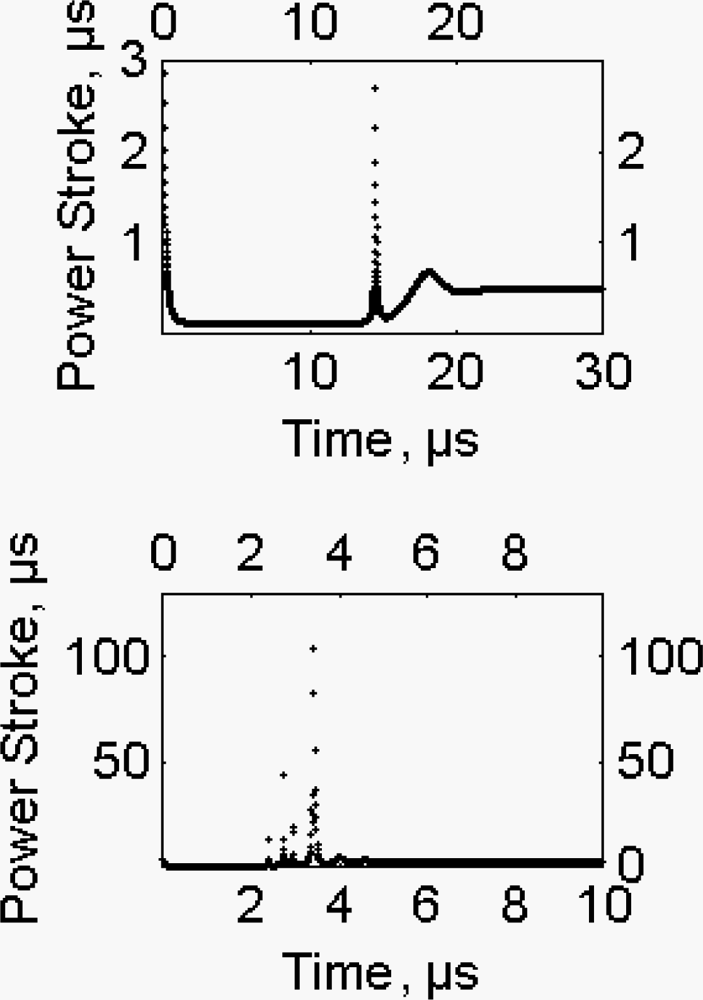
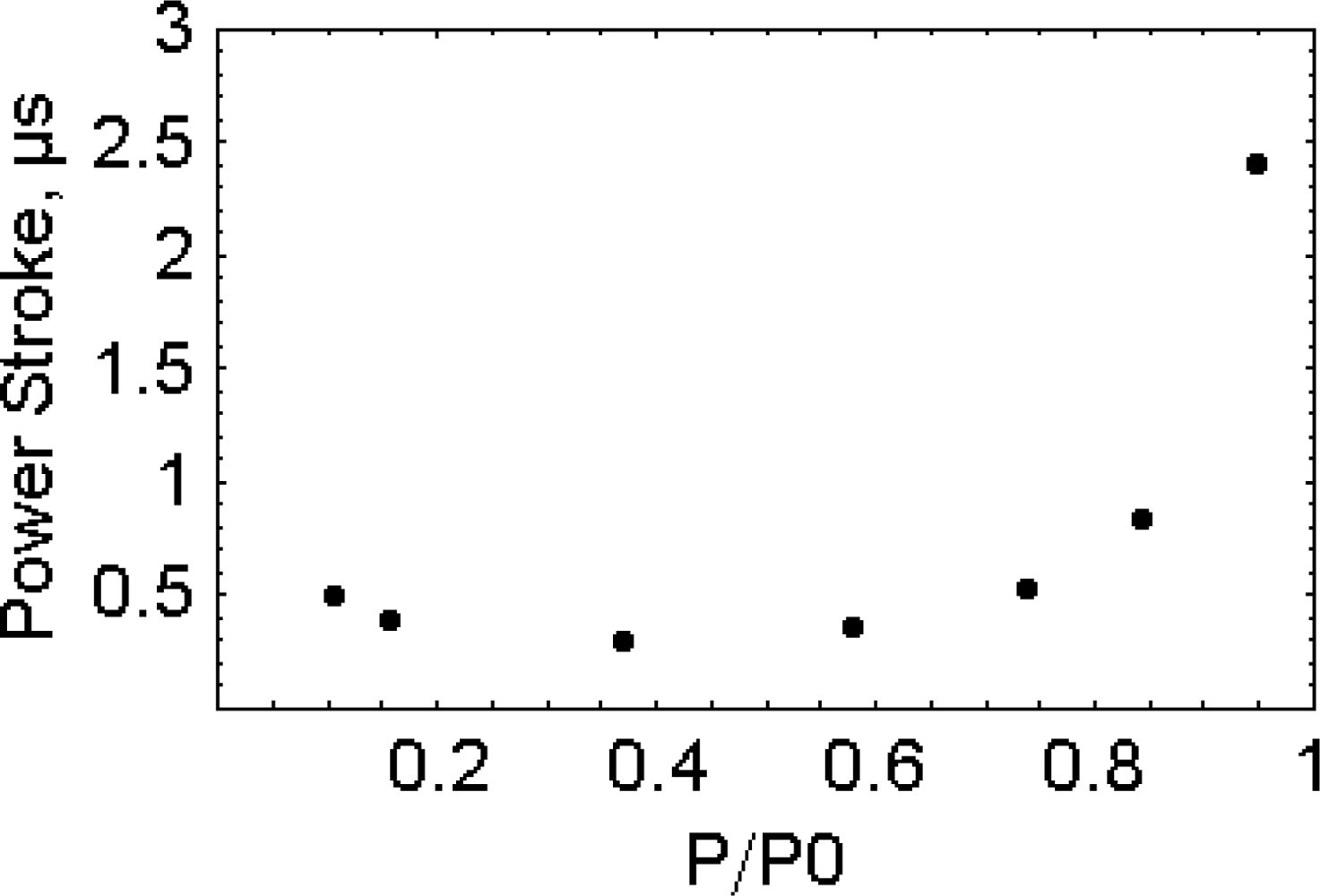
2.6. The working stroke
2.7. Viscosity, an inseparable partner of muscle contraction
3. Conclusions
3.1. The non ideality of the contractile system
3.2. The viscous properties of the system
3.3. The response time of the system
Acknowledgments
Appendix A
Ideal and non ideal solutions
The chemical potential of a protein as a function of the osmotic pressure
Appendix B
Calculation of the interfilament distance of the actin filament and of the S1-decorated actin filament
Appendix C
The compression of the actin filament subjected to an osmotic stress
References
- Huxley, AF; Niedergerke, R. Structural changes in muscle during contraction. Nature 1954, 173, 971–973. [Google Scholar]
- Huxley, HE; Hanson, EJ. Changes in the cross-striation of muscle during contraction and stretch and their strucrural interpretations. Nature 1954, 173, 973–976. [Google Scholar]
- Lymn, RW; Taylor, EW. Mechanism of adenosine triphosphate hydrolysis by actomyosin. Biochemistry 1971, 10, 4617–4624. [Google Scholar]
- Trentham, DR; Eccleston, JF; Bagshaw, CR. Kinetic analysis of ATPase mechanisms. Q. Rev. Biophys 1976, 9, 217–281. [Google Scholar]
- Grazi, E. A highly non ideal solution: the contractile system of skeletal muscle. Eur. Biophys. J 2000, 29, 535–541. [Google Scholar]
- Grazi, E; Di Bona, C. Viscosity as an inseparable partner of muscle contraction. J. Theor. Biol 2006, 242, 853–861. [Google Scholar]
- Grazi, E; Cintio, O. Thermodynamic features of myosin filament suspensions: Implications for the modeling of muscle contraction. Biophys. J 2001, 81, 313–320. [Google Scholar]
- Grazi, E; Adami, R; Cintio, O; Cuneo, P; Magri, E; Trombetta, G. Dissecting the free energy of formation of the 1:1 actomyosin complex. Biophys. Chem 2001, 89, 181–191. [Google Scholar]
- Maughan, D; Gorman, T. In Joint Meeting of the European Club for Muscle Motility and the European Cytoskeletal Club on Cellular Dynamics. Tyberias: Israel, 1987; p. SP-14. [Google Scholar]
- Brokaw, CJ. Computer simulation of movement-generating crossbridges. Biophys. J 1976, 16, 1013–1027. [Google Scholar]
