
Multiplex Detection of Fluorescent Chemokine Binding to CXC Chemokine Receptors by NanoBRET


Abstract
:1. Introduction
2. Results
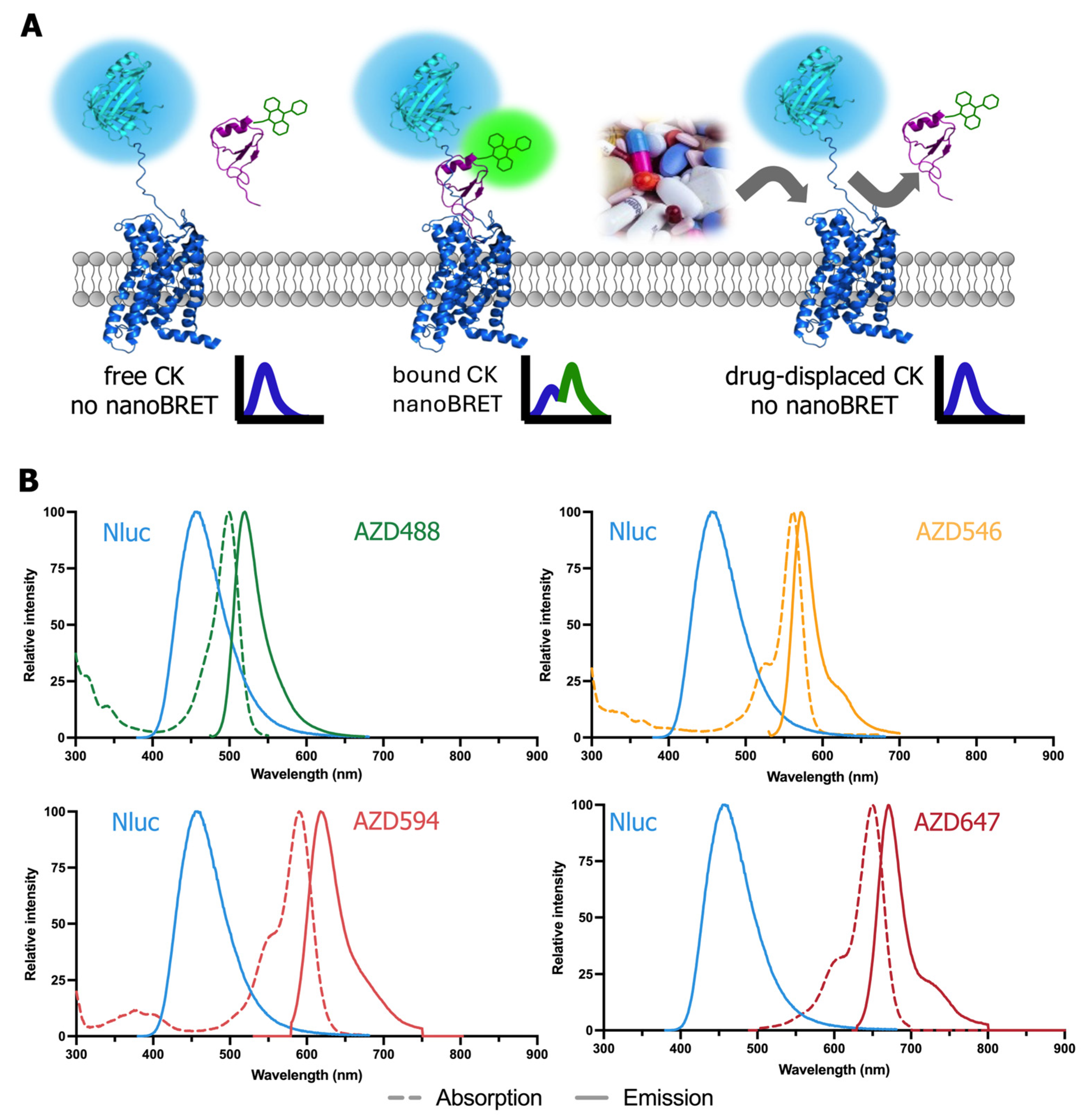
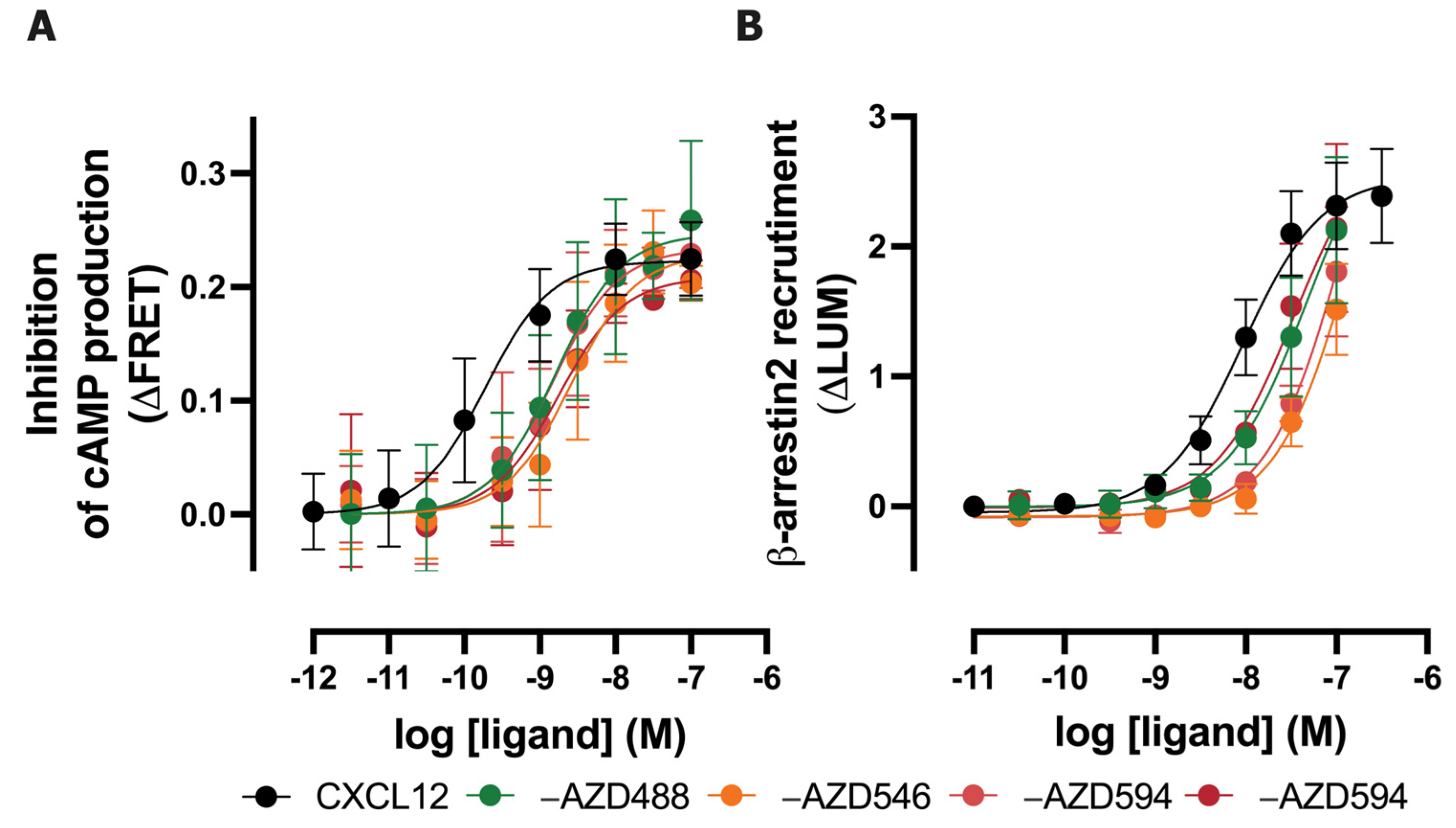
2.1. Fluorescent AZDye-Labeled CXCL12 Analogs Act as CXCR4 and ACKR3 Agonists
2.2. Fluorescent AZDye-Labeled CXCL12 Analogs Display Comparable Binding Affinities for ACKR3 or CXCR4
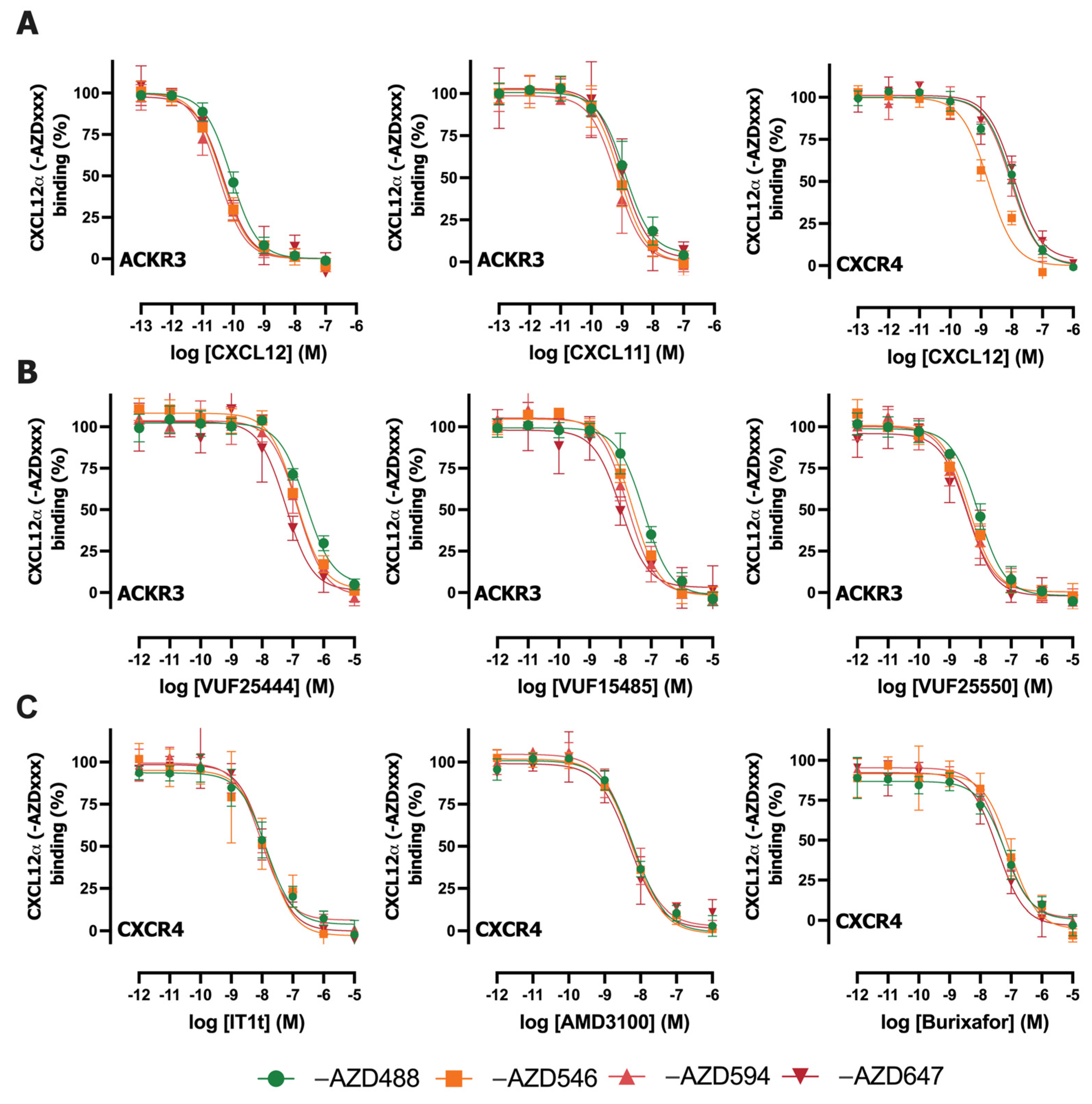
2.3. Inhibition of CXCL12-AZDxxx Binding to NLuc-ACKR3 or NLuc-CXCR4 by Various Ligands
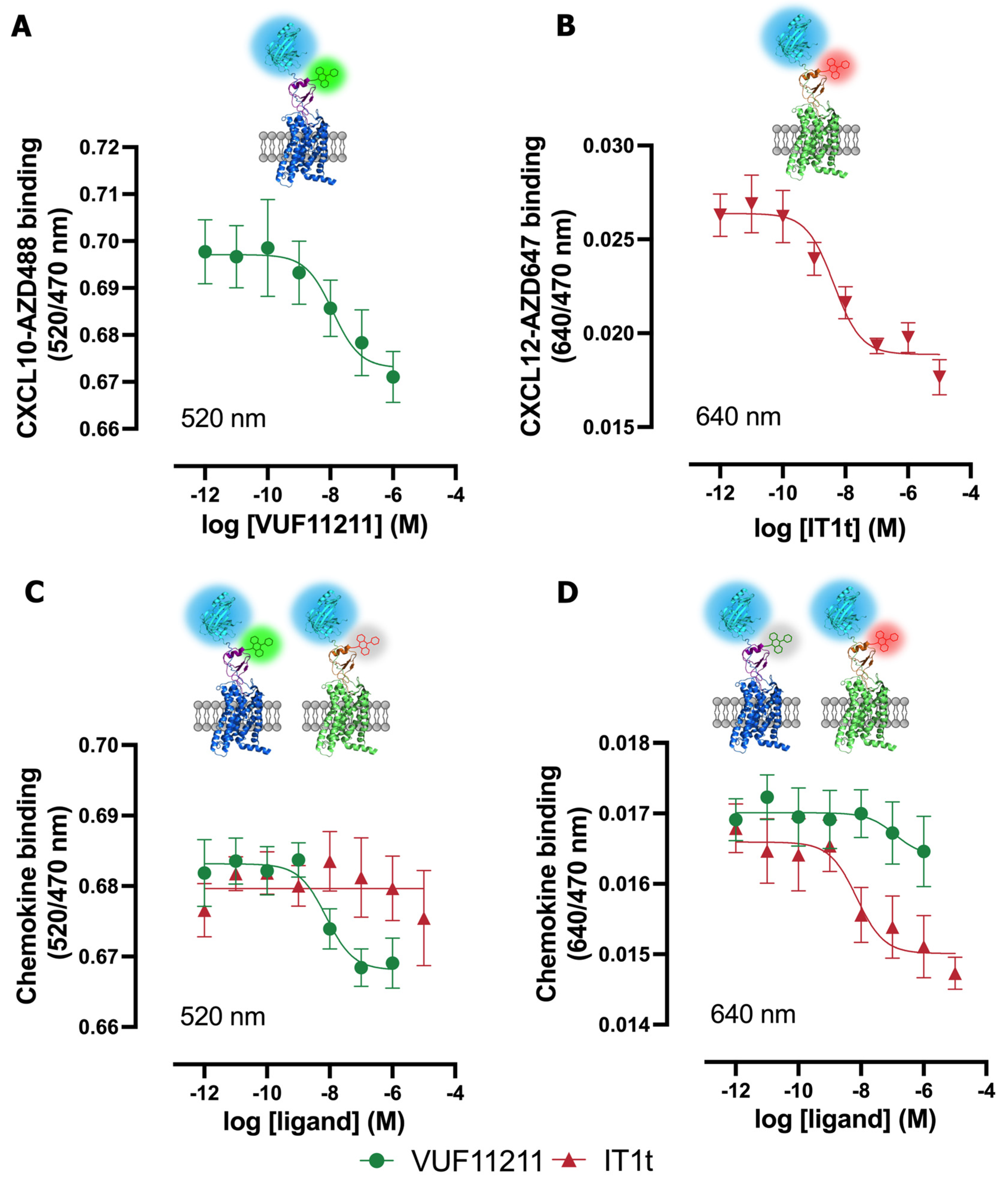
2.4. Multiplexed NanoBRET-Based Binding Assay to Simultaneously Detect Ligand Binding to Two Different Chemokine Receptors in the Same Sample
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Constructs
4.3. Cell Culture
4.4. β-arrestin2 Recruitment (NanoBit)
4.5. Intracellular cAMP Measurement by EPAC Sensor
4.6. NLuc-ACKR3, NLuc-CXCR4, NLuc-CXCR3 HEK293T Membrane Preparation
4.7. NanoBRET-Based Binding Assay
4.8. Multiplex NanoBRET-Based Binding Assay to CXCR4 and CXCR3
4.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hatse, S.; Princen, K.; Liekens, S.; Vermeire, K.; De Clercq, E.; Schols, D. Fluorescent CXCL12AF647 as a novel probe for nonradioactive CXCL12/CXCR4 cellular interaction studies. Cytom. Part A 2004, 61, 178–188. [Google Scholar] [CrossRef]
- Zwier, J.M.; Roux, T.; Cottet, M.; Durroux, T.; Douzon, S.; Bdioui, S.; Gregor, N.; Bourrier, E.; Oueslati, N.; Nicolas, L.; et al. A fluorescent ligand-binding alternative using tag-lite technology. J. Biomol. Screen. 2010, 15, 1248–1259. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, L.A.; Johnstone, E.K.M.; Wheal, A.J.; Goulding, J.; Robers, M.B.; MacHleidt, T.; Wood, K.V.; Hill, S.J.; Pfleger, K.D.G. Application of BRET to monitor ligand binding to GPCRs. Nat. Methods 2015, 12, 661–663. [Google Scholar] [CrossRef]
- Stoddart, L.A.; Kilpatrick, L.E.; Hill, S.J. NanoBRET Approaches to Study Ligand Binding to GPCRs and RTKs. Trends Pharmacol. Sci. 2018, 39, 136–147. [Google Scholar] [CrossRef] [PubMed]
