
Transcriptomic Analysis Reveals the Inability of Recombinant AAV8 to Activate Human Monocyte-Derived Dendritic Cells
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results and Discussion
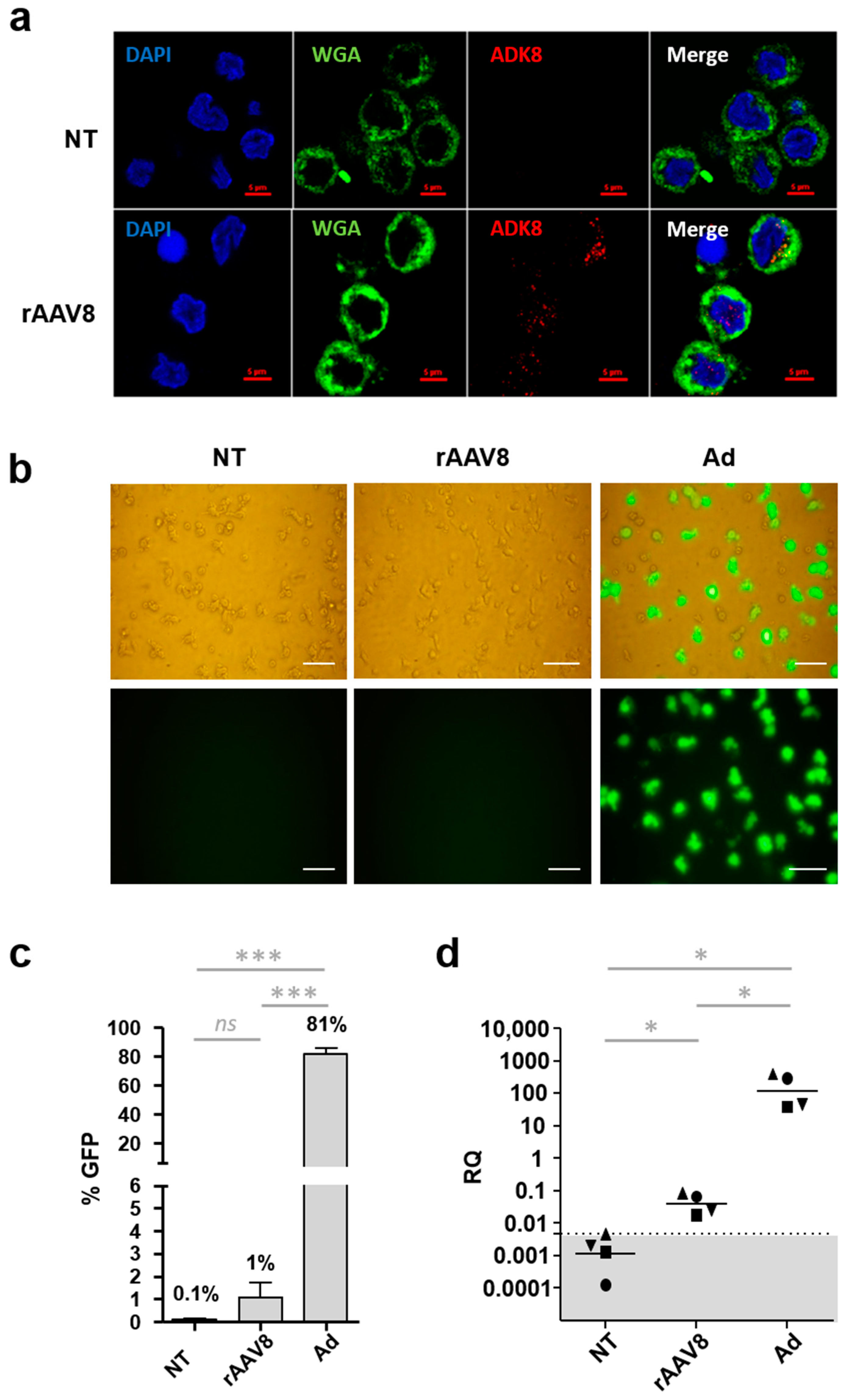
2.1. AAV8 Is Efficiently Internalized into Human moDCs
2.2. Recombinant AAV8 Fails to Activate Human moDC Inflammatory Pathways In Vitro
3. Materials and Methods
3.1. Generation of Monocyte-Derived Dendritic Cells (moDCs)
3.2. Vector Production
3.3. Staining of rAAV8 Particles by Immunohistochemistry
3.4. Image Analysis
3.5. Quantification of Transgene mRNA by RT-qPCR
3.6. Quantification of Vector Genome Extracted with or without Proteinase-Capsid Degradation
3.7. Cell Phenotyping by Flow Cytometry
3.8. 3′Sequencing RNA Profiling (3′SRP)
3.9. Bioinformatics Protocol
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Au, H.K.E.; Isalan, M.; Mielcarek, M. Gene Therapy Advances: A Meta-Analysis of AAV Usage in Clinical Settings. Front. Med. 2022, 8, 2746. [Google Scholar] [CrossRef] [PubMed]
- Salmon, F.; Grosios, K.; Petry, H. Safety Profile of Recombinant Adeno-Associated Viral Vectors: Focus on Alipogene Tiparvovec (Glybera®). Expert Rev. Clin. Pharmacol. 2014, 7, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Duan, D. Systemic Delivery of Adeno-Associated Viral Vectors. Curr. Opin. Virol. 2016, 21, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, V.; Petry, H.; Salmon, F. Immune Responses to AAV-Vectors, The Glybera Example from Bench to Bedside. Front. Immunol. 2014, 5, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendell, J.R.; Al-Zaidy, S.; Shell, R.; Arnold, W.D.; Rodino-Klapac, L.R.; Prior, T.W.; Lowes, L.; Alfano, L.; Berry, K.; Church, K.; et al. Single-Dose Gene-Replacement Therapy for Spinal Muscular Atrophy. N. Engl. J. Med. 2017, 377, 1713–1722. [Google Scholar] [CrossRef]
- Keeler, A.M.; Flotte, T.R. Recombinant Adeno-Associated Virus Gene Therapy in Light of Luxturna (and Zolgensma and Glybera): Where Are We, and How Did We Get Here? Annu. Rev. Virol. 2019, 6, 601–621. [Google Scholar] [CrossRef]
- Blair, H.A. Valoctocogene Roxaparvovec: First Approval. Drugs 2022, 82, 1505–1510. [Google Scholar] [CrossRef]
- FDA Approves HEMGENIX for Hemophilia B. Available online: https://www.cslbehring.com/newsroom/2022/fda-hemgenix (accessed on 16 December 2022).
- Keam, S.J. Eladocagene Exuparvovec: First Approval. Drugs 2022, 82, 1427–1432. [Google Scholar] [CrossRef]
- Manno, C.S.; Arruda, V.R.; Pierce, G.F.; Glader, B.; Ragni, M.; Rasko, J.; Ozelo, M.C.; Hoots, K.; Blatt, P.; Konkle, B.; et al. Successful Transduction of Liver in Hemophilia by AAV-Factor IX and Limitations Imposed by the Host Immune Response. Nat. Med. 2006, 12, 342–347. [Google Scholar] [CrossRef]
- Mingozzi, F.; Maus, M.V.; Hui, D.J.; Sabatino, D.E.; Murphy, S.L.; Rasko, J.E.J.; Ragni, M.V.; Manno, C.S.; Sommer, J.; Jiang, H.; et al. CD8+ T-Cell Responses to Adeno-Associated Virus Capsid in Humans. Nat. Med. 2007, 13, 419–422. [Google Scholar] [CrossRef]
