
Metabolic Activation and DNA Interactions of Carcinogenic N-Nitrosamines to Which Humans Are Commonly Exposed


1
Masonic Cancer Center, University of Minnesota, Minneapolis, MN 55455, USA
2
Department of Medicinal Chemistry, College of Pharmacy, University of Minnesota, Minneapolis, MN 55455, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(9), 4559; https://doi.org/10.3390/ijms23094559
Submission received: 29 March 2022
/
Revised: 15 April 2022
/
Accepted: 18 April 2022
/
Published: 20 April 2022
(This article belongs to the Special Issue N-Nitroso Compounds: Biological and Environmental Significance and Defense Mechanisms)
Abstract
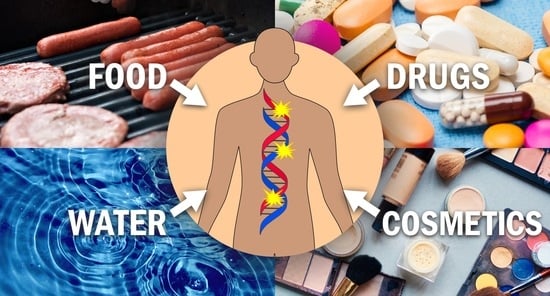
:Carcinogenic N-nitrosamine contamination in certain drugs has recently caused great concern and the attention of regulatory agencies. These carcinogens—widely detectable in relatively low levels in food, water, cosmetics, and drugs—are well-established and powerful animal carcinogens. The electrophiles resulting from the cytochrome P450-mediated metabolism of N-nitrosamines can readily react with DNA and form covalent addition products (DNA adducts) that play a central role in carcinogenesis if not repaired. In this review, we aim to provide a comprehensive and updated review of progress on the metabolic activation and DNA interactions of 10 carcinogenic N-nitrosamines to which humans are commonly exposed. Certain DNA adducts such as O6-methylguanine with established miscoding properties play central roles in the cancer induction process, whereas others have been linked to the high incidence of certain types of cancers. We hope the data summarized here will help researchers gain a better understanding of the bioactivation and DNA interactions of these 10 carcinogenic N-nitrosamines and facilitate further research on their toxicologic and carcinogenic properties.
1. Introduction
Peter Magee and John Barnes reported in 1956 that N-nitrosodimethylamine 1 (NDMA, Figure 1), a simple water-soluble compound with only 11 atoms, readily induced liver tumors in rats [1]. This was remarkable because most carcinogenesis studies at the time had been performed with higher molecular weight non-water-soluble compounds such as polycyclic aromatic hydrocarbons. Sakshaug et al., and Ender and Ceh made the connection between NDMA formation from nitrite-treated herring meal and liver toxicity in farm animals and provided evidence for the occurrence of this carcinogen in smoked fish and meat [2,3]. Thus, concern arose that N-nitrosamines in food treated with nitrite could be a carcinogenic hazard to humans. Sen and colleagues demonstrated the presence of N-nitrosamines in cured meat products [4] and this was followed by a surge in interest in the N-nitrosamine contamination of foods, which persists to the present.
In the meantime, cancer researchers investigated the powerful carcinogenic properties of multiple structurally diverse N-nitrosamines. Druckrey and co-workers and Lijinsky and colleagues demonstrated the carcinogenicity and frequent organoselectivity of multiple N-nitrosamines [5,6]. A review published in 1984 by Preussmann and Stewart summarizes the carcinogenicity of more than 200 N-nitrosamines, which commonly affect specific organs in laboratory animals, independent of the route of administration [7]. A book by Lijinsky also summarizes the extensive carcinogenicity data [5]. Bogovski and Bogovski published a summary of the carcinogenic activity of nitroso compounds in different animal species; NDMA induced tumors in 16 different animal species and NDEA in 26 ranging from rainbow trout to cynomolgus monkey [8]. Such a remarkable database hardly exists for any other type of carcinogen.
Thus, there was intense interest in the possible role of N-nitrosamines in human cancer in the latter part of the 20th century. The International Agency for Research on Cancer (IARC) held a series of regular meetings dedicated to this subject between 1966 and 1991. Figure 2 shows the participants at the 1983 meeting in Banff, Canada, a clear indication of the high interest in the topic. Fortunately, methods were developed to decrease levels of N-nitrosamines in food, beer, and other consumer products and interest in the topic waned somewhat in the early part of this century. Recently, however, concern regarding N-nitrosamine contamination of consumer products has re-emerged as they were found in certain pharmaceutical agents and drinking water [9]. It is worth noting that the carcinogenic potency of most N-nitrosamines is so great that they are excluded from the widely used Threshold of Toxicological Concern concept in the risk assessment of exposure to potential carcinogens in food and other consumer products [10].
All N-nitrosamines require metabolism to exert their carcinogenic properties. The electrophiles produced in these simple metabolic pathways, generally catalyzed by cytochrome P450 enzymes, readily alkylate DNA initiating the carcinogenic process. These critical pathways are the subject of this review of the 10 N-nitrosamines illustrated in Figure 1.
2. Overview of Carcinogenic N-Nitrosamines to Which Humans Are Commonly Exposed
N-Nitrosamines are the products of nitrosation reactions occurring on the N atoms of secondary and tertiary amines. They can be formed during water and food processing, tobacco curing, and drug and cosmetics manufacturing; they can also be formed endogenously. The compounds shown in Figure 1 represent an important family of carcinogens that are closely related to our daily lives [9,11].
2.1. Carcinogenic N-Nitrosamines Occurring in Food
The total N-nitrosamines occurring in food was estimated to be an average of 6.7 ± 0.8 ng/g, ranging from 0 to 120.8 ng/g [9]. NDEA was most frequently detected in 387 samples of agricultural food, whereas NDMA occurred at the highest concentration in seasoning, especially in processed fish (12.6–322.9 ng/g) and some oils (>10 ng/g) [12]. The average estimated concentrations of some common N-nitrosamines detected in food follows the descending order as NDMA (2.2 ± 0.3 ng/g), NDBA (1.5 ± 0.5 ng/g), NPYR (1.5 ± 0.2 ng/g), NDEA (0.9 ± 0.3 ng/g), NPIP (0.5 ± 0.1 ng/g), NMOR (0.05 ± 0.01 ng/g), NMEA (0.04 ± 0.01 ng/g), and NDPA (0.02 ± 0.01 ng/g) [9].
2.2. Carcinogenic N-Nitrosamines Occurring in Water
NDMA has been detected in potable water and gave rise to regulatory concerns regarding its presence in drinking water. The occurrence of NDMA is considered to be due to chloramination during water disinfection [13,14]. NDMA is the most prevalent N-nitrosamine contaminant in drinking water (average: 17.7 ± 4.7 ng/L), accounting for 5–13% of the total observed N-nitrosamines (average: 39.4 ± 10.5 ng/L; range: 2.8–309.0 ng/L) in water. The other carcinogenic N-nitrosamines detected in potable water include NPIP (7.9 ± 4.0 ng/L), NPYR (5.5 ± 2.6 ng/L), NDEA (4.2 ± 0.8 ng/L), NDBA (1.7 ± 0.6 ng/L), NMOR (0.9 ± 0.2 ng/L), NMEA (0.6 ± 0.1 ng/L), and NDPA (0.4 ± 0.03 ng/L) [9].
2.3. Carcinogenic N-Nitrosamines Occurring in Drugs
Drug contamination with N-nitrosamines has attracted great attention due to the recent recalls of more than 1400 lots of drugs from the U.S. market [15]. The presence of NDMA has been confirmed by the U.S. Food and Drug Administration (FDA) in some batches of ranitidine, nizatidine, metformin, and valsartan since 2018 [16]. Similarly, the detection of NDEA has caused the withdrawal of some types of irbesartan, losartan, and valsartan from the market [16]. The discovery of NDMA and NDEA has led regulatory agencies to conduct further analysis of the N-nitrosamine impurities in the affected drug products [17]. The U.S. FDA has identified 5 N-nitrosamines that were detected in drugs; they are NDMA, NDEA, N-nitroso-N-methyl-4-aminobutanoic acid (NMBA), N-nitroso-isopropylethylamine (NIPEA), and N-nitrodiisopropylamine (NDIPA). Two N-nitrosamines—NDBA and N-nitrosomethylphenylamine (NMPA)—are also considered to be theoretically present in drug products. The FDA-recommended acceptable intake limits for the carcinogenic N-nitrosamines discussed in this review are 96 ng/day (NDMA) and 26.5 ng/day (NDEA) [17].
2.4. Carcinogenic N-Nitrosamines Occurring in Cosmetics
The total N-nitrosamines occurring in cosmetics was estimated to be 1507 ± 752 ng/g, ranging from 0 to 49,000 ng/g. The relatively high concentrations of N-nitrosamines observed in cosmetics including hair care products, soaps, shampoos, lotions, and others are predominantly contributed by NDELA. It accounted for 99% of total N-nitrosamines in these products. The other minor N-nitrosamine contaminants are NMOR (~0.99%) and NDMA (~0.01%) [9]. NDEA has also been detected in cosmetics at levels ranging from 0–40.9 ng/g [18]. NDELA is formed by nitrosation of triethanolamine and diethanolamine [19] since they are readily present in cosmetics together with the nitrosating agents such as nitrite [18].
3. Metabolic Activation and DNA Interactions of Carcinogenic Acyclic N-Nitrosamines
As shown in Figure 1, there are seven carcinogenic acyclic N-nitrosamines (NDMA, NMEA, NSAR, NDEA, NDPA, NDELA, and NDBA) to which humans are commonly exposed. We discuss them in order based on their increasing structural complexity.
3.1. N-Nitrosodimethylamine (NDMA)
3.1.1. Exposure and Carcinogenicity
NDMA (1, Figure 1) is easily formed by nitrosation with sodium nitrite of an acidified solution of dimethylamine. It was used in industry for several purposes, the most common of which was as a precursor to the rocket fuel 1,1-dimethylhydrazine [20]. However, such uses of NDMA essentially ended after the study of Magee and Barnes [1]. Exposure to NDMA in humans occurs in low concentrations in daily life from food, water, and cosmetics to some contaminated drugs as noted in Section 2.
The IARC summarized the laboratory animal data of NDMA comprehensively in 1978 and reaffirmed its Group 2A classification in 1987. NDMA is a strong carcinogen primarily targeting the liver but it can also induce kidney tumors when given at high doses [7]. The cancer risk of NDMA exceeds many known strong carcinogens including asbestos, benzo[a]pyrene, and polychlorinated biphenyls [9]. A linear dose-response relationship was observed at low dose rates (lower than 1 ppm) in a study with 4080 rats. No indicated safe threshold concentration was observed for NDMA in the rats [21,22]. NDMA is tumorigenic in multiple animal models including Syrian golden hamsters, mastomys, guinea pigs, rabbits, and ducks [7]. The toxicity of NDMA in humans has been demonstrated by some unfortunate poisonings [20]. The currently available data suggest that the IARC classification of NDMA should be re-evaluated. The U.S. government classifies NDMA as “reasonably anticipated to be a human carcinogen” [23].
3.1.2. Metabolism
The bioactivation of NDMA to reactive intermediates is primarily catalyzed by P450 2E1 in human liver microsomes [24]. The oxidation of the methyl group (α-methyl hydroxylation) leads to α-hydroxyNDMA (11, Scheme 1), an unstable and mutagenic intermediate that spontaneously decomposes generating two reactive species—formaldehyde 12 and methyl diazohydroxide 14 [24,25]. Formaldehyde can be oxidized sequentially producing formic acid 13 and CO2 [26,27]; methyl diazohydroxide will spontaneously form the highly electrophilic methyldiazonium ion 15 and alkylate DNA [28] or be solvolyzed to methanol [29]. The evolution of 15N2 from 15N-labeled NDMA metabolism suggested that approximately 33–67% of NDMA was metabolized in vitro via α-methyl hydroxylation [30,31,32]. A deuterium isotope effect has been observed with a slight reduction of the NDMA metabolic oxidation rate for NDMA-d6 with the ratio of VH/VD = ~1.2. The effect was more prominent based on the concentrations of DNA adducts formed in the liver and kidneys after oral administration [33]. The methylating species or its immediate precursor 11, although highly reactive, were sufficiently stable to pass out of rat hepatocytes [34]. The enzymatic denitrosation of NDMA has also been observed in liver microsomes and is considered a detoxification pathway. The major denitrosation product of NDMA is methylamine 17, formed via the proposed intermediate methyliminium ion 16; the other product is formaldehyde [35,36,37].
3.1.3. Methyl DNA Adducts Formed by NDMA Metabolism
The mutagenicity and genotoxicity of NDMA are well-established [38]. It alkylates DNA and protein via the two reactive intermediates—methyldiazonium ion 15 and formaldehyde 12. Methyl DNA adducts formed by the methyldiazonium ion are considered to play a major role in carcinogenesis by NDMA [39]. Formaldehyde, on the other hand, can also form DNA adducts such as cross-links or hydroxymethylene adducts [40].
A comprehensive analysis of methyl DNA adducts in liver DNA was conducted in rats treated with NDMA by a single i.p. dose of 10 mg/kg [41]. Adducts were detected after deglycosylation. Methyl DNA adducts (Figure 3), including N3- and N7-Me-Ade (19 and 20), O2- and N3-Me-Cyt (21 and 22), N3-, O6-, N7-Me-Gua (23, 24 and 27) and Me-Fapy-Gua (29), O2-, N3-, and O4-Me-Thy (31, 33 and 35), and methyl DNA phosphate adduct 36, were formed 2 h after administration. The preferential methylation sites in liver DNA were Gua-N7, Gua-O6, and Ade-N3. The half-lives of methyl DNA adducts were relatively short, ranging from 4 to 17 h for the methyl base adducts and up to 7 days for the methyl phosphate adduct Tp(Me)T 37 [41]. The adduct profile in NDMA-exposed Syrian golden hamsters was different from that in the rats. O6-Me-Gua was the most persistent adduct, whereas N7-Me-Gua was extensively excised, especially in the liver. Other minor methyl adducts including N1-, N3-, N7-Me-Ade, and N3-Me-Gua were also detectable in most hamster tissues [42].
In the rat study, after a 2 mg/kg dose of NDMA by stomach tubing daily (workdays only) for up to 24 weeks, N7-Me-Gua 27 predominated in the liver, exceeding that in the kidneys and lungs [43] by approximately 16 times. This is consistent with the liver being the primary tumor site observed in carcinogenicity studies with low doses of NDMA. The other most commonly quantified adduct, O6-Me-Gua 24, accounted for 6.7% and 12.0% of N7-Me-Gua, 4 h and 24 h after a single s.c. dose of 0.055 mmol/kg NDMA, respectively [44]. Although the levels of N7-Me-Gua remained constant in the rat liver, increased excision of O6-Me-Gua was observed after chronic administration of NDMA [45,46]. This was due to the enhanced activity of O6-alkylguanine-DNA alkyltransferease (AGT) in the rat liver after repeated low-dose exposure to this carcinogen [47,48,49]. However, a clear interspecies difference has been noted, with a partially depleted AGT activity observed in mice chronically treated with low doses of NDMA [49]. The formation and persistence of O6-Me-Gua were also greatly affected by the co-treatment with ethanol, a known inhibitor of NDMA hepatic metabolism [50]. A remarkable 10-fold increase was observed in mammary gland DNA adducts in rats co-exposed to NDMA and ethanol. This is probably due to a reduced hepatic first-pass effect, which also resulted in slightly decreased concentrations of the liver DNA adducts [51].
3.1.4. Mutagenicity and Genotoxicity of Methyl DNA Adducts
DNA methylation, especially as related to the activation of proto-oncogenes, has been linked to the induction of pulmonary neoplasia by N-nitrosomethylamines [52]. There is convincing evidence that the formation and removal of O6-Me-Gua 24 (Figure 3) in DNA is related to the risk of tumor induction in different organs [53,54]. O6-Me-Gua, formed from alkylating agents such as NDMA, is possibly one of the most extensively studied of all DNA adducts. Its persistence in specific rat tissues where tumors developed after treatment with N-methyl-N-nitrosourea was established in early studies, supporting the hypothesis that it caused miscoding in DNA [55]. Elegant experiments by the Essigmann group demonstrated unequivocally that O6-Me-Gua causes G to A transition mutations, which is consistent with G-A transition mutations in the ras oncogene in laboratory animals treated with NDMA and other methylating carcinogens [56,57]. The repair enzyme AGT can remove the methyl group or other alkyl groups, returning the DNA to its unmodified structure [58].
Methylated thymidines such as O4-Me-Thd 34 (Figure 3) are poorly repaired [59]. The persistence of the Thd adducts likely contributes to the mutagenicity and carcinogenicity of NDMA. Two adducts, O2-Me-Thd 30 and O4-Me-Thd, blocked DNA synthesis and induced A to G transitions mediated by human DNA polymerase κ (pol κ) [60]. This result echoes that of Singer et al. published in 1983 [61].
The methyl Fapy-dGuo adduct 28 blocked eukaryotic high-fidelity polymerases but can be efficiently bypassed by translesion polymerases. Misreplication products, accounting for 8–29% of total extension products, included C to T/G/A mutations and an interesting one-nucleotide deletion [62].
Wang et al. first investigated the mutagenicity of methyl DNA phosphate adduct 36 (Figure 3) [63]. Due to the stereochemistry of the phosphorus atom, two diastereomers are formed upon DNA phosphate methylation. The (S)-isomer can be efficiently bypassed, causing TT to GT and GC mutations in the flanking TT dinucleotide site. This mutation was induced in an AGT (also termed as Ada)-dependent manner. On the other hand, the (R)-isomer moderately blocked the replication of the DNA synthesis [63].
3.1.5. Methyl DNA Adducts in Human Tissues
The major DNA adducts caused by NDMA incubated with cultured esophagus from human patients were O6-Me-Gua 24 and N7-Me-Gua 27, in a ratio of 0.3. A 10-fold interindividual variation was observed for adduct levels due to NDMA in human esophageal DNA [64]. In the liver DNA of an NDMA-poisoning victim, O6-Me-Gua and N7-Me-Gua were detected at levels of 273–317 and 1363–1373 μmol/mol Gua, respectively. These adducts were not detected in the liver and kidney DNA from unrelated cases [65].
Methyl DNA adducts have been widely detected in human tissues likely due to exposure to various methylating agents that can originate from multiple sources both endogenously and exogenously [66,67,68]. For example, Foiles et al. developed a monoclonal antibody for O6-Me-Gua and used it in a competitive enzyme-linked immunosorbent assay coupled with HPLC resulting in the first identification of O6-Me-Gua in human placental DNA [69]. In Japanese donors, this adduct was detected in both leukocyte and liver DNA. In contrast, O4-Me-Thy 35 was only detected in the liver DNA [70]. Similarly, in the liver and leukocyte DNA of humans who were not exposed to known alkylating agents, O6-Me-Gua and O4-Me-Thy were detected in nearly all the liver DNA samples, at levels of 1.1–6.7 and 0.1–14 adducts/107 nucleotides, respectively. Only O6-Me-Gua was detected in peripheral leukocyte DNA, accounting for 3.6% of that in the liver [71]. Using a radioimmunoassay with a monoclonal antibody against O6-Me-dGuo, this adduct has been detected at relatively high levels in the esophagus and liver DNA of esophageal cancer patients in China who were considered exposed to high levels of dietary N-nitrosamines [72].
Peter Magee would frequently conclude his lectures by noting that NDMA is a potent hepatocarcinogen in rats, but we have insufficient data to conclude that it is a human carcinogen. Now we have massive biochemical, molecular biological, and laboratory animal testing data in support of its potential human carcinogenicity, resulting in increased regulatory vigilance that has decreased NDMA exposure in most situations to extremely low levels. This appears to be an excellent example of cancer prevention through research and application.
3.2. N-Nitrosomethylethylamine (NMEA)
3.2.1. Exposure and Carcinogenicity
NMEA (2, Figure 1) has been found in a variety of processed foods [73,74]. It can be formed as a byproduct during water disinfection with chlorine [75]. It is also one of the contaminants found in some drug products at trace levels [76]. NMEA has been documented as a tobacco constituent at very low concentrations [73,77].
The carcinogenicity of NMEA was demonstrated in rats in 1967. Hepatocellular carcinomas are the primary cancer type induced by NMEA administered in drinking water at doses of 1 or 2 mg/kg body weight per day [6]. As part of an extensive program on the chemistry and biology of N-nitroso compound [5], Lijinsky et al. showed that deuterated NMEA-d3 38 (Figure 4) led to a high incidence of esophageal tumors when compared to undeuterated NMEA administered at an identical dose [78,79].
3.2.2. Metabolism
It is generally considered that NMEA is bioactivated by two α-hydroxylation pathways (Scheme 2). When the α-hydroxylation occurs on the ethyl group (as illustrated by 41), NMEA is converted to methyldiazonium ion 15, the same intermediate as formed from NDMA metabolism; the other product is acetaldehyde 42. When the α-hydroxylation occurs on the methyl group (as illustrated by 43), the ethyldiazonium ion 45 is formed along with formaldehyde 12. The two diazonium ions 15 and 45 can alkylate DNA and form methyl or ethyl DNA adducts correspondingly. A detailed discussion of the pathway by which α-hydroxylation occurs in the ethyl group can be found in the metabolism study of NDEA in Section 3.4.
3.2.3. DNA Adducts Formed by NMEA Metabolism
After a single i.p. injection of NMEA to F344 rats, N7-Me-Gua 27 (Figure 3) was readily detected in the liver, kidneys, and esophageal DNA. However, O6-Me-Gua 24 was only quantifiable in the liver and kidneys. N7-Et-Gua 54 (Figure 5) was also detected in the hepatic DNA [80]. The level of N7-Me-Gua in hepatic DNA exceeded N7-Et-Gua by 170–200 times. N7-Me-Gua occurred at the highest concentration in the liver, followed by the kidneys (15-fold lower), esophagus (100-fold lower), and lung (200-fold lower) [81]. In another study in which F344 rats were administered [1-ethyl-14C]NMEA 39 (Figure 4) by a single i.p. injection, N7-Et-Gua and O6-Et-Gua 52 (Figure 5) were the two major ethyl DNA adducts identified in the liver after 4 h exposure. N3-Et-Gua 51, N3-Et-Ade 47, and N7-Et-Ade 48 were also identified as minor products resulting from NMEA metabolism [82].
Quantitative differences of methylation and/or ethylation in hepatic DNA were clearly observed in F344 rats administered NMEA or the related N-nitrosamines NDMA and NDEA by i.p. injections. Although DNA methylation by NMEA was comparable to that observed upon the administration of an equimolar mixture of NDMA plus NDEA, DNA ethylation by NMEA was ~4-fold lower [83]. Thus, the ratio changes of methyl/ethyl DNA adducts formed by NMEA in the tissue DNA of rats may explain the shift of organ specificity of NMEA-d3 carcinogenicity reported by Lijinsky et al. [78,79]. After a single i.p. injection of [methyl-14C]NMEA-d3 40 (Figure 4) to F344 rats, the levels of N7-Me-Gua decreased by ~30% in the liver but were 160% greater in the esophagus compared to non-deuterated [methyl-14C]NMEA. The increase in esophageal DNA methylation seems to correlate with the increased esophageal carcinogenicity of NMEA-d3 because of the increased α-hydroxylation of its ethyl group since pharmacokinetic differences between NMEA and NMEA-d3 have been ruled out [80].
3.3. N-Nitrososarcosine (NSAR)
3.3.1. Exposure and Carcinogenicity
NSAR (3, Figure 1) has been detected in food such as smoked meat (2–56 μg/kg) [73,84], malt (5.6–11.3 ppb), and beer (trace—6.0 ppb) [85]. It is also one of the carcinogenic components of tobacco smoke (22–460 ng/cigarette) [84] and some smokeless tobacco products (30–550 ng/g) [86,87]. It may also be formed endogenously from nitrosation of sarcosine and dimethylglycine (but not creatine) [88,89,90]. Human exposure to NSAR has been demonstrated by the detection of urinary NSAR, ranging from 0.1 to 3.4 μg/day, in multiple countries [73].
The carcinogenicity of NSAR has been demonstrated in mice and rats. The dietary administration of NSAR caused nasal tumors in mice; oral exposure from drinking water caused esophageal tumors in rats. Liver tumors were observed in newborn mice administered NSAR by i.p. injection [73,84]. Rat studies administering a combination of sarcosine ethyl ester hydrochloride and NaNO2 suggested esophageal carcinogenicity of the NSAR derivative N-nitrososarcosine ethyl ester 70 (Figure 6) [91,92].