- White, C.W.; Caspar, B.; Vanyai, H.K.; Pfleger, K.D.G.; Hill, S.J. CRISPR-Mediated Protein Tagging with Nanoluciferase to Investigate Native Chemokine Receptor Function and Conformational Changes. Cell Chem. Biol. 2020, 27, 499–510.e7. [Google Scholar] [CrossRef] [PubMed]
- Cole, N.B. Site-specific protein labeling with SNAP-tags. Curr. Protoc. Protein Sci. 2013, 2013, 30.1.1–30.1.16. [Google Scholar] [CrossRef]
- van den Bor, J.; Bergkamp, N.D.; Anbuhl, S.M.; Dekker, F.; Comez, D.; Perez Almeria, C.V.; Bosma, R.; White, C.W.; Kilpatrick, L.E.; Hill, S.J.; et al. NanoB2 to monitor interactions of ligands with membrane proteins by combining nanobodies and NanoBRET. Cell Rep. Methods 2023, 3, 100422. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, S.; Comez, D.; Karsai, N.; Arimont-Segura, M.; Canals, M.; Caspar, B.; de Graaf, C.; Kilpatrick, L.E.; Leurs, R.; Kellam, B.; et al. Small Molecule Fluorescent Ligands for the Atypical Chemokine Receptor 3 (ACKR3). ACS Med. Chem. Lett. 2024, 15, 143–148. [Google Scholar] [CrossRef]
- Boon, K.; Vanalken, N.; Szpakowska, M.; Chevigné, A.; Schols, D.; Van Loy, T. Systematic assessment of chemokine ligand bias at the human chemokine receptor CXCR2 indicates G protein bias over β-arrestin recruitment and receptor internalization. Cell Commun. Signal. 2024, 22, 43. [Google Scholar] [CrossRef]
- Chevigné, A.; Janji, B.; Meyrath, M.; Reynders, N.; D’uonnolo, G.; Uchański, T.; Xiao, M.; Berchem, G.; Ollert, M.; Kwon, Y.J.; et al. Cxcl10 is an agonist of the cc family chemokine scavenger receptor ackr2/d6. Cancers 2021, 13, 1054. [Google Scholar] [CrossRef]
- Scholten, D.J.; Canals, M.; Maussang, D.; Roumen, L.; Smit, M.J.; Wijtmans, M.; De Graaf, C.; Vischer, H.F.; Leurs, R. Pharmacological modulation of chemokine receptor function. Br. J. Pharmacol. 2012, 165, 1617–1643. [Google Scholar] [CrossRef] [PubMed]
- Szpakowska, M.; D’Uonnolo, G.; Luís, R.; Alonso Bartolomé, A.; Thelen, M.; Legler, D.F.; Chevigné, A. New pairings and deorphanization among the atypical chemokine receptor family—Physiological and clinical relevance. Front. Immunol. 2023, 14, 1133394. [Google Scholar] [CrossRef] [PubMed]