- Nathwani, A.C.; Tuddenham, E.G.D.; Rangarajan, S.; Rosales, C.; McIntosh, J.; Linch, D.C.; Chowdary, P.; Riddell, A.; Pie, A.J.; Harrington, C.; et al. Adenovirus-Associated Virus Vector-Mediated Gene Transfer in Hemophilia B. N. Engl. J. Med. 2011, 365, 2357–2365. [Google Scholar] [CrossRef] [PubMed]
- Flotte, T.R.; Trapnell, B.C.; Humphries, M.; Carey, B.; Calcedo, R.; Rouhani, F.; Campbell-Thompson, M.; Yachnis, A.T.; Sandhaus, R.A.; McElvaney, N.G.; et al. Phase 2 Clinical Trial of a Recombinant Adeno-Associated Viral Vector Expressing α1-Antitrypsin: Interim Results. Hum. Gene Ther. 2011, 22, 1239–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, C.; Chulay, J.D.; Trapnell, B.C.; Humphries, M.; Carey, B.; Sandhaus, R.A.; McElvaney, N.G.; Messina, L.; Tang, Q.; Rouhani, F.N.; et al. Human Treg Responses Allow Sustained Recombinant Adeno-Associated Virus-Mediated Transgene Expression. J. Clin. Investig. 2013, 123, 5310–5318. [Google Scholar] [CrossRef] [PubMed]
- Mueller, C.; Gernoux, G.; Gruntman, A.M.; Borel, F.; Reeves, E.P.; Calcedo, R.; Rouhani, F.N.; Yachnis, A.; Humphries, M.; Campbell-Thompson, M.; et al. 5 Year Expression and Neutrophil Defect Repair after Gene Therapy in Alpha-1 Antitrypsin Deficiency. Mol. Ther. 2017, 25, 1387–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gernoux, G.; Gruntman, A.M.; Blackwood, M.; Zieger, M.; Flotte, T.R.; Mueller, C. Muscle-Directed Delivery of an AAV1 Vector Leads to Capsid-Specific T Cell Exhaustion in Nonhuman Primates and Humans. Mol. Ther. 2020, 28, 747–757. [Google Scholar] [CrossRef]
- Guilbaud, M.; Devaux, M.; Couzinié, C.; Le Duff, J.; Toromanoff, A.; Vandamme, C.; Jaulin, N.; Gernoux, G.; Larcher, T.; Moullier, P.; et al. Five Years of Successful Inducible Transgene Expression Following Locoregional Adeno-Associated Virus Delivery in Nonhuman Primates with No Detectable Immunity. Hum. Gene Ther. 2019, 30, 802. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, J.; Chan, Y.K.; Samulski, R.J. Adeno-Associated Virus (AAV) versus Immune Response. Viruses 2019, 11, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhuri, M.; Maeda, Y.; Ma, H.; Ram, S.; Fitzgerald, K.A.; Tai, P.W.L.; Gao, G. Overcoming Innate Immune Barriers That Impede AAV Gene Therapy Vectors. J. Clin. Investig. 2021, 131, e143780. [Google Scholar] [CrossRef]
- Zhu, J.; Huang, X.; Yang, Y. The TLR9-MyD88 Pathway Is Critical for Adaptive Immune Responses to Adeno-Associated Virus Gene Therapy Vectors in Mice. J. Clin. Investig. 2009, 119, 2388–2398. [Google Scholar] [CrossRef] [Green Version]
- Rogers, G.L.; Shirley, J.L.; Zolotukhin, I.; Kumar, S.R.P.; Sherman, A.; Perrin, G.Q.; Hoffman, B.E.; Srivastava, A.; Basner-Tschakarjan, E.; Wallet, M.A.; et al. Plasmacytoid and Conventional Dendritic Cells Cooperate in Crosspriming AAV Capsid-Specific CD8+ T Cells. Blood 2017, 129, 3184–3195. [Google Scholar] [CrossRef] [Green Version]