3.3.2. Metabolism
NSAR is not extensively metabolized in rats; 88% of dosed NSAR is excreted unchanged in the urine [93]. Metabolic activation of NSAR starts from α-methyl hydroxylation (Scheme 3). The reactive intermediate carboxymethyldiazonium ion 78 and formaldehyde 12 are formed after the spontaneous decomposition of α-hydroxyNSAR 76. Both products resulting from NSAR α-hydroxylation are able to react with DNA [94]. By analogy to N-nitroso-2-oxopropylpropylamine metabolism [95,96,97,98], it also seems possible for NSAR to form the methyldiazonium ion 15 via the intramolecular attack by the diazotate oxygen of 77 on the carbonyl carbon and consecutive decomposition. The proposed methylating agent 15 is likely to be responsible for the observed methyl DNA adducts in DNA exposed to nitrosated glycine derivatives [99,100,101,102,103].
3.3.3. Carboxymethylating and Methylating Intermediates Formed by NSAR Metabolism
O6-Carboxymethylguanine 81 (O6-CM-Gua, Figure 7) has been detected in DNA treated with mesyloxyacetic acid 71 (Figure 6) [99] and glycine reacted with nitric oxide [104] and related compounds including N-(N′-acetyl-L-prolyl)-N-nitrosoglycine 72 (APNG), azaserine 73 (AS), potassium diazoacetate 74 (KDA), and N-nitrosoglycocholic acid 75 [99,100,101,102,103,105]. However, O6-CM-Gua was not detected in physiological concentrations of glycine and nitric oxide, which does not support the hypothesis that the resulting carboxymethylating species is an etiological agent for human gastrointestinal tumors [104]. In addition, O6-Me-Gua 24 (Figure 3) has been detected concomitantly with O6-CM-Gua in vitro; it is proposed to be formed via the putative methyldiazonium ion 15 after decarboxylation (Scheme 3) [99,100,101,102,103,104]. This may also partially explain the reported relatively high occurrence of O6-Me-Gua in human gastrointestinal DNA [72,106,107,108,109,110] since nitrosation of glycine—abundant in food sources—could occur there [111,112].
The other carboxymethyl DNA adducts N6-(carboxymethyl)-2′-deoxyadenosine 79 (N6-CM-dAdo, Figure 7), N4-(carboxymethyl)-2′-deoxycytidine 80 (N4-CM-dCyd), N3-(carboxymethyl)thymidine 83 (N3-CM-Thd), and O4-(carboxymethyl)thymidine 84 (O4-CM-Thd) have also been detected in vitro in 74 (KDA)-treated calf thymus DNA [113,114]. In cultured human skin fibroblasts and human colorectal carcinoma cells exposed to azaserine 73, O6-(carboxymethyl)-2′-deoxyguanosine 82 (O6-CM-dGuo), N6-CM-dAdo, and O6-Me-dGuo were simultaneously measured, with O6-CM-dGuo predominating among the three determined adducts [115].
3.3.4. Mutagenicity and Genotoxicity of Carboxymethyl DNA Adducts
KDA 74 (Figure 6) caused GC to AT transitions in the p53 gene in equal amounts to GC to TA and AT to TA transversions. This contrasts with the methylating agent methylnitrosourea which causes predominantly GC to AT transitions. The difference is hypothesized to be due to the different mutagenicity of O6-CM-dGuo 82 compared with O6-Me-dGuo [105].
O6-CM-dGuo is not a substrate of bacterial and mammalian AGT [103] but can be repaired by human O6-methylguanine-DNA methyltransferase (MGMT) [116]. It may also be repaired by nucleotide excision repair (NER) [117]. In E. coli cells, O6-CM-dGuo strongly impeded DNA replication. It caused exclusive G to A transitions during the bypass that uniquely requires the involvement of DNA polymerases IV and V [118]. Pol η and ζ were also likely involved in bypassing O6-CM-dGuo lesions [119].
In a primer extension assay, N4-CM-dCyd 80 (Figure 7) strongly blocked the extension reaction, causing relatively few C to A or T mutations; N6-CM-dAdo 79 blocked the extension reaction to a lesser extent but caused a higher frequency of A to T mutations [114]. Pol η was able to readily bypass N6-CM-dAdo lesions with high fidelity. However, it bypassed N4-CM-dCyd lesions inefficiently with a substantial frequency of dCMP and dAMP misincorporation [120]. N3-CM-Thd 83 and O4-CM-Thd 84 were highly resistant to the bypass of the yeast Pol η. Preferential T to C mutations caused by those two lesions were observed [120].
The five carboxymethyl DNA adducts N6-CM-dAdo, N4-CM-dCyd, O6-CM-dGuo, N3-CM-Thd, and O4-CM-Thd (Figure 7) were investigated individually for their mutagenicity and genotoxicity in human HEK293T cells. Among the five adducts, O6-CM-dGuo was moderately genotoxic by blocking DNA replication and weakly mutagenic by inducing 6.4% G to A mutations. In contrast, two Thd adducts, N3-CM-Thd and O4-CM-Thd, were strongly mutagenic, inducing 81% T to A mutations and 68% T to C mutations, respectively [119].
3.3.5. Carboxymethyl DNA Adducts in Human Tissues
Based on the evidence of human exposure to NSAR and its potential metabolic activation mechanism, it is reasonable to anticipate the detection of carboxymethyl DNA adducts in humans. In the blood DNA of healthy volunteers restricted to a standardized high-meat diet, O6-CM-Gua 81 (Figure 7) was detected at levels of 35–80 adducts/108 nucleotides [104]. The formation of O6-CM-Gua in the colonic exfoliated cells of healthy volunteers has been positively associated with the consumption of red meat. Levels of O6-CM-Gua were also positively correlated with the concentrations of apparent total fecal N-nitroso compounds [121]. These results provide some evidence supporting the etiological role of dietary N-nitroso compounds such as NSAR or nitrosated glycine derivatives, which can decompose to form carboxymethylating and methylating agents, in human gastrointestinal carcinogenesis. However, further studies are required.
Interestingly, we could not detect N7-(carboxymethyl)guanine 85 (N7-CM-Gua, Figure 8) in any human liver samples but its analog N7-(2′-carboxyethyl)guanine 86 (N7-CE-Gua) occurred at a relatively high mean level of 373 ± 320 fmol/μmol Gua in 100% of the same human livers. One possible source of N7-CE-Gua is 3-(methylnitrosamino)propionic acid 87 (MNPA) metabolism [94].
3.4. N-Nitrosodiethylamine (NDEA)
3.4.1. Exposure and Carcinogenicity
Human exposure to NDEA (4, Figure 1) occurs through water, food, and cosmetics [18,73]. Some batches of drugs including irbesartan, losartan, and valsartan have been recalled due to contamination with NDEA since 2018 [16]. It has also been identified in tobacco smoke, with greater concentrations found in sidestream smoke compared to mainstream smoke [73]. However, the levels of NDEA in current cigarette smoke and smokeless tobacco products are extremely low [77].
The carcinogenicity of NDEA has been demonstrated in multiple laboratory animal species including mice, rats, Syrian golden hamsters, guinea pigs, rabbits, dogs, pigs, and monkeys [73]. Tumors caused by NDEA primarily occurred in the liver, often with lung metastases in mice and rats after oral administration [73]. Peto et al. conducted a large study of rats treated with NDEA in drinking water for their lifetime starting from 6 weeks of age. Various types of liver cancers were observed primarily in the NDEA-treated rats; esophageal cancers were also observed exclusively with NDEA but not NDMA treatment. A few nasopharyngeal tumors were also caused by NDEA in the rats [21,22]. The IARC upgraded NDEA to Group 2A in 1987, but as is the case with NDMA, re-evaluation may be appropriate.
3.4.2. Metabolism
NDEA metabolic activation for carcinogenicity is principally catalyzed by P450 2E1 and P450 2A6 [24,122,123]. When the P450-catalyzed hydroxylation occurs at the α-carbon of the ethyl group of NDEA, the electrophilic ethyldiazonium ion 45 (Scheme 4) is formed after decomposition of the unstable intermediate ethyl diazohydroxide 44. Intermediate 44 reacts with DNA producing ethyl DNA adducts such as N7-Et-Gua and O6-Et-Gua. Acetaldehyde 42 is also formed by NDEA α-hydroxylation [124]. When the β-carbon is hydroxylated, the reactive intermediate 2-hydroxyethyldiazonium ion 100 can be formed by a secondary α-hydroxylation on the other ethyl group of NDEA (as illustrated by 90). The 2-hydroxyethyldiazonium ion 100 alkylates DNA and forms DNA adducts such as N7-HOEt-Gua 69 (Figure 5) that have been detected in the hepatic DNA of NDEA-treated rats [82]. The denitrosation reaction competes with bioactivation in NDEA metabolism, suggesting a dual role played by P450s in the biotransformation of NDEA [35].
3.4.3. Ethyl DNA Adducts Formed by NDEA Metabolism
Ethyl DNA adducts are readily formed by NDEA metabolism in vivo after a single administration of the carcinogen [125,126]. Although the main ethyl DNA adducts (as shown in Figure 5) were ethyl DNA phosphate adducts B1p(Et)B2 61 (58%), ethyl DNA base adducts after deglycosylation were also produced as expected. N7-Et-Gua 54 (12%), O6-Et-Gua 52 (8%), O2-Et-Thy 55 (7%), and N3-Et-Ade 47 (4%) represented the most abundant ethyl DNA base adducts in the same sample mixture. The other minor base adducts observed were N1-Et-Ade 46, N7-Et-Ade 48, O2-Et-Cyt 49, N3-Et-Cyt 50, N3-Et-Gua 51, N3-Et-Thy 57, and O4-Et-Thy 59 [125,126]. In addition, trace levels of N7-HOEt-Gua 69 were also detected in the hepatic DNA of rats exposed to NDEA, comprising less than 2% of N7-Et-Gua 54 [82].
Variation in ethyl DNA base adduct accumulation has been investigated in the liver DNA of rats exposed to NDEA in drinking water for 77 days [127,128]. O4-Et-Thd 60 accumulated in the first 28 days and persisted at such levels until the end of the experiment. O6-Et-dGuo 53, in contrast, did not accumulate after repeated exposure to NDEA. The concentration of O6-Et-dGuo was highest after 2 days of administration of NDEA and decreased throughout the 77-day study course [127,128]. This was attributed to a rapid repair mechanism for O6-Et-dGuo when compared with O4-Et-Thd [129].
3.4.4. Mutagenicity and Genotoxicity of Ethyl DNA Adducts
The dynamic change in the accumulation of O6-Et-dGuo and O4-Et-Thd after continued exposure to NDEA clearly indicates the different repair mechanisms of these ethyl DNA adducts. O6-Et-dGuo can be repaired by AGT without the excision of the DNA base [130]. On the contrary, O4-Et-Thd is not a good substrate of rat liver AGT; the half-life of O4-Et-Thd reaches 11–19 days in vivo [41,129,131,132]. A similar result was also observed for O2-Et-Thd 56 (Figure 5) [133]. The DNA repair protein AGT repairs O4-Et-Thd, however, to a lesser extent compared to its analog O4-Me-Thd 34 (Figure 3) [134,135,136].
Diverse mutagenic consequences have been observed for the 3 regioisomers of ethyl thymidine adducts. O4-Et-Thd, a major-groove lesion, induces a large number of A to G transitions when incorporated into polynucleotides. However, O2-Et-Thd (a minor-groove lesion) was only slightly mutagenic and N3-Et-Thd 58 (Figure 5) did not induce mutations [137]. O4-Et-Thd is also genotoxic by moderately blocking DNA replication with a bypass efficiency of 20–33% in human cells [138].
The DNA polymerases responsible for bypassing the three regioisomeric ethyl thymidine lesions have been extensively investigated. DNA polymerase I and RNA polymerase II can recognize the ethyl Thd lesions and direct the dGMP misincorporation opposite to O4-Et-Thd but not O2-Et-Thd [139,140]. Human Pol η similarly can bypass all 3 ethyl Thd lesions and generate the replication product with a substantial frequency of A to G transitions caused by O4-Et-Thd [141,142,143]. In E. coli cells, both Pol IV and Pol V are essential for the misincorporation of dCMP opposite to O2-Et-Thd, whereas Pol V is necessary for the T to A transversions caused by this lesion [144]. In human cells, Pol η and ζ are the predominant polymerases mainly responsible for bypassing O2- and O4-Et-Thd, and causing the respective T to A/G and T to C mutations [138,145].
3.4.5. Ethyl DNA Adducts in Human Tissues
Ethyl DNA adducts have been detected in various human tissues. They may arise from the metabolism of the N-nitrosamines NDEA and NMEA, or from structurally unknown directly acting ethylating agents present in tobacco and tobacco smoke [146]. Levels of O2-, N3-, and O4-Et-Thd (56, 58, and 60, Figure 5) in the leukocyte DNA of 20 smokers were 44.8 ± 52.0, 41.1 ± 43.8, and 48.3 ± 53.9 adducts/108 nucleotides, respectively, significantly exceeding those in the 20 nonsmokers. The formation of each ethyl thymidine adduct was statistically associated with that of the other two ethyl Thd adducts [147]. O4-Et-Thd was detected in the lower respiratory tract DNA of smokers but not in nonsmokers [148]. O4-Et-Thd levels in the lung DNA of smokers (3.8 adducts/108 nucleotides) were higher (p < 0.01) than in nonsmokers (1.6 adducts/108 nucleotides) [149]. The smoking-related formation of this adduct in lung DNA was further confirmed in Hungarian lung cancer patients [150].
Similarly, levels of N3-Et-Ade 47 and N7-Et-Gua 54 (Figure 5) were also significantly higher in the leukocyte DNA from smokers (16.0 ± 7.8 and 9.7 ± 8.3 adducts/108 nucleotides, respectively) than those from nonsmokers (5.4 ± 2.6 and 0.3 ± 0.8 adducts/108 nucleotides, respectively). The levels of N3-Et-Ade and N7-Et-Gua were positively correlated [151]. N3-Et-Ade and N7-Et-Gua were also detected in the salivary DNA of smokers and nonsmokers. The occurrence of N7-Et-Gua in the saliva of smokers (14.1 ± 8.2 adducts/108 nucleotides) was significantly higher than that of nonsmokers (3.8 ± 2.8 adducts/108 nucleotides, p < 0.0001). The levels of N7-Et-Gua were also strongly associated with tobacco smoking [152]. However, there are some contradictory results that question the robustness of applying ethyl DNA adducts as biomarkers for smoking-related cancer etiology studies. We have quantified the levels of N7-Et-Gua in human leukocyte DNA from 30 smokers and 30 nonsmokers. No statistical difference was observed in the levels of this adduct in the leukocyte DNA from the two subject groups (smokers: 49.6 ± 43.3 fmol/μmol Gua; nonsmokers: 41.3 ± 34.9 fmol/μmol Gua) [153].
In addition to human tissue DNA, ethyl DNA adducts have been detected in human urine showing a potential correlation with smoking status. Urinary excretion of N3-Et-Ade has been observed to increase 5–8-fold with tobacco smoking [154,155] but not from dietary origin [156]. At 50%, the mean level of N-terminal N-ethylvaline in the hemoglobin of smokers was also significantly higher than in the nonsmokers [157].
Other than smoking, dietary exposure to ethylating agents derived from N-nitrosamines or possibly other sources may play an important role in human carcinogenesis, especially in some particular geographic areas. In Linxian, the incidence of esophageal cancer was comparably higher than its surrounding area in the same region of China. In the esophageal and hepatic DNA of esophageal cancer patients from Linxian, relatively high levels of O6-Et-dGuo 53 (Figure 5) were detected by radioimmunoassay. This appeared to be consistent with the relatively high exposure levels of dietary N-nitrosamines in those who lived in this area [72].
The endogenous formation of ethylating agents may be important in the formation of ethyl DNA adducts in the human liver. In the liver DNA of 15 autopsy specimens, O4-Et-Thd occurred at levels of 0.5–140 adducts/107 nucleotides [71]. It was not detected in the peripheral leukocyte DNA in the same study [70,71]. N7-Et-Gua was detected in 25 of 26 human hepatic DNA samples, occurring at a level of 42.2 ± 43.0 fmol/μmol Gua [158].
3.5. N-Nitroso-di-n-propylamine (NDPA)
3.5.1. Exposure and Carcinogenicity
3.5.2. Metabolism
NDPA is metabolized via α-, β-, and γ-hydroxylation of the propyl group (Scheme 5). Of the three metabolic pathways, α-hydroxylation is regarded as the primary route for NDPA bioactivation [159].
Catalyzed primarily by P450 2E1 and P450 2B1 [160,161], N-nitroso-1-hydroxypropylpropylamine 101 is formed by the α-hydroxylation of NDPA. It decomposes to generate the reactive intermediates propyl diazohydroxide 102 and propionaldehyde 106 [162]. The diazohydroxide further reacts forming the electrophilic carbocations 104 and 107 via the intermediate propyldiazonium ion 103. The solvolysis products of the two carbocations—1-propanol 105 and 2-propanol 108—have been detected in vitro [163,164].
The β-hydroxylation of NDPA generates N-nitroso-2-hydroxypropylpropylamine 109 (NHPPA); its glucuronide is excreted, accounting for 5% of the administered NDPA in 24 h rat urine [165]. NHPPA can be further oxidized to N-nitroso-2-oxopropylpropylamine 110 (NOPPA) [163,165]. NOPPA can be reduced back to NHPPA, accounting for nearly 50% of the total dose of NOPPA in 24 h rat urine [165,166]; carbonyl reduction was similarly observed in the metabolism of relevant metabolites such as N-nitroso-(2-hydroxypropyl)-(2-oxopropyl)amine 118 (Figure 9) [167]. NOPPA also undergoes a secondary α-hydroxylation (as illustrated by 111), generating the methyldiazonium ion 15 and acetic acid 113 via an oxadiazoline intermediate resulting from a spontaneous intramolecular attack by the diazotate oxygen on the carbonyl carbon of 112 [95,96,97,98]. P450 2E1 and P450 2B1 play a major role in the consecutive hydroxylation of NOPPA [97].
The γ-hydroxylation of NDPA forms N-nitroso-3-hydroxypropylpropylamine 114 and N-nitrosopropyl-(carboxyethyl)amine 115. They were detected as minor metabolites of NDPA in isolated rat hepatocytes [168].
3.5.3. DNA Adducts Formed by NDPA Metabolism
In 1971 and 1973, Kruger first investigated the alkylation of nucleic acids using [14C]NDPA. The simultaneous detection of N7-Me-Gua 27 (Figure 3) and N7-(n-propyl)guanine 119 (N7-n-Pr-Gua, Figure 10) in the liver RNA of rats treated with NDPA strongly suggested the metabolism of NDPA by both α- and β-hydroxylation pathways [96]. [14C]N7-Me-Gua was only detectable in rat liver RNA and DNA when the rats were treated with [α-14C]NDPA, in agreement with the methylating agent arising from the β-hydroxylation of NDPA [170]. When the rats were treated with [β-14C]NDPA, only [14C]N7-n-Pr-Gua was detectable, consistent with the metabolism mechanism of NDPA α-hydroxylation [96]. Similarly, [3H]N7-Me-Gua was detected in the rat liver after application with [3H]NOPPA [171].
Alkylation of DNA has been clearly demonstrated using [α-14C]NDPA in vitro [172]. However, the chemical characterization of NDPA-DNA adducts is limited. Only some related studies have been reported. Kokkinakis in 1992 reported a study of methyl and hydroxypropyl DNA adducts in the tissues of hamsters and rats after a single s.c. dose of 3H-labeled N-nitroso-bis(2-hydroxypropyl)amine 116 (Figure 9). Methyl DNA adducts were preferentially formed over hydroxypropyl DNA adducts at low doses (100–500 mg/kg body weight) but became secondary at higher doses. Both adducts occurred at their highest concentrations in the liver, the primary metabolic activation site. The methyl DNA adducts identified in this study were N7-Me-Gua 27 and O6-Me-Gua 24 (Figure 3); the hydroxypropyl DNA adducts were N7-(2-hydroxypropyl)guanine 120, O6-(2-hydroxypropyl)guanine 121, and O6-(1-methyl-2-hydroxyethyl)guanine 122 (Figure 10) [173]. The formation of N7-Me-Gua and O6-Me-Gua was also confirmed in hamsters and rats treated with the NDPA derivatives N-nitroso-bis(2-oxopropyl)amine 117 and N-nitroso-(2-hydroxypropyl)-(2-oxopropyl)amine 118 (Figure 9), both of which are pancreatic carcinogens in hamsters [174,175]. Adduct 120 was also detected in the tissues of hamsters and rats treated with 117 and 118 [175].
3.6. N-Nitrosodiethanolamine (NDELA)
3.6.1. Exposure and Carcinogenicity
NDELA (6, Figure 1) is an environmentally prevalent N-nitrosamine found in cosmetics. It can also be detected in some food products, synthetic cutting fluids, and tobacco and tobacco smoke [176,177,178,179].
The carcinogenicity of NDELA has been extensively studied in rats, mice, and hamsters [73,179]. After oral administration, NDELA induced primarily liver tumors in rats. It also induced lung tumors in mice and some nasal tumors in rats. In hamsters, NDELA treatment resulted in nasal cavity tumors and tracheal tumors regardless of the administration pathways (s.c. injection, topical application, and oral swabbing) [73,179].
3.6.2. Metabolism
The distribution of NDELA in Osborne-Mendel rats has been studied using two administration pathways. After oral administration, NDELA was absorbed and distributed rapidly and reached a peak concentration at 8 h; after topical application, NDELA was slowly absorbed but rapidly distributed as when dosed orally. NDELA was excreted mainly in the urine as the unchanged form and one metabolite [180,181]. The excreted NDELA also represented a high percentage (60–90%) in the urine of male Sprague-Dawley rats administered NDELA in drinking water [182]. A similar high urinary excretion rate (73–89%) of unchanged NDELA was also determined in rats treated percutaneously and intratracheally [19]. However, the absorption rate of NDELA was significantly lower in Syrian golden hamsters. After s.c. injection, 49% and 11% of the dose appeared in the urine and feces, respectively, in 16 h; 34% and 6% after oral swabbing; and only 21% and 4% were detected after skin application [183].
An investigation of the rat urinary metabolites of NDELA suggested only one compound containing the nitroso moiety, which was identified as N-nitroso-(2-hydroxyethyl)glycine 135 (NHEG, Scheme 6). It represented 6% of the dosed NDELA in the rats [184]. The glucuronide of NDELA was also identified in rat urine after gavage [185]. However, no sulfate derivatives were observed even though NDELA sulfate was considered a possible activated metabolite that could react with DNA to form 2-hydroxyethyl adducts (as shown in Figure 5) [185,186,187]. N-Nitroso-2-hydroxymorpholine 137 (NHMOR) was also observed as a minor metabolite in vitro in rat liver S9 supernatant [188,189].
The formation of NHEG 135 and NHMOR 137 strongly implies the important role that β-hydroxylation may play in the bioactivation of NDELA in vivo (Scheme 6). This reaction is catalyzed primarily by P450 2E1 [188]. NHMOR can be further metabolized by α-hydroxylation on the two methylene groups. When the α-hydroxylation occurs on the 3-carbon, the major metabolite has been identified as glyoxal 133 formed via 139 and 140; when the α-hydroxylation occurs on the 5-carbon, the major metabolite is 2-acetoxyacetaldehyde 144 formed via 142 and 143 [188]. It is noteworthy that NHMOR also arises from NMOR metabolism which is discussed in detail in Section 4.3.2.
However, several lines of evidence raise questions regarding the importance of β-hydroxylation in the carcinogenesis of NDELA. Although it is a stable precursor to potential DNA alkylating agents such as glyoxal, NHMOR was inactive or marginally carcinogenic to rats or mice when administered in drinking water [190]. Only the glyoxal-deoxyguanosine adduct 145 (N1,N2-glyoxal-dGuo, Figure 11) was observed in vitro in NHMOR-incubated calf thymus DNA (up to 48 h) and in vivo in the liver DNA of rats given NHMOR at a single dose by gavage for 4 h [191]. The levels of N1,N2-glyoxal-dGuo formed by NHMOR were also lower than those formed by NDELA [192]. Those findings taken together suggest that additional metabolic activation pathways such as α-hydroxylation are involved in the carcinogenesis of NDELA [193].
Catalyzed primarily by P450 2E1, hydroxylation occurs on the α-carbon of NDELA and forms the 2-hydroxyethyldiazonium ion 127 and glycolaldehyde 130 after spontaneous decomposition [188,194]. The 2-hydroxyethyldiazonium ion forms the carbocation 128 after the loss of H2O and subsequently yields the solvolysis product ethylene glycol 129, which can also undergo microsome-mediated oxidation to glyoxal 133. The carbocation 128 also undergoes elimination and forms acetaldehyde 42 [188,195]. O6-(2-Hydroxyethyl)-2′-deoxyguanosine 68 (O6-HOEt-dGuo, Figure 5) arising from NDELA metabolism has been detected in rat liver DNA; it is not derived from NHMOR metabolism [191]. Deuteration on the α-carbon of NDELA greatly decreased the formation of glycolaldehyde 130 and O6-HOEt-dGuo 68, whereas β-deuteration oppositely affected the formation of those products. This further suggests that O6-HOEt-dGuo mainly results from NDELA α-hydroxylation [191,194]. One unexpected finding was that glycolaldehyde 130, other than being classically converted to glycolic acid 131 and oxalic acid 132, was also transformed to glyoxal 133 via the catalysis of P450 2E1 [188,194]. The collective evidence indicates the necessity of the α-hydroxylation of NDELA for DNA adduct formation.