- Poeta, V.M.; Massara, M.; Capucetti, A.; Bonecchi, R. Chemokines and chemokine receptors: New targets for cancer immunotherapy. Front. Immunol. 2019, 10, 379. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.M.; Summers, B.C.; Wang, Y.; Melikian, A.; Berahovich, R.; Miao, Z.; Penfold, M.E.T.; Sunshine, M.J.; Littman, D.R.; Kuo, C.J.; et al. A novel chemokine receptor for SDF-1 and I-TAC involved in cell survival, cell adhesion, and tumor development. J. Exp. Med. 2006, 203, 2201–2213. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Riese, D.J.; Shen, J. The Role of the CXCL12/CXCR4/CXCR7 Chemokine Axis in Cancer. Front. Pharmacol. 2020, 11, 574667. [Google Scholar] [CrossRef] [PubMed]
- Do, H.T.T.; Lee, C.H.; Cho, J. Chemokines and their receptors: Multifaceted roles in cancer progression and potential value as cancer prognostic markers. Cancers 2020, 12, 287. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.; Murphy, P.M. Adaptive immunodeficiency in WHIM syndrome. Int. J. Mol. Sci. 2019, 20, 3. [Google Scholar] [CrossRef] [PubMed]
- Richard-Bildstein, S.; Aissaoui, H.; Pothier, J.; Schäfer, G.; Gnerre, C.; Lindenberg, E.; Lehembre, F.; Pouzol, L.; Guerry, P. Discovery of the Potent, Selective, Orally Available CXCR7 Antagonist ACT-1004-1239. J. Med. Chem. 2020, 63, 15864–15882. [Google Scholar] [CrossRef]
- Shelke, N.B.; Kadam, R.; Tyagi, P.; Rao, V.R.; Kompella, U.B. Intravitreal poly(l-lactide) microparticles sustain retinal and choroidal delivery of TG-0054, a hydrophilic drug intended for neovascular diseases. Drug Deliv. Transl. Res. 2011, 1, 76–90. [Google Scholar] [CrossRef]
- Skerlj, R.T.; Bridger, G.J.; Kaller, A.; McEachern, E.J.; Crawford, J.B.; Zhou, Y.; Atsma, B.; Langille, J.; Nan, S.; Veale, D.; et al. Discovery of novel small molecule orally bioavailable C-X-C chemokine receptor 4 antagonists that are potent inhibitors of T-tropic (X4) HIV-1 replication. J. Med. Chem. 2010, 53, 3376–3388. [Google Scholar] [CrossRef] [PubMed]
- Zarca, A.; Adlere, I.; Viciano, C.P.; Arimont-Segura, M.; Meyrath, M.; Simon, I.A.; Bebelman, J.P.; Laan, D.; Custers, H.G.J.; Janssen, E.; et al. Pharmacological characterization and radiolabeling of VUF15485, a high-affinity small-molecule agonist for the atypical chemokine receptor ACKR3. Mol. Pharmacol. 2024, 105, 301–312. [Google Scholar] [CrossRef]