- Hösel, M.; Broxtermann, M.; Janicki, H.; Esser, K.; Arzberger, S.; Hartmann, P.; Gillen, S.; Kleeff, J.; Stabenow, D.; Odenthal, M.; et al. Toll-like Receptor 2-Mediated Innate Immune Response in Human Nonparenchymal Liver Cells toward Adeno-Associated Viral Vectors. Hepatology 2012, 55, 287–297. [Google Scholar] [CrossRef]
- Shao, W.; Earley, L.F.; Chai, Z.; Chen, X.; Sun, J.; He, T.; Deng, M.; Hirsch, M.L.; Ting, J.; Samulski, R.J.; et al. Double-Stranded RNA Innate Immune Response Activation from Long-Term Adeno-Associated Virus Vector Transduction. JCI Insight 2018, 3, e120474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathwani, A.C.; Reiss, U.M.; Tuddenham, E.G.D.; Rosales, C.; Chowdary, P.; McIntosh, J.; Della Peruta, M.; Lheriteau, E.; Patel, N.; Raj, D.; et al. Long-Term Safety and Efficacy of Factor IX Gene Therapy in Hemophilia B. N. Engl. J. Med. 2014, 371, 1994–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, A.; Okada, H.; Hayashita-Kinoh, H.; Shin, J.H.; Tamaoka, A.; Okada, T.; Takeda, S. RAAV8 and RAAV9-Mediated Long-Term Muscle Transduction with Tacrolimus (FK506) in Non-Human Primates. Mol. Ther. Methods Clin. Dev. 2020, 18, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Toromanoff, A.; Adjali, O.; Larcher, T.; Hill, M.; Guigand, L.; Chenuaud, P.; Deschamps, J.Y.; Gauthier, O.; Blancho, G.; Vanhove, B.; et al. Lack of Immunotoxicity After Regional Intravenous (RI) Delivery of RAAV to Nonhuman Primate Skeletal Muscle. Mol. Ther. 2010, 18, 151–160. [Google Scholar] [CrossRef]
- Vandenberghe, L.H.; Wang, L.; Somanathan, S.; Zhi, Y.; Figueredo, J.; Calcedo, R.; Sanmiguel, J.; Desai, R.A.; Chen, C.S.; Johnston, J.; et al. Heparin Binding Directs Activation of T Cells against Adeno-Associated Virus Serotype 2 Capsid. Nat. Med. 2006, 12, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, S.; Shin, J.H.; Yuasa, K.; Nishiyama, A.; Kira, J.; Okada, T.; Takeda, S. Transduction Efficiency and Immune Response Associated With the Administration of AAV8 Vector Into Dog Skeletal Muscle. Mol. Ther. 2009, 17, 73–80. [Google Scholar] [CrossRef]
- Mays, L.E.; Vandenberghe, L.H.; Xiao, R.; Bell, P.; Nam, H.-J.; Agbandje-McKenna, M.; Wilson, J.M. Adeno-Associated Virus Capsid Structure Drives CD4-Dependent CD8+ T Cell Response to Vector Encoded Proteins. J. Immunol. 2009, 182, 6051–6060. [Google Scholar] [CrossRef] [Green Version]
- Le Guiner, C.; Montus, M.; Servais, L.; Cherel, Y.; Francois, V.; Thibaud, J.L.; Wary, C.; Matot, B.; Larcher, T.; Guigand, L.; et al. Forelimb Treatment in a Large Cohort of Dystrophic Dogs Supports Delivery of a Recombinant AAV for Exon Skipping in Duchenne Patients. Mol. Ther. 2014, 22, 1923–1935. [Google Scholar] [CrossRef] [Green Version]
- Sonntag, F.; Köther, K.; Schmidt, K.; Weghofer, M.; Raupp, C.; Nieto, K.; Kuck, A.; Gerlach, B.; Böttcher, B.; Müller, O.J.; et al. The Assembly-Activating Protein Promotes Capsid Assembly of Different Adeno-Associated Virus Serotypes. J. Virol. 2011, 85, 12686–12697. [Google Scholar] [CrossRef] [Green Version]