3.6.3. DNA Adducts Formed by NDELA Metabolism
O6-HOEt-dGuo 68 and N7-HOEt-Gua 69 (Figure 5) were characterized in the reaction mixture of N-nitroso-3-acetoxy-2-hydroxymorpholine 138 (Scheme 6) with dGuo. However, neither of these adducts was detected in the reaction mixture of N-nitroso-5-acetoxy-2-hydroxymorpholine 141 with dGuo. This suggested that the α-hydroxylation of NHMOR on the 5-carbon was unlikely to produce DNA adducts, whereas NHMOR α-hydroxylation on the 3-carbon might yield a carcinogenic outcome [195]. However, this seems not to be supported by an in vivo study, in which no 2-hydroxyethyl guanine adducts were detected in the liver DNA of rats treated with NHMOR [188]. In contrast, 2-hydroxyethyl guanine adducts were tentatively identified in the hydrolysates of the liver DNA of rats treated with NDELA by gavage [185]. One adduct was later characterized as O6-HOEt-dGuo using the synthesized authentic standard. It was detected in vivo in the liver DNA of rats treated with NDELA [191,194,196]. Taken together, 2-hydroxyethyl adducts formed by NDELA are reasonably considered to result from the α-hydroxylation pathway rather than the β-hydroxylation pathway.
Glyoxal DNA adducts formed by NDELA and its analogs were readily detected in vitro and in vivo. In the reaction with dGuo in vitro, N1,N2-glyoxal-dGuo 145 (Figure 11) was formed as the major adduct (65%) by N-nitroso-3-acetoxy-2-hydroxymorpholine 138; N1,N2-etheno-dGuo 146 was formed as the major adduct (44%) by the less-reactive N-nitroso-5-acetoxy-2-hydroxymorpholine 141 [195]. N1,N2-glyoxal-dGuo was also detected in the liver DNA of rats treated with NDELA or NHMOR [192,196]. A few analogs of NDELA also caused the same N1,N2-glyoxal-dGuo adduct in the rat liver DNA [192]. However, due to the complex potential origins of glyoxal, the formation of N1,N2-glyoxal-dGuo does not necessarily reflect the preference of metabolic pathways of NDELA.
3.7. N-Nitrosodi-n-butylamine (NDBA)
3.7.1. Exposure and Carcinogenicity
NDBA (7, Figure 1) has been found in agricultural products, fish, processed meats, seasonings, and contaminated water [12,73]. It was also formed at trace levels during the production of the drug ranitidine [197]. Some early data suggested the presence of NDBA in tobacco smoke but with no clear evidence in recent studies [73].
NDBA was classified by IARC in 1978 and reaffirmed in 1987 as a group 2B carcinogen. It is carcinogenic to the esophagus and bladder of laboratory animals including mice, rats, Syrian golden hamsters, and guinea pigs. It also causes liver and forestomach tumors [73,198,199,200,201,202]. In 1983, Lijinsky and Reuber reported that NDBA, even though much weaker than NDPA, induced liver tumors in 60% of rats administered by gavage for 83 weeks. It also induced forestomach (50%) and bladder tumors (35%) [203]. Even after a short-term (2 weeks) exposure to NDBA, preneoplastic lesions were positively found in the liver, esophagus, forestomach, and bladder of rats 52 weeks post-treatment [204]. Mice with a p53 gene knockout had increased susceptibility to esophageal and bladder carcinogenesis caused by NDBA [205].
3.7.2. Metabolism
By analogy to other N-nitrosodialkylamines, NDBA requires metabolic activation to exert its carcinogenicity. Four metabolic pathways can occur in NDBA metabolism (Scheme 7). They are α-, β-, γ-, and δ-hydroxylation of the butyl group of NDBA, among which α-hydroxylation has been suggested to be primarily involved in NDBA carcinogenesis [206].
In an incubation mixture of NDBA with rat liver microsomes, the principal metabolite retaining the N-nitroso group was the γ-hydroxylation product N-nitroso-(3-hydroxybutyl)butylamine 157. The α-hydroxylation products butyraldehyde 152 and/or 1-butanol 154 and 2-butanol 155 were also detected, indicating the formation of the butyl carbocations 150 and 151 [206,207]. In rats, NDBA was extensively metabolized with no unchanged compounds being detected in the urine. Two major metabolites N-nitroso-(3-carboxypropyl)butylamine 161 and N-nitroso-(3-hydroxybutyl)butylamine 157 resulted from δ- and γ-hydroxylation, respectively. The third minor metabolite was the β-hydroxylation product N-nitroso-(2-hydroxybutyl)butylamine 156. All metabolites were detected in the urine as such and as their gluconides [208]. One of the major urinary metabolites 161 arises from the consecutive oxidation of the initial δ-hydroxylation product N-nitroso-(4-hydroxybutyl)butylamine 160 (BBN), a urothelial carcinogen [209]. It can be further converted to several minor metabolites such as 162, 163, and 164 by β-oxidation and subsequent biotransformations [210,211]. In contrast to rats, the primary metabolite of NDBA in hamsters was the glucuronide of N-nitroso-(3-hydroxybutyl)butylamine 157 [212].
In the urine of rats treated with NDBA, the end products of NDBA-GSH conjugates were also detected. They are N-acetyl-S-butyl-L-cysteine 153 resulting from α-hydroxylation of NDBA and N-acetyl-S-3-hydroxybutyl-L-cysteine 158 and N-acetyl-S-3-oxobutyl-L-cysteine 159 that are hypothesized to result from γ-hydroxylation of NDBA followed by a secondary α-hydroxylation [213].
3.7.3. DNA Adducts Formed by NDBA Metabolism
The butyl carbocation 150 (Scheme 7) resulting from the α-hydroxylation of NDBA is considered to be the alkylating agent which attacks DNA forming n-butyl DNA adducts. In the liver DNA of rats treated with 185 mg/kg NDBA by a single i.p. dose, O6-(n-butyl)guanine 123 (O6-n-Bu-Gua, Figure 10) has been detected at a concentration of 0.34 μmol/mol Gua [214].
The sequential α-hydroxylation of NDBA metabolites such as BBN 160 can form the other alkylating species and react with DNA to cause lesions. In the urothelial and hepatic DNA of rats, both O6-n-Bu-Gua 123 and O6-(4-hydroxybutyl)guanine 124 (O6-(4-OH-n-Bu)-Gua, Figure 10) were detected at 17.9 ± 7.23 and 12.2 ± 7.01 μmol/mol Gua, respectively, after 24 h treatment with a single oral dose of 120 mg BBN. O6-n-Bu-Gua did not accumulate after repeated exposure to a lower dose of BBN; O6-(4-OH-n-Bu)-Gua was not detected in the same study [215].
4. Metabolic Activation and DNA Interactions of Carcinogenic Cyclic N-Nitrosamines
Three carcinogenic cyclic N-nitrosamines (NPYR, NPIP, and NMOR, Figure 1) are discussed here due to their common human exposure. They are introduced in order based on their increasing structural complexity.
4.1. N-Nitrosopyrrolidine (NPYR)
4.1.1. Exposure and Carcinogenicity
NPYR (8, Figure 1) is a simple symmetric cyclic N-nitrosamine. It has been extensively investigated as a model compound for studies of N′-nitrosonornicotine (NNN), and in its own right due to its common occurrence in food (1.5 ± 0.2 ng/g) and water (5.5 ± 2.6 ng/L) [9]. It is also present in tobacco smoke and smokeless tobacco products [73,77,216]. Endogenous nitrosation is considered to be another possible pathway to form NPYR, due to the relatively high human exposure to its precursor pyrrolidine, which is excreted to the extent of ~20 mg per day in the urine [217].
NPYR is a strong hepatic carcinogen in mice and rats. The IARC reviewed some early data on NPYR carcinogenicity in 1978 [73]. Since then, some new data have been reported. In rats administered NPYR in drinking water, it primarily caused hepatocellular carcinomas, many of which metastasized [218,219]. A dose-response relationship of hepatic tumor formation was observed in rats administered NPYR in drinking water [220]. NPYR also induced lung tumors in A/J mice after i.p. injection [221]. In Syrian golden hamsters, NPYR induced tracheal and nasal cavity tumors when administered intraperitoneally [222] but tracheal papillomas and hepatic neoplastic nodules when fed in the diet [223].
4.1.2. Metabolism
NPYR is metabolized extensively in rats, with only ~1% of unchanged NPYR detected in the urine [224]. It is ultimately excreted as volatiles such as CO2 and N2 [224,225]. To exert its carcinogenicity, NPYR requires metabolic activation primarily catalyzed by P450 2E1 [224,226]. The identification of 2-hydroxytetrahydrofuran 171 (2-hydroxyTHF, Scheme 8) [227,228,229], crotonaldehyde 177 [230], and 3-hydroxy-1-nitrosopyrrolidine 178 (3-hydroxyNPYR) [231,232] as metabolites of NPYR in rats indicated the occurrence of the α- and β-hydroxylation pathways of NPYR metabolism. The nearly diminished carcinogenicity of the NPYR analog 2,5-dimethyl-N-nitrosopyrrolidine in rats suggested the importance of α-hydroxylation for the metabolic activation of NPYR to express its ultimate carcinogenicity.
Catalyzed primarily by P450 2E1 [224,226], NPYR is metabolized via α-hydroxylation to form 2-hydroxyNPYR 167, which decomposes spontaneously to the 4-oxobutyldiazonium ion 169 through the intermediate 4-oxobutyl diazohydroxide 168. After the loss of one molecule of N2 from the diazonium ion, carbocations 172 and 174 and the oxonium ion 170 are suggested to be the reactive intermediates in forming the solvolysis products 4-hydroxybutanal 173 in equilibrium with 2-hydroxyTHF 171 and 3-hydroxybutanal 175 [227,228,229] and the elimination product crotonaldehyde 177 [230,233,234,235]. Among those metabolites, 2-hydroxyTHF 171 is the most prevalent one [227].
α-Hydroxylation of NPYR predominated in explanted esophagus from rats and humans [236], cultured human colon [237], and cultured bladder cells from rats and humans [238]. The rate of NPYR α-hydroxylation by rat hepatic microsomes was 1.43 nmol/min/mg protein [228]; this rate was lower in human hepatic microsomes (0.68 nmol/min/mg protein) [239]. The apparent Km for rat lung microsomes and post-microsomal supernatant was approximately 20 mM but was much smaller for rat liver microsomes (0.36 mM) [229,240]. NPYR α-hydroxylation was inducible in rats and hamsters that were pre-treated with Aroclor or ethanol [228,241,242].
4.1.3. DNA Adducts Formed by NPYR Metabolism
As depicted in Scheme 8, the butanal carbocation 172 is considered to form the oxonium ion 170 and the isomeric carbocation 174 after rearrangement. All 3 reactive species can react with DNA and form the corresponding DNA adducts. Crotonaldehyde 177, the elimination product of the isomeric carbocation 174, also reacts with DNA.
DNA Adducts Formed by NPYR-Derived Carbocations
In 1982, Hunt and Shank first reported the detection of a fluorescent adduct, which was formed dose-dependently in the liver DNA of rats treated with NPYR [243]. We later structurally characterized this adduct to be 2-amino-6,7,8,9-tetrahydro-9-hydroxypyrido [2,1-f]purine-4(3H)-one 179 (N7,8-butano-Gua, Figure 12) [244], which was likely formed by simultaneous alkylation at Gua-N7 and cyclization at Gua-C8 [245,246]. N7,8-Butano-Gua was the predominant adduct formed among the NPYR-derived DNA adducts in the liver DNA of rats (see Table 1) [247]. It occurred most abundantly in the target organ liver but was also detected in the kidneys and lungs of NPYR-treated mice, rats, and hamsters [248]. In rats treated with NPYR by intragastric administration, this adduct peaked at 12–24 h after dosing in a dose-dependent manner and persisted for at least 3 days [248]. N7,8-Butano-Gua occurred to a higher extent in the RNA than in the DNA of rat liver (see Table 1), suggesting that RNA could be superior for biomarker studies of this lesion [249]. It was also detected in rat urine [249].
Two acyclic adducts—N7-(4-oxobutyl)guanine 180 and N7-(3-carboxypropyl)guanine 181—can also be formed by the carbocation resulting from NPYR α-hydroxylation. They were both identified in vitro in calf thymus DNA incubated with α-acetoxy-N-nitrosopyrrolidine 165 (α-acetoxyNPYR, Scheme 8); N7-(4-oxobutyl)guanine predominated among these two adducts [245,246]. There was little evidence in support of the cyclization of N7-(4-oxobutyl)guanine to N7,8-butano-Gua 179 [245]. N7-(4-Oxobutyl)guanine occurred at about one-third of the level of N7,8-butano-Gua, at a concentration of 643 ± 9 μmol/mol Gua in the hepatic DNA of rats administered NPYR by i.p. injection [250]. A similar concentration (603 μmol/mol Gua) was also observed in the liver DNA but not the liver RNA of rats administered NPYR by gavage [249].
DNA Adducts Formed by an NPYR-Derived Oxonium Ion
The NPYR-derived oxonium ion 170 reacts with DNA forming a group of tetrahydrofuranyl DNA adducts, some of which have been identified both in vitro and in vivo. Quantitation of those adducts has been performed by reducing them to the corresponding 4-hydroxybutyl adducts (see Table 1).
In the reaction of dGuo and calf thymus DNA with α-acetoxyNPYR 165, N2-(tetrahydrofuran-2-yl)-2′-deoxyguanosine 185 (N2-THF-dGuo, Figure 12) was characterized as the first identified tetrahydrofuranyl DNA adduct derived from NPYR metabolism [253,254]. The levels of this adduct exceeded those of other adducts formed by α-acetoxyNPYR in the in vitro DNA hydrolysate samples [253]. After neutral thermal hydrolysis, 2-hydroxyTHF 171 (Scheme 8) was released as the major product from the treated DNA in vitro; N2-THF-dGuo 185 is considered the major precursor of this product [235]. The reduction of N2-THF-dGuo forms N2-(4-hydroxybutyl)-2′-deoxyguanosine 190 (N2-(4-HOB)-dGuo, Figure 12), which occurred in relatively low abundance compared to other types of NPYR DNA adducts in the liver DNA of rats. However, it remains one of the most abundant adducts among those formed by the oxonium ion 170 [247,252].
The other tetrahydrofuranyl DNA adducts N6-(tetrahydrofuran-2-yl)-2′-deoxyadenosine 184 (N6-THF-dAdo) and N4-(tetrahydrofuran-2-yl)-2′-deoxycytidine 186 (N4-THF-dCyd) were also identified in calf thymus DNA incubated with α-acetoxyNPYR. Two unstable Thd adducts, O2-(tetrahydrofuran-2-yl)thymidine 187 (O2-THF-Thd) and O4-(tetrahydrofuran-2-yl)thymidine 188 (O4-THF-Thd), were also formed and verified by their reduction products [255]. Some of the tetrahydrofuranyl DNA adducts were also observed in the reactions of oxidized THF with deoxyribonucleosides in vitro [256]. In a study in which F344 rats were gavaged with a single dose of NPYR and sacrificed after 16 h, O2-THF-Thd 187 (quantified in the reduced form 192) predominated among the tetrahydrofuranyl DNA adducts at all 3 doses (see Table 1) [247]. In the chronic rat study with doses of NPYR administered in drinking water at 600 ppm for 1 week or 200 ppm for 4 or 13 weeks, dGuo and Thd adducts persisted in much higher abundance than the dAdo adduct. However, they did not seem to accumulate over the study course [252].
DNA Adducts Formed by NPYR-Derived Crotonaldehyde
In the reactions of dGuo with two stable regiochemically activated compounds, α-acetoxyNPYR 165 and 4-(carbethoxynitrosamino)butanal 166 (Scheme 8), two diasteromeric DNA adducts, (6S,8S)- and (6R,8R)-N1,N2-propanodeoxyguanosine 194 and 195 (N1,N2-propano-dGuo, Figure 12), were the major products, identical to the products formed by crotonaldehyde 177 [257]. Those two adducts were also formed by the reaction of α-acetoxyNPYR with dGuo, DNA, or RNA in vitro [244,245,249]. In the hepatic DNA of rats treated with NPYR in drinking water for 14 days, N1,N2-propano-dGuo (194 and 195 together) occurred at the level of 0.06 μmol/mol Gua. This adduct was also formed in the skin DNA of mice topically treated with crotonaldehyde. However, the concentration of N1,N2-propano-dGuo (194 and 195 together) in the rat liver DNA was significantly lower (~10,000-fold) than N7,8-butano-Gua 179 [251]. The diastereomer (6S,8S)-N1,N2-propano-dGuo 194 was preferentially formed over its (6R,8R)-counterpart 195 by crotonaldehyde [233].
In addition to the methyl-substituted N1,N2-propano-dGuo adducts, two new exocyclic 7,8-guanine adducts, cis- and trans-2-amino-7,8-dihydro-8-hydroxy-6-methyl-3H-pyrrolo [2,1-f]purine-4(6H)-one 196 and 197 (N7,8-Cro-Gua, Figure 12), were also identified in vitro in calf thymus DNA treated with α-acetoxyNPYR or crotonaldehyde but not in vivo in rat liver DNA [244,245]. They are likely formed via initial Michael addition followed by cyclization, the same mechanism for the selective formation of N1,N2-propano-dGuo [258]. The same two diasteromeric adducts 196 and 197 were also detected in crotonaldehyde-treated DNA in vitro and in vivo [244,251].
Other DNA Adducts Related to NPYR Metabolism
As depicted in Scheme 8, the metabolite 3-hydroxybutanal 175 can dimerize to form paraldol 176, which has been detected in the hydrolysates of DNA treated with α-acetoxyNPYR. Paraldol was also released from crotonaldehyde-treated DNA after neutral thermal hydrolysis but in much higher abundance, suggesting the presence of the unstable paraldol-releasing adduct(s) formed by α-acetoxyNPYR is likely via the intermediate crotonaldehyde [235]. Two paraldol-dGuo adducts were characterized in crotonaldehyde-treated DNA. They are N2-(2-(2-hydroxypropyl)-6-methyl-1,3-dioxan-4-yl)-2′-deoxyguanosine 198 (N2-paraldol-dGuo, Figure 13) and N2-(2-(2-hydroxypropyl)-6-methyl-1,3-dioxan-4-yl)deoxyguanylyl-(5′–3′)-thymidine 199 (N2-paraldol-dGuo-(5′–3′)-Thd) [233]. The major paraldol-releasing DNA adduct was later characterized to be N2-(3-hydroxybutylidene)-2′-deoxyguanosine 200, a Schiff base that was unstable at the nucleoside level but appeared to be stable in DNA. The level of 200 exceeded the Michael addition products of tricyclic N1,N2-propano-dGuo adducts 194 and 195 in crotonaldehyde- and α-acetoxyNPYR-treated DNA [234,259].
In the reaction of α-acetoxyNPYR 165 with dGuo, N7-(N-nitrosopyrrolidin-2-yl)guanine 202 (Figure 13) was identified as the first example of a nitrosamine adduct retaining the N-nitroso moiety [259]. It was considered to be formed through the intermediacy of the nitrosiminium ion 201 [260]. A new cyclized adduct 2-(2-hydroxypyrrolidin-1-yl)-2′-deoxyinosine 203 (N2-Py(OH)-dI) was for the first time identified and confirmed by its synthetic standard in the reaction of α-acetoxyNPYR with both dGuo and DNA; it can be reduced to 2-(pyrrolidin-1-yl)-2′-deoxyinosine 204 (N2-Py-dI) [259].
4.1.4. Mutagenicity and Genotoxicity of NPYR-Derived DNA Adducts
NPYR is mutagenic towards E. coli and S. typhinurium after hepatic microsomal activation [261,262]. A few DNA adducts formed by NPYR metabolism such as N1,N2-propano-dGuo can also arise from crotonaldehyde. The mutagenicity and genotoxicity of these types of DNA adducts have been extensively reviewed before [263,264,265]. However, the mutagenicity and genotoxicity of the other types of NPYR DNA adducts, such as 184–186 and their reduced forms 189–193, still warrant investigation.
4.1.5. Human DNA Adducts Related to NPYR Metabolism
The N1,N2-propano-dGuo adducts have been detected in human tissues [266,267]. We have quantified (6S,8S)-N1,N2-propano-dGuo 194 and (6R,8R)-N1,N2-propano-dGuo 195 in human liver, lung, and white blood cells [268]. (6S,8S)-N1,N2-Propano-dGuo 194 occurred at a mean (± SD) concentration of 6.70 ± 2.92 and 7.19 ± 4.14 fmol/μmol dGuo, respectively, in liver and lung DNA. However, the detection rates were low; only 4 out of 23 liver samples and 16 out of 45 lung samples were positive. Similarly, (6R,8R)-N1,N2-propano-dGuo 195 occurred at mean (±SD) concentrations of 7.87 ± 4.47 and 12.8 ± 7.6 fmol/μmol dGuo, respectively, in the same liver and lung DNA. No statistically significant difference was observed between the occurrence of the two diastereomers. Neither of the two isomers was detected in the DNA of 11 human white blood cell samples [268]. However, considering the fact that the most abundant NPYR DNA adduct, N7,8-butano-Gua 179, has not been detected in the human tissues, the origin of N1,N2-propano-dGuo is likely not due to the exposure of NPYR but rather to other pathways such as the endogenous formation of crotonaldehyde from lipid peroxidation [269]. Presently, there is no direct evidence for the presence of NPYR-DNA adducts specifically in human tissues. However, the studies on NPYR-DNA damage did lead to research on crotonaldehyde-DNA interactions and their detection in human tissue samples as noted above.
4.2. N-Nitrosopiperidine (NPIP)
4.2.1. Exposure and Carcinogenicity
NPIP (9, Figure 1) has been reported in water, spices, and foods such as cheese, smoked fish, and processed meat [73,270]. The average concentration of NPIP was 0.5 ± 0.1 ng/g in food and 7.9 ± 4.0 ng/L in potable water [9]. The formation of NPIP by the nitrosation of amine precursors such as piperidine has been associated with concentrations of sodium nitrite during the production of dry fermented sausages [271]. The presence of NPIP in tobacco products has been documented but is currently at extremely low levels [73,77].
The IARC reviewed the carcinogenicity data of NPIP in 1978 and reaffirmed its Group 2B classification in 1987. It has shown carcinogenic effects in multiple laboratory animals such as mice, rats, hamsters, and monkeys [73]. In mice, NPIP induced primarily liver and lung tumors when administered in the diet and caused lung adenomas when administered in the drinking water or by i.p. injection. In rats, esophageal and hepatic tumors were induced by NPIP administered in drinking water. However, the target organ shifted to the nasal cavity when administered by s.c. injection. Carcinomas of the esophagus and pharynx were mainly observed by i.v. injection of NPIP [73]. The ability of NPIP to cause esophageal tumors is striking when compared to its close analog NPYR, which never causes esophageal tumors in rats. This was confirmed by Gray et al. in a large 2-year study. The incidence of hepatic and esophageal tumors in rats was dose-responsive to the concentration of NPIP in the drinking water [220]. In hamsters, the trachea appears to be the primary target organ by NPIP after s.c. injection. In monkeys, hepatocellular carcinomas were observed after oral dosing with NPIP [73].
4.2.2. Metabolism
NPIP is primarily biotransformed by hepatic P450s with the additional contribution of cytosolic proteins [272,273,274]. Catalyzed primarily by P450 2As [226,275,276], NPIP undergoes α-hydroxylation to form the unstable α-hydroxyNPIP 206 (Scheme 9). This intermediate has been trapped in the form of α-hydroxyNPIP phosphate ester 221 under near-ultraviolet irradiation conditions [277]. After the decomposition of α-hydroxyNPIP and the spontaneous loss of H2O and N2, the electrophilic intermediate carbocation 213 is formed. The carbocation 213 can form the oxonium ion 216 via intramolecular cyclization or the isomeric carbocation 211 via a 1,2-H shift. Their solvolysis products, 5-hydroxypentanal 214 (in equilibrium with 2-hydroxytetrahydropyran 217 (THP-OH)) [278,279,280] and the reduced form 1,5-pentanediol 215 [272] and 4-hydroxypentanal 212 (in equilibrium with 2-hydroxy-5-methyltetrahydrofuran 210) [278] or the potential elimination/oxidation product 4-oxopent-2-enal 209 (acetylacrolein) [278], have been identified in support of the proposed mechanism of NPIP α-hydroxylation. It is noteworthy that 4-oxopent-2-enal 209 may also result from the metabolism of 2-methylfuran [281].
Beta- and γ-hydroxylation of NPIP have also been suggested as potential metabolic pathways (Scheme 9). N-Nitroso-4-hydroxypiperidine 219 and N-nitroso-4-piperidone 220 were detected in vitro in early studies with rat liver microsomes [282,283]. N-Nitroso-3-hydroxypiperidine 218 and N-nitroso-4-hydroxypiperidine 219 were minor products compared to 5-hydroxypentanal 214 in studies with guinea pig liver microsomes [280].
The metabolic bioactivation pathways illustrated in Scheme 9 were also supported by the structure-carcinogenicity studies. Methyl substitution at the α-carbons of NPIP significantly decreased tumor formation in rats administered NPIP or its methyl analogs in drinking water [284]. The β- or γ-substitutions (methyl and hydroxy) did not affect the carcinogenicity of NPIP in rats [284,285].
4.2.3. DNA Adducts Formed by NPIP Metabolism