- Scholten, D.J.; Wijtmans, M.; Van Senten, J.R.; Custers, H.; Stunnenberg, A.; De Esch, I.J.P.; Smit, M.J.; Leurs, R. Pharmacological characterization of [3H]VUF11211, a novel radiolabeled small-molecule inverse agonist for the chemokine receptor CXCR3. Mol. Pharmacol. 2015, 87, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Adlere, I.; Sun, S.; Zarca, A.; Roumen, L.; Gozelle, M.; Viciano Perpiñá, C.; Caspar, B.; Arimont, M.; Bebelman, J.P.; Briddon, S.J.; et al. Structure-based exploration and pharmacological evaluation of N-substituted piperidin-4-yl-methanamine CXCR4 chemokine receptor antagonists. Eur. J. Med. Chem. 2019, 162, 631–649. [Google Scholar] [CrossRef] [PubMed]
- Wedemeyer, M.J.; Mahn, S.A.; Getschman, A.E.; Crawford, K.S.; Peterson, F.C.; Marchese, A.; McCorvy, J.D.; Volkman, B.F. The chemokine X-factor: Structure-function analysis of the CXC motif at CXCR4 and ACKR3. J. Biol. Chem. 2020, 295, 13927–13939. [Google Scholar] [CrossRef] [PubMed]
- Fretz, H.; Gude, M.; Guerry, P.; Kimmerlin, T.; Lehembre, F.; Pfeifer, T.; Valdenaire, A. 1-[m-carboxamido(hetero)aryl-methyl]-heterocyclyl-carboxamide Derivatives. U.S. Patent 9,428,456, 15 September 2020. [Google Scholar]
- Thoma, G.; Streiff, M.B.; Kovarik, J.; Glickman, F.; Wagner, T.; Beerli, C.; Zerwes, H.G. Orally bioavailable isothioureas block function of the chemokine receptor CXCR4 in vitro and in vivo. J. Med. Chem. 2008, 51, 7915–7920. [Google Scholar] [CrossRef] [PubMed]
- Schols, D.; Este, A.; Henson, G.; De Clercq, E. Bicyclams, a class of potent anti-HIV agents, are targeted at the HIV coreceptor fusin/CXCR-4. Antivir. Res. 1997, 35, 147–156. [Google Scholar] [CrossRef]
- Bérenger, S.; Adamska, J.M.; Deflorian, F.; De Graaf, C.; Christie, B.; Szpakowska, M.; Chevigné, A.; De Esch, I.J.P.; Zarzycka, B.; Vischer, H.F.; et al. Design, synthesis and pharmacological characterization of the first photoswitchable small-molecule agonist for the Atypical Chemokine Receptor 3. bioRxiv 2024, 1–25. [Google Scholar] [CrossRef]