- Gurda, B.L.; Raupp, C.; Popa-Wagner, R.; Naumer, M.; Olson, N.H.; Ng, R.; McKenna, R.; Baker, T.S.; Kleinschmidt, J.A.; Agbandje-McKenna, M. Mapping a Neutralizing Epitope onto the Capsid of Adeno-Associated Virus Serotype 8. J. Virol. 2012, 86, 7739–7751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.E.; Storm, T.A.; Huang, Z.; Kay, M.A. Rapid Uncoating of Vector Genomes Is the Key to Efficient Liver Transduction with Pseudotyped Adeno-Associated Virus Vectors. J. Virol. 2004, 78, 3110–3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jooss, K.; Yang, Y.; Fisher, K.J.; Wilson, J.M. Transduction of Dendritic Cells by DNA Viral Vectors Directs the Immune Response to Transgene Products in Muscle Fibers. J. Virol. 1998, 72, 4212–4223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veron, P.; Allo, V.; Rivière, C.; Bernard, J.; Douar, A.-M.; Masurier, C. Major Subsets of Human Dendritic Cells Are Efficiently Transduced by Self-Complementary Adeno-Associated Virus Vectors 1 and 2. J. Virol. 2007, 81, 5385–5394. [Google Scholar] [CrossRef] [Green Version]
- Ponnazhagan, S.; Mahendra, G.; Curiel, D.T.; Shaw, D.R. Adeno-Associated Virus Type 2-Mediated Transduction of Human Monocyte-Derived Dendritic Cells: Implications for Ex Vivo Immunotherapy. J. Virol. 2001, 75, 9493–9501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, K.-Q.; Mizukami, H.; Urabe, M.; Toda, Y.; Shinoda, K.; Yoshida, A.; Oomura, K.; Kojima, Y.; Ichino, M.; Klinman, D.; et al. Induction of Robust Immune Responses against Human Immunodeficiency Virus Is Supported by the Inherent Tropism of Adeno-Associated Virus Type 5 for Dendritic Cells. J. Virol. 2006, 80, 11899–11910. [Google Scholar] [CrossRef] [Green Version]
- Rossi, A.; Dupaty, L.; Aillot, L.; Zhang, L.; Gallien, C.; Hallek, M.; Odenthal, M.; Adriouch, S.; Salvetti, A.; Büning, H. Vector Uncoating Limits Adeno-Associated Viral Vector-Mediated Transduction of Human Dendritic Cells and Vector Immunogenicity. Sci. Rep. 2019, 9, 3631. [Google Scholar] [CrossRef] [Green Version]
- Gernoux, G.; Guilbaud, M.; Dubreil, L.; Larcher, T.; Babarit, C.; Ledevin, M.; Jaulin, N.; Planel, P.; Moullier, P.; Adjali, O. Early Interaction of Adeno-Associated Virus Serotype 8 Vector with the Host Immune System Following Intramuscular Delivery Results in Weak but Detectable Lymphocyte and Dendritic Cell Transduction. Hum. Gene Ther. 2015, 26, 1–13. [Google Scholar] [CrossRef]
- Wang, Z.; Storb, R.; Tapscott, S.J.; Riddell, S. Analyzing Cellular Immunity to AAV in a Canine Model Using ELISpot Assay. In Handbook of ELISPOT; Part of the Methods in Molecular Biology Book Series (MIMB, Volume 792); Humana Press: Totowa, NJ, USA, 2012; pp. 65–74. [Google Scholar] [CrossRef] [Green Version]