In the reaction mixture of α-acetoxy-N-nitrosopiperidine 205 (α-acetoxyNPIP, Scheme 9) with dGuo, a peak corresponding to 7-(2-oxopropyl)-5,9-dihydro-9-oxo-3-β-D-deoxyribofuranosylimidazo [1,2-a]purine 222 (7-(2-oxopropyl)-N1,N2-etheno-dGuo, Figure 14) was observed. Its formation was proposed to occur via the intermediate 4-oxopent-2-enal 209 [254,278]. The unstable hemiaminal precursor for 7-(2-oxopropyl)-NN1,N2-etheno-dGuo has been identified in vitro, as such or in its reduced form. It was characterized as 7-(2-oxopropyl)-5-hydroxy-5,6,7,9-tetrahydro-9-oxo-3-β-D-deoxyribofuranosylimidazo [1,2-a]purine 223 [286].
A pair of two diastereomeric N2-(3,4,5,6-tetrahydro-2H-pyran-2-yl)-2′-deoxyguanosine 224 (N2-THP-dGuo, Figure 14) were identified as the major products in the same reaction mixture of dGuo compared to 7-(2-oxopropyl)-N1,N2-etheno-dGuo 222 [254]. Similarly, in the reaction mixture of α-acetoxyNPIP 205 with calf thymus DNA, N2-THP-dGuo was clearly formed, whereas 7-(2-oxopropyl)-N1,N2-etheno-dGuo was minimally observable. THP-OH 217 (Scheme 9) was released in the DNA hydrolysate after neutral thermal hydrolysis, probably resulting from N2-THP-dGuo [253].
4.2.4. Human DNA Adducts Related to NPIP Metabolism
In 2019, Totsuka et al. reported their DNA adductome investigation of esophageal cancer patients in China [287]. They found that a distinctive pattern of NPIP-derived DNA adduct N2-THP-dGuo 224 (Figure 14) formed in the tissues of esophageal cancer patients in the high-incidence area versus those in the low-incidence area. The difference in the occurrence of this adduct was also statistically significant (p < 0.01) in the peripheral blood samples of patients from the two areas. The exposure to NPIP was not likely due to smoking or drinking since all the samples were from patients who were nonsmokers and nondrinkers. Drinking water and vegetables were potential sources that have been preliminarily investigated [287].
NPIP induced preferentially AT to CG transversions in gpt delta transgenic rats. However, the predominant somatic mutation of esophageal tumor samples was CG to TA transitions; no distinctive mutational pattern was identified to associate with esophageal cancer in the high-incidence area. In the p53 genes of ~90% of patients, GC to AT transitions predominated, in agreement with the findings from their previous study in 2005 [288]. There was only a weak correlation observed between one of the mutational signatures (signature 17 in the COSMIC database) and the N2-THP-dGuo levels among 19 subjects (r = 0.44; p < 0.05). Taken together, this study suggests that NPIP-induced DNA adducts can at least partially contribute to esophageal carcinogenesis [287].
4.3. N-Nitrosomorpholine (NMOR)
4.3.1. Exposure and Carcinogenicity
NMOR (10, Figure 1) has been detected in water, food, cosmetics, and occupational airspaces [289,290,291,292,293,294,295]. It has also been documented to occur in smokeless tobacco products but at very low concentrations [77]. Similar to other N-nitrosamines, NMOR can be formed easily by nitrosation of its parent amine morpholine with NaNO2. It is one of the most rapidly formed nitrosamine products [296]. When morpholine was administered in combination with nitrite to rats by gavage, the formation of NMOR was readily measured in the urine, with the extent of morpholine nitrosation being 0.5–12% depending on the doses [297]. Mice exposed to 15NO2 and administered morpholine by gavage also formed NMOR with the highest concentrations detected in the skin followed by the stomach [298]. Human exposure to morpholine and its analogs has been considered substantial from food and drugs [270,299,300], tobacco usage [301], and rubber and tire manufacturing [302,303]. Thus, the in vivo formation of NMOR has been considered likely to occur in humans. Low levels of NMOR have been detected in human urine and gastric juice [304,305,306].
The carcinogenicity data of NMOR was reviewed by IARC in 1978 and 1987, and it was classified as a group 2B carcinogen. NMOR primarily induces liver, bile duct, and kidney tumors in mice and rats when given in the drinking water or by i.v. injection; it causes tumors of the respiratory system (trachea and nasal cavity) of hamsters by s.c. injection [73]. After 1978, there were some new carcinogenicity data reported. When NMOR was administered in the drinking water to A/J mice, lung tumors occurred at a relatively low total dose of 53–55 μmol/mouse. It induced 100% liver tumor incidence in F344 rats at a high total dose of 1.1 mmol/rat [190]. It is interesting to note that 2-/6-methylation of NMOR shifts its organospecificity. In Syrian golden hamsters treated by gavage, NMOR induced primarily nasal cavity tumors whereas N-nitroso-2-methylmorpholine induced tumors in the nasal cavity and liver [307]; N-nitroso-2,6-dimethylmorpholine caused liver and pancreas tumors [308]. In rats, NMOR is a potent liver carcinogen whereas N-nitroso-2,6-dimethylmorpholine is carcinogenic to the esophagus and nasal cavity [7,309].
4.3.2. Metabolism
After a single i.p. injection, NMOR was rapidly distributed throughout the rat tissues and metabolized to a relatively high extent with 24% of the dose excreted unchanged in the urine after 18 h [310]. It is metabolized via both α- and β-hydroxylation pathways (Scheme 10).
Hydroxylation on the 2-carbons of NMOR appears to be the major biotransformation pathway, since NHEG 135, derived from the β-hydroxylation product NHMOR 137 via the intermediate 136, has been detected predominantly among the rat urinary metabolites, accounting for 33% or 37% of total metabolites [311]. It has been used as a reliable biomarker to monitor the in vivo formation of NMOR [297]. However, NHEG can also be formed by NDELA and N,N-dinitrosopiperazine, although to the lesser extent of <10% and 22% from each compound, respectively [184,312]. It is noteworthy that the group 2B carcinogen NDELA 6 is also formed by the reduction of intermediate 136 during NMOR metabolism, accounting for 12% of total urinary metabolites [311,313]. NDELA can be reversibly oxidized to form the same products 135, 136, and 137 (see Scheme 6) as in the case of NMOR.
When the hydroxylation occurs on the 3-carbon of NMOR, the unstable product N-nitroso-3-hydroxymorpholine 228 (α-hydroxyNMOR) is formed. By analogy to NPIP metabolism, a potential α-hydroxyNMOR acetate ester 234 has been suggested to be similarly formed under near-ultraviolet light irradiation conditions [314]. α-HydroxyNMOR decomposes quickly to form the diazohydroxide 229, which can also result from the regiochemically activated precursors α-acetoxy-N-nitrosomorpholine 225, N-nitrosomorpholine hydroperoxide 226, and (2-(carbethoxynitrosamino)ethoxy)ethanal 227 [315,316,317]. The diazohydroxide spontaneously loses one molecule of H2O and N2 and forms the carbocation 231 and its isomeric forms, 232 and 233. The carbocation 231 forms the solvolysis product, 2-(2-hydroxyethoxy)acetaldehyde 235, in equilibrium with 2-hydroxy-1,4-dioxane 236; the carbocation 232 forms acetaldehyde 42 and glycolaldehyde 130 after solvolysis; the other carbocation 233 forms glyoxal 133 [311,318]. The major ultimate product arising from NMOR α-hydroxylation was (2-hydroxyethoxy)acetic acid 237 in rat urine. A significant isotope effect was observed in the formation of 237 when the α-carbons of NMOR were deuterated (16% vs. 3.4%) [311]. It has been shown that 1,4-dioxan-2-one 238 exists in aqueous solutions in equilibrium with 237 with an equilibrium constant KOA of 0.034 ± 0.002 M. It appears to be a common non-carcinogenic metabolite of the carcinogens NMOR, dioxane, and diethylene glycol [319].
The β-hydroxylation product of NMOR—NHMOR 137 (Scheme 10)—is highly mutagenic [189,320,321] and forms DNA adducts in vitro [315]. It is, however, non-tumorigenic or marginally tumorigenic to laboratory animals [190]. To explain the apparent lack of carcinogenicity of NHMOR, Loeppky et al. investigated the metabolic profiles of two NHMOR α-hydroxylation metabolites—N-nitroso-2,3-dihydroxymorpholine 139 and N-nitroso-2,5-dihydroxymorpholine 142—using their stable precursors, N-nitroso-3-acetoxy-2-hydroxymorpholine 138 and N-nitroso-5-acetoxy-2-hydroxymorpholine 141. The hydrolytic decomposition products of 139 were glyoxal 133 (95%), ethylene glycol 129 (55%), acetaldehyde 42 (10%), and acetic acid; the hydrolytic decomposition products of 142 were 2-acetoxyacetaldehyde 144 (65%), glycol aldehyde 130 (15%), glyoxal 133 (trace), and acetic acid. The high yield of 2-acetoxyacetaldehyde 144 may be responsible for the low carcinogenicity of NHMOR if it is primarily hydroxylated at the 2-carbons [195].
4.3.3. DNA Adducts Formed by NMOR Metabolism
A study by Stewart et al. in 1974 using [14C]NMOR suggested that six radioactive DNA adducts are formed by NMOR metabolism. One of those compounds was putatively identified to be N7-HOEt-Gua 69 (Figure 5) [310]. Fishbein et al. incubated nucleosides and calf thymus DNA with NMOR hydroperoxide 226 (Scheme 10) and identified a panel of 2-ethoxyacetaldehyde purine adducts as shown in Figure 15. These include N3-, N6-, and N7-(2-oxoethoxyethyl)adenine (245, 246, and 248) and N1-, N2-, O6-, and N7-(2-oxoethoxyethyl)guanine (249, 250, 252 and 253), among which N7-(2-oxoethoxyethyl)guanine 253 and O6-(2-oxoethoxyethyl)guanine 252 are the two most abundant and N3-(2-oxoethoxyethyl)adenine 245 is the third, however, in much lower concentrations. Those 2-oxoethoxyethyl adducts slowly decayed to the corresponding hydroxyethyl adducts (62–67 and 69, Figure 5) with a faster rate in duplex DNA than in nucleosides. Interestingly, the two 2-oxoethoxyethyl adducts 246 and 250, occurring at the exocyclic amino groups, also formed the corresponding intramolecular ring closure products 247 and 251, respectively, during chemical standard synthesis. However, 247 and 251 were not detected in the treated nucleosides or calf thymus DNA [317]. The same group also identified the first cross-link DNA adduct 6-(2-(2-((9H-purin-6-yl)amino)ethoxy)ethoxy)-9H-purin-2-amine 254 (Figure 15) derived from α-hydroxyNMOR upon reduction. It was proposed to be formed by the carbocation 231 (Scheme 10) attacking the O6-OH of guanine first, followed by the pendant aldehyde reacting with N6-NH2 of adenine [322].
We conducted an in vitro study of [2-(carbethoxynitrosamino)ethoxy]ethanal 227 (Scheme 10) reacting with dGuo and demonstrated the formation of N1,N2-glyoxal-dGuo 145 (Figure 11) probably via glyoxal resulting from NMOR α-hydroxylation [309]. In the reaction mixture of NHMOR with dGuo, adduct 145 was also formed. The proposed mechanism of the formation of this adduct by NHMOR is depicted in Scheme 11 [315].
Consistent with their metabolic profiles, the two precursors of α-hydroxyNHMOR 138 and 141 (Scheme 10) showed different reactivity toward dGuo. In the in vitro incubation mixture with dGuo, compound 141 was less reactive compared to 138. The dGuo adduct formed by 141 was N1,N2-etheno-dGuo 146 (Figure 11). The dGuo adduct primarily formed by 138 was N1,N2-glyoxal-dGuo 145 (Figure 11) along with two minor adducts, N7-HOEt-Gua 69 and O6-HOEt-Gua 67 (Figure 5) [195].
5. Concluding Remarks
The structural simplicity of the powerful carcinogen NDMA provided a unique opportunity to understand the role of metabolism and DNA adduct formation in carcinogenesis. This research has clearly demonstrated the main requisite steps in tumor formation by N-nitrosamines and other related organic carcinogens: formation of electrophilic intermediates catalyzed by cytochrome P450 enzymes, binding of those intermediates to sites in DNA, the role of DNA repair systems, mutagenesis resulting from certain unrepaired DNA adducts such as O6-Me-Gua, and consequent permanent mutations in critical growth control genes. This sequence of events is also quite well-established for NDEA and NMEA and forms the basis for the investigation of more structurally complex N-nitrosamines as described here. For these other N-nitrosamines, there are still gaps in our understanding of their metabolic activation pathways and the DNA adduct structures, repair mechanisms, and mutagenic properties of the persistent adducts. This review has presented an overview of the current status of this research.
Studies of the metabolic activation and DNA adduct formation of carcinogenic N-nitrosamines have long been part of the critical mass of research on these compounds. The focus of many cancer researchers on N-nitrosamines produced an overall heightened awareness of their ease of formation, levels of contamination in consumer products, and carcinogenic effects among other topics. This heightened awareness led to significant decreases in human N-nitrosamine exposures from the 1970s to the present. However, constant vigilance is necessary to maintain these lower exposure levels. The recent manufacturing errors leading to N-nitrosamine contamination of drugs would have been unimaginable in the 1980s and 1990s when the ease of N-nitrosamine formation under certain conditions was prominent in the eyes of chemists and toxicologists.
There are approximately 200 different N-nitrosamines that have been documented to be carcinogenic in more than 30 animal species [323]. Humans exposed to these compounds are, no doubt, susceptible to their carcinogenic effects. The continuous evaluation of potential N-nitrosamine contamination such as N-nitrosomethyl-n-alkylamines in medicines is in progress by other groups [324]. Among those many N-nitrosamines, the compounds listed in Figure 1 comprise the most commonly detected carcinogenic N-nitrosamines from human daily exposure, occurring in food, water, drugs, and cosmetics. An understanding of the metabolism of these carcinogens and their mechanisms of DNA interactions is necessary basic knowledge for evaluating and controlling their potential carcinogenic effects in humans, and some metabolites and/or DNA adducts that are specific to carcinogen exposure in humans may serve as biomarkers for cancer etiology studies.
With advances in bioanalytical methods, especially in the development of mass spectrometry, metabolism studies have evolved from radioisotope-labeling technology to high-resolution mass spectrometry. Furthermore, mass spectrometry-based metabolomics studies may provide rich chemical information on structures of potential metabolites of interest [325]. Similarly, DNA adductomics [326] is also emerging as a useful tool to investigate DNA adducts that are potential biomarkers for cancer etiology studies, as illustrated in the NPIP section [287].
In summary, the recent occurrence of NDMA and NDEA in some batches of FDA-approved drugs has heightened global awareness of the carcinogenic effects of N-nitrosamines, which comprise a significant number of carcinogens to which humans are exposed on a daily basis through food, water, drugs, cosmetics, and tobacco products. In this review, we provide a comprehensive and updated review of 10 N-nitrosamine carcinogens with a focus on their mechanisms of metabolic activation and DNA interactions. A better understanding of the metabolism and DNA adduct formation of N-nitrosamines can hopefully provide clues for relevant cancer etiology and prevention studies.
Author Contributions
Conceptualization, Y.L. and S.S.H.; writing—original draft preparation, Y.L.; writing—review and editing, Y.L. and S.S.H. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support for tobacco-specific N-nitrosamine studies in our laboratory was provided by the U.S. National Cancer Institute through grant CA-81301. Mass spectrometry was carried out in the Analytical Biochemistry Shared Resource of the Masonic Cancer Center, University of Minnesota, funded in part by Cancer Center Support grant CA-77598.
Acknowledgments
Editorial assistance from Robert (Bob) Carlson is greatly appreciated.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| α-AcetoxyNPYR | α-acetoxy-N-nitrosopyrrolidine |
| AGT | O6-alkylguanine-DNA alkyltransferease |
| APNG | N-(N′-acetyl-L-prolyl)-N-nitrosoglycine |
| AS | azaserine |
| BBN | N-nitroso-(4-hydroxybutyl)butylamine |
| E. coli | Escherichia coli |
| FDA | Food and Drug Administration |
| GSH | glutathione |
| α-hydroxyNMOR | N-nitroso-3-hydroxymorpholine |
| IARC | International Agency for Research on Cancer |
| i.p. | intraperitoneal |
| i.v. | intravenous |
| KDA | potassium diazoacetate |
| MGMT | O6-methylguanine-DNA methyltransferase |
| MNPA | 3-(methylnitrosamino)propionic acid |
| N2-THF-dGuo | N2-(tetrahydrofuran-2-yl)-2′-deoxyguanosine |
| N2-(4-HOB)-dGuo | N2-(4-hydroxybutyl)-2′-deoxyguanosine |
| N7-CE-Gua | N7-(2′-carboxyethyl)guanine |
| N7-CM-Gua | N7-(carboxymethyl)guanine |
| N7-Et-Gua | N7-ethylguanine |
| N7-Me-Gua | N7-methylguanine |
| N7-n-Pr-Gua | N7-(n-propyl)guanine |
| N7,8-butano-Gua | 2-amino-6,7,8,9-tetrahydro-9-hydroxypyrido [2,1-f]purine-4(3H)-one |
| N7,8-Cro-Gua | 2-amino-7,8-dihydro-8-hydroxy-6-methyl-3H-pyrrolo [2,1-f]purine-4(6H)-one |
| NDBA | N-nitrosodi-n-butylamine |
| NDEA | N-nitrosodiethylamine |
| NDELA | N-nitrosodiethanolamine |
| NDIPA | N-nitrodiisopropylamine |
| NDMA | N-nitrosodimethylamine |
| NDPA | N-nitrosodi-n-propylamine |
| NER | nucleotide excision repair |
| NHEG | N-nitroso-(2-hydroxyethyl)glycine |
| NHMOR | N-Nitroso-2-hydroxymorpholine |
| NHPPA | N-nitroso-2-hydroxypropylpropylamine |
| NIPEA | N-nitrosoisopropylethylamine |
| NMBA | N-nitroso-N-methyl-4-aminobutanoic acid |
| NMEA | N-nitrosomethylethylamine |
| NMOR | N-nitrosomorpholine |
| NMPA | N-nitrosomethylphenylamine |
| NOPPA | N-nitroso-2-oxopropylpropylamine |
| NPIP | N-nitrosopiperidine |
| NPYR | N-nitrosopyrrolidine |
| NSAR | N-nitrososarcosine |
| 7-(2-oxopropyl)-N1,N2-etheno-dGuo | 7-(2-oxopropyl)-5,9-dihydro-9-oxo-3-β-D-deoxyribofuranosylimidazo [1,2-a]purine |
| O2-Me-Thd | O2-methylthymidine |
| O4-Me-Thd | O4-methylthymidine |
| O6-n-Bu-Gua | O6-(n-butyl)guanine |
| O6-CM-Gua | O6-(carboxymethyl)guanine |
| O6-Et-Gua | O6-ethylguanine |
| O6-(4-OH-n-Bu)-Gua | O6-(4-hydroxybutyl)guanine |
| O6-HOEt-dGuo | O6-(2-hydroxyethyl)-2′-deoxyguanosine |
| O6-Me-Gua | O6-methylguanine |
| pol κ | polymerase κ |
| ppm | parts per million |
| s.c. | subcutaneous |
| S. typhinurium | Salmonella typhinurium |
| THP-OH | 2-hydroxytetrahydropyran |
References
- Magee, P.N.; Barnes, J.M. The production of malignant primary hepatic tumours in the rat by feeding dimethylnitrosamine. Br. J. Cancer 1956, 10, 114–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakshaug, J.; Sognen, E.; Hansen, M.A.; Koppang, N. Dimethylnitrosamine; its hepatotoxic effect in sheep and its occurrence in toxic batches of herring meal. Nature 1965, 206, 1261–1262. [Google Scholar] [CrossRef] [PubMed]
- Ender, F.; Ceh, L. Occurrence of nitrosamines in foodstuffs for human and animal consumption. Food Cosmet. Toxicol. 1968, 6, 569–571. [Google Scholar] [CrossRef]
- Sen, N.P. The evidence for the presence of dimethylnitrosamine in meat products. Food Cosmet. Toxicol. 1972, 10, 219–223. [Google Scholar] [CrossRef]
- Lijinsky, W. Chemistry and Biology of N-Nitroso Compounds; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Druckrey, H.; Preussmann, R.; Ivankovic, S.; Schmähl, D. Organotropic carcinogenic effects of 65 various N-nitroso- compounds on BD rats. Z. Krebsforsch. 1967, 69, 103–201. [Google Scholar] [CrossRef]
- Preussmann, R.; Stewart, B.W. N-Nitroso Carcinogens. In Chemical Carcinogens, 2nd ed.; ACS Monograph 182; Searle, C.E., Ed.; American Chemical Society: Washington, DC, USA, 1984; Volume 2, pp. 643–828. [Google Scholar]
- Bogovski, P.; Bogovski, S. Animal species in which N-nitroso compounds induce cancer. Int. J. Cancer 1981, 27, 471–474. [Google Scholar] [CrossRef]
- Gushgari, A.J.; Halden, R.U. Critical review of major sources of human exposure to N-nitrosamines. Chemosphere 2018, 210, 1124–1136. [Google Scholar] [CrossRef]
- Kroes, R.; Kleiner, J.; Renwick, A. The threshold of toxicological concern concept in risk assessment. Toxicol. Sci. 2005, 86, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Tricker, A.R. N-nitroso compounds and man: Sources of exposure, endogenous formation and occurrence in body fluids. Eur. J. Cancer Prev. 1997, 6, 226–268. [Google Scholar] [CrossRef]
- Park, J.E.; Seo, J.E.; Lee, J.Y.; Kwon, H. Distribution of seven N-nitrosamines in food. Toxicol. Res. 2015, 31, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Mitch, W.A.; Sharp, J.O.; Trussell, R.R.; Valentine, R.L.; Alvarez-Cohen, L.; Sedlak, D.L. N-nitrosodimethylamine (NDMA) as a drinking water contaminant: A review. Environ. Eng. Sci. 2003, 20, 389–404. [Google Scholar] [CrossRef] [Green Version]
- Beard, J.C.; Swager, T.M. An organic chemist’s guide to N-nitrosamines: Their structure, reactivity, and role as contaminants. J. Org. Chem. 2021, 86, 2037–2057. [Google Scholar] [CrossRef] [PubMed]
- Bharate, S.S. Critical analysis of drug product recalls due to nitrosamine impurities. J. Med. Chem. 2021, 64, 2923–2936. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Drug Recalls. Available online: https://www.fda.gov/drugs/drug-safety-and-availability/drug-recalls (accessed on 21 September 2021).
- U.S. Food and Drug Administration. Control of Nitrosamine Impurities in Human Drugs: Guidance for Industry; Center for Drug Evaluation and Research: Silver Spring, MD, USA, 2021.
- Lim, D.S.; Roh, T.H.; Kim, M.K.; Kwon, Y.C.; Choi, S.M.; Kwack, S.J.; Kim, K.B.; Yoon, S.; Kim, H.S.; Lee, B.M. Risk assessment of N-nitrosodiethylamine (NDEA) and N-nitrosodiethanolamine (NDELA) in cosmetics. J. Toxicol. Environ. Health A 2018, 81, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Preussmann, R.; Spiegelhalder, B.; Eisenbrand, G.; Wurtele, G.; Hofmann, I. Urinary excretion of N-nitrosodiethanolamine in rats following its epicutaneous and intratracheal administration and its formation in vivo following skin application of diethanolamine. Cancer Lett. 1981, 13, 227–231. [Google Scholar] [CrossRef]
- American Chemical Society. Molecule of the Week Archive: N-Nitrosodimethylamine. ACS Molecule of the Week, 14 January 2019. [Google Scholar]
- Peto, R.; Gray, R.; Brantom, P.; Grasso, P. Effects on 4080 rats of chronic ingestion of N-nitrosodiethylamine or N-nitrosodimethylamine: A detailed dose-response study. Cancer Res. 1991, 51 Pt 2, 6415–6451. [Google Scholar]
- Peto, R.; Gray, R.; Brantom, P.; Grasso, P. Dose and time relationships for tumor induction in the liver and esophagus of 4080 inbred rats by chronic ingestion of N-nitrosodiethylamine or N-nitrosodimethylamine. Cancer Res. 1991, 51 Pt 2, 6452–6469. [Google Scholar]
- National Toxicology Program. Report on Carcinogens, 15th ed.; U.S. Department of Health and Human Services, Public Health Service: Research Triangle Park, NC, USA, 2021.
- Yamazaki, H.; Inui, Y.; Yun, C.H.; Guengerich, F.P.; Shimada, T. Cytochrome P450 2E1 and 2A6 enzymes as major catalysts for metabolic activation of N-nitrosodialkylamines and tobacco-related nitrosamines in human liver microsomes. Carcinogenesis 1992, 13, 1789–1794. [Google Scholar] [CrossRef]
- Mochizuki, M.; Osabe, M.; Anjo, T.; Suzuki, E.; Okada, M. Mutagenicity of α-hydroxy N-nitrosamines in V79 Chinese hamster cells. J. Cancer Res. Clin. Oncol. 1984, 108, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, G.; Calcutt, M.W.; Guengerich, F.P. Oxidation of N-nitrosoalkylamines by human cytochrome P450 2A6: Sequential oxidation to aldehydes and carboxylic acids and analysis of reaction steps. J. Biol. Chem. 2010, 285, 8031–8044. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, G.; Calcutt, M.W.; Nagy, L.D.; Guengerich, F.P. Oxidation of methyl and ethyl nitrosamines by cytochrome P450 2E1 and 2B1. Biochemistry 2012, 51, 9995–10007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godoy, H.M.; Diaz Gomez, M.I.; Castro, J.A. Mechanism of dimethylnitrosamine metabolism and activation in rats. J. Natl. Cancer Inst. 1978, 61, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Lake, B.G.; Minski, M.J.; Phillips, J.C.; Gangolli, S.D.; Lloyd, A.G. Investigations into the hepatic metabolism of dimethylnitrosamine in the rat. Life Sci. 1975, 17, 1599–1606. [Google Scholar] [CrossRef]
