
Non-Coding RNAs Regulate Spontaneous Abortion: A Global Network and System Perspective
by
 , , and
, , and
Jianyu Gan
1, 
Ting Gu
1,
Huaqiang Yang
1,
Zheng Ao
2,
Gengyuan Cai
1,3,4,
Linjun Hong
1,3,4,* and
Zhenfang Wu
1,3,4,* 1
National Engineering Research Center for Breeding Swine Industry, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
2
Key Laboratory of Animal Genetics, Breeding and Reproduction in the Plateau Mountainous Region, Ministry of Education, College of Animal Science, Guizhou University, Guiyang 550025, China
3
Guangdong Provincial Key Laboratory of Agro-Animal Genomics and Molecular Breeding, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
4
State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources, Guangzhou 510642, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(8), 4214; https://doi.org/10.3390/ijms23084214
Submission received: 16 March 2022
/
Revised: 6 April 2022
/
Accepted: 8 April 2022
/
Published: 11 April 2022
(This article belongs to the Special Issue Non-coding RNAs in Pathogens and Associated Diseases)
Abstract
:Spontaneous abortion is a common pregnancy complication that negatively impacts women’s health and commercial pig production. It has been demonstrated that non-coding RNA (ncRNA) is involved in SA by affecting cell proliferation, invasion, apoptosis, epithelial-mesenchymal transformation (EMT), migration, and immune response. Over the last decade, research on ncRNAs in SA has primarily concentrated on micro RNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs). In this review, we discuss recent ncRNA studies focused on the function and mechanism of miRNAs, lncRNAs, and circRNAs in regulating SA. Meanwhile, we suggest that a ceRNA regulatory network exists in the onset and development of SA. A deeper understanding of this network will accelerate the process of the quest for potential RNA markers for SA diagnosis and treatment.
Keywords:
spontaneous abortion; miRNA; lncRNA; circRNA; competing endogenous RNA; systematic network1. Introduction
Spontaneous abortion (SA) is clinically defined as the loss of pregnancy before 24 weeks of gestation without any external intervention [1,2]. Two or more consecutive SA in the first 20 weeks are defined as recurrent spontaneous abortions (RSA) [3]. SA not only has a negative impact on women’s health and reproduction but also poses a substantial risk to global commercial pig production (about 20–30% of pig embryos develop SA during 12–30 days of pregnancy) [4]. It is estimated that approximately 5% of women of childbearing age have RSA [5], which causes psychological distress and has a negative impact on their welfare. Understanding the pathogenesis of SA and its treatment are critical for both human reproductive health and commercial porcine production. It has recently been determined that endocrine, immune, environmental, infectious, and genetic factors all play a role in SA development [6,7]. Nevertheless, about half of patients cannot be identified as having the pathogenesis of SA (defined as unexplained recurrent spontaneous abortion (URSA)), posing significant challenges in treating SA. URSA remains a substantial challenge in obstetrics due to a lack of safe and effective treatment options and reliable early detection tools [8]. Hence, studies and research that can explain the mechanism of SA are required.
Non-coding RNAs (ncRNAs) are RNAs that do not encode proteins and include but are not limited to small nuclear RNA (snRNA), small nucleolar RNA (snoRNA), ribosomal RNA (rRNA), transfer RNA (tRNA), PIWI-interacting RNA (piRNA), circular RNA (circRNA), long non-coding RNA (lncRNA), and microRNA (miRNA). ncRNAs play essential biological roles even though they do not encode proteins. For instance, they can regulate a wide range of paramount life activities by participating in chromosomal remodeling, gene transcription, and post-transcriptional modification [9]. Over the last decade, research on ncRNAs in SA has primarily concentrated on miRNAs, lncRNAs, and circRNAs.
Herein, we discuss recent ncRNA studies focused on the function and mechanism of miRNAs, lncRNAs, and circRNAs in regulating SA. Since the competing endogenous RNA (ceRNA) network hypothesis was proposed in 2011, researchers have become increasingly aware of the possible potential application of ceRNA network regulation in diagnosing and treating a wide range of diseases [10]. Based on the findings of these studies on pregnant diseases [11,12,13,14,15], we hypothesize that there may be a broad and complex ceRNA regulatory network that will induce SA when network regulation is disrupted. We anticipate that our viewpoint will be helpful as a reference and will aid future research into the pathogenesis of SA and the search for potential RNA markers for diagnosis and treatment.
2. Pathogenesis of SA
There are different pathogenesis of SA, including genetic factors [16], physiological factors [17], immune factors [18], and environmental factors [19] (Figure 1). Genetic factors are considered the main contributors to sporadic SA and RSA due to fetal chromosome abnormalities accounting for 50–60% of cases [2]. However, about 40–50% of RSA patients demonstrate normal karyotypes, and the etiologies are still unclear, which is thought to have URSA [20,21,22]. In the early stage of normal pregnancy, the correct performance of the functions of placental trophoblast cells directly affects the survival of embryos. In humans and mice, cytotrophoblast cells differentiate into syncytiotrophoblast and extravillous trophoblast (EVT) cells after implantation [23]. Subsequently, EVT cells invade the maternal endometrium, fixing the placenta to the uterine wall and leading to the remodeling of the maternal spiral artery to provide nutrition for the developing fetus [23]. Any abnormal step in this process may lead to placenta-related pathological pregnancy, including SA [24].
In recent years, with the increasing research of ncRNAs in SA, researchers have a more thorough understanding of the mechanisms of ncRNAs regulating SA. Indeed, miRNAs [25,26], lncRNAs [27,28,29], and circRNAs [30,31] are all involved in the modulation of the pathogenesis of SA. Afterward, researchers also found some ncRNAs that can be regarded as markers of SA, which play a critical role in the diagnosis of SA. However, regarding how these ncRNAs participate in the regulation of SA, the specific mechanism is still unclear, resulting in great limitations in the use of these ncRNAs in clinical diagnosis, treatment, and porcine production.
3. miRNAs and Spontaneous Abortion
3.1. Overview of miRNAs
miRNAs is an endogenous, short, non-coding, and highly conserved calss of molecule that play a role in the mechanism of post-transcriptional gene expression by inhibiting the translation of protein-encoded genes or cutting target mRNAs [32,33]. The first miRNA, LIN-4, was found in Caenorhabditis elegans in 1993 [34]. Afterward, studies on miRNA have emerged continuously and even become a research hotspot. miRNAs can bind to the mRNAs of target downstream genes, affecting the stability and transcription of the target mRNAs. A special feature of miRNAs is that one kind of miRNA can regulate the expression of several kinds of mRNAs, while one kind of mRNA can also be targeted by multiple kinds of miRNAs, which means that a small number of miRNAs can regulate about 60% of the protein-coding genes [17,35]. Over 1000 kinds of miRNAs have been revealed to participate in modulating complex processes in vivo, such as cellular growth, differentiation, immune response, and tissue remodeling, and some of them play a critical role in multiple diseases states, including SA [36,37]. As a result, some miRNAs in the maternal circulatory system have been screened as good biomarkers for monitoring the progression of normal pregnancy and the existence of pregnant diseases, which can be used to prevent and treat some reproductive diseases [38].