- Rayment, Y; Rypnievski, WR; Schmidt-Base, K; Smith, R; Tomchick, DB; Benning, MM; Winkelmann, DA; Wesenberg, G; Holden, HM. Three-dimensional structure of myosin subfragment-1: A molecular motor. Science 1993, 261, 50–58. [Google Scholar]
- Lebovitz, JL; Helfand, E; Praestgaard, E. Scaled particle theory of fluid mixtures. J. Chem. Phys 1965, 43, 774–779. [Google Scholar]
- Gibbson, RM. The scaled particle theory for mixtures of hard convex particles. Mol. Phys 1970, 18, 809–816. [Google Scholar]
- Minton, AP. Excluded volume as a determinant of macromolecular structure and reactivity. Biopolymers 1981, 20, 2093–2120. [Google Scholar]
- Glasstone, S. Thermodynamics for Chemists; Van Norstrand: Princeton, NJ, 1947; p. 365. [Google Scholar]
- Morel, JE; Garrigos, M. Dimerization of the myosin heads in solution. Biochemistry 1982, 21, 2679–2686. [Google Scholar]
- Grussaute, H; Ollagnon, F; Morel, JE. F-actin-myosin-subfragment-1 (S1) interactions. Identification of the refractory state of S1 with the S1 dimer. Eur. J. Biochem 1995, 228, 524–529. [Google Scholar]
- Grazi, E; Magri, E; Schwienbacher, C; Trombetta, G. Osmotic properties of myosin subfragment-1: Implications of the mechanism of muscle contraction. Archiv. Biochem. Biophys 1995, 322, 97–102. [Google Scholar]
- Parsegian, VA; Rand, RP; Fuller, NL; Rau, DC. Osmotic stress for the direct measurement of intermolecular forces. Methods Enzymol 1986, 127, 400–416. [Google Scholar]
- Oplatka, A. On the mechanochemistry of muscle contraction. J. Theor. Biol 1972, 54, 437–447. [Google Scholar]
- Merah, Z; Morel, JE. Isometric tension exerted by a myofibril of the frog at 0 °C; geometrical considerations. J. Muscle Res. Cell Motil 1993, 14, 552–553. [Google Scholar]
- Grazi, E; Magri, E; Schwienbacher, C; Trombetta, G. The stiffness of the crossbridge is a function of the intrinsic protein osmotic pressure generated by the crossbridge itself. FEBS Lett 1996a, 387, 101–104. [Google Scholar]
- Grazi, E; Magri, E; Schwienbacher, C; Trombetta, G. A model relating protein osmotic pressure to the stiffness of the cross-bridge components and the contractile force of skeletal muscle. Eur. J. Biochem 1996b, 241, 25–31. [Google Scholar]