- Dekkers, S.; Caspar, B.; Goulding, J.; Kindon, N.D.; Kilpatrick, L.E.; Stoddart, L.A.; Briddon, S.J.; Kellam, B.; Hill, S.J.; Stocks, M.J. Small-Molecule Fluorescent Ligands for the CXCR4 Chemokine Receptor. J. Med. Chem. 2023, 66, 5208–5222. [Google Scholar] [CrossRef]
- White, C.W.; Platt, S.; Kilpatrick, L.E.; Dale, N.; Abhayawardana, R.S.; Dekkers, S.; Kindon, N.D.; Kellam, B.; Stocks, M.J.; Pfleger, K.D.G.; et al. CXCL17 is an allosteric inhibitor of CXCR4 through a mechanism of action involving glycosaminoglycans. Sci. Signal. 2024, 17, eabl3758. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Liu, Y.-C.; Yen, C.-F.; Chen, H.-C.; Chen, S.-J.; King, C.-H.R.; Hsu, M.-C. Rapid Mobilization of Murine Hematopoietic Stem and Progenitor Cells with TG-0054, a Novel CXCR4 Antagonist. Blood 2009, 114, 3542. [Google Scholar] [CrossRef]
- Harms, M.; Gilg, A.; Ständker, L.; Beer, A.J.; Mayer, B.; Rasche, V.; Gruber, C.W.; Münch, J. Microtiter plate-based antibody-competition assay to determine binding affinities and plasma/blood stability of CXCR4 ligands. Sci. Rep. 2020, 10, 16036. [Google Scholar] [CrossRef] [PubMed]
- Scholten, D.J.; Canals, M.; Wijtmans, M.; De Munnik, S.; Nguyen, P.; Verzijl, D.; De Esch, I.J.P.; Vischer, H.F.; Smit, M.J.; Leurs, R. Pharmacological characterization of a small-molecule agonist for the chemokine receptor CXCR3. Br. J. Pharmacol. 2012, 166, 898–911. [Google Scholar] [CrossRef] [PubMed]
- Yen, Y.C.; Schafer, C.T.; Gustavsson, M.; Eberle, S.A.; Dominik, P.K.; Deneka, D.; Zhang, P.; Schall, T.J.; Kossiakoff, A.A.; Tesmer, J.J.G.; et al. Structures of atypical chemokine receptor 3 reveal the basis for its promiscuity and signaling bias. Sci. Adv. 2022, 8, eabn8063. [Google Scholar] [CrossRef] [PubMed]
- Ngo, T.; Stephens, B.S.; Gustavsson, M.; Holden, L.G.; Abagyan, R.; Handel, T.M.; Kufareva, I. Crosslinking-guided geometry of a complete CXC receptor-chemokine complex and the basis of chemokine subfamily selectivity. PLoS Biol. 2020, 18, e3000656. [Google Scholar] [CrossRef] [PubMed]