3.2. miRNAs Affect SA by Regulating Apoptosis
In 1998, Kokawa et al. [39] first proposed that apoptosis balance disruption could be one of the primary causes of SA. Studies confirmed that some miRNAs could indeed regulate apoptosis by affecting different pathways, thus triggering or inhibiting SA. For instance, Liu et al. [40] discovered that miR-93 expression was increased in the chorionic villi of patients with RSA, and the expression of targeted BCL2-like 2 (BCL2L2) disrupted the balance of apoptosis, cell proliferation, migration, and invasion and promoted the occurrence of SA.
In addition, miR-19b and miR-494 can regulate the expression of phosphatase and tensin homolog (PTEN) in a synergistic manner, influencing SA occurrence [41]. PTEN is a well-known tumor suppressor gene that regulates apoptosis in various cells and is involved in the cell cycle, growth, migration, and death through the PI3K/AKT signaling pathway [42]. RSA occurrence can be effectively prevented with miR-19b upregulation and miR-494 downregulation, which inhibits cell apoptosis through the expression of their common target gene PTEN [41].
Furthermore, miRNA-365, an upstream regulator of MDM2/P53 expression, cell cycle progression, and apoptosis in trophoblast cells, was discovered to regulate trophoblast apoptosis through serum/glucocorticoid regulatory kinase 1 (SGK1) [43]. Since SGK1 is the direct target gene of miR-365, silencing SGK1 can cause trophoblast cell cycle arrest and apoptosis, and SGK1 overexpression can reduce the effect of miR-365 on apoptosis and MDM2/P53 expression [43]. Both these mechanisms have a direct bearing on SA. Furthermore, when Dong et al. [44] investigated the role of miRNA-199B-5p and SGK1 in pregnancy maintenance, they discovered that miR-199B-5p expression was significantly upregulated in the aborted decidua, while miR-199B-5p overexpression in human endometrial stromal cells and transgenic mice reduced SGK1 expression, and the in vitro and in vivo results were consistent and comparable. Therefore, miR-199b-5p, like miR-365, may control apoptosis by targeting SGK1. Another study revealed that by targeting poly (ADP- ribose) polymerase 1 (PARP1), miR-520 could facilitate trophoblast apoptosis induced by DNA damage, thereby contributing to the onset and progression of RSA [45].
3.3. miRNAs Are Key Regulators in the Physiological Processes of the Placenta
SA is directly linked to placental physiological processes of invasion, migration, EMT, and angiogenesis. When these processes are hampered and overburdened, uterine-placental dysfunction occurs, increasing the risk of SA [46]. The Wnt signaling pathway governs numerous cellular functions, including migration and invasion, and can be regulated by ubiquitin-specific peptidase 25 (USP25) [47]. Ding et al. [48] found that miR-27A-3p can regulate the expression of USP25 by binding to the 3′ untranslated region of USP25 in trophoblast cells, thus regulating the migration and invasion of EVT and trophoblast cells. EMT, an evolutionarily conserved developmental program, has been linked to carcinogenesis and confers metastatic properties on cancer cells. When miR-27A-3p was transfected with microRNA mimics or inhibitors, the expression of USP25 in RSA’s patients’ placental villi was down-regulated, and the processes of EMT, invasion, and migration were inhibited [48].
Furthermore, miR-16 can regulate placental angiogenesis and development by regulating vascular endothelial growth factor (VEGF) expression and participating in the pathogenesis of RSA [49]. VEGF plays a pivotal role in regulating the invasion and proliferation of trophoblast cells. When the VEGF signal pathway is inhibited, it may suppress trophoblast cell proliferation and invasion, resulting in SA [50,51]. The vascular endothelial growth factor receptor-2 (VEGFR2) is the receptor for VEGF and contributes to the proliferation, migration, and invasion of trophoblasts by binding to core proteins [52]. MiR-219a can inhibit the function of VEGFR2 and its downstream PI3K/AKT/NF-κB signal pathway by targeting the expression of VEGFR2, which inhibits the excessive proliferation and invasion of trophoblast cells by negative regulation [26]. Meanwhile, Zhang et al. [25] identified that miR-155-5p regulated SA by activating the NF-κB signaling pathway. MiR-155-5p overexpression can reduce the release of inflammatory cytokines such as IL-6, IFN-γ, TNF-α, and IL-10 by decidual stromal cells and inhibit the apoptosis of decidual stromal cells.
3.4. miRNAs Participate in the Regulation of Immunity
In early pregnancy, it is necessary to experience the processes of placental development and spiral artery remodeling, which require the participation of immune cells and immune molecules; thus, SA is largely related to the imbalance of the maternal immune system [53,54,55]. Natural killer (NK) cells and macrophages surround the spiral artery, while interstitial EVT cells are recruited by NK cells and macrophages to replace vascular endothelial cells in the spiral artery thus initiating the process of remodeling [55,56]. Considering the indispensable role of NK cells and macrophages, many scholars have studied miRNAs and their targets on NK cells and M1 macrophages in patients with SA.
In the NK cells of decidual tissue of patients with URSA, Li et al. [57] studied the expression levels of six kinds of miRNA in decidual NK cells of URSA patients and revealed that miR-34a-3p/5p, miR-141-3p/5p, and miR-24 might link to URSA. These miRNAs target P53, PTEN, and IFN-γ (IFNG), respectively, to regulate apoptosis, proliferation, and angiogenesis [58,59,60,61]. Considering the effects of P53, IFNG, and PTEN on placental villi, trophoblast invasion, migration, EMT, and apoptosis have been discussed previously, it is reasonable to deem that these miRNAs in NK cells of URSA patients play a fundamental role in regulating SA.
Moreover, Li et al. [62] used miRNA microarray to study the miRNAs map of NK cells in decidual tissues of patients with URSA. Once again, it was found that there was a close relationship between abnormal expression of miRNAs and URSA, and 50 differentially expressed miRNAs were identified to construct miRNA-Gene-Network, miRNA-Go-Network, and miRNA-Gene-TF-Network. In miRNA-Go-Network, the key miRNA is hsa-miR-5787, which inhibits cell growth by targeting eukaryotic translation initiation factor 5 (EIF5) [63]. From the results of GO analysis of down-regulated miRNAs, the Wnt receptor signal pathway is a key functional pathway, and the prediction of transcription factors indicates that Krüppel-like factor 6 (KLF6, also known as CPBP) may be the key gene regulating these genes and miRNAs network [62].