- Huxley, AF. Muscle structure and theories of contraction. Prog. Biophys. Biophys. Chem 1957, 7, 255–318. [Google Scholar]
- Worthington, CR. Conceptual model for the force–velocity relation of muscle (Hill’s equation). Nature 1962, 193, 1283–1284. [Google Scholar]
- Worthington, CR; Elliott, GF. Muscle contraction: the step size distance and the impulse time per ATP. Int. J. Biol. Macromol 1996, 18, 123–131. [Google Scholar]
- Reconditi, M; Linari, M; Lucii, L; Stewart, A; Sun, Y; Boesecke, P; Narayanan, T; Fischetti, RF; Irving, T; Piazzesi, G; Irving, M; Lombardi, V. The myosin motor in muscle generates a smaller and slower working stroke at higher load. Nature 2004, 428, 578–581. [Google Scholar]
- He, ZH; Rodney, RK; Chillinworth, RK; Brune, M; Corrie, JET; Webb, MR; Ferenczy, MA. The efficiency of contraction in rabbit skeletal muscle fibres, determined from the rate of release of inorganic phosphate. J. Physiol 1999, 517(3), 839–854. [Google Scholar]
- Duke, TAJ. Biophysics molecular model of muscle contraction. Proc. Natl. Acad. Sci. USA 1999, 96, 2770–2775. [Google Scholar]
- Huxley, HE. Past, present and future experiments on muscle. Phil.Trans. R. Soc. Lond. B 2000, 355, 539–543. [Google Scholar]
- Kushmerick, MJ. Appendix. Free energy of ATP hydrolysis in the sacoplasm. Proc. Roy. Soc. Lond. B 1969, 174, 348–353. [Google Scholar]
- Hill, AV. The maximum work and mechanical efficiency of human muscles and their most economical speed. J. Physiol. (London) 1922, 56, 19–41. [Google Scholar]
- Gasser, HS; Hill, AV. The dynamics of muscular contraction. Proc. R. Soc. (London) 1924, 96, 398–437. [Google Scholar]
- Fenn, WO. A quantitative comparison between the energy liberated and the work performed by the isolate sartorius muscle of the frog. J. Physiol. (London) 1923, 58, 175–203. [Google Scholar]
- Ernst, E. Sliding friction contra sliding hypothesis. Acta Biochim. Biophys. Acad. Sci. Hung 1977, 12, 83–85. [Google Scholar]
- Huxley, AF. Reflections on Muscle.; Liverpool University Press: Liverpool, UK, 1980. [Google Scholar]