- Kroeger-Koepke, M.B.; Koepke, S.R.; McClusky, G.A.; Magee, P.N.; Michejda, C.J. α-Hydroxylation pathway in the in vitro metabolism of carcinogenic nitrosamines: N-nitrosodimethylamine and N-nitroso-N-methylaniline. Proc. Natl. Acad. Sci. USA 1981, 78, 6489–6493. [Google Scholar] [CrossRef] [Green Version]
- Cottrell, R.C.; Lake, B.G.; Phillips, J.C.; Gangolli, S.D. The hepatic metabolism of 15N labelled dimethylnitrosamine in the rat. Biochem. Pharmacol. 1977, 26, 809–813. [Google Scholar] [CrossRef]
- Koepke, S.R.; Tondeur, Y.; Farrelly, J.G.; Stewart, M.L.; Michejda, C.J.; Kroeger-Koepke, M.B. Metabolism of 15N-labelled N-nitrosodimethylamine and N-nitroso-N-methylaniline by isolated rat hepatocytes. Biochem. Pharmacol. 1984, 33, 1509–1513. [Google Scholar] [CrossRef]
- Swann, P.F.; Mace, R.; Angeles, R.M.; Keefer, L.K. Deuterium isotope effect on metabolism of N-nitrosodimethylamine in vivo in rat. Carcinogenesis 1983, 4, 821–825. [Google Scholar] [CrossRef]
- Umbenhauer, D.R.; Pegg, A.E. Alkylation of intracellular and extracellular DNA by dimethylnitrosamine following activation by isolated rat hepatocytes. Cancer Res. 1981, 41 Pt 1, 3471–3474. [Google Scholar]
- Yoo, J.S.; Ishizaki, H.; Yang, C.S. Roles of cytochrome P450IIE1 in the dealkylation and denitrosation of N-nitrosodimethylamine and N-nitrosodiethylamine in rat liver microsomes. Carcinogenesis 1990, 11, 2239–2243. [Google Scholar] [CrossRef]
- Lorr, N.A.; Tu, Y.Y.; Yang, C.S. The nature of nitrosamine denitrosation by rat liver microsomes. Carcinogenesis 1982, 3, 1039–1043. [Google Scholar] [CrossRef]
- Lee, V.M.; Keefer, L.K.; Archer, M.C. An evaluation of the roles of metabolic denitrosation and alpha-hydroxylation in the hepatotoxicity of N-Nitrosodimethylamine. Chem. Res. Toxicol. 1996, 9, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Negishi, T.; Shiotani, T.; Fujikawa, K.; Hayatsu, H. The genotoxicities of N-nitrosamines in Drosophila melanogaster in vivo: The correlation of mutagenicity in the wing spot test with the DNA damages detected by the DNA-repair test. Mutat. Res. 1991, 252, 119–128. [Google Scholar] [CrossRef]
- Singer, B. Alkylation of the O6 of guanine is only one of many chemical events that may initiate carcinogenesis. Cancer Investig. 1984, 2, 233–238. [Google Scholar] [CrossRef]
- Li, Y.; Hecht, S.S. Metabolism and DNA adduct formation of tobacco-specific N-nitrosamines. Int. J. Mol. Sci. 2022, Manuscript submitted.
- Den Engelse, L.; Menkveld, G.J.; De Brij, R.J.; Tates, A.D. Formation and stability of alkylated pyrimidines and purines (including imidazole ring-opened 7-alkylguanine) and alkylphosphotriesters in liver DNA of adult rats treated with ethylnitrosourea or dimethylnitrosamine. Carcinogenesis 1986, 7, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Margison, G.P.; Margison, J.M.; Montesano, R. Methylated purines in the deoxyribonucleic acid of various Syrian-golden-hamster tissues after administration of a hepatocarcinogenic dose of dimethylnitrosamine. Biochem. J. 1976, 157, 627–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margison, G.P.; Margison, J.M.; Montesano, R. Accumulation of O6-methylguanine in non-target-tissue deoxyribonucleic acid during chronic administration of dimethylnitrosamine. Biochem. J. 1977, 165, 463–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, S.S.; Trushin, N.; Castonguay, A.; Rivenson, A. Comparative tumorigenicity and DNA methylation in F344 rats by 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone and N-nitrosodimethylamine. Cancer Res. 1986, 46, 498–502. [Google Scholar]
- Montesano, R.; Brésil, H.; Margison, G.P. Increased excision of O6-methylguanine from rat liver DNA after chronic administration of dimethylnitrosamine. Cancer Res. 1979, 39, 1798–1802. [Google Scholar]
- Souliotis, V.L.; Chhabra, S.; Anderson, L.M.; Kyrtopoulos, S.A. Dosimetry of O6-methylguanine in rat DNA after low-dose, chronic exposure to N-nitrosodimethylamine (NDMA). Implications for the mechanism of NDMA hepatocarcinogenesis. Carcinogenesis 1995, 16, 2381–2387. [Google Scholar] [CrossRef]
- Montesano, R.; Brésil, H.; Planche-Martel, G.; Margison, G.P.; Pegg, A.E. Stability and capacity of dimethylnitrosamine-induced O6-methylguanine repair system in rat liver. Cancer Res. 1983, 43 Pt 1, 5808–5814. [Google Scholar]
- Hall, J.; Brésil, H.; Serres, M.; Martel-Planche, G.; Wild, C.P.; Montesano, R. Modulation of O6-methylguanine-DNA methyltransferase in rat and hamster liver after treatment with dimethylnitrosamine. Cancer Res. 1990, 50, 5426–5430. [Google Scholar] [PubMed]
- Lindamood, C., 3rd; Bedell, M.A.; Billings, K.C.; Dyroff, M.C.; Swenberg, J.A. Dose response for DNA alkylation, [3H]thymidine uptake into DNA, and O6-methylguanine-DNA methyltransferase activity in hepatocytes of rats and mice continuously exposed to dimethylnitrosamine. Cancer Res. 1984, 44, 196–200. [Google Scholar] [PubMed]
- Anderson, L.M.; Harrington, G.W.; Pylypiw, H.M., Jr.; Hagiwara, A.; Magee, P.N. Tissue levels and biological effects of N-nitrosodimethylamine in mice during chronic low or high dose exposure with or without ethanol. Drug Metab. Dispos. 1986, 14, 733–739. [Google Scholar] [PubMed]
- Chhabra, S.K.; Anderson, L.M.; Perella, C.; Desai, D.; Amin, S.; Kyrtopoulos, S.A.; Souliotis, V.L. Coexposure to ethanol with N-nitrosodimethylamine or 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone during lactation of rats: Marked increase in O6-methylguanine-DNA adducts in maternal mammary gland and in suckling lung and kidney. Toxicol. Appl. Pharmacol. 2000, 169, 191–200. [Google Scholar] [CrossRef]
- Belinsky, S.A.; Devereux, T.R.; Anderson, M.W. Role of DNA methylation in the activation of proto-oncogenes and the induction of pulmonary neoplasia by nitrosamines. Mutat. Res. 1990, 233, 105–116. [Google Scholar] [CrossRef]
- Nicoll, J.W.; Swann, P.F.; Pegg, A.E. Effect of dimethylnitrosamine on persistence of methylated guanines in rat liver and kidney DNA. Nature 1975, 254, 261–262. [Google Scholar] [CrossRef]
- Pegg, A.E.; Hui, G. Formation and subsequent removal of O6-methylguanine from deoxyribonucleic acid in rat liver and kidney after small doses of dimethylnitrosamine. Biochem. J. 1978, 173, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Margison, G.P.; Kleihues, P. Chemical carcinogenesis in the nervous system. Preferential accumulation of O6-methylguanine in rat brain deoxyribonucleic acid during repetitive administration of N-methyl-N-nitrosourea. Biochem. J. 1975, 148, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Loechler, E.L.; Green, C.L.; Essigmann, J.M. In vivo mutagenesis by O6-methylguanine built into a unique site in a viral genome. Proc. Natl. Acad. Sci. USA 1984, 81, 6271–6275. [Google Scholar] [CrossRef] [Green Version]
- Delaney, J.C.; Essigmann, J.M. Biological properties of single chemical-DNA adducts: A twenty year perspective. Chem. Res. Toxicol. 2008, 21, 232–252. [Google Scholar] [CrossRef] [Green Version]
- Pegg, A.E. Multifaceted roles of alkyltransferase and related proteins in DNA repair, DNA damage, resistance to chemotherapy, and research tools. Chem. Res. Toxicol. 2011, 24, 618–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samson, L.; Han, S.; Marquis, J.C.; Rasmussen, L.J. Mammalian DNA repair methyltransferases shield O4MeT from nucleotide excision repair. Carcinogenesis 1997, 18, 919–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, N.; Wang, J.; Wang, P.; Jiang, Y.; Wang, Y. In-vitro replication studies on O2-methylthymidine and O4-methylthymidine. Chem. Res. Toxicol. 2012, 25, 2523–2531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, B.; Sági, J.; Kuśmierek, J.T. Escherichia coli polymerase I can use O2-methyldeoxythymidine or O4-methyldeoxythymidine in place of deoxythymidine in primed poly(dA-dT).poly(dA-dT) synthesis. Proc. Natl. Acad. Sci. USA 1983, 80, 4884–4888. [Google Scholar] [CrossRef] [Green Version]
- Christov, P.P.; Yamanaka, K.; Choi, J.Y.; Takata, K.; Wood, R.D.; Guengerich, F.P.; Lloyd, R.S.; Rizzo, C.J. Replication of the 2,6-diamino-4-hydroxy-N5-(methyl)-formamidopyrimidine (MeFapy-dGuo) adduct by eukaryotic DNA polymerases. Chem. Res. Toxicol. 2012, 25, 1652–1661. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wang, P.; Wang, Y. Cytotoxic and mutagenic properties of alkyl phosphotriester lesions in Escherichia coli cells. Nucleic Acids Res. 2018, 46, 4013–4021. [Google Scholar] [CrossRef] [Green Version]
- Harris, C.C.; Autrup, H.; Stoner, G.D.; Trump, B.F.; Hillman, E.; Schafer, P.W.; Jeffrey, A.M. Metabolism of benzo(a)pyrene, N-nitrosodimethylamine, and N-nitrosopyrrolidine and identification of the major carcinogen-DNA adducts formed in cultured human esophagus. Cancer Res. 1979, 39, 4401–4406. [Google Scholar]
- Herron, D.C.; Shank, R.C. Methylated purines in human liver DNA after probable dimethylnitrosamine poisoning. Cancer Res. 1980, 40, 3116–3117. [Google Scholar]
- Kyrtopoulos, S.A. DNA adducts in humans after exposure to methylating agents. Mutat. Res. 1998, 405, 135–143. [Google Scholar] [CrossRef]
- Phillips, D.H. Smoking-related DNA and protein adducts in human tissues. Carcinogenesis 2002, 23, 1979–2004. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Stepanov, I.; Hecht, S.S. Recent studies on DNA adducts resulting from human exposure to tobacco smoke. Toxics 2019, 7, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foiles, P.G.; Miglietta, L.M.; Akerkar, S.A.; Everson, R.B.; Hecht, S.S. Detection of O6-methyldeoxyguanosine in human placental DNA. Cancer Res. 1988, 48, 4184–4188. [Google Scholar] [PubMed]
- Kang, H.I.; Konishi, C.; Eberle, G.; Rajewsky, M.F.; Kuroki, T.; Huh, N.H. Highly sensitive, specific detection of O6-methylguanine, O4-methylthymine, and O4-ethylthymine by the combination of high-performance liquid chromatography prefractionation, 32P postlabeling, and immunoprecipitation. Cancer Res. 1992, 52, 5307–5312. [Google Scholar] [PubMed]
- Kang, H.; Konishi, C.; Kuroki, T.; Huh, N. Detection of O6-methylguanine, O4-methylthymine and O4-ethylthymine in human liver and peripheral blood leukocyte DNA. Carcinogenesis 1995, 16, 1277–1280. [Google Scholar] [CrossRef]
- Umbenhauer, D.; Wild, C.P.; Montesano, R.; Saffhill, R.; Boyle, J.M.; Huh, N.; Kirstein, U.; Thomale, J.; Rajewsky, M.F.; Lu, S.H. O6-methyldeoxyguanosine in oesophageal DNA among individuals at high risk of oesophageal cancer. Int. J. Cancer 1985, 36, 661–665. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. Some N-Nitroso Compounds. In IARC Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans; IARC: Lyon, France, 1978; Volume 17. [Google Scholar]
- Campillo, N.; Viñas, P.; Martínez-Castillo, N.; Hernández-Córdoba, M. Determination of volatile nitrosamines in meat products by microwave-assisted extraction and dispersive liquid-liquid microextraction coupled to gas chromatography-mass spectrometry. J. Chromatogr. A 2011, 1218, 1815–1821. [Google Scholar] [CrossRef]
- West, D.M.; Wu, Q.; Donovan, A.; Shi, H.; Ma, Y.; Jiang, H.; Wang, J. N-nitrosamine formation by monochloramine, free chlorine, and peracetic acid disinfection with presence of amine precursors in drinking water system. Chemosphere 2016, 153, 521–527. [Google Scholar] [CrossRef]
- Lim, H.H.; Oh, Y.S.; Shin, H.S. Determination of N-nitrosodimethylamine and N-nitrosomethylethylamine in drug substances and products of sartans, metformin and ranitidine by precipitation and solid phase extraction and gas chromatography-tandem mass spectrometry. J. Pharm. Biomed. Anal. 2020, 189, 113460. [Google Scholar] [CrossRef]
- Oldham, M.J.; DeSoi, D.J.; Rimmer, L.T.; Wagner, K.A.; Morton, M.J. Insights from analysis for harmful and potentially harmful constituents (HPHCs) in tobacco products. Regul. Toxicol. Pharmacol. 2014, 70, 138–148. [Google Scholar] [CrossRef]
- Lijinsky, W.; Reuber, M.D. Carcinogenicity in rats of nitrosomethylethylamines labeled with deuterium in several positions. Cancer Res. 1980, 40, 19–21. [Google Scholar]
- Lijinsky, W.; Saavedra, J.E.; Reuber, M.D.; Singer, S.S. Esophageal carcinogenesis in F344 rats by nitrosomethylethylamines substituted in the ethyl group. J. Natl. Cancer Inst. 1982, 68, 681–684. [Google Scholar] [PubMed]
- von Hofe, E.; Schmerold, I.; Nims, R.W.; Keefer, L.K.; Reist, E.J.; Kleihues, P. ß-deuteration of N-nitrosoethylmethylamine causes a shift in DNA methylation from rat liver to esophagus. Carcinogenesis 1991, 12, 545–549. [Google Scholar] [CrossRef] [PubMed]
- von Hofe, E.; Grahmann, F.; Keefer, L.K.; Lijinsky, W.; Nelson, V.; Kleihues, P. Methylation versus ethylation of DNA in target and nontarget tissues of Fischer 344 rats treated with N-nitrosomethylethylamine. Cancer Res. 1986, 46, 1038–1042. [Google Scholar] [PubMed]
- von Hofe, E.; Kleihues, P.; Keefer, L.K. Extent of DNA 2-hydroxyethylation by N-nitrosomethylethylamine and N-nitrosodiethylamine in vivo. Carcinogenesis 1986, 7, 1335–1337. [Google Scholar] [CrossRef] [Green Version]
- von Hofe, E.; Kleihues, P. Comparative studies on hepatic DNA alkylation in rats by N-nitrosomethylethylamine and N-nitrosodimethylamine plus N-nitrosodiethylamine. J. Cancer Res. Clin. Oncol. 1986, 112, 205–209. [Google Scholar] [CrossRef]
- National Toxicology Program. Report on Carcinogens, 12th ed.; U.S. Department of Health and Human Services, Public Health Service: Research Triangle Park, NC, USA, 2011.
- Sen, N.P.; Tessier, L.; Seaman, S.W. Determination of N-nitrosoproline and N-nitrososarcosine in malt and beer. J. Agric. Food Chem. 1983, 31, 1033–1036. [Google Scholar] [CrossRef]
- Wu, J.C.; Rickert, W.S.; Masters, A.; Joza, P. Determination of N-nitrososarcosine in tobacco and smokeless tobacco products using isotope dilution liquid chromatography tandem mass spectrometry. Anal. Methods 2012, 4, 3448–3452. [Google Scholar] [CrossRef]
- Werneth, M.; Pani, J.; Hofbauer, L.; Pummer, S.; Weber, M.T.; Pour, G.; Kahlig, H.; Mayer-Helm, B.; Stepan, H. Stereospecific response of E/Z-isomers of N-nitrososarcosine in LC-ESI-MS/MS. J. Chromatogr. Sci. 2021, 59, 813–822. [Google Scholar] [CrossRef]
- Derave, W.; Vanden Eede, E.; Hespel, P.; Carmella, S.G.; Hecht, S.S. Oral creatine supplementation in humans does not elevate urinary excretion of the carcinogen N-nitrososarcosine. Nutrition 2006, 22, 332–333. [Google Scholar] [CrossRef]
- Friedman, M.A. Reaction of sodium nitrite with dimethlglycine produces nitrososarcosine. Bull. Environ. Contam. Toxicol. 1975, 13, 226–232. [Google Scholar] [CrossRef]
- Friedman, M.A. Nitrosation of sarcosine: Chemical kinetics and gastric assay. Bull. Environ. Contam. Toxicol. 1972, 8, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.Y.; Wang, D.Y.; Tanaka, M.; Igarashi, H.; Kamo, T.; Shen, Q.; Sugimura, H.; Kino, I. Efficient and specific induction of esophageal tumors in rats by precursors of N-nitrososarcosine ethyl ester. Pathol. Int. 1995, 45, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Shubin, A.V.; Lesovaya, E.A.; Kirsanov, K.I.; Antoshina, E.E.; Trukhanova, L.S.; Gorkova, T.G.; Belitsky, G.A.; Yakubovskaya, M.G.; Demidyuk, I.V. Re-examination of the esophageal squamous cell carcinoma model in rats induced by N-nitrososarcosine ethyl ester precursors. Bull. Exp. Biol. Med. 2018, 164, 676–679. [Google Scholar] [CrossRef]
- Ohshima, H.; Bereziat, J.C.; Bartsch, H. Monitoring N-nitrosamino acids excreted in the urine and feces of rats as an index for endogenous nitrosation. Carcinogenesis 1982, 3, 115–120. [Google Scholar] [CrossRef]
- Cheng, G.; Wang, M.; Villalta, P.W.; Hecht, S.S. Detection of 7-(2′-carboxyethyl)guanine but not 7-carboxymethylguanine in human liver DNA. Chem. Res. Toxicol. 2010, 23, 1089–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, K.H.; Archer, M.C. Studies on the metabolic activation of ß-keto nitrosamines: Mechanisms of DNA methylation by N-(2-oxopropyl)-N-nitrosourea and N-nitroso-N-acetoxymethyl-N-2-oxopropylamine. Cell Biol. Int. 1984, 48, 169–179. [Google Scholar]
- Krüger, F.W. Metabolism of nitrosamines in vivo. I. Evidence for -oxydation of aliphatic di-n-alkylnitrosamines: The stimultaneous formation of 7-methylguanine besides 7-propyl- or 7-butylguanine after application of di-n-propyl- or di-n-butylnitrosamine. Z. Krebsforsch. Klin. Onkol. Cancer Res. Clin. Oncol. 1971, 76, 145–154. [Google Scholar] [CrossRef]
- Teiber, J.F.; Macé, K.; Hollenberg, P.F. Metabolism of the ß-oxidized intermediates of N-nitrosodi-n-propylamine: N-nitroso-ß-hydroxypropylpropylamine and N-nitroso-ß-oxopropylpropylamine. Carcinogenesis 2001, 22, 499–506. [Google Scholar] [CrossRef] [Green Version]
- Leung, K.H.; Archer, M.C. Mechanism of DNA methylation by N-nitroso(2-oxopropyl)propylamine. Carcinogenesis 1985, 6, 189–191. [Google Scholar] [CrossRef]
- Harrison, K.L.; Jukes, R.; Cooper, D.P.; Shuker, D.E. Detection of concomitant formation of O6-carboxymethyl- and O6-methyl-2′-deoxyguanosine in DNA exposed to nitrosated glycine derivatives using a combined immunoaffinity/HPLC method. Chem. Res. Toxicol. 1999, 12, 106–111. [Google Scholar] [CrossRef]
- Anderson, D.; Blowers, S.D. Limited cancer bioassay to test a potential food chemical. Lancet 1994, 344, 343–344. [Google Scholar] [CrossRef]
- Zurlo, J.; Curphey, T.J.; Hiley, R.; Longnecker, D.S. Identification of 7-carboxymethylguanine in DNA from pancreatic acinar cells exposed to azaserine. Cancer Res. 1982, 42, 1286–1288. [Google Scholar] [PubMed]
- Harrison, K.L.; Fairhurst, N.; Challis, B.C.; Shuker, D.E. Synthesis, characterization, and immunochemical detection of O6-(carboxymethyl)-2′-deoxyguanosine: A DNA adduct formed by nitrosated glycine derivatives. Chem. Res. Toxicol. 1997, 10, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Shuker, D.E.; Margison, G.P. Nitrosated glycine derivatives as a potential source of O6-methylguanine in DNA. Cancer Res. 1997, 57, 366–369. [Google Scholar] [PubMed]
- Cupid, B.C.; Zeng, Z.; Singh, R.; Shuker, D.E. Detection of O6-carboxymethyl-2′-deoxyguanosine in DNA following reaction of nitric oxide with glycine and in human blood DNA using a quantitative immunoslot blot assay. Chem. Res. Toxicol. 2004, 17, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Gottschalg, E.; Scott, G.B.; Burns, P.A.; Shuker, D.E. Potassium diazoacetate-induced p53 mutations in vitro in relation to formation of O6-carboxymethyl- and O6-methyl-2′-deoxyguanosine DNA adducts: Relevance for gastrointestinal cancer. Carcinogenesis 2007, 28, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Jackson, P.E.; Hall, C.N.; Badawi, A.F.; O’Connor, P.J.; Cooper, D.P.; Povey, A.C. Frequency of Ki-ras mutations and DNA alkylation in colorectal tissue from individuals living in Manchester. Mol. Carcinog. 1996, 16, 12–19. [Google Scholar] [CrossRef]
- The EUROGAST Study Group. O6-Methylguanine in blood leucocyte DNA: An association with the geographic prevalence of gastric cancer and with low levels of serum pepsinogen A, a marker of severe chronic atrophic gastritis. Carcinogenesis 1994, 15, 1815–1820. [Google Scholar]
- Hall, C.N.; Badawi, A.F.; O’Connor, P.J.; Saffhill, R. The detection of alkylation damage in the DNA of human gastrointestinal tissues. Br. J. Cancer 1991, 64, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Kyrtopoulos, S.A.; Vrotsou, B.; Golematis, B.; Bonatsos, M.; Lakiotis, G. O6-Methylguanine-DNA transmethylase activity in extracts of human gastric mucosa. Carcinogenesis 1984, 5, 943–947. [Google Scholar] [CrossRef]
- Povey, A.C.; Hall, C.N.; Badawi, A.F.; Cooper, D.P.; O’Connor, P.J. Elevated levels of the pro-carcinogenic adduct, O6-methylguanine, in normal DNA from the cancer prone regions of the large bowel. Gut 2000, 47, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.P.; Reed, P.I. N-nitroso compounds in fresh gastric juice and their relation to intragastric pH and nitrite employing an improved analytical method. Carcinogenesis 1993, 14, 2547–2551. [Google Scholar] [CrossRef] [PubMed]
- Bingham, S.A.; Pignatelli, B.; Pollock, J.R.; Ellul, A.; Malaveille, C.; Gross, G.; Runswick, S.; Cummings, J.H.; O’Neill, I.K. Does increased endogenous formation of N-nitroso compounds in the human colon explain the association between red meat and colon cancer? Carcinogenesis 1996, 17, 515–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, Y. Chemical synthesis of oligodeoxyribonucleotides containing N3- and O4-carboxymethylthymidine and their formation in DNA. Nucleic Acids Res. 2009, 37, 336–345. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y. Synthesis and characterization of oligodeoxyribonucleotides containing a site-specifically incorporated N6-carboxymethyl-2′-deoxyadenosine or N4-carboxymethyl-2′-deoxycytidine. Nucleic Acids Res. 2010, 38, 6774–6784. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Wang, J.; Wang, P.; Wang, Y. Quantification of azaserine-induced carboxymethylated and methylated DNA lesions in cells by nanoflow liquid chromatography-nanoelectrospray ionization tandem mass spectrometry coupled with the stable isotope-dilution method. Anal. Chem. 2016, 88, 8036–8042. [Google Scholar] [CrossRef] [Green Version]
- Senthong, P.; Millington, C.L.; Wilkinson, O.J.; Marriott, A.S.; Watson, A.J.; Reamtong, O.; Eyers, C.E.; Williams, D.M.; Margison, G.P.; Povey, A.C. The nitrosated bile acid DNA lesion O6-carboxymethylguanine is a substrate for the human DNA repair protein O6-methylguanine-DNA methyltransferase. Nucleic Acids Res. 2013, 41, 3047–3055. [Google Scholar] [CrossRef]
- O’Driscoll, M.; Macpherson, P.; Xu, Y.Z.; Karran, P. The cytotoxicity of DNA carboxymethylation and methylation by the model carboxymethylating agent azaserine in human cells. Carcinogenesis 1999, 20, 1855–1862. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Leng, J.; Wang, Y. DNA replication studies of N-nitroso compound-induced O6-alkyl-2′-deoxyguanosine lesions in Escherichia coli. J. Biol. Chem. 2019, 294, 3899–3908. [Google Scholar] [CrossRef]
- Wu, J.; Wang, P.; Li, L.; Williams, N.L.; Ji, D.; Zahurancik, W.J.; You, C.; Wang, J.; Suo, Z.; Wang, Y. Replication studies of carboxymethylated DNA lesions in human cells. Nucleic Acids Res. 2017, 45, 7276–7284. [Google Scholar] [CrossRef] [Green Version]
- Swanson, A.L.; Wang, J.; Wang, Y. In vitro replication studies of carboxymethylated DNA lesions with Saccharomyces cerevisiae polymerase η. Biochemistry 2011, 50, 7666–7673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewin, M.H.; Bailey, N.; Bandaletova, T.; Bowman, R.; Cross, A.J.; Pollock, J.; Shuker, D.E.; Bingham, S.A. Red meat enhances the colonic formation of the DNA adduct O6-carboxymethyl guanine: Implications for colorectal cancer risk. Cancer Res. 2006, 66, 1859–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.S.; Yoo, J.S.; Ishizaki, H.; Hong, J.Y. Cytochrome P450IIE1: Roles in nitrosamine metabolism and mechanisms of regulation. Drug Metab. Rev. 1990, 22, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Camus, A.M.; Geneste, O.; Honkakoski, P.; Béréziat, J.C.; Henderson, C.J.; Wolf, C.R.; Bartsch, H.; Lang, M.A. High variability of nitrosamine metabolism among individuals: Role of cytochromes P450 2A6 and 2E1 in the dealkylation of N-nitrosodimethylamine and N-nitrosodiethylamine in mice and humans. Mol. Carcinog. 1993, 7, 268–275. [Google Scholar] [CrossRef]
- Verna, L.; Whysner, J.; Williams, G.M. N-nitrosodiethylamine mechanistic data and risk assessment: Bioactivation, DNA-adduct formation, mutagenicity, and tumor initiation. Pharmacol. Ther. 1996, 71, 57–81. [Google Scholar] [CrossRef]
- Singer, B. In vivo formation and persistence of modified nucleosides resulting from alkylating agents. Environ. Health Perspect. 1985, 62, 41–48. [Google Scholar] [CrossRef]
- Pegg, A.E. Alkylation and subsequent repair of DNA after exposure to dimethylnitrosamine and related carcinogens. Rev. Biochem. Toxicol. 1983, 38–133. [Google Scholar]
- Swenberg, J.A.; Dyroff, M.C.; Bedell, M.A.; Popp, J.A.; Huh, N.; Kirstein, U.; Rajewsky, M.F. O4-ethyldeoxythymidine, but not O6-ethyldeoxyguanosine, accumulates in hepatocyte DNA of rats exposed continuously to diethylnitrosamine. Proc. Natl. Acad. Sci. USA 1984, 81, 1692–1695. [Google Scholar] [CrossRef] [Green Version]
- Boucheron, J.A.; Richardson, F.C.; Morgan, P.H.; Swenberg, J.A. Molecular dosimetry of O4-ethyldeoxythymidine in rats continuously exposed to diethylnitrosamine. Cancer Res. 1987, 47, 1577–1581. [Google Scholar]