M1 macrophages also play a crucial role in placental immunity and spiral artery remodeling. MiR-103 can negatively regulate the polarization of M1 macrophages by inhibiting the STAT1/IRF1 signal pathway, resulting in inhibiting the occurrence of SA. Overexpressed miR-103 can effectively reduce the absorption and M1 macrophages polarization of mouse embryos. Therefore, when miR-103 is reduced, it is possible to induce RSA by promoting the STAT1/IRF1 signaling pathway to increase the polarization of M1 macrophages. Moreover, miR-103 can be used to distinguish between RSA patients and normal pregnant subjects because the downregulation of miR-103 in RSA is very sensitive, which makes miR-103 become a promising diagnostic marker and therapeutic target for RSA [64]. In addition, Ding et al. [65] discovered that M1 macrophages could also inhibit the EMT process of trophoblast cells in vitro by secreting extracellular vesicles (EVs). Through miRNA sequencing, miR-146a-5p and miR-146b-5p were identified as the highest expression of miRNAs in trophoblast cells treated with M1-EVs [65]. Further functional experiments suggested that miR-146a-5p and miR-146b-5p in EV directly inhibited TNF receptor-associated factor 6 (TRAF6) expression at the post-transcriptional level, which inhibited the EMT of trophoblast cells in mechanism and played an important role in the communication with trophoblast cells [65].
To summarize, the potential mechanisms of miRNAs regulating SA are varied and may involve different target genes and binding sites (Table 1). In recent years, some studies have found statistical differences in miRNAs in exosomes and plasma between patients with SA and normal pregnant subjects [8,66], which suggests that there are still broad potential pathways in which miRNAs regulate SA that we need to discover. A substantial body of evidence suggests that a miRNA–mRNA regulation network and complicated interactions and crosstalk between miRNAs are involved in the pathogenesis of SA. If this network is thoroughly investigated, it will significantly aid in the diagnosis and treatment of SA.
4. lncRNAs and Spontaneous Abortion
4.1. Overview of lncRNAs
lncRNAs were once considered to be part of transcriptional noise, but now they represent a new “style” of transcriptional and post-transcriptional gene regulation. Structurally, lncRNA is similar to mRNA, with a 5′ cap and 3′ PolyA tail structure, longer than 200 nt and has a complex secondary or tertiary structure without a highly conserved sequence [67,68]. Paradoxically, although the coding regions of lncRNAs are short or non-existent and have a low expression level in cells, they can not only be transcribed from any part of the genome but also play an imperative role in various biological activities such as cell signaling, cell differentiation, transcriptional regulation, epigenetics, and immune response [17,69,70,71,72]. lncRNAs not only regulate the expression of protein-coding genes at the transcriptional and post-transcriptional levels but also are the precursor of smaller regulatory RNAs, such as miRNAs and piwiRNAs [73]. Moreover, as one kind of ceRNA, lncRNAs can also regulate miRNA levels and mRNA stability and translation through homologous base pairing [10].
The research on lncRNAs in SA is increasingly on the rise in recent years. Previously, researchers have identified several lncRNAs subsets that are differentially expressed in the villi of SA patients and normal pregnant women, providing evidence for the involvement of lncRNAs in the physiology and pathogenesis of SA [73,74,75,76]. These studies found that lncRNAs might be mainly involved in the pathogenesis of SA through inflammation, extracellular matrix (ECM)-receptor interaction, and apoptosis pathways, and contributed to the follow-up screening of lncRNA as a marker for the diagnosis of SA. lncRNAs are mainly involved in SA generation and regulation through two ways (Figure 2): (1) regulate the expression of target mRNA directly; (2) regulate the transcription and expression of downstream target mRNA by targeting miRNAs.
4.2. lncRNAs Regulate Target mRNAs Directly
Like miRNAs, lncRNAs can also regulate cellular proliferation, invasion, migration, and apoptosis by modifying target mRNAs. H19 is the first lncRNA discovered by humans, and it represents a milestone in the history of understanding and investigating ncRNA [77]. Several investigations have confirmed that H19 is a significantly expressed gene throughout embryonic development and has a role in tumor invasion and migration [78,79]. Even though the mechanism of embryo implantation in the endometrium is similar to tumor invasion, few studies on H19 in SA have been conducted. Bai et al. [80] recently observed that the expressions of H19 and GPX4 in the SA group were lower than in the standard group. Silencing H19 can downregulate the expression of GPX4 at mRNA and protein levels in HTR8/SVneo cells. A previous study found that GPX4 is a critical regulator in the ferroptosis pathway [80]. Therefore, the correlation between the expression of H19 and GPX4 also implies that ferroptosis might occur in SA, opening up a new research avenue for better comprehension of the pathogenesis of SA.
Some transcription factors and genes directly regulate the expression of lncRNA, affecting its downstream regulatory pathways and controlling the occurrence of SA. Ying Yang 1 (YY1), a transcription factor with the dual role of transcriptional inhibition and activation, is a transcription factor that participates in SA by modulating the expression of cytoskeleton-related proteins to alter the invasion of trophoblast cells [81]. In RSA patients, the YY1mRNA level of primary trophoblast cells was considerably lower than healthy controls [82]. Furthermore, YY1 may directly bind to the PVT1 promoter and subsequently control apoptosis, adhesion, and invasion through the mTOR pathway, thereby affecting the invasion and adhesion of trophoblast cells [83]. Similarly, the HOX transcriptional antisense RNA HOTAIR is a YY1 target lncRNA, and the HOTAIR promoter has two binding sites that can be bound by YY1 [84]. HOTAIR overexpression promotes the production of matrix metalloproteinase 2 and enhances trophoblast cell invasion. Moreover, the tumor suppressor gene, P53, has been elucidated to regulate the expression of numerous lncRNAs [85]. Meanwhile, P53 overexpression would reduce cellular proliferation, migration, and invasion while increasing apoptosis, resulting in SA [86,87]. Wang et al. [88] identified that the expression level of P53 protein was significantly negatively correlated with the expression level of MALAT1 and confirmed that P53 inhibited MALAT1 expression by directly binding to the promoter region. Subsequently, RNA pull-down and co-immunoprecipitation assays demonstrated that MALAT1 causes CRY2 ubiquitin-mediated degradation by engaging E3 ubiquitin ligase FBXW7, resulting in inhibition of SA by participating in the trophoblast cell migration and invasion [89]. Based on the above research, it is assumed that the P53-MALAT1-FBXW7-CRY2 axis plays a dominant role in the pathogenesis of SA.