- Jaracz-Ros, A.; Bernadat, G.; Cutolo, P.; Gallego, C.; Gustavsson, M.; Cecon, E.; Baleux, F.; Kufareva, I.; Handel, T.M.; Bachelerie, F.; et al. Differential activity and selectivity of N-terminal modified CXCL12 chemokines at the CXCR4 and ACKR3 receptors. J. Leukoc. Biol. 2020, 107, 1123–1135. [Google Scholar] [CrossRef] [PubMed]
- Szpakowska, M.; Nevins, A.M.; Meyrath, M.; Rhainds, D.; D’huys, T.; Guité-Vinet, F.; Dupuis, N.; Gauthier, P.A.; Counson, M.; Kleist, A.; et al. Different contributions of chemokine N-terminal features attest to a different ligand binding mode and a bias towards activation of ACKR3/CXCR7 compared with CXCR4 and CXCR3. Br. J. Pharmacol. 2018, 175, 1419–1438. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Stephens, B.; Qin, L.; Yin, X.; Dores, M.R.; Smith, T.H.; Grimsey, N.; Abagyan, R.; Trejo, J.A.; Kufareva, I.; et al. A general method for site specific fluorescent labeling of recombinant chemokines. PLoS ONE 2014, 9, e81454. [Google Scholar] [CrossRef]
- Picciocchi, A.; Šiaučiunaitee-Gaubard, L.; Petit-Hartlein, I.; Sadir, R.; Revilloud, J.; Caro, L.; Vivaudou, M.; Fieschi, F.; Moreau, C.; Vivès, C. C-terminal engineering of CXCL12 and CCL5 chemokines: Functional characterization by electrophysiological recordings. PLoS ONE 2014, 9, e87394. [Google Scholar] [CrossRef]
- Kotliar, I.B.; Ceraudo, E.; Kemelmakher-Liben, K.; Oren, D.A.; Lorenzen, E.; Dodig-Crnković, T.; Horioka-Duplix, M.; Huber, T.; Schwenk, J.M.; Sakmar, T.P. Itch receptor MRGPRX4 interacts with the receptor activity–modifying proteins. J. Biol. Chem. 2023, 299, 104664. [Google Scholar] [CrossRef]
- Dahl, L.; Kotliar, I.B.; Bendes, A.; Dodig-Crnković, T.; Fromm, S.; Elofsson, A.; Uhlén, M.; Sakmar, T.P.; Schwenk, J.M. Multiplexed selectivity screening of anti-GPCR antibodies. Sci. Adv. 2023, 9, eadf9297. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Wang, Y.; Yu, X.; He, Y.; Zheng, X.; Qin, J.; Gu, Y.; Li, X.; Shi, Y.; Ma, X.; et al. An image-based Abplex method for high-throughput GPCRs antibody discovery. Biotechnol. J. 2024, 19, e2300336. [Google Scholar] [CrossRef] [PubMed]