- Richardson, F.C.; Dyroff, M.C.; Boucheron, J.A.; Swenberg, J.A. Differential repair of O4-alkylthymidine following exposure to methylating and ethylating hepatocarcinogens. Carcinogenesis 1985, 6, 625–629. [Google Scholar] [CrossRef]
- Pegg, A.E. Properties of the O6-Alkylguanine-DNA Repair System of Mammalian Cells. In IARC Scientific Publication No. 57: N-Nitroso Compounds: Occurrence, Biological Effects and Relevance to Human Cancer; O’Neill, I., von Borstel, R., Miller, C., Long, J., Bartsch, H., Eds.; IARC: Lyon, France, 1984; pp. 575–580. [Google Scholar]
- Scherer, E.; Timmer, A.P.; Emmelot, P. Formation by diethylnitrosamine and persistence of O4-ethylthymidine in rat liver DNA in vivo. Cancer Lett. 1980, 10, 1–6. [Google Scholar] [CrossRef]
- Dolan, M.E.; Scicchitano, D.; Singer, B.; Pegg, A.E. Comparison of repair of methylated pyrimidines in poly(dT) by extracts from rat liver and Escherichia coli. Biochem. Biophys. Res. Commun. 1984, 123, 324–330. [Google Scholar] [CrossRef]
- Dragan, Y.P.; Hully, J.R.; Nakamura, J.; Mass, M.J.; Swenberg, J.A.; Pitot, H.C. Biochemical events during initiation of rat hepatocarcinogenesis. Carcinogenesis 1994, 15, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Amato, N.J.; Zhai, Q.; Wang, Y. Cytotoxic and mutagenic properties of O4-alkylthymidine lesions in Escherichia coli cells. Nucleic Acids Res. 2015, 43, 10795–10803. [Google Scholar] [CrossRef] [Green Version]
- Graves, R.J.; Li, B.F.; Swann, P.F. Repair of O6-methylguanine, O6-ethylguanine, O6-isopropylguanine and O4-methylthymine in synthetic oligodeoxynucleotides by Escherichia coli ada gene O6-alkylguanine-DNA-alkyltransferase. Carcinogenesis 1989, 10, 661–666. [Google Scholar] [CrossRef]
- Wilkinson, M.C.; Potter, P.M.; Cawkwell, L.; Georgiadis, P.; Patel, D.; Swann, P.F.; Margison, G.P. Purification of the E. coli ogt gene product to homogeneity and its rate of action on O6-methylguanine, O6-ethylguanine and O4-methylthymine in dodecadeoxyribonucleotides. Nucleic Acids Res. 1989, 17, 8475–8484. [Google Scholar] [CrossRef]
- Saffhill, R. In vitro miscoding of alkylthymines with DNA and RNA polymerases. Cell Biol. Int. 1985, 53, 121–130. [Google Scholar] [CrossRef]
- Wu, J.; Li, L.; Wang, P.; You, C.; Williams, N.L.; Wang, Y. Translesion synthesis of O4-alkylthymidine lesions in human cells. Nucleic Acids Res. 2016, 44, 9256–9265. [Google Scholar]
- Singer, B.; Spengler, S.J.; Fraenkel-Conrat, H.; Kuśmierek, J.T. O4-Methyl, -ethyl, or -isopropyl substituents on thymidine in poly(dA-dT) all lead to transitions upon replication. Proc. Natl. Acad. Sci. USA 1986, 83, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Wang, W.; Wu, J.; Shin, J.H.; Wang, P.; Unarta, I.C.; Chong, J.; Wang, Y.; Wang, D. Mechanism of DNA alkylation-induced transcriptional stalling, lesion bypass, and mutagenesis. Proc. Natl. Acad. Sci. USA 2017, 114, e7082–e7091. [Google Scholar] [CrossRef] [Green Version]
- Andersen, N.; Wang, P.; Wang, Y. Replication across regioisomeric ethylated thymidine lesions by purified DNA polymerases. Chem. Res. Toxicol. 2013, 26, 1730–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, N.L.; Wang, P.; Wang, Y. Replicative bypass of O2-alkylthymidine lesions in vitro. Chem. Res. Toxicol. 2016, 29, 1755–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, N.L.; Wang, P.; Wu, J.; Wang, Y. In vitro lesion bypass studies of O4-alkylthymidines with human DNA polymerase η. Chem. Res. Toxicol. 2016, 29, 669–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Q.; Wang, P.; Cai, Q.; Wang, Y. Syntheses and characterizations of the in vivo replicative bypass and mutagenic properties of the minor-groove O2-alkylthymidine lesions. Nucleic Acids Res. 2014, 42, 10529–10537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Wang, P.; Li, L.; You, C.; Wang, Y. Cytotoxic and mutagenic properties of minor-groove O2-alkylthymidine lesions in human cells. J. Biol. Chem. 2018, 293, 8638–8644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Kaur, B.; Farmer, P.B. Detection of DNA damage derived from a direct acting ethylating agent present in cigarette smoke by use of liquid chromatography-tandem mass spectrometry. Chem. Res. Toxicol. 2005, 18, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Wang, Y.C.; Lin, W.P. Analysis of ethylated thymidine adducts in human leukocyte DNA by stable isotope dilution nanoflow liquid chromatography-nanospray ionization tandem mass spectrometry. Anal. Chem. 2012, 84, 2521–2527. [Google Scholar] [CrossRef]
- Godschalk, R.; Nair, J.; Kliem, H.C.; Wiessler, M.; Bouvier, G.; Bartsch, H. Modified immunoenriched 32P-HPLC assay for the detection of O4-ethylthymidine in human biomonitoring studies. Chem. Res. Toxicol. 2002, 15, 433–437. [Google Scholar] [CrossRef]
- Godschalk, R.; Nair, J.; van Schooten, F.J.; Risch, A.; Drings, P.; Kayser, K.; Dienemann, H.; Bartsch, H. Comparison of multiple DNA adduct types in tumor adjacent human lung tissue: Effect of cigarette smoking. Carcinogenesis 2002, 23, 2081–2086. [Google Scholar] [CrossRef] [Green Version]
- Anna, L.; Kovács, K.; Gyorffy, E.; Schoket, B.; Nair, J. Smoking-related O4-ethylthymidine formation in human lung tissue and comparisons with bulky DNA adducts. Mutagenesis 2011, 26, 523–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.J.; Liu, Y.F. Simultaneous quantitative analysis of N3-ethyladenine and N7-ethylguanine in human leukocyte deoxyribonucleic acid by stable isotope dilution capillary liquid chromatography-nanospray ionization tandem mass spectrometry. J. Chromatogr. A 2013, 1271, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Lin, C.R. Noninvasive measurement of smoking-associated N3-ethyladenine and N7-ethylguanine in human salivary DNA by stable isotope dilution nanoflow liquid chromatography-nanospray ionization tandem mass spectrometry. Toxicol. Lett. 2014, 225, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Balbo, S.; Villalta, P.W.; Hecht, S.S. Quantitation of 7-ethylguanine in leukocyte DNA from smokers and nonsmokers by liquid chromatography-nanoelectrospray-high resolution tandem mass spectrometry. Chem. Res. Toxicol. 2011, 24, 1729–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prevost, V.; Shuker, D.E. Cigarette smoking and urinary 3-alkyladenine excretion in man. Chem. Res. Toxicol. 1996, 9, 439–444. [Google Scholar] [CrossRef]
- Kopplin, A.; Eberle-Adamkiewicz, G.; Glüsenkamp, K.H.; Nehls, P.; Kirstein, U. Urinary excretion of 3-methyladenine and 3-ethyladenine after controlled exposure to tobacco smoke. Carcinogenesis 1995, 16, 2637–2641. [Google Scholar] [CrossRef]
- Prevost, V.; Shuker, D.E.; Friesen, M.D.; Eberle, G.; Rajewsky, M.F.; Bartsch, H. Immunoaffinity purification and gas chromatography-mass spectrometric quantification of 3-alkyladenines in urine: Metabolism studies and basal excretion levels in man. Carcinogenesis 1993, 14, 199–204. [Google Scholar] [CrossRef]
- Carmella, S.G.; Chen, M.; Villalta, P.W.; Gurney, J.G.; Hatsukami, D.K.; Hecht, S.S. Ethylation and methylation of hemoglobin in smokers and non-smokers. Carcinogenesis 2002, 23, 1903–1910. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, M.; Villalta, P.W.; Hecht, S.S. Liquid chromatography-electrospray ionization tandem mass spectrometry analysis of 7-ethylguanine in human liver DNA. Chem. Res. Toxicol. 2007, 20, 1498–1502. [Google Scholar] [CrossRef]
- Agency for Toxic Substances and Disease Registry. Toxicological Profile for N-Nitrosodi-n-Propylamine; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2019.
- Shu, L.; Hollenberg, P.F. Identification of the cytochrome P450 isozymes involved in the metabolism of N-nitrosodipropyl-,N-nitrosodibutyl- and N-nitroso-n-butyl-n-propylamine. Carcinogenesis 1996, 17, 839–848. [Google Scholar] [CrossRef]
- Teiber, J.F.; Hollenberg, P.F. Identification of the human liver microsomal cytochrome P450s involved in the metabolism of N-nitrosodi-n-propylamine. Carcinogenesis 2000, 21, 1559–1566. [Google Scholar]
- Farrelly, J.G.; Stewart, M.L.; Lijinsky, W. The metabolism of nitrosodi-n-propylamine, nitrosodiallylamine and nitrosodiethanolamine. Carcinogenesis 1984, 5, 1015–1019. [Google Scholar] [CrossRef] [PubMed]
- Park, K.K.; Archer, M.C. Microsomal metabolism of N-nitrosodi-n-propylamine: Formation of products resulting from alpha- and beta-oxidation. Cell Biol. Int. 1978, 22, 83–90. [Google Scholar] [CrossRef]
- Park, K.K.; Wishnok, J.S.; Archer, M.C. Mechanism of alkylation by N-nitroso compounds: Detection of rearranged alcohol in the microsomal metabolism of N-nitrosodi-n-propylamine and base-catalyzed decomposition of N-n-propyl-N-nitrosourea. Cell Biol. Int. 1977, 18, 349–354. [Google Scholar] [CrossRef]
- Leung, K.H.; Archer, M.C. Urinary metabolites of N-nitrosodipropylamine, N-nitroso-2-hydroxypropylpropylamine and N-nitroso-2-oxopropylpropylamine in the rat. Carcinogenesis 1981, 2, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Park, K.K.; Archer, M.C. Metabolism of N-nitroso-2-oxopropylpropylamine by rat liver: Formation of products resulting from both oxidation and reduction. Cancer Biochem. Biophys. 1978, 3, 37–40. [Google Scholar]
- Kokkinakis, D.M.; Hollenberg, P.F.; Scarpelli, D.G. Major urinary metabolites in hamsters and rats treated with N-nitroso(2-hydroxypropyl)(2-oxopropyl)amine. Cancer Res. 1985, 45, 3586–3592. [Google Scholar]
- Bauman, P.A.; Hotchkiss, J.H.; Parker, R.S. Metabolism of N-nitrosodi-n-propylamine and N-nitrosodiallylamine by isolated rat hepatocytes. Cancer Lett. 1985, 28, 229–236. [Google Scholar] [CrossRef]
- Appel, K.E.; Graf, H. Metabolic nitrite formation from N-nitrosamines: Evidence for a cytochrome P-450 dependent reaction. Carcinogenesis 1982, 3, 293–296. [Google Scholar] [CrossRef]
- Krüger, F.W. Metabolism of nitrosamines in vivo. II. On the methylation of nucleic acids by aliphatic di-n-alkyl-nitrosamines in vivo, caused by beta-oxydation: The increased formation of 7-methyl-guanine after application of beta-hydroxypropyl-propyl-nitrosamine compared to that after application of di-n-propyl-nitrosamine. Z. Krebsforsch. Klin. Onkol. Cancer Res. Clin. Oncol. 1973, 79, 90–97. [Google Scholar]
- Krüger, F.W.; Bertram, B. Metabolism of nitrosamines in vivo. 3. On the methylation of nucleic acids by aliphatic Di-n-alkyl-nitrosamines in vivo resulting from ß-oxidation: The formation of 7-methylguanine after application of 2-oxo-propyl-propyl-nitrosamine and methyl-propyl-nitro-samine. Z. Krebsforsch. Klin. Onkol. Cancer Res. Clin. Oncol. 1973, 80, 189–196. [Google Scholar]
- Shu, L.; Hollenberg, P.F. Alkylation of cellular macromolecules and target specificity of carcinogenic nitrosodialkylamines: Metabolic activation by cytochromes P450 2B1 and 2E1. Carcinogenesis 1997, 18, 801–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokkinakis, D.M. Alkylation of rodent tissue DNA induced by N-nitrosobis(2-hydroxypropyl)amine. Carcinogenesis 1992, 13, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Kokkinakis, D.M. Differences between pancreatropic nitrosamine carcinogens and N-nitrosodimethylamine in methylating DNA in various tissues of hamsters and rats. Cell Biol. Int. 1991, 78, 167–181. [Google Scholar] [CrossRef]
- Kokkinakis, D.M. Differences in DNA-guanine alkylation between male Sprague-Dawley rats and Syrian hamsters following exposure to a single dose of pancreatic nitrosamine carcinogens. Chem. Res. Toxicol. 1990, 3, 150–156. [Google Scholar] [CrossRef]
- Fan, T.Y.; Morrison, J.; Rounbehler, D.P.; Ross, R.; Fine, D.H.; Miles, W.; Sen, N.P. N-Nitrosodiethanolamine in synthetic cutting fluids: A part-per-hundred impurity. Science 1977, 196, 70–71. [Google Scholar] [CrossRef]
- Fan, T.Y.; Goff, U.; Song, L.; Fine, D.H.; Arsenault, G.P.; Biemann, K. N-Nitrosodiethanolamine in cosmetics, lotions and shampoos. Food Cosmet. Toxicol. 1977, 15, 423–430. [Google Scholar] [CrossRef]
- Brunnemann, K.D.; Hoffmann, D. Assessment of the carcinogenic N-nitrosodiethanolamine in tobacco products and tobacco smoke. Carcinogenesis 1981, 2, 1123–1127. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. Some Industrial Chemicals. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 2000; Volume 77. [Google Scholar]
- Lethco, E.J.; Wallace, W.C.; Brouwer, E. The fate of N-nitrosodiethanolamine after oral and topical administration to rats. Food Chem. Toxicol. 1982, 20, 401–406. [Google Scholar] [CrossRef]
- Preussmann, R.; Wurtele, G.; Eisenbrand, G.; Spiegelhalder, B. Urinary excretion of N-nitrosodiethanolamine administered orally to rats. Cancer Lett. 1978, 4, 207–209. [Google Scholar] [CrossRef]
- Preussmann, R.; Habs, M.; Habs, H.; Schmähl, D. Carcinogenicity of N-nitrosodiethanolamine in rats at five different dose levels. Cancer Res. 1982, 42, 5167–5171. [Google Scholar]
- Hoffmann, D.; Rivenson, A.; Adams, J.D.; Juchatz, A.; Vinchkoski, N.; Hecht, S.S. A study of tobacco carcinogenesis. 24. Effects of route of administration and dose on the carcinogenicity of N-nitrosodiethanolamine in the Syrian Golden hamster. Cancer Res. 1983, 43, 2521–2524. [Google Scholar]
- Airoldi, L.; Bonfanti, M.; Benfenati, E.; Tavecchia, P.; Fanelli, R. Identification of an acidic metabolite of N-nitrosodiethanolamine isolated from rat urine. Biomed. Mass. Spectrom. 1983, 10, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Farrelly, J.G.; Thomas, B.J.; Lijinsky, W. Metabolism and Cellular Interactions of N-Nitrosodiethanolamine. In IARC Scientific Publication No. 84: The Relevance of N-Nitroso Compounds to Human Cancer: Exposures and Mechanisms; Bartsch, H., O’Neill, I., Schulte-Hermann, R., Eds.; IARC: Lyon, France, 1987; pp. 87–90. [Google Scholar]
- Sterzel, W.; Eisenbrand, G. N-nitrosodiethanolamine is activated in the rat to an ultimate genotoxic metabolite by sulfotransferase. J. Cancer Res. Clin. Oncol. 1986, 111, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Michejda, C.J.; Andrews, A.W.; Koepke, S.R. Derivatives of side-chain hydroxylated nitrosamines direct acting mutagens in Salmonella typhimurium. Mutat. Res. 1979, 67, 301–308. [Google Scholar] [CrossRef]
- Loeppky, R.N.; Goelzer, P. Microsome-mediated oxidation of N-nitrosodiethanolamine (NDELA), a bident carcinogen. Chem. Res. Toxicol. 2002, 15, 457–469. [Google Scholar] [CrossRef]
- Hecht, S.S. N-Nitroso-2-hydroxymorpholine, a mutagenic metabolite of N-nitrosodiethanolamine. Carcinogenesis 1984, 5, 1745–1747. [Google Scholar] [CrossRef]
- Hecht, S.S.; Lijinsky, W.; Kovatch, R.M.; Chung, F.L.; Saavedra, J.E. Comparative tumorigenicity of N-nitroso-2-hydroxymorpholine, N-nitrosodiethanolamine and N-nitrosomorpholine in A/J mice and F344 rats. Carcinogenesis 1989, 10, 1475–1477. [Google Scholar] [CrossRef]
- Loeppky, R.N.; Ye, Q.; Goelzer, P.; Chen, Y. DNA adducts from N-nitrosodiethanolamine and related beta-oxidized nitrosamines in vivo: 32P-postlabeling methods for glyoxal- and O6-hydroxyethyldeoxyguanosine adducts. Chem. Res. Toxicol. 2002, 15, 470–482. [Google Scholar] [CrossRef]
- Loeppky, R.N.; Cui, W.; Goelzer, P.; Park, M.; Ye, Q. Glyoxal-guanine DNA Adducts: Detection, Stability and Formation in vivo from Nitrosamines. In IARC Scientific Publication No. 150: Exocyclic DNA Adducts in Mutagenesis and Carcinogenesis; Singer, B., Bartsch, H., Eds.; IARC: Lyon, France, 1999; pp. 155–168. [Google Scholar]
- Loeppky, R.N.; Fuchs, A.; Janzowski, C.; Humberd, C.; Goelzer, P.; Schneider, H.; Eisenbrand, G. Probing the mechanism of the carcinogenic activation of N-nitrosodiethanolamine with deuterium isotope effects: In Vivo induction of DNA single-strand breaks and related in vitro assays. Chem. Res. Toxicol. 1998, 11, 1556–1566. [Google Scholar] [CrossRef]
- Loeppky, R.N. The mechanism of bioactivation of N-nitrosodiethanolamine. Drug Metab. Rev. 1999, 31, 175–193. [Google Scholar] [CrossRef]
- Loeppky, R.N.; Sukhtankar, S.; Gu, F.; Park, M. The carcinogenic significance of reactive intermediates derived from 3-acetoxy- and 5-acetoxy-2-hydroxy-N-nitrosomorpholine. Chem. Res. Toxicol. 2005, 18, 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Dennehy, M.K.; Loeppky, R.N. Mass spectrometric methodology for the determination of glyoxaldeoxyguanosine and O6-hydroxyethyldeoxyguanosine DNA adducts produced by nitrosamine bident carcinogens. Chem. Res. Toxicol. 2005, 18, 556–565. [Google Scholar] [CrossRef]
- Giménez-Campillo, C.; Pastor-Belda, M.; Campillo, N.; Hernández-Córdoba, M.; Viñas, P. Development of a new methodology for the determination of N-nitrosamines impurities in ranitidine pharmaceuticals using microextraction and gas chromatography-mass spectrometry. Talanta 2021, 223 Pt 2, 121659. [Google Scholar] [CrossRef] [PubMed]
- Imaida, K.; Wang, C.Y. Effect of sodium phenobarbital and sodium saccharin in AIN-76A diet on carcinogenesis initiated with N-[4-(5-nitro-2-furyl)-2-thiazolyl]formamide and N,N-dibutylnitrosamine in male F344 rats. Cancer Res. 1986, 46 Pt 1, 6160–6164. [Google Scholar] [PubMed]
- Fukushima, S.; Sakata, T.; Tagawa, Y.; Shibata, M.A.; Hirose, M.; Ito, N. Different modifying response of butylated hydroxyanisole, butylated hydroxytoluene, and other antioxidants in N,N-dibutylnitrosamine esophagus and forestomach carcinogenesis of rats. Cancer Res. 1987, 47, 2113–2116. [Google Scholar]
- Imaida, K.; Fukushima, S.; Inoue, K.; Masui, T.; Hirose, M.; Ito, N. Modifying effects of concomitant treatment with butylated hydroxyanisole or butylated hydroxytoluene on N,N-dibutylnitrosamine-induced liver, forestomach and urinary bladder carcinogenesis in F344 male rats. Cancer Lett. 1988, 43, 167–172. [Google Scholar] [CrossRef]
- Hirose, M.; Uwagawa, S.; Ozaki, K.; Takaba, K.; Ito, N. Effects of butylated hydroxyanisole pretreatment on low dose N-methyl-N′-nitro-N-nitrosoguanidine- or N,N-dibutylnitrosamine-induced rat forestomach or esophageal carcinogenesis. Carcinogenesis 1991, 12, 1773–1776. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.A.; Fukushima, S.; Takahashi, S.; Hasegawa, R.; Ito, N. Enhancing effects of sodium phenobarbital and N,N-dibutylnitrosamine on tumor development in a rat wide-spectrum organ carcinogenesis model. Carcinogenesis 1990, 11, 1027–1031. [Google Scholar] [CrossRef]
- Lijinsky, W.; Reuber, M.D. Carcinogenesis in Fischer rats by nitrosodipropylamine, nitrosodibutylamine and nitrosobis(2-oxopropyl)amine given by gavage. Cancer Lett. 1983, 19, 207–213. [Google Scholar] [CrossRef]
- Tsuda, H.; Mera, Y.; Seki, K.; Aoki, T.; Fukushima, S.; Ito, N. Induction of tumors in the liver, urinary bladder, esophagus and forestomach by short-term treatment with different doses of N,N′-dibutylnitrosamine in rats. Jpn. J. Cancer Res. 1987, 78, 227–234. [Google Scholar]
- Nishikawa, T.; Salim, E.I.; Morimura, K.; Kaneko, M.; Ogawa, M.; Kinoshita, A.; Osugi, H.; Kinoshita, H.; Fukushima, S. High susceptibility of p53 knockout mice to esophageal and urinary bladder carcinogenesis induced by N,N-dibutylnitrosamine. Cancer Lett. 2003, 194, 45–54. [Google Scholar] [CrossRef]
- Suzuki, E.; Mochizuki, M.; Wakabayashi, Y.; Okada, M. In vitro metabolic activation of N,N-dibutylnitrosamine in mutagenesis. Gan 1983, 74, 51–59. [Google Scholar]
- Suzuki, E.; Mochizuki, M.; Shibuya, K.; Okada, M. N,N-dibutylnitrosamine metabolism by rat liver S9 fraction and oxidation by chemical model systems. Gan 1983, 74, 41–50. [Google Scholar] [PubMed]
- Suzuki, E.; Okada, M. Metabolic fate of N,N-dibutylnitrosamine in the rat. Gan 1980, 71, 863–870. [Google Scholar] [PubMed]
- Vasconcelos-Nóbrega, C.; Colaço, A.; Lopes, C.; Oliveira, P.A. Review: BBN as an urothelial carcinogen. In Vivo 2012, 26, 727–739. [Google Scholar] [PubMed]
- Suzuki, E.; Okada, M. Metabolic fate of N-butyl-N-(4-hydroxybutyl)nitrosamine in the rat. Gan 1980, 71, 856–862. [Google Scholar] [PubMed]
- Bonfanti, M.; Magagnotti, C.; Bonati, M.; Fanelli, R.; Airoldi, L. Pharmacokinetic profile and metabolism of N-nitrosobutyl-(4-hydroxybutyl)amine in rats. Cancer Res. 1988, 48, 3666–3669. [Google Scholar] [PubMed]
- Suzuki, E.; Aoki, J.; Okada, M. Metabolic fate of N-butyl-N-(4-hydroxybutyl) nitrosamine and N,N-dibutylnitrosamine in the guinea pig, with reference to their carcinogenic effects on the urinary bladder. Gan 1981, 72, 547–551. [Google Scholar]
- Suzuki, E.; Osabe, M.; Okada, M.; Ishizaka, T.; Suzuki, T. Urinary metabolites of N-nitrosodibutylamine and N-nitrodibutylamine in the rat: Identification of N-acetyl-S-alkyl-L-cysteines. Jpn. J. Cancer Res. 1987, 78, 382–385. [Google Scholar]
- Bonfanti, M.; Magagnotti, C.; Galli, A.; Bagnati, R.; Moret, M.; Gariboldi, P.; Fanelli, R.; Airoldi, L. Determination of O6-butylguanine in DNA by immunoaffinity extraction/gas chromatography-mass spectrometry. Cancer Res. 1990, 50, 6870–6875. [Google Scholar]
- Airoldi, L.; Magagnotti, C.; Bonfanti, M.; Chiappetta, L.; Lolli, M.; Medana, C.; De Gregorio, G.; Fanelli, R. Detection of O6-butyl- and O6-(4-hydroxybutyl)guanine in urothelial and hepatic DNA of rats given the bladder carcinogen N-nitrosobutyl(4-hydroxybutyl)amine. Carcinogenesis 1994, 15, 2297–2301. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, D.; Brunnemann, K.D.; Adams, J.D.; Hecht, S.S. Formation and Analysis of N-Nitrosamines in Tobacco Products and their Endogenous Formation in Consumers. In IARC Scientific Publication No. 57: N-Nitroso Compounds: Occurrence, Biological Effects and Relevance to Human Cancer; O’Neill, I., von Borstel, R., Miller, C., Long, J., Bartsch, H., Eds.; IARC: Lyon, France, 1984; pp. 743–762. [Google Scholar]
- Tricker, A.R.; Pfundstein, B.; Kälble, T.; Preussmann, R. Secondary amine precursors to nitrosamines in human saliva, gastric juice, blood, urine and faeces. Carcinogenesis 1992, 13, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Lijinsky, W.; Reuber, M.D. Carcinogenic effect of nitrosopyrrolidine, nitrosopiperidine and nitrosohexamethyleneimine in Fischer rats. Cancer Lett. 1981, 12, 99–103. [Google Scholar] [CrossRef]
- Chung, F.L.; Tanaka, T.; Hecht, S.S. Induction of liver tumors in F344 rats by crotonaldehyde. Cancer Res. 1986, 46, 1285–1289. [Google Scholar] [PubMed]
- Gray, R.; Peto, R.; Brantom, P.; Grasso, P. Chronic nitrosamine ingestion in 1040 rodents: The effect of the choice of nitrosamine, the species studied, and the age of starting exposure. Cancer Res. 1991, 51 Pt 2, 6470–6491. [Google Scholar] [PubMed]
- Hecht, S.S.; Abbaspour, A.; Hoffman, D. A study of tobacco carcinogenesis XLII. Bioassay in mice of some structural analogues of tobacco-specific nitrosamines. Cancer Lett. 1988, 42, 141–145. [Google Scholar] [CrossRef]
- McCoy, G.D.; Hecht, S.S.; Katayama, S.; Wynder, E.L. Differential effect of chronic ethanol consumption on the carcinogenicity of N-nitrosopyrrolidine and N′-nitrosonornicotine in male Syrian Golden hamsters. Cancer Res. 1981, 41, 2849–2854. [Google Scholar]
- McCoy, G.D.; Hecht, S.S.; Furuya, K. The effect of chronic ethanol consumption on the tumorigenicity of N-nitrosopyrrolidine in male Syrian golden hamsters. Cancer Lett. 1986, 33, 151–159. [Google Scholar] [CrossRef]
- Hecht, S.S.; McCoy, G.D.; Chen, C.-H.B.; Hoffmann, D. The Metabolism of Cyclic Nitrosamines. In N-Nitroso Compounds; Scanlan, R.A., Tannenbaum, S.R., Eds.; American Chemical Society: Washington, DC, USA, 1981; Volume 174, pp. 49–75. [Google Scholar]
- Snyder, C.M.; Farrelly, J.G.; Lijinsky, W. Metabolism of three cyclic nitrosamines in Sprague-Dawley rats. Cancer Res. 1977, 37, 3530–3532. [Google Scholar]
- Wong, H.L.; Murphy, S.E.; Hecht, S.S. Cytochrome P450 2A-catalyzed metabolic activation of structurally similar carcinogenic nitrosamines: N′-nitrosonornicotine enantiomers, N-nitrosopiperidine, and N-nitrosopyrrolidine. Chem. Res. Toxicol. 2005, 18, 61–69. [Google Scholar] [CrossRef]
- Hecht, S.S.; Chen, C.B.; Hoffmann, D. Evidence for metabolic α hydroxylation of N-nitrosopyrrolidine. Cancer Res. 1978, 38, 215–218. [Google Scholar] [PubMed]
- Chen, C.B.; McCoy, G.D.; Hecht, S.S.; Hoffmann, D.; Wynder, E.L. High pressure liquid chromatographic assay for alpha hydroxylation of N-nitrosopyrrolidine by isolated rat liver microsomes. Cancer Res. 1978, 38 Pt 1, 3812–3816. [Google Scholar] [PubMed]
- Hecker, L.I.; Farrelly, J.G.; Smith, J.H.; Saavedra, J.E.; Lyon, P.A. Metabolism of the liver carcinogen N-nitrosopyrrolidine by rat liver microsomes. Cancer Res. 1979, 39 Pt 1, 2679–2686. [Google Scholar] [PubMed]