Immune imbalance is one of the primary causes of SA. In decidual natural killing (DNK) cells, lncRNA has also been demonstrated to have a critical role in the pathogenesis of SA. Human DNK cells constitute the most numerous lymphocyte populations in early pregnancy at the maternal-fetal interface. They act as a regulator of maternal immunological tolerance, facilitating embryo implantation and placenta development [76]. Therefore, it is worthwhile to investigate the differential expression and regulation mechanisms of lncRNAs in the DNK cells of SA patients. Using RNA-seq technology, Li et al. [76] identified that 276 mRNAs and 67 lncRNAs were differentially expressed in the DNK of SA patients versus normal pregnant women. Based on these differentially expressed RNAs, a lncRNA-mRNA regulatory network with small-world characteristics was built, which is involved in a variety of biological processes (immune response, inflammatory response, cell adhesion, and ECM organization) and signal pathways (cytokine-cytokine receptor interaction, ECM-receptor interaction, Toll-like receptor signaling pathway, and phosphatidylinositol signaling system).
Furthermore, lnc-49a, a novel lncRNA identified recently, is a positive regulator of CD49a in DNK cells and is involved in regulating DNK cell functions, such as cellular activity, migration, and adhesion. Li et al. [90] discovered that the expression of CD49a was downregulated in RSA patients. In contrast, the expressions of perforin, granzyme B, and IFNG were upregulated, implying that lnc-49a can impede SA through an unknown mechanism by regulating the expression of the targeted gene CD49a [90]. In peripheral blood, some lncRNAs are related to the occurrence of SA. For example, Liu et al. [74] found that RP11-115N4.1 was the most abundant lncRNA in the peripheral blood of URSA patients. The activation of RP11-115N4.1 can significantly increase the binding protein of HSP70 and HNRNPH3, which may be one of its mechanisms for regulating immune response, and it can trigger the occurrence of SA via inhibiting the migration of trophoblast cells.
Moreover, lncRNA can influence SA through imprinting. During placental development, imprinted lncRNA has a dynamic temporal and spatial expression. Sheng et al. [91] found that the imprinted lncRNA Rian might play a crucial role in placenta development because its homologous sequence lncRNA MEG8 (RIAN) was unusually highly expressed in human SA villi. Upregulation of MEG8 expression inhibited cell proliferation and invasion in trophoblast cell lines, whereas downregulation of MEG8 expression had the opposite effect. These findings suggest that MEG8 plays an important regulatory role in trophoblast cell growth and function and that incorrect MEG8 expression may result in dysfunctional trophoblast cells, which may play a role in the formation of URSA.
Furthermore, enhancer RNAs, a group of lncRNAs transcribed from enhancers, have been linked to URSA. The regulatory effect of enhancers on gene expression is a new area of study, and LNC-SLC4A1-1, transcribed from active enhancers labeled with H3K27ac and H3K4me1, has been found to bind to CXCL8 by recruiting NF-κB, resulting in CXCL8 activation. The activated CXCL8 induces an increase in TNF-α and IL-1β, intensifying trophoblast cell inflammation [92]. These studies provide new insights into understanding the role of lncRNA in pathological pregnancy and URSA research.
4.3. lncRNAs Regulate Target miRNAs to Regulate Their Downstream Target mRNAs
A considerable number of lncRNAs, such as ceRNA, regulate gene expression through sponging miRNA [93]. For example, H19 has been identified to compete with miRNA let-7 to control the target gene Integrinβ3 (ITGB3), critical in endometrial receptivity [94]. When H19 was knocked out, the expression of ITGB3 was reduced, resulting in a downregulation of the adhesion and invasion. Further experiments have demonstrated that H19 suppresses the impact of let-7 by acting as a molecular sponge to avoid the degradation of ITGB3mRNA and improves the EVT adherence to the endometrium, thereby promoting EVT invasion during early pregnancy [94]. Moreover, PVT1 can also regulate the function of trophoblast cells through the PVT1/miR-424/eIF5A pathway [83]. When PVT1 is overexpressed, it suppresses miR-424, which in turn decreases the production of miR-424’s target gene, eIF5A, favoring cell survival, proliferation, and migration [83]. Xiang et al. [95] also noticed SNHG7-1, a specific lncRNA, regulated the proliferation and invasion of trophoblast cells and promoted the occurrence of RSA via the Wnt/β-catenin signaling pathway, which was targeted by binding to miR-34a, hence encouraging the onset of RSA. Huang et al. [28] recently discovered an innovative lncRNA called lnc-HZ04, which can directly and specifically bind to miR-hz04, weakening the decreasing the effect of miR-hz04 on inositol 1,4,5-trisphosphate receptor type 1 (IP3R1) mRNA expression and stability, thus activating the IP3R1/p-CaMKII/SGCB pathway mediated by Ca2+ and further promoting trophoblast cell apoptosis.
It is still rare to find research on the regulation of target genes and pathways by lncRNA through sponging miRNA. Previous studies have indicated that this regulatory pathway appears to play an indispensable role in the occurrence and progression of SA (Table 2), implying a better understanding of the mechanism of lncRNA involved in SA.
5. circRNAs and Spontaneous Abortion
5.1. Overview of circRNAs
circRNAs are a new class of endogenous ncRNA, which were first discovered in 1979 [97]. However, they were subsequently regarded as “junk products” of gene expression and did not get people’s attention. Until Memczak et al. [98] and Hansen et al. [99] discovered the unique functions of circRNAs, circRNAs quickly became a new generation of star molecules in the following years. This type of RNA has a stable structure and is known for its unique closed-loop and single-stranded structure. Since there is no free end as the binding point of exonuclease, circRNA has strong resistance to exonuclease hydrolysis, which results in some unique characteristics of circRNA that other ncRNAs do not have [100,101], such as longer half-life [102]. Currently, the biological functions of circRNAs are being studied extensively. For instance, they act as miRNA sponges to regulate the function of miRNAs [103], as transcription or translation regulators to affect protein expression [98,104], and interact with proteins to regulate gene expression [105,106]. Unexpectedly, some circRNAs even have the potential to encode proteins [107]. Moreover, circRNA has been determined to be involved in the pathogenesis of many diseases, such as atherosclerosis, neurological disorders, diabetes, and cancer [101,108].