- French, A.R.; Meqbil, Y.J.; van Rijn, R.M. ClickArr: A novel, high-throughput assay for evaluating β-arrestin isoform recruitment. Front. Pharmacol. 2023, 14, 1295518. [Google Scholar] [CrossRef] [PubMed]
- Mella, R.M.; Kortazar, D.; Roura-Ferrer, M.; Salado, C.; Valcárcel, M.; Castilla, A.; Villacé, P. Nomad Biosensors: A New Multiplexed Technology for the Screening of GPCR Ligands. SLAS Technol. 2018, 23, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, A.; Lage, S.; Cadavid, M.I.; Loza, M.I.; Brea, J. Development of a Multiplex Assay for Studying Functional Selectivity of Human Serotonin 5-HT2A Receptors and Identification of Active Compounds by High-Throughput Screening. J. Biomol. Screen. 2016, 21, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Watson, J.; Chen, M.; Shen, D.R.; Yarde, M.; Agler, M.; Burford, N.; Alt, A.; Jayachandra, S.; Cvijic, M.E.; et al. Integrating high-content analysis into a multiplexed screening approach to identify and characterize GPCR agonists. J. Biomol. Screen. 2014, 19, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Watts, A.O.; Van Lipzig, M.M.H.; Jaeger, W.C.; Seeber, R.M.; Van Zwam, M.; Vinet, J.; Van Der Lee, M.M.C.; Siderius, M.; Zaman, G.J.R.; Boddeke, H.W.G.M.; et al. Identification and profiling of CXCR3-CXCR4 chemokine receptor heteromer complexes. Br. J. Pharmacol. 2013, 168, 1662–1674. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.; Martínez-Burgo, B.; Sepuru, K.M.; Rajarathnam, K.; Kirby, J.A.; Sheerin, N.S.; Ali, S. Regulation of chemokine function: The roles of GAG-binding and post-translational nitration. Int. J. Mol. Sci. 2017, 18, 1692. [Google Scholar] [CrossRef]
- Ma, X.; Leurs, R.; Vischer, H.F. NanoLuc-Based Methods to Measure β-Arrestin2 Recruitment to G Protein-Coupled Receptors. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2021; Volume 2268. [Google Scholar] [CrossRef]
- Reyes-Alcaraz, A.; Lee, Y.N.; Yun, S.; Hwang, J.I.; Seong, J.Y. Conformational signatures in β-arrestin2 reveal natural biased agonism at a G-protein-coupled receptor. Commun. Biol. 2018, 1, 128. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Excitation Maximum (nm) | Emission Maximum (nm) | cAMP by CXCR4 (pEC50) | β-arrestin2 Recruitment to ACKR3 (pEC50) | pKD for CXCR4 c | pKD for ACKR3 c |