- Wang, M.Y.; Chung, F.L.; Hecht, S.S. Identification of crotonaldehyde as a hepatic microsomal metabolite formed by α-hydroxylation of the carcinogen N-nitrosopyrrolidine. Chem. Res. Toxicol. 1988, 1, 28–31. [Google Scholar] [CrossRef]
- Krüger, F.W.; Bertram, B. Metabolism of nitrosamines in vivo IV. Isolation of 3-hydroxy-1-nitrosopyrrolidine from rat urine after application of 1-nitrosopyrrolidine. Z. Krebsforsch. Klin. Onkol. Cancer Res. Clin. Oncol. 1975, 83, 255–260. [Google Scholar] [CrossRef]
- Cottrell, R.C.; Walters, D.G.; Young, P.J.; Phillips, J.C.; Lake, B.G.; Gangolli, S.D. Studies of the urinary metabolites of N-nitrosopyrrolidine in the rat. Toxicol. Appl. Pharmacol. 1980, 54, 368–376. [Google Scholar] [CrossRef]
- Wang, M.; McIntee, E.J.; Cheng, G.; Shi, Y.; Villalta, P.W.; Hecht, S.S. Identification of paraldol-deoxyguanosine adducts in DNA reacted with crotonaldehyde. Chem. Res. Toxicol. 2000, 13, 1065–1074. [Google Scholar] [CrossRef]
- Wang, M.; McIntee, E.J.; Cheng, G.; Shi, Y.; Villalta, P.W.; Hecht, S.S. A Schiff base is a major DNA adduct of crotonaldehyde. Chem. Res. Toxicol. 2001, 14, 423–430. [Google Scholar] [CrossRef]
- Wang, M.; Upadhyaya, P.; Dinh, T.T.; Bonilla, L.E.; Hecht, S.S. Lactols in hydrolysates of DNA treated with α-acetoxy-N-nitrosopyrrolidine or crotonaldehyde. Chem. Res. Toxicol. 1998, 11, 1567–1573. [Google Scholar] [CrossRef]
- Autrup, H.; Stoner, G.D. Metabolism of N-nitrosamines by cultured human and rat esophagus. Cancer Res. 1982, 42, 1307–1311. [Google Scholar]
- Autrup, H.; Harris, C.C.; Trump, B.F. Metabolism of acyclic and cyclic N-nitrosamines by cultured human colon. Proc. Soc. Exp. Biol. Med. 1978, 159, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Autrup, H.; Grafstrom, R.C.; Christensen, B.; Kieler, J. Metabolism of chemical carcinogens by cultured human and rat bladder epithelial cells. Carcinogenesis 1981, 2, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Hecht, S.S.; Chen, C.H.; McCoy, G.D.; Hoffmann, D.; Domellof, L. α-hydroxylation of N-nitrosopyrrolidine and N′-nitrosonornicotine by human liver microsomes. Cancer Lett. 1979, 8, 35–41. [Google Scholar] [CrossRef]
- Hecker, L.I. In vitro metabolism of N-nitrosopyrrolidine by rat lung subcellular fractions: Alpha-hydroxylation in a non-target tissue. Cell Biol. Int. 1980, 30, 57–65. [Google Scholar] [CrossRef]
- McCoy, G.D.; Chen, C.H.; Hecht, S.S.; McCoy, E.C. Enhanced metabolism and mutagenesis of nitrosopyrrolidine in liver fractions isolated from chronic ethanol-consuming hamsters. Cancer Res. 1979, 39, 793–796. [Google Scholar] [PubMed]
- McCoy, G.D.; Hecht, S.S.; McCoy, E.C. Comparison of microsomal inducer pretreatment on the in vitro alpha-hydroxylation and mutagenicity of N-Nitrosopyrrolidine in rat and hamster liver. Environ. Mutagen. 1982, 4, 221–230. [Google Scholar] [CrossRef]
- Hunt, E.J.; Shank, R.C. Evidence for DNA adducts in rat liver after adminstration of N-nitrosopyrrolidine. Biochem. Biophys. Res. Commun. 1982, 104, 1343–1348. [Google Scholar] [CrossRef]
- Chung, F.L.; Wang, M.Y.; Hecht, S.S. Detection of exocyclic guanine adducts in hydrolysates of hepatic DNA of rats treated with N-nitrosopyrrolidine and in calf thymus DNA reacted with alpha-acetoxy-N-nitrosopyrrolidine. Cancer Res. 1989, 49, 2034–2041. [Google Scholar]
- Wang, M.; Chung, F.L.; Hecht, S.S. Formation of acyclic and cyclic guanine adducts in DNA reacted with alpha-acetoxy-N-nitrosopyrrolidine. Chem. Res. Toxicol. 1989, 2, 423–428. [Google Scholar] [CrossRef]
- Wang, M.; Nishikawa, A.; Chung, F.L. Differential effects of thiols on DNA modifications via alkylation and Michael addition by α-acetoxy-N-nitrosopyrrolidine. Chem. Res. Toxicol. 1992, 5, 528–531. [Google Scholar] [CrossRef]
- Loureiro, A.P.; Zhang, W.; Kassie, F.; Zhang, S.; Villalta, P.W.; Wang, M.; Hecht, S.S. Mass spectrometric analysis of a cyclic 7,8-butanoguanine adduct of N-nitrosopyrrolidine: Comparison to other N-nitrosopyrrolidine adducts in rat hepatic DNA. Chem. Res. Toxicol. 2009, 22, 1728–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, E.J.; Shank, R.C. Formation and persistence of a DNA adduct in rodents treated with N-nitrosopyrrolidine. Carcinogenesis 1991, 12, 571–575. [Google Scholar] [CrossRef]
- Wang, M.; Hecht, S.S. A cyclic N7,C-8 guanine adduct of N-nitrosopyrrolidine (NPYR): Formation in nucleic acids and excretion in the urine of NPYR-treated rats. Chem. Res. Toxicol. 1997, 10, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chung, F.L.; Hecht, S.S. Formation of 7-(4-oxobutyl)guanine in hepatic DNA of rats treated with N-nitrosopyrrolidine. Carcinogenesis 1992, 13, 1909–1911. [Google Scholar] [CrossRef] [PubMed]
- Chung, F.L.; Young, R.; Hecht, S.S. Detection of cyclic 1,N2-propanodeoxyguanosine adducts in DNA of rats treated with N-nitrosopyrrolidine and mice treated with crotonaldehyde. Carcinogenesis 1989, 10, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Lao, Y.; Cheng, G.; Shi, Y.; Villalta, P.W.; Nishikawa, A.; Hecht, S.S. Analysis of adducts in hepatic DNA of rats treated with N-nitrosopyrrolidine. Chem. Res. Toxicol. 2007, 20, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Young-Sciame, R.; Chung, F.L.; Hecht, S.S. Formation of N2-tetrahydrofuranyl and N2-tetrahydropyranyl adducts in the reactions of α-acetoxy-N-nitrosopyrrolidine and α-acetoxy-N-nitrosopiperidine with DNA. Chem. Res. Toxicol. 1995, 8, 617–624. [Google Scholar] [CrossRef]
- Young-Sciame, R.; Wang, M.; Chung, F.L.; Hecht, S.S. Reactions of α-acetoxy-N-nitrosopyrrolidine and α-acetoxy-N-nitrosopiperidine with deoxyguanosine: Formation of N2-tetrahydrofuranyl and N2-tetrahydropyranyl adducts. Chem. Res. Toxicol. 1995, 8, 607–616. [Google Scholar] [CrossRef]
- Wang, M.; Lao, Y.; Cheng, G.; Shi, Y.; Villalta, P.W.; Hecht, S.S. Identification of adducts formed in the reaction of α-acetoxy-N-nitrosopyrrolidine with deoxyribonucleosides and DNA. Chem. Res. Toxicol. 2007, 20, 625–633. [Google Scholar] [CrossRef] [Green Version]
- Hermida, S.A.; Possari, E.P.; Souza, D.B.; de Arruda Campos, I.P.; Gomes, O.F.; Di Mascio, P.; Medeiros, M.H.; Loureiro, A.P. 2′-Deoxyguanosine, 2′-deoxycytidine, and 2′-deoxyadenosine adducts resulting from the reaction of tetrahydrofuran with DNA bases. Chem. Res. Toxicol. 2006, 19, 927–936. [Google Scholar] [CrossRef]
- Chung, F.L.; Hecht, S.S. Formation of cyclic 1,N2-adducts by reaction of deoxyguanosine with alpha-acetoxy-N-nitrosopyrrolidine, 4-(carbethoxynitrosamino)butanal, or crotonaldehyde. Cancer Res. 1983, 43, 1230–1235. [Google Scholar] [PubMed]
- Chung, F.L.; Roy, K.R.; Hecht, S.S. A study of chemical carcinogenesis. 104. A study of reactions of α,ß-unsaturated carbonyl-compounds with deoxyguanosine. J. Org. Chem. 1988, 53, 14–17. [Google Scholar] [CrossRef]
- Wang, M.; McIntee, E.J.; Shi, Y.; Cheng, G.; Upadhyaya, P.; Villalta, P.W.; Hecht, S.S. Reactions of α-acetoxy-N-nitrosopyrrolidine with deoxyguanosine and DNA. Chem. Res. Toxicol. 2001, 14, 1435–1445. [Google Scholar] [CrossRef] [PubMed]
- Chahoua, L.; Cai, H.; Fishbein, J.C. Cyclic α-acetoxynitrosamines: Mechanisms of decomposition and stability of α-hydroxynitrosamine and nitrosiminium ion reactive intermediates. J. Am. Chem. Soc. 1999, 121, 5161–5169. [Google Scholar] [CrossRef]
- Elespuru, R.K.; Lijinsky, W. Mutagenicity of cyclic nitrosamines in Escherichia coli following activation with rat liver microsomes. Cancer Res. 1976, 36 Pt 1, 4099–4101. [Google Scholar]
- Gomez, R.F.; Johnston, M.; Sinskey, A.J. Activation of nitrosomorpholine and nitrosopyrrolidine to bacterial mutagens. Mutat. Res. 1974, 24, 5–7. [Google Scholar] [CrossRef]
- Wang, H.T.; Zhang, S.; Hu, Y.; Tang, M.S. Mutagenicity and sequence specificity of acrolein-DNA adducts. Chem. Res. Toxicol. 2009, 22, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.S.; Wang, H.T.; Hu, Y.; Chen, W.S.; Akao, M.; Feng, Z.; Hu, W. Acrolein induced DNA damage, mutagenicity and effect on DNA repair. Mol. Nutr. Food. Res. 2011, 55, 1291–1300. [Google Scholar] [CrossRef] [Green Version]
- Minko, I.G.; Kozekov, I.D.; Harris, T.M.; Rizzo, C.J.; Lloyd, R.S.; Stone, M.P. Chemistry and biology of DNA containing 1,N2-deoxyguanosine adducts of the α,ß-unsaturated aldehydes acrolein, crotonaldehyde, and 4-hydroxynonenal. Chem. Res. Toxicol. 2009, 22, 759–778. [Google Scholar] [CrossRef] [Green Version]
- Nair, U.; Bartsch, H.; Nair, J. Lipid peroxidation-induced DNA damage in cancer-prone inflammatory diseases: A review of published adduct types and levels in humans. Free Radic. Biol. Med. 2007, 43, 1109–1120. [Google Scholar] [CrossRef]
- Chung, F.L.; Nath, R.G.; Nagao, M.; Nishikawa, A.; Zhou, G.D.; Randerath, K. Endogenous formation and significance of 1,N2-propanodeoxyguanosine adducts. Mutat. Res. 1999, 424, 71–81. [Google Scholar] [CrossRef]
- Zhang, S.; Villalta, P.W.; Wang, M.; Hecht, S.S. Analysis of crotonaldehyde- and acetaldehyde-derived 1,N2-propanodeoxyguanosine adducts in DNA from human tissues using liquid chromatography electrospray ionization tandem mass spectrometry. Chem. Res. Toxicol. 2006, 19, 1386–1392. [Google Scholar] [CrossRef] [Green Version]
- Chung, F.L.; Chen, H.J.; Nath, R.G. Lipid peroxidation as a potential endogenous source for the formation of exocyclic DNA adducts. Carcinogenesis 1996, 17, 2105–2111. [Google Scholar] [CrossRef] [PubMed]
- De Mey, E.; De Klerck, K.; De Maere, H.; Dewulf, L.; Derdelinckx, G.; Peeters, M.C.; Fraeye, I.; Vander Heyden, Y.; Paelinck, H. The occurrence of N-nitrosamines, residual nitrite and biogenic amines in commercial dry fermented sausages and evaluation of their occasional relation. Meat Sci. 2014, 96 Pt A, 821–828. [Google Scholar] [CrossRef]
- De Mey, E.; De Maere, H.; Goemaere, O.; Steen, L.; Peeters, M.C.; Derdelinckx, G.; Paelinck, H.; Fraeye, I. Evaluation of N-nitrosopiperidine formation from biogenic amines during the production of dry fermented sausages. Food Bioproc. Technol. 2013, 7, 1269–1280. [Google Scholar] [CrossRef]
- Gilbert, P.J.; Rollmann, B.; Rondelet, J.; Mercier, M.; Poncelet, F. Mutagenicity and alpha-hydroxylation of N-nitrosopyrrolidine and N-nitrosopiperidine: A possible correlation. Toxicology 1981, 22, 345–352. [Google Scholar] [CrossRef]
- Ayrton, A.D.; Smith, J.N.; Ioannides, C. Bioactivation of N-nitrosopiperidine to mutagens: Role of hepatic cytochrome P-450 proteins and contribution of cytosolic fraction. Carcinogenesis 1987, 8, 1691–1695. [Google Scholar] [CrossRef]
- Lai, D.Y.; Arcos, J.C.; Argus, M.F. Factors influencing the microsome- and mitochondria-catalyzed in vitro binding of diethylnitrosamine and N-nitrosopiperidine to deoxyribonucleic acid. Biochem. Pharmacol. 1979, 28, 3545–3550. [Google Scholar] [CrossRef]
- Wong, H.L.; Murphy, S.E.; Hecht, S.S. Preferential metabolic activation of N-nitrosopiperidine as compared to its structural homologue N-nitrosopyrrolidine by rat nasal mucosal microsomes. Chem. Res. Toxicol. 2003, 16, 1298–1305. [Google Scholar] [CrossRef]
- Wong, H.L.; Murphy, S.E.; Wang, M.; Hecht, S.S. Comparative metabolism of N-nitrosopiperidine and N-nitrosopyrrolidine by rat liver and esophageal microsomes and cytochrome P450 2A3. Carcinogenesis 2003, 24, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Arimoto, S.; Shimada, H.; Ukawa, S.; Mochizuki, M.; Hayatsu, H. Formation of directly mutagenic alpha-hydroxy-N-nitrosopiperidine phosphate ester by near-ultraviolet irradiation of N-nitrosopiperidine in phosphate buffer. Biochem. Biophys. Res. Commun. 1989, 162, 1140–1146. [Google Scholar] [CrossRef] [Green Version]
- Hecht, S.S.; Young-Sciame, R.; Chung, F.L. Reaction of α-acetoxy-N-nitrosopiperidine with deoxyguanosine: Oxygen-dependent formation of 4-oxo-2-pentenal and a 1,N2-ethenodeoxyguanosine adduct. Chem. Res. Toxicol. 1992, 5, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.H.; Park, K.K.; Archer, M.C. Alpha-hydroxylation in the metabolism of N-nitrosopiperidine by rat liver microsomes: Formation of 5-hydroxypentanal. Res. Commun. Chem. Pathol. Pharmacol. 1978, 19, 201–211. [Google Scholar] [PubMed]
- Nakamura, M.; Horiguchi, Y.; Kawabata, T. Effect of Ascorbic Acid and Some Reducing Agents on N-Nitrosopiperidine Metabolism by Liver Microsomes. In IARC Scientific Publication No. 57: N-Nitroso Compounds: Occurrence, Biological Effects and Relevance to Human Cancer; O’Neill, I., von Borstel, R., Miller, C., Long, J., Bartsch, H., Eds.; IARC: Lyon, France, 1984; pp. 547–552. [Google Scholar]
- Ravindranath, V.; Burka, L.T.; Boyd, M.R. Reactive metabolites from the bioactivation of toxic methylfurans. Science 1984, 224, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Challis, B.C. Oxidation of N-nitrosopiperidine in the Udenfriend model system and its metabolism by rat-liver microsomes. Biochem. Pharmacol. 1975, 24, 621–626. [Google Scholar] [CrossRef]
- Challis, B.C.; Rayman, M.P. Potential alkylating agents from the oxidation of carcinogenic cyclic N-nitrosamines. Br. J. Cancer 1973, 28, 84. [Google Scholar] [CrossRef] [Green Version]
- Lijinsky, W.; Taylor, H.W. Carcinogenicity of methylated nitrosopiperidines. Int. J. Cancer 1975, 16, 318–322. [Google Scholar] [CrossRef]
- Lijinsky, W.; Taylor, H.W. Tumorigenesis by oxygenated nitrosopiperidines in rats. J. Natl. Cancer Inst. 1975, 55, 705–708. [Google Scholar] [CrossRef]
- Liu, Z.; Young-Sciame, R.; Hecht, S.S. Liquid chromatography-electrospray ionization mass spectrometric detection of an ethenodeoxyguanosine adduct and its hemiaminal precursors in DNA reacted with α-acetoxy-N-nitrosopiperidine and cis-4-oxo-2-pentenal. Chem. Res. Toxicol. 1996, 9, 774–780. [Google Scholar] [CrossRef]
- Totsuka, Y.; Lin, Y.; He, Y.; Ishino, K.; Sato, H.; Kato, M.; Nagai, M.; Elzawahry, A.; Totoki, Y.; Nakamura, H.; et al. DNA adductome analysis identifies N-nitrosopiperidine involved in the etiology of esophageal cancer in Cixian, China. Chem. Res. Toxicol. 2019, 32, 1515–1527. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, S.; Yokokawa, Y.; Hou, J.; Zhang, X.L.; Li, X.P.; Li, S.S.; Li, X.X.; Zhu, D.C.; Kamijima, M.; Yamanoshita, O.; et al. Environmental exposure and p53 mutations in esophageal cancer patients in areas of low and high incidence of esophageal cancer in China. Tohoku J. Exp. Med. 2005, 207, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiegelhalder, B.; Preussmann, R. Contamination of toiletries and cosmetic products with volatile and nonvolatile N-nitroso carcinogens. J. Cancer Res. Clin. Oncol. 1984, 108, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Sanches Filho, P.J.; Rios, A.; Valcárcel, M.; Zanin, K.D.; Caramão, E.B. Determination of nitrosamines in preserved sausages by solid-phase extraction-micellar electrokinetic chromatography. J. Chromatogr. A 2003, 985, 503–512. [Google Scholar] [CrossRef]
- Charrois, J.W.; Arend, M.W.; Froese, K.L.; Hrudey, S.E. Detecting N-nitrosamines in drinking water at nanogram per liter levels using ammonia positive chemical ionization. Environ. Sci. Technol. 2004, 38, 4835–4841. [Google Scholar] [CrossRef]
- Reh, B.D.; Fajen, J.M. Worker exposures to nitrosamines in a rubber vehicle sealing plant. Am. Ind. Hyg. Assoc. J. 1996, 57, 918–923. [Google Scholar] [CrossRef]
- Fajen, J.M.; Carson, G.A.; Rounbehler, D.P.; Fan, T.Y.; Vita, R.; Goff, U.E.; Wolf, M.H.; Edwards, G.S.; Fine, D.H.; Reinhold, V.; et al. N-nitrosamines in the rubber and tire industry. Science 1979, 205, 1262–1264. [Google Scholar] [CrossRef]
- Oury, B.; Limasset, J.C.; Protois, J.C. Assessment of exposure to carcinogenic N-nitrosamines in the rubber industry. Int. Arch. Occup. Environ. Health 1997, 70, 261–271. [Google Scholar] [CrossRef]
- Monarca, S.; Feretti, D.; Zanardini, A.; Moretti, M.; Villarini, M.; Spiegelhalder, B.; Zerbini, I.; Gelatti, U.; Lebbolo, E. Monitoring airborne genotoxicants in the rubber industry using genotoxicity tests and chemical analyses. Mutat. Res. 2001, 490, 159–169. [Google Scholar] [CrossRef]
- Mirvish, S.S. Formation of N-nitroso compounds: Chemistry, kinetics, and in vivo occurrence. Toxicol. Appl. Pharmacol. 1975, 31, 325–351. [Google Scholar] [CrossRef]
- Hecht, S.S.; Morrison, J.B. A sensitive method for detecting in vivo formation of N-nitrosomorpholine and its application to rats given low doses of morpholine and sodium nitrite. Cancer Res. 1984, 44, 2873–2877. [Google Scholar]
- Van Stee, E.W.; Sloane, R.A.; Simmons, J.E.; Moorman, M.P.; Brunnemann, K.D. Endogenous formation of N-nitrosomorpholine in mice from 15NO2 by inhalation and morpholine by gavage. Carcinogenesis 1995, 16, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Singer, G.M.; Lijinsky, W. Naturally occurring nitrosatable compounds. I. Secondary amines in foodstuffs. J. Agric. Food Chem. 1976, 24, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Börzsönyi, M.; Surján, A.; Pintér, A.; Török, G.; Csik, M.; Tamás, J.; Fetter, J.; Ferencz, A. Endogenous Formation of N-Nitrosomorpholine from a New Fungicide. In IARC Scientific Publication No. 31: N-Nitroso Compounds: Analysis, Formation and Occurrence; Walker, E., Griciute, L., Castegnaro, M., Borzsonyi, M., Eds.; IARC: Lyon, France, 1980; pp. 677–683. [Google Scholar]
- Brunnemann, K.D.; Scott, J.C.; Hoffmann, D. N-Nitrosomorpholine and other volatile N-nitrosamines in snuff tobacco. Carcinogenesis 1982, 3, 693–696. [Google Scholar] [CrossRef] [PubMed]
- McGlothlin, J.D.; Wilcox, T.C.; Fajen, J.M.; Edwards, G.S. A Health Hazard Evaluation of Nitrosamines in a Tire Manufacturing Plant. In Chemical Hazards in the Workplace; American Chemical Society: Washington, DC, USA, 1981; Volume 149, pp. 283–299. [Google Scholar]
- Spiegelhalder, B.; Preussmann, R. Occupational nitrosamine exposure. 1. Rubber and tyre industry. Carcinogenesis 1983, 4, 1147–1152. [Google Scholar] [CrossRef]
- Kakizoe, T.; Wang, T.T.; Eng, V.W.; Furrer, R.; Dion, P.; Bruce, W.R. Volatile N-nitrosamines in the urine of normal donors and of bladder cancer patients. Cancer Res. 1979, 39, 829–832. [Google Scholar]
- van Maanen, J.M.; Pachen, D.M.; Dallinga, J.W.; Kleinjans, J.C. Formation of nitrosamines during consumption of nitrate- and amine-rich foods, and the influence of the use of mouthwashes. Cancer Detect. Prev. 1998, 22, 204–212. [Google Scholar] [CrossRef]
- Dallinga, J.W.; Pachen, D.M.; Lousberg, A.H.; van Geel, J.A.; Houben, G.M.; Stockbrügger, R.W.; van Maanen, J.M.; Kleinjans, J.C. Volatile N-nitrosamines in gastric juice of patients with various conditions of the gastrointestinal tract determined by gas chromatography-mass spectrometry and related to intragastric pH and nitrate and nitrite levels. Cancer Lett. 1998, 124, 119–125. [Google Scholar] [CrossRef]
- Lijinsky, W.; Kovatch, R.M.; Knutsen, G.L. Carcinogenesis by nitrosomorpholines, nitrosooxazolidines and nitrosoazetidine given by gavage to Syrian golden hamsters. Carcinogenesis 1984, 5, 875–878. [Google Scholar] [CrossRef]
- Rao, M.S.; Scarpelli, D.G.; Lijinsky, W. Carcinogenesis in Syrian hamsters by N-nitroso-2,6-dimethylmorpholine, its cis and trans isomers, and the effect of deuterium labeling. Carcinogenesis 1981, 2, 731–735. [Google Scholar] [CrossRef]
- Palladino, G.F.; Chung, F.L.; Hecht, S.S. 3-(2-Deoxy-ß-D-erythropentofuranosyl)-6,7-dihydro-6,7-dihydroxyimidazo[1,2-a]purin-9(3H)-one, a major deoxyguanosine adduct formed from a novel diazo hydroxide product of α-hydroxylation of the carcinogen N-nitrosomorpholine. J. Am. Chem. Soc. 1986, 108, 6066–6068. [Google Scholar] [CrossRef]
- Stewart, B.W.; Swann, P.F.; Holsman, J.W.; Magee, P.N. Cellular injury and carcinogenesis. Evidence for the alkylation of rat liver nucleic acids in vivo by N-nitrosomorpholine. Z. Krebsforsch. Klin. Onkol. Cancer Res. Clin. Oncol. 1974, 82, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hecht, S.S.; Young, R. Metabolic α-hydroxylation of N-nitrosomorpholine and 3,3,5,5-tetradeutero-N-nitrosomorpholine in the F344 rat. Cancer Res. 1981, 41 Pt 1, 5039–5043. [Google Scholar] [PubMed]
- Hecht, S.S.; Morrison, J.B.; Young, R. N-Nitroso(2-hydroxyethyl)glycine, a urinary metabolite of N,N-dinitrosopiperazine with potential utility as a monitor for its formation in vivo from piperazine. Carcinogenesis 1984, 5, 979–981. [Google Scholar] [CrossRef] [PubMed]
- Manson, D.; Cox, P.J.; Jarman, M. Metabolism of N-nitrosomorpholine by the rat in vivo and by rat liver microsomes and its oxidation by the Fenton system. Cell Biol. Int. 1978, 20, 341–354. [Google Scholar] [CrossRef]
- Arimoto-Kobayashi, S.; Hayatsu, H. Formation of direct-acting mutagens from mixtures of N-nitrosomorpholine and carboxylates by UVA irradiation. Environ. Mol. Mutagen. 1998, 31, 163–168. [Google Scholar] [CrossRef]
- Chung, F.L.; Hecht, S.S. Formation of the cyclic 1,N2-glyoxal-deoxyguanosine adduct upon reaction of N-nitroso-2-hydroxymorpholine with deoxyguanosine. Carcinogenesis 1985, 6, 1671–1673. [Google Scholar] [CrossRef] [PubMed]
- Zink, C.N.; Kim, H.J.; Fishbein, J.C. Synthesis and aqueous chemistry of α-acetoxy-N-nitrosomorpholine: Reactive intermediates and products. J. Org. Chem. 2006, 71, 202–209. [Google Scholar] [CrossRef]
- Zink, C.N.; Soissons, N.; Fishbein, J.C. Products of the direct reaction of the diazonium ion of a metabolite of the carcinogen N-nitrosomorpholine with purines of nucleosides and DNA. Chem. Res. Toxicol. 2010, 23, 1223–1233. [Google Scholar] [CrossRef]
- Kim, H.J.; Fishbein, J.C. Reexamination of the aqueous chemistry of N-nitroso-3-hydroxymorpholine, a metabolite of the carcinogen N-nitrosomorpholine. Chem. Res. Toxicol. 2003, 16, 715–720. [Google Scholar] [CrossRef]
- Koissi, N.; Shah, N.H.; Ginevan, B.; Eck, W.S.; Roebuck, B.D.; Fishbein, J.C. Lactone metabolite common to the carcinogens dioxane, diethylene glycol, and N-nitrosomorpholine: Aqueous chemistry and failure to mediate liver carcinogenesis in the F344 rat. Chem. Res. Toxicol. 2012, 25, 1022–1028. [Google Scholar] [CrossRef] [Green Version]
- Eisenbrand, G.; Denkel, E.; Pool, B. Alcoholdehydrogenase as an activating enzyme for N-nitrosodiethanolamine (NDELA): In vitro activation of NDELA to a potent mutagen in Salmonella typhimurium. J. Cancer Res. Clin. Oncol. 1984, 108, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Denkel, E.; Pool, B.L.; Schlehofer, J.R.; Eisenbrand, G. Biological activity of N-nitrosodiethanolamine and of potential metabolites which may arise after activation by alcohol dehydrogenase in Salmonella typhimurium, in mammalian cells, and in vivo. J. Cancer Res. Clin. Oncol. 1986, 111, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Koissi, N.; Fishbein, J.C. Trapping of a cross-link formed by a major purine adduct of a metabolite of the carcinogen N-nitrosomorpholine by inorganic and biological reductants. Chem. Res. Toxicol. 2013, 26, 732–740. [Google Scholar] [CrossRef] [Green Version]
- Hecht, S.S. Approaches to cancer prevention based on an understanding of N-nitrosamine carcinogenesis. Proc. Soc. Exp. Biol. Med. 1997, 216, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Ricker, K.; Tsai, F.C.; Hsieh, C.J.; Osborne, G.; Sun, M.; Marder, M.E.; Elmore, S.; Schmitz, R.; Sandy, M.S. Estimated cancer risks associated with nitrosamine contamination in commonly used medications. Int. J. Environ. Res. Public Health 2021, 18, 9465. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Balbo, S.; Turesky, R.J.; Villalta, P.W. DNA adductomics. Chem. Res. Toxicol. 2014, 27, 356–366. [Google Scholar] [CrossRef]
Figure 1.
Chemical structures of carcinogenic N-nitrosamines to which humans are commonly exposed. NDMA, N-nitrosodimethylamine; NMEA, N-nitrosomethylethylamine; NSAR, N-nitrososarcosine; NDEA, N-nitrosodiethylamine; NDPA, N-nitrosodi-n-propylamine; NDELA, N-nitrosodiethanolamine; NDBA, N-nitrosodi-n-butylamine; NPYR, N-nitrosopyrrolidine; NPIP, N-nitrosopiperidine; NMOR, N-nitrosomorpholine.
Figure 1.
Chemical structures of carcinogenic N-nitrosamines to which humans are commonly exposed. NDMA, N-nitrosodimethylamine; NMEA, N-nitrosomethylethylamine; NSAR, N-nitrososarcosine; NDEA, N-nitrosodiethylamine; NDPA, N-nitrosodi-n-propylamine; NDELA, N-nitrosodiethanolamine; NDBA, N-nitrosodi-n-butylamine; NPYR, N-nitrosopyrrolidine; NPIP, N-nitrosopiperidine; NMOR, N-nitrosomorpholine.