5.2. circRNAs and SA
circRNAs have been shown in recent studies to play a critical role in tumor regulation [109,110,111]. Whereas there are multiple similarities between tumorigenesis and embryo implantation in humans, circRNA may also play an indispensable role in cellular invasion, EMT, migration, and apoptosis. Indeed, several researchers have begun to pay attention to the correlation between circRNAs and SA (Table 2). Li et al. [112] discovered 123 significantly differentially expressed circRNAs in early RSA patients’ decidual tissues compared to normal pregnant women. Based on these significantly differentially expressed circRNAs, a miRNA-mRNA network targeted by circRNA was constructed, including three circRNAs (has_circRNA_104179, has_circRNA_103093, and has_circRNA_103092), 27 miRNAs, and 82 mRNAs. Another study analyzed the expression of circRNAs in chorionic villi of RSA patients and healthy controls by using circRNA chips and found 594 differently expressed circRNAs [113]. Further analysis revealed that the aberrant production of circRNAs in RSA villi contributes to RSA pathogenesis via sponging miRNA.
Some experiments specifically focus on the circRNA regulatory pathways in SA. Li et al. [96] investigated the effect of CIRC-ZUFSP overexpression and downregulation on trophoblast cell activity in vitro. They discovered that CIRC-ZUFSP influences the molecular mechanism of trophoblast cell migration and invasion through modulating miR-203. STOX1 is the target gene of miR-203. Thus CIRC-ZUFSP may induce SA through the CIRC-ZUFSP/miR-203/STOX1 pathway, which inhibits the migration and invasion of trophoblast cells [96]. In addition, Zhu et al. [31] discovered that circPUM1 could promote trophoblast cell processes and anti-inflammatory effects through the miR-30a-5p/JunB axis, hence preventing the formation and progression of SA. CircPUM1 gene knockout reduces trophoblast cell proliferation, migration, and invasion, increases trophoblast cell apoptosis, and promotes pro-apoptotic protein (cleaved caspases-3) levels and pro-inflammatory factors (TNF-α, IL-6, and IL-8) secretion, similar to the role of lncRNA CXCL8 discussed above.
Furthermore, circFOXP1 (hsa_circ_0008234), which is generated from the exon region of the FOXP1 gene, may regulate the cell activity of many disorders, and Gao et al. [30] demonstrated for the first time that it could regulate the function of trophoblast cells via the miR-143-3p/S100A11 cascade pathway. Mechanically, circFOXP1 regulates S100 calcium binding protein A11 (S100A11) expression by binding to miR-143- 3p competitively. These studies have shown that circRNA has similarities to lncRNA, which can be combined with the corresponding miRNAs to regulate the expression of their downstream target genes, thereby controlling the occurrence and progression of SA. Concurrently, RNA studies have provided new ideas and research directions for the diagnosis and treatment of SA.
6. ceRNA Network in SA: A Systematic Perspective to Elucidate the Regulation of SA by ncRNAs
Based on the preceding explanation, it can be concluded that there is an intricate RNA regulatory network connected through a complex network of feedback mechanisms in the case of SA. In 2011, Salmena et al. [10] proposed the ceRNA regulatory network concept, claiming that mRNAs, transcribed pseudogenes, and lncRNA could interact via a unique “language” established by miRNA binding sites (“miRNA response elements” or “MRE”). This concept is currently being refined and is widely accepted. Non-coding RNAs, such as circRNA, were later discovered and added to this network [93]. In recent years, some studies have clarified the role of ceRNA networks in cancer development [114,115,116] and pregnancy diseases such as recurring implantation failure (RIF) [117]. More directly, Huang et al. [118] and Zang et al. [119] validated the presence of this network in SA patients in humans and pigs, respectively. However, there is a paucity of in-depth research on the role of the ceRNA network in the pathogenesis of SA. Based on the findings of previous studies of miRNA, lncRNA, and circRNA in SA, we anticipate that several ceRNA networks have the potential to serve as a guide and reference for more in-depth research of the mechanism and role of ceRNA in SA (Figure 3).
6.1. LncRNAs-miRNAs-mRNAs Network
According to the ceRNA hypothesis, lncRNAs operate as molecular sponges to absorb miRNAs, resulting in decreased binding to their downstream target mRNAs and restoration of mRNA activity or expression [10,93]. Huang et al. [118] and Zang et al. [119] created ceRNA networks in SA patients from humans and pigs. This network in human SA comprises 31 lncRNAs, 1 miRNA (hsa-miR-210-5p), and 3 genes (NTNG2, GRIA1, and AQP1) [118]. While in pigs, it consists of 4 LncRNAs (TCONS_00051274, TCONS_00161675, TCONS_00108310, TCONS_00177102), 11 miRNAs and 13 genes [119]. Some sporadic lncRNAs, such as GAS5, have also been confirmed to be correlated with miR140-5p as a ceRNA [120]. In addition, UCA has been found to operate as a ceRNA through the uptake of miR-455, which leads to the upregulation of RUNX family transcription factor 2 (RUNX2) expression in HTR-8/SVneo cells [121]. The H19, PVT1, SNHG7, and lnc-HZ04 described above can hypothetically be utilized as ceRNAs to absorb miRNAs and contribute to the occurrence and development of SA. However, the ceRNAs that we have identified are only the tip of the iceberg regarding the entire ceRNA regulatory network, and the comprehensive network control mechanism that underpins it is still mysterious.
6.2. CircRNAs-miRNAs-mRNAs Network
CircRNA contains multiple miRNA binding sites acting on miRNA sponges. These sponges work as CeRNAs to improve miRNA-induced target gene suppression by boosting their expression levels and through disease-related miRNAs interaction [122]. There are currently no relevant data to elucidate the role of the circRNA-mediated ceRNA regulatory network in the occurrence and progression of SA. Other studies, however, have shown that the circRNA-mediated ceRNA regulatory network plays a critical role in the average growth and development of embryonic organs and tissues in pig and mouse embryos [123,124]. These studies have possible implications for the universality of circRNA-mediated ceRNA networks in various organisms.
Similarly, some researchers have already discovered circRNA-mediated ceRNA networks in pregnancy diseases, such as preeclampsia (PE) and RIF [125,126]. Contemporary research on the role of circRNA in SA has recently shown that circRNAs regulate SA through the circRNA-miRNA-mRNA axis. Based on a solid body of evidence, we have reason to believe that the circRNA-miRNA-mRNA network is essential in the occurrence and regulation of SA, which suggests that we should focus our efforts in the future on ceRNA to reveal the mechanism of circRNA in SA.