|---|---|---|---|---|---|---|
| CXCL12 | - | - | 9.7 ± 0.2 a | 8.0 ± 0.1 a | - | - |
| -AZD488 | 494 | 517 | 8.8 ± 0.3 | 7.3 ± 0.2 | 8.2 ± 0.1 | 9.9 ± 0.1 |
| -AZD546 | 554 | 570 | 8.6 ± 0.3 | 6.8 ± 0.3 | 8.0 ± 0.0 | 9.5 ± 0.0 |
| -AZD594 | 590 | 617 | 8.9 ± 0.2 | 6.9 ± 0.2 | 7.6 ± 0.1 | 9.7 ± 0.1 |
| -AZD647 | 649 | 671 | 8.8 ± 0.2 | 7.5 ± 0.1 b | 7.9 ± 0.1 | 9.4 ± 0.2 |
| Receptor | Ligand | -AZD488 | -AZD546 | -AZD594 | -AZD647 |
|---|---|---|---|---|---|
| ACKR3 | CXCL12 | 10.6 ± 0.1 | 10.7 ± 0.1 | 10.8 ± 0.2 | 10.5 ± 0.1 |
| CXCL11 | 9.4 ± 0.3 | 9.4 ± 0.3 | 9.5 ± 0.3 | 9.2 ± 0.3 | |
| VUF25444 | 7.1 ± 0.1 | 7.2 ± 0.0 | 7.2 ± 0.2 | 7.5 ± 0.2 a | |
| VUF15485 | 7.9 ± 0.1 b | 7.9 ± 0.1 | 8.1 ± 0.2 | 8.2 ± 0.1 | |
| VUF25550 | 8.6 ± 0.1 | 8.7 ± 0.1 | 8.8 ± 0.3 | 8.7 ± 0.4 | |
| CXCR4 | CXCL12 | 8.4 ± 0.1 | 9.0 ± 0.1 c | 8.2 ± 0.2 | 8.4 ± 0.5 |
| IT1t | 8.2 ± 0.2 | 8.2 ± 0.4 | 8.2 ± 0.0 | 8.1 ± 0.2 | |
| AMD3100 | 8.6 ± 0.1 | 8.5 ± 0.0 | 8.5 ± 0.2 | 8.6 ± 0.2 | |
| Burixafor | 7.6 ± 0.2 | 7.3 ± 0.2 | 7.5 ± 0.1 | 7.7 ± 0.1 |
| Ligand | CXCR3 Only at 520 nm | CXCR4 Only at 640 nm | Multiplex at 520 nm | Multiplex at 640 nm |
|---|---|---|---|---|
| VUF11211 | 8.8 ± 0.2 | ND | 8.8 ± 0.1 | ND |
| IT1t | ND | 8.6 ± 0.3 | ND | 8.4 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamska, J.M.; Leftheriotis, S.; Bosma, R.; Vischer, H.F.; Leurs, R. Multiplex Detection of Fluorescent Chemokine Binding to CXC Chemokine Receptors by NanoBRET. Int. J. Mol. Sci. 2024, 25, 5018. https://doi.org/10.3390/ijms25095018
Adamska JM, Leftheriotis S, Bosma R, Vischer HF, Leurs R. Multiplex Detection of Fluorescent Chemokine Binding to CXC Chemokine Receptors by NanoBRET. International Journal of Molecular Sciences. 2024; 25(9):5018. https://doi.org/10.3390/ijms25095018
Chicago/Turabian StyleAdamska, Justyna M., Spyridon Leftheriotis, Reggie Bosma, Henry F. Vischer, and Rob Leurs. 2024. "Multiplex Detection of Fluorescent Chemokine Binding to CXC Chemokine Receptors by NanoBRET" International Journal of Molecular Sciences 25, no. 9: 5018. https://doi.org/10.3390/ijms25095018