Figure 2.
Participants in the conference on N-Nitroso Compounds: Occurrence, Biological Effects and Relevance to Human Cancer, sponsored by the International Agency for Research on Cancer (IARC), Agriculture Canada, and the Alberta Heritage Foundation for Medical Research in Banff, Canada, 5–9 September 1983. (Reprinted with permission of the IARC).
Figure 2.
Participants in the conference on N-Nitroso Compounds: Occurrence, Biological Effects and Relevance to Human Cancer, sponsored by the International Agency for Research on Cancer (IARC), Agriculture Canada, and the Alberta Heritage Foundation for Medical Research in Banff, Canada, 5–9 September 1983. (Reprinted with permission of the IARC).

Scheme 1.
NDMA metabolism by α-hydroxylation and denitrosation.

Figure 3.
Structures of methyl DNA base and phosphate adducts.

Figure 4.
Structures of isotope-labeled NMEAs.

Scheme 2.
Mechanisms of NMEA metabolic activation.

Figure 5.
Structures of ethyl and hydroxyethyl DNA adducts.

Figure 6.
Structures of agents that carboxymethylate DNA.

Scheme 3.
Mechanisms of NSAR metabolic activation.

Figure 7.
Structures of carboxymethyl DNA adducts.

Figure 8.
Chemical structures of N7-CM-Gua, N7-CE-Gua, and MNPA.

Scheme 4.
Mechanisms of NDEA metabolic activation.

Scheme 5.
Mechanisms of NDPA metabolic activation.

Figure 9.
Structures of NDPA analogs that form DNA adducts.

Figure 10.
Structures of propyl/hydroxypropyl and butyl/hydroxybutyl DNA adducts.

Scheme 6.
Mechanisms of NDELA metabolic activation.

Figure 11.
Structures of glyoxal and etheno DNA adducts.

Scheme 7.
Mechanisms of NDBA metabolic activation.

Scheme 8.
Mechanisms of NPYR metabolic activation.

Figure 12.
Structures of DNA adducts formed by NPYR metabolism.

Figure 13.
Chemical structures of other DNA adducts related to NPYR metabolism.

Scheme 9.
Mechanisms of NPIP metabolism.

Figure 14.
Structures of DNA adducts formed by NPIP metabolism.

Scheme 10.
Mechanisms of NMOR and NHMOR metabolism.

Figure 15.
Structures of DNA adducts formed by NMOR metabolism.

Scheme 11.
Proposed mechanism of N1,N2-glyoxal-dGuo formation by NHMOR.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Levels of DNA adducts in the tissues of laboratory animals exposed to NPYR.
| Animal Species | Administration Pathway | Exposure Amount | Exposure Time | Target Tissue | DNA Adducts (μmol/mol dGuo unless otherwise Noted) Formed by NPYR | Ref. | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N7,8-Butanoguanine | N7-(4-Oxobutyl)-dGuo | N1,N2-Propano-Gua a | N6-(4-HOB)-dAdo b | N2-(4-HOB)-dGuo b | O2-(4-HOB)-Thd b | O4-(4-HOB)-Thd b | ||||||
| Male F344 rats | Single dose by gavage | 3.1 mg/kg (8 mCi) [3,4-3H]NPYR | 16 h | Liver | 0.7 pmol/mg DNA | [244] | ||||||
| 6.7 mg/kg (15 mCi) [3,4-3H]NPYR | ND | |||||||||||
| 175 mg/kg NPYR | 320 pmol/mg DNA | |||||||||||
| 350 mg/kg NPYR | 390 pmol/mg DNA | |||||||||||
| 450 mg/kg NPYR | 24 h | Liver RNA | 2860 ± 332 | ND | [249] | |||||||
| Liver DNA | 1430 ± 56 | 603 | ||||||||||
| 46 mg/kg NPYR | 16 h | Liver | 952 ± 180 | 0.231 ± 0.078 | 0.013 ± 0.006 | 0.369 ± 0.134 | 1.065 ± 0.349 | [247] | ||||
| 92 mg/kg NPYR | 1742 ± 245 | 0.499 ± 0.091 | 0.008 ± 0.004 | 0.478 ± 0.114 | 2.049 ± 0.361 | |||||||
| 184 mg/kg NPYR | 3032 ± 855 | 0.905 ± 0.240 | 0.016 ± 0.008 | 0.951 ± 0.193 | 3.767 ± 0.840 | |||||||
| Single dose by i.p. injection | 450 mg/kg NPYR | 16 h | Liver | 1792 ± 87 | 643 ± 9 | ND | [250] | |||||
| In the drinking water | 6 mM NPYR | 14 days | Liver | 0.06 | [251] | |||||||
| 600 ppm NPYR | 1 week | 0.02 ± 0.01 | 3.41 ± 0.60 | 2.56 ± 0.84 | 2.28 ± 0.47 | [252] | ||||||
| 200 ppm NPYR | 4 weeks | 0.03 ± 0.01 | 4.84 ± 0.48 | 3.87 ± 0.97 | 3.83 ± 0.57 | |||||||
| 200 ppm NPYR | 13 weeks | 0.04 ± 0.02 | 5.39 ± 0.39 | 3.52 ± 0.43 | 5.05 ± 1.68 | |||||||
| Male Sprague-Dawley rats | Single dose by intubation | 14–900 mg/kg NPYR | 12 h | Liver | Dose-dependently formed; 0.1–3 equivalents of N7-Me-Gua | [243] | ||||||
| 0.63 mg [14C]NPYR/ 1 rat (207 g) | 8- or 42.6-fold of N7-Me-Gua radioactivity | |||||||||||
| Intragastric administration | 900 mg/kg NPYR | 12 h | Liver | 3983 | [248] | |||||||
| Kidney | 159 | |||||||||||
| Lung | 159 | |||||||||||
| Male Swiss Webster mice | Intragastric administration | 900 mg/kg NPYR | 24 h | Liver | 9987 | [248] | ||||||
| Kidney | 699 | |||||||||||
| Lung | 499 | |||||||||||
| Male Syrian golden hamsters | Intragastric administration | 900 mg/kg NPYR | 24 h | Liver | 9000 | [248] | ||||||
| Kidney | 900 | |||||||||||
| Lung | 1170 | |||||||||||
a The levels reported here contain the two diastereomers (6S,8S)- and (6R,8R)-N1,N2-propano-Gua. b The 4-hydroxybutyl (4-HOB) DNA adducts were the reduction products of their tetrahydrofuranyl precursors. They were more stable for quantitation than their precursors.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Y.; Hecht, S.S. Metabolic Activation and DNA Interactions of Carcinogenic N-Nitrosamines to Which Humans Are Commonly Exposed. Int. J. Mol. Sci. 2022, 23, 4559. https://doi.org/10.3390/ijms23094559
AMA Style
Li Y, Hecht SS. Metabolic Activation and DNA Interactions of Carcinogenic N-Nitrosamines to Which Humans Are Commonly Exposed. International Journal of Molecular Sciences. 2022; 23(9):4559. https://doi.org/10.3390/ijms23094559
Chicago/Turabian StyleLi, Yupeng, and Stephen S. Hecht. 2022. "Metabolic Activation and DNA Interactions of Carcinogenic N-Nitrosamines to Which Humans Are Commonly Exposed" International Journal of Molecular Sciences 23, no. 9: 4559. https://doi.org/10.3390/ijms23094559
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.