6.3. lncRNAs/circRNAs-miRNAs-mRNAs Network
Both lncRNA and circRNA contain miRNA binding sites and bind the same miRNA. These miRNA binding sites exist in a competitive interaction, complicating the connection and regulation mechanism of this ceRNA network. As a result, searching for such a ceRNA network necessitates the use of more advanced and mature RNA-seq technology and bioinformatics analysis methods. Several researchers have discovered that ceRNA networks mediated by lncRNA and circRNA exist in post-menopausal complications and PE [127,128]. These networks are usually more extensive and complex than the single lncRNA-mediated or circRNA-mediated ceRNA networks.
Based on the above research results, we believe that there are at least one or more ceRNA regulatory networks in SA, and these networks have critical regulatory effects on the onset and progression of SA. Identifying these networks and RNA markers based on them is crucial in understanding the specific etiology diagnosis and treatment of SA.
7. Conclusions and Perspective
This study discusses the most recent advances in ncRNAs research, the ceRNA regulatory network, and potential molecular pathways relevant to the incidence and progression of SA. Through these studies, we have established that a complex ceRNA regulatory mechanism exists in the organism that regulates cell proliferation, invasion, migration, EMT, apoptosis, and angiogenesis to regulate the occurrence and development of SA. With advancements in technology such as next-generation and the maturation of RNA chips, ncRNAs with differential expression will be identified in SA. These ncRNAs will provide reference and insights into the construction of ceRNA networks and the in-depth study of the regulation of differentially expressed ncRNAs in SA.
SA in pigs is a major problem that impairs the economic efficiency of commercial pig production. Given the highly conservative nature of some ncRNAs, such as some miRNAs and circRNAs, studies of ncRNAs in humans and mice may provide valuable references for researching the regulatory mechanisms of ncRNAs in porcine SA occurrence. Likewise, owing to the similarity with humans in size, physiology, and genomic characteristics, pigs are generally considered to be the best animal model for researching human diseases [129], which addresses the part of the research on SA that is difficult to conduct in humans due to the ethical problem. This can provide a deeper understanding for researchers of the occurrence mechanism of SA and provide a theoretical basis for treating and preventing SA.
Moreover, ncRNAs are becoming increasingly important in biomedical research as new biomarkers in the diagnosis, prediction, prognosis, and therapeutic response to diseases [130]. ncRNA expression profiles are more effective than mRNA expression profiles in distinguishing normal and pathological tissues [131]. Therefore, searching for the differentially expressed ncRNAs in SA will not only provide us with a more comprehensive understanding of the pathogenesis of URSA but also enable the selection of certain ncRNAs as early predictors of SA, which will improve the prevention and treatment of the disease.
Although recent studies have partially explained the role and mechanism of ncRNAs in SA, we still lack a comprehensive understanding of this process, and there are still some challenges and mechanisms of ncRNAs in SA that need to be resolved and explored. Undoubtfully, the pathogenesis and regulatory network of ncRNA on SA will be elucidated with an in-depth study of ncRNA, which will also accelerate the process of the quest for potential RNA markers for SA diagnosis and treatment. These regulatory networks have potential applications in obstetric practice and pork production, improving women’s health and commercial pig production efficiency.
Author Contributions
Conceptualization, J.G.; investigation, J.G. and T.G; writing—original draft preparation, J.G. and T.G.; writing—review and editing, J.G. and H.Y.; funding acquisition, Z.A. and G.C.; project administration, L.H. and Z.W. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Department of Science and Technology of Guangdong Province (grant number 2018B030313011) and the Foundation of Key Laboratory of Animal Genetics, Breeding and Reproduction in The Plateau Mountainous Region, Ministry of Education, Guizhou University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
Some figures in this review were created with the help of BioRender software. The authors declare no conflict of interest.
References
- Regan, L.; Rai, R. Epidemiology and the medical causes of miscarriage. Baillieres Best Pract. Res. Clin. Obstet. Gynaecol. 2000, 14, 839–854. [Google Scholar] [CrossRef] [PubMed]
- Tur-Torres, M.H.; Garrido-Gimenez, C.; Alijotas-Reig, J. Genetics of recurrent miscarriage and fetal loss. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 42, 11–25. [Google Scholar] [CrossRef]
- Practice Committee of the American Society for Reproductive Medicine. Electronic address, a.a.o. Definitions of infertility and recurrent pregnancy loss: A committee opinion. Fertil. Steril. 2020, 113, 533–535. [Google Scholar] [CrossRef]
- Ross, J.W.; Ashworth, M.D.; Stein, D.R.; Couture, O.P.; Tuggle, C.K.; Geisert, R.D. Identification of differential gene expression during porcine conceptus rapid trophoblastic elongation and attachment to uterine luminal epithelium. Physiol. Genom. 2009, 36, 140–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido-Gimenez, C.; Alijotas-Reig, J. Recurrent miscarriage: Causes, evaluation and management. Postgrad. Med. J. 2015, 91, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, O.B.; Steffensen, R.; Nielsen, H.S.; Varming, K. Multifactorial etiology of recurrent miscarriage and its scientific and clinical implications. Gynecol. Obstet. Investig. 2008, 66, 257–267. [Google Scholar] [CrossRef]
- Warren, J.E.; Silver, R.M. Genetics of pregnancy loss. Clin. Obstet. Gynecol. 2008, 51, 84–95. [Google Scholar] [CrossRef]