- Cecchi, G; Bagni, MA; Cecchini, E; Colombini, B; Colomo, F. Crossbridge viscosity in activated frog muscle fibres. Biophys. Chem 1997, 68, 1–8. [Google Scholar]
- Bagni, MA; Cecchi, G; Cecchini, E; Colombini, B; Colomo, F. Force responses to fast ramp stretches in stimulated frog skeletal muscle fibres. J. Muscle Res. Cell Motil 1998, 19, 33–42. [Google Scholar]
- Elliott, GF; Worthington, CR. Muscle contraction: Viscous-like frictional forces and the impulsive model. Int. J. Biol. Macromol 2001, 29, 213–218. [Google Scholar]
- Muller, W. Zum Problem der Anlaufstromung einer Flussigkeit im geraden Rohr mot Kreisring und Kreisquerschnitt. Z. Angevw. Math. Mech 1932, 16, 227–238. [Google Scholar]
- Szymanski, PJ. Math. Pures Appl. Ser 1932, 9(II), 67–107.
- Schwienbacher, C; Magri, E; Trombetta, G; Grazi, E. The osmotic properties of the calcium regulated actin filament. Biochemistry 1995, 34, 1090–1095. [Google Scholar]
- Hanson, J; Lowy, J. The structure of F-actin and of actin filaments isolated from muscle. J. Mol. Biol 1963, 6, 46–60. [Google Scholar]
- Grazi, E; Cuneo, P; Magri, E; Adami, R; Trombetta, G. Protein cross talking through osmotic work: the free energy of formation of the MgADP-myosin complexes at the muscle protein osmotic pressure. Biochim Biophys Acta 1998, 1388, 419–427. [Google Scholar]
- Rasgado-Flores, H; Taylor, SR; Pena-Rasgado, C; Orgel, J; Gonzalez-Serratos, H. Muscle contraction and cell volume changes in skeletal muscle. J. Muscle Res. Cell Motil 2004, 25, 592–595. [Google Scholar]
- MacKay, RS; MacKay, DJC. Ergodic pumping: A mechanism to drive biomolecular conformation changes. Physica D 2006, 216, 220–234. [Google Scholar]





© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Grazi, E. Water and Muscle Contraction. Int. J. Mol. Sci. 2008, 9, 1435-1452. https://doi.org/10.3390/ijms9081435
Grazi E. Water and Muscle Contraction. International Journal of Molecular Sciences. 2008; 9(8):1435-1452. https://doi.org/10.3390/ijms9081435
Chicago/Turabian StyleGrazi, Enrico. 2008. "Water and Muscle Contraction" International Journal of Molecular Sciences 9, no. 8: 1435-1452. https://doi.org/10.3390/ijms9081435