- Berry, G.E.; Asokan, A. Cellular Transduction Mechanisms of Adeno-Associated Viral Vectors. Curr. Opin. Virol. 2016, 21, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Mays, L.E.; Wang, L.; Lin, J.; Bell, P.; Crawford, A.; Wherry, E.J.; Wilson, J.M. AAV8 Induces Tolerance in Murine Muscle as a Result of Poor APC Transduction, T Cell Exhaustion, and Minimal MHCI Upregulation on Target Cells. Mol. Ther. 2014, 22, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Zabaleta, N.; Dai, W.; Bhatt, U.; Hérate, C.; Maisonnasse, P.; Chichester, J.A.; Sanmiguel, J.; Estelien, R.; Michalson, K.T.; Diop, C.; et al. An AAV-Based, Room-Temperature-Stable, Single-Dose COVID-19 Vaccine Provides Durable Immunogenicity and Protection in Non-Human Primates. Cell Host Microbe 2021, 29, 1437–1453.e8. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, E.; Cornec, M.; Dumont, S.; Meistermann, D.; Bordron, P.; David, L.; Redon, R.; Bonnaud, S.; Bihouée, A. 3′ RNA Sequencing for Robust and Low-Cost Gene Expression Profiling. Res. Sq. 2021, preprint. [Google Scholar] [CrossRef]
- Hoene, V.; Peiser, M.; Wanner, R. Human Monocyte-Derived Dendritic Cells Express TLR9 and React Directly to the CpG-A Oligonucleotide D19. J. Leukoc. Biol. 2006, 80, 1328–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreibelt, G.; Tel, J.; Sliepen, K.H.E.W.J.; Benitez-Ribas, D.; Figdor, C.G.; Adema, G.J.; De Vries, I.J.M. Toll-like Receptor Expression and Function in Human Dendritic Cell Subsets: Implications for Dendritic Cell-Based Anti-Cancer Immunotherapy. Cancer Immunol. Immunother. 2010, 59, 1573–1582. [Google Scholar] [CrossRef]
- Motwani, M.; Pesiridis, S.; Fitzgerald, K.A. DNA Sensing by the CGAS–STING Pathway in Health and Disease. Nat. Rev. Genet. 2019, 20, 657–674. [Google Scholar] [CrossRef]
- Schiroli, G.; Conti, A.; Ferrari, S.; della Volpe, L.; Jacob, A.; Albano, L.; Beretta, S.; Calabria, A.; Vavassori, V.; Gasparini, P.; et al. Precise Gene Editing Preserves Hematopoietic Stem Cell Function Following Transient P53-Mediated DNA Damage Response. Cell Stem Cell 2019, 24, 551–565.e8. [Google Scholar] [CrossRef] [Green Version]
- Zaiss, A.-K.; Liu, Q.; Bowen, G.P.; Wong, N.C.W.; Bartlett, J.S.; Muruve, D.A. Differential Activation of Innate Immune Responses by Adenovirus and Adeno-Associated Virus Vectors. J. Virol. 2002, 76, 4580–4590. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Allen, J.M.; Riddell, S.R.; Gregorevic, P.; Store, R.; Tapscott, S.J.; Chamberlain, J.S.; Kuhr, C.S. Immunity to Adeno-Associated Virus-Mediated Gene Transfer in a Random-Bred Canine Model of Duchenne Muscular Dystrophy. Hum. Gene Ther. 2007, 18, 18–26. [Google Scholar] [CrossRef]
- Abe, T.; Marutani, Y.; Shoji, I. Cytosolic DNA-Sensing Immune Response and Viral Infection. Microbiol. Immunol. 2019, 63, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Flotte, T.R. Revisiting the ‘New’ Inflammatory Toxicities of Adeno-Associated Virus Vectors. Hum. Gene Ther. 2020, 31, 398–399. [Google Scholar] [CrossRef]