- Qin, W.; Tang, Y.; Yang, N.; Wei, X.; Wu, J. Potential role of circulating microRNAs as a biomarker for unexplained recurrent spontaneous abortion. Fertil. Steril. 2016, 105, 1247–1254.e3. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Li, N.; Sun, L.; Zheng, D.; Shao, G. Non-coding RNAs: The key detectors and regulators in cardiovascular disease. Genomics 2021, 113, 1233–1246. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.; Gupta, P.; Sahoo, S.; Mallick, B. Non-coding RNAs and their cross-talks impacting reproductive health of women. Wiley Interdiscip. Rev. RNA 2021, e1695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, P.; Zhang, L.; Huang, C.; Gao, J.; Li, Y.; Yang, B. Identification of Key Genes and Long Noncoding RNA-Associated Competing Endogenous RNA (ceRNA) Networks in Early-Onset Preeclampsia. Biomed Res. Int. 2020, 2020, 1673486. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Wang, L.; Li, S.; Wang, H.; Huang, S.; Zhang, Z.; Wang, R.; Guan, H.; Huang, Y. Identification of Key Molecules and lncRNA-miRNA-mRNA ceRNA Network in Preeclampsia. Int. J. Gen. Med. 2021, 14, 7579–7590. [Google Scholar] [CrossRef]
- Fu, X.; Cong, H.; Zhao, S.; Li, Y.; Liu, T.; Sun, Y.; Lv, N. Construction of Glycometabolism- and Hormone-Related lncRNA-Mediated Feedforward Loop Networks Reveals Global Patterns of lncRNAs and Drug Repurposing in Gestational Diabetes. Front. Endocrinol. 2020, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Leng, L.; Zhang, C.; Ren, L.; Li, Q. Construction of a long noncoding RNA-mediated competitive endogenous RNA network reveals global patterns and regulatory markers in gestational diabetes. Int. J. Mol. Med. 2019, 43, 927–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogasawara, M.; Aoki, K.; Okada, S.; Suzumori, K. Embryonic karyotype of abortuses in relation to the number of previous miscarriages. Fertil. Steril. 2000, 73, 300–304. [Google Scholar] [CrossRef]
- Chen, X.; Guo, D.Y.; Yin, T.L.; Yang, J. Non-Coding RNAs Regulate Placental Trophoblast Function and Participate in Recurrent Abortion. Front. Pharmacol. 2021, 12, 646521. [Google Scholar] [CrossRef]
- McNamee, K.; Dawood, F.; Farquharson, R. Recurrent miscarriage and thrombophilia: An update. Curr. Opin. Obstet. Gynecol. 2012, 24, 229–234. [Google Scholar] [CrossRef]
- Kumar, S. Occupational, environmental and lifestyle factors associated with spontaneous abortion. Reprod. Sci. 2011, 18, 915–930. [Google Scholar] [CrossRef]
- Vasconcelos, S.; Ramalho, C.; Marques, C.J.; Doria, S. Altered expression of epigenetic regulators and imprinted genes in human placenta and fetal tissues from second trimester spontaneous pregnancy losses. Epigenetics 2019, 14, 1234–1244. [Google Scholar] [CrossRef]
- Nikitina, T.V.; Sazhenova, E.A.; Tolmacheva, E.N.; Sukhanova, N.N.; Kashevarova, A.A.; Skryabin, N.A.; Vasilyev, S.A.; Nemtseva, T.N.; Yuriev, S.Y.; Lebedev, I.N. Comparative Cytogenetic Analysis of Spontaneous Abortions in Recurrent and Sporadic Pregnancy Losses. Biomed. Hub 2016, 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sugiura-Ogasawara, M.; Ozaki, Y.; Suzumori, N. Management of recurrent miscarriage. J. Obstet. Gynaecol. Res. 2014, 40, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Pollheimer, J.; Vondra, S.; Baltayeva, J.; Beristain, A.G.; Knofler, M. Regulation of Placental Extravillous Trophoblasts by the Maternal Uterine Environment. Front. Immunol. 2018, 9, 2597. [Google Scholar] [CrossRef]
- Wu, L.; Cheng, B.; Liu, Q.; Jiang, P.; Yang, J. CRY2 suppresses trophoblast migration and invasion in recurrent spontaneous abortion. J. Biochem. 2020, 167, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Tian, P.; Xu, H. MicroRNA-155-5p regulates survival of human decidua stromal cells through NF-kappaB in recurrent miscarriage. Reprod. Biol. 2021, 21, 100510. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Li, Z.; Hu, P.; Wang, J.; Fu, J.; Wei, B.; Zhang, Y. miR-219a suppresses human trophoblast cell invasion and proliferation by targeting vascular endothelial growth factor receptor 2 (VEGFR2). J. Assist. Reprod. Genet. 2021, 38, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.X.; Tang, Z.Y. Long non-coding RNA H19 regulates Bcl-2, Bax and phospholipid hydroperoxide glutathione peroxidase expression in spontaneous abortion. Exp. Ther. Med. 2021, 21, 41. [Google Scholar] [CrossRef]
- Huang, W.; Dai, M.; Qiu, T.; Liang, T.; Xie, J.; Mi, C.; Zhao, J.; Chen, W.; Tian, P.; Zhang, S.; et al. Novel lncRNA-HZ04 promotes BPDE-induced human trophoblast cell apoptosis and miscarriage by upregulating IP(3) R(1) /CaMKII/SGCB pathway by competitively binding with miR-hz04. FASEB J. 2021, 35, e21789. [Google Scholar] [CrossRef]
- Xu, Z.; Tian, P.; Guo, J.; Mi, C.; Liang, T.; Xie, J.; Huang, W.; Dai, M.; Chen, W.; Zhang, H. Lnc-HZ01 with m6A RNA methylation inhibits human trophoblast cell proliferation and induces miscarriage by up-regulating BPDE-activated lnc-HZ01/MXD1 positive feedback loop. Sci. Total Environ. 2021, 776. [Google Scholar] [CrossRef]
- Gao, Y.; Tang, Y.; Sun, Q.; Guan, G.; Wu, X.; Shi, F.; Zhou, Z.; Yang, W. Circular RNA FOXP1 relieves trophoblastic cell dysfunction in recurrent pregnancy loss via the miR-143-3p/S100A11 cascade. Bioengineered 2021, 12, 9081–9093. [Google Scholar] [CrossRef]
- Zhu, L.; Shi, L.; Ye, W.; Li, S.; Liu, X.; Zhu, Z. Circular RNA PUM1 (CircPUM1) attenuates trophoblast cell dysfunction and inflammation in recurrent spontaneous abortion via the MicroRNA-30a-5p (miR-30a-5p)/JUNB axis. Bioengineered 2021, 12, 6878–6890. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, D.; Villén, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The impact of microRNAs on protein output. Nature 2008, 455, 64–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Sayed, D.; Abdellatif, M. MicroRNAs in development and disease. Physiol. Rev. 2011, 91, 827–887. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Elovitz, M.A.; Anton, L.; Bastek, J.; Brown, A.G. Can microRNA profiling in maternal blood identify women at risk for preterm birth? Am. J. Obstet. Gynecol. 2015, 212, 782.e1–782.e5. [Google Scholar] [CrossRef]