- Nayak, S.; Herzog, R.W. Progress and Prospects: Immune Responses to Viral Vectors. Gene Ther. 2009, 17, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaiss, A.K.; Cotter, M.J.; White, L.R.; Clark, S.A.; Wong, N.C.W.; Holers, V.M.; Bartlett, J.S.; Muruve, D.A. Complement Is an Essential Component of the Immune Response to Adeno-Associated Virus Vectors. J. Virol. 2008, 82, 2727–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.J.; Ross, N.; Kamal, A.; Kim, K.Y.; Kropf, E.; Deschatelets, P.; Francois, C.; Quinn, W.J.I.; Singh, I.; Majowicz, A.; et al. Pre-Existing Humoral Immunity and Complement Pathway Contribute to Immunogenicity of Adeno-Associated Virus (AAV) Vector in Human Blood. Front. Immunol. 2022, 13, 999021. [Google Scholar] [CrossRef]
- Ayuso, E.; Blouin, V.; Lock, M.; Mcgorray, S.; Leon, X.; Alvira, M.R.; Auricchio, A.; Bucher, S.; Chtarto, A.; Clark, K.R.; et al. Manufacturing and Characterization of a Recombinant Adeno-Associated Virus Type 8 Reference Standard Material. Hum. Gene Ther. 2014, 25, 977–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Costa, S.; Blouin, V.; Broucque, F.; Penaud-Budloo, M.; Fçranois, A.; Perez, I.C.; Le Bec, C.; Moullier, P.; Snyder, R.O.; Ayuso, E. Practical Utilization of Recombinant AAV Vector Reference Standards: Focus on Vector Genomes Titration by Free ITR QPCR. Mol. Ther. Methods Clin. Dev. 2016, 5, 16019. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji—An Open Source Platform for Biological Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Köster, J.; Mölder, F.; Jablonski, K.P.; Letcher, B.; Hall, M.B.; Tomkins-Tinch, C.H.; Sochat, V.; Forster, J.; Lee, S.; Twardziok, S.O.; et al. Sustainable Data Analysis with Snakemake. F1000Research 2021, 10, 33. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor | Number of Analyzed Cells | Number of Cells with Internalized rAAV8 | % of Cell with Internalized rAAV8 |
|---|---|---|---|
| A | 1796 | 1774 | 98.7% |
| B | 221 | 183 | 82.8% |
| C | 345 | 183 | 53% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masri, S.; Carré, L.; Jaulin, N.; Vandamme, C.; Couzinié, C.; Guy-Duché, A.; Dupont, J.-B.; Pereira, A.; Charpentier, E.; David, L.; et al. Transcriptomic Analysis Reveals the Inability of Recombinant AAV8 to Activate Human Monocyte-Derived Dendritic Cells. Int. J. Mol. Sci. 2023, 24, 10447. https://doi.org/10.3390/ijms241310447
Masri S, Carré L, Jaulin N, Vandamme C, Couzinié C, Guy-Duché A, Dupont J-B, Pereira A, Charpentier E, David L, et al. Transcriptomic Analysis Reveals the Inability of Recombinant AAV8 to Activate Human Monocyte-Derived Dendritic Cells. International Journal of Molecular Sciences. 2023; 24(13):10447. https://doi.org/10.3390/ijms241310447
Chicago/Turabian StyleMasri, Samer, Laure Carré, Nicolas Jaulin, Céline Vandamme, Célia Couzinié, Aurélien Guy-Duché, Jean-Baptiste Dupont, Allwyn Pereira, Eric Charpentier, Laurent David, and et al. 2023. "Transcriptomic Analysis Reveals the Inability of Recombinant AAV8 to Activate Human Monocyte-Derived Dendritic Cells" International Journal of Molecular Sciences 24, no. 13: 10447. https://doi.org/10.3390/ijms241310447