- Fu, G.; Brkić, J.; Hayder, H.; Peng, C. MicroRNAs in Human Placental Development and Pregnancy Complications. Int. J. Mol. Sci. 2013, 14, 5519–5544. [Google Scholar] [CrossRef] [Green Version]
- Kokawa, K.; Shikone, T.; Nakano, R. Apoptosis in human chorionic villi and decidua during normal embryonic development and spontaneous abortion in the first trimester. Placenta 1998, 19, 21–26. [Google Scholar] [CrossRef]
- Liu, H.N.; Tang, X.M.; Wang, X.Q.; Gao, J.; Li, N.; Wang, Y.Y.; Xia, H.F. MiR-93 Inhibits Trophoblast Cell Proliferation and Promotes Cell Apoptosis by Targeting BCL2L2 in Recurrent Spontaneous Abortion. Reprod. Sci. 2020, 27, 152–162. [Google Scholar] [CrossRef]
- Tian, S.; Yu, J.; Zhang, Y.; Bian, Y.; Ma, J.; Yan, J. Overexpression of PTEN regulated by miR-19b and miR-494 in the villous of recurrent spontaneous abortion patients. J. Reprod. Immunol. 2020, 140, 103133. [Google Scholar] [CrossRef] [PubMed]
- Gimm, O.; Perren, A.; Weng, L.-P.; Marsh, D.J.; Yeh, J.J.; Ziebold, U.; Gil, E.; Hinze, R.; Delbridge, L.; Lees, J.A.; et al. Differential Nuclear and Cytoplasmic Expression of PTEN in Normal Thyroid Tissue, and Benign and Malignant Epithelial Thyroid Tumors. Am. J. Pathol. 2000, 156, 1693–1700. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Shen, W.W.; Cao, X.M.; Ding, W.Y.; Yan, L.P.; Gao, L.J.; Li, X.L.; Zhong, T.Y. Novel mechanism of miRNA-365-regulated trophoblast apoptosis in recurrent miscarriage. J. Cell. Mol. Med. 2017, 21, 2412–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lv, Y.; Wang, L.; Gong, C.; Sun, J.; Chen, X.; Chen, Y.; Yang, L.; Zhang, Y.; Yang, X.; et al. MicroRNAome in decidua: A new approach to assess the maintenance of pregnancy. Fertil. Steril. 2015, 103, 980–989.e6. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Yang, L.; Wang, H. miR-520 promotes DNA-damage-induced trophoblast cell apoptosis by targeting PARP1 in recurrent spontaneous abortion (RSA). Gynecol. Endocrinol. 2017, 33, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Powe, C.E.; Levine, R.J.; Karumanchi, S.A. Preeclampsia, a disease of the maternal endothelium: The role of antiangiogenic factors and implications for later cardiovascular disease. Circulation 2011, 123, 2856–2869. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Liu, J.; Fu, T.; Shan, B.; Qian, L.; Pan, L.; Yuan, J. USP25 regulates Wnt signaling by controlling the stability of tankyrases. Genes Dev. 2017, 31, 1024–1035. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Cheng, Y.; Zhang, Y.; Liao, S.; Yin, T.; Yang, J. The miR-27a-3p/USP25 axis participates in the pathogenesis of recurrent miscarriage by inhibiting trophoblast migration and invasion. J. Cell. Physiol. 2019, 234, 19951–19963. [Google Scholar] [CrossRef]
- Zhu, Y.; Lu, H.; Huo, Z.; Ma, Z.; Dang, J.; Dang, W.; Pan, L.; Chen, J.; Zhong, H. MicroRNA-16 inhibits feto-maternal angiogenesis and causes recurrent spontaneous abortion by targeting vascular endothelial growth factor. Sci. Rep. 2016, 6, 35536. [Google Scholar] [CrossRef] [Green Version]
- Chai, L.; Ling, K.; He, X.; Yang, R. Expression of ATF4 and VEGF in chorionic villus tissue in early spontaneous abortion. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 434–438. [Google Scholar] [CrossRef]
- Lala, P.K. The effects of angiogenic growth factors on extravillous trophoblast invasion and motility. Placenta 2000, 21, 593–595. [Google Scholar] [CrossRef] [PubMed]
- Khan, G.A.; Girish, G.V.; Lala, N.; Di Guglielmo, G.M.; Lala, P.K. Decorin is a novel VEGFR-2-binding antagonist for the human extravillous trophoblast. Mol. Endocrinol. 2011, 25, 1431–1443. [Google Scholar] [CrossRef] [PubMed]
- Chatuphonprasert, W.; Jarukamjorn, K.; Ellinger, I. Physiology and Pathophysiology of Steroid Biosynthesis, Transport and Metabolism in the Human Placenta. Front. Pharmacol. 2018, 9, 1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyall, F.; Bulmer, J.N.; Duffie, E.; Cousins, F.; Theriault, A.; Robson, S.C. Human Trophoblast Invasion and Spiral Artery Transformation. Am. J. Pathol. 2001, 158, 1713–1721. [Google Scholar] [CrossRef]
- Smith, S.D.; Dunk, C.E.; Aplin, J.D.; Harris, L.K.; Jones, R.L. Evidence for immune cell involvement in decidual spiral arteriole remodeling in early human pregnancy. Am. J. Pathol. 2009, 174, 1959–1971. [Google Scholar] [CrossRef] [Green Version]
- Wallace, A.E.; Fraser, R.; Cartwright, J.E. Extravillous trophoblast and decidual natural killer cells: A remodelling partnership. Hum. Reprod. Update 2012, 18, 458–471. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Li, J. Association of miR-34a-3p/5p, miR-141-3p/5p, and miR-24 in Decidual Natural Killer Cells with Unexplained Recurrent Spontaneous Abortion. Med. Sci. Monit. 2016, 22, 922–929. [Google Scholar] [CrossRef] [Green Version]
- He, X.; He, L.; Hannon, G.J. The guardian’s little helper: microRNAs in the p53 tumor suppressor network. Cancer Res. 2007, 67, 11099–11101. [Google Scholar] [CrossRef] [Green Version]
- Fayyad-Kazan, H.; Hamade, E.; Rouas, R.; Najar, M.; Fayyad-Kazan, M.; El Zein, N.; ElDirani, R.; Hussein, N.; Fakhry, M.; Al-Akoum, C.; et al. Downregulation of microRNA-24 and -181 parallels the upregulation of IFN-γ secreted by activated human CD4 lymphocytes. Hum. Immunol. 2014, 75, 677–685. [Google Scholar] [CrossRef]
- Fiedler, J.; Jazbutyte, V.; Kirchmaier, B.C.; Gupta, S.K.; Lorenzen, J.; Hartmann, D.; Galuppo, P.; Kneitz, S.; Pena, J.T.; Sohn-Lee, C.; et al. MicroRNA-24 regulates vascularity after myocardial infarction. Circulation 2011, 124, 720–730. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Gao, R.; Chen, X.; Zhang, H.; Zheng, A.; Yang, D.; Ding, Y.; Wang, Y.; He, J. Possible roles of mmu-miR-141 in the endometrium of mice in early pregnancy following embryo implantation. PLoS ONE 2013, 8, e67382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Li, J.; Jia, B.; Wang, Y.; Zhang, J.; Liu, G. Genome-wide identification of microRNAs in decidual natural killer cells from patients with unexplained recurrent spontaneous abortion. Am. J. Reprod. Immunol. 2018, 80, e13052. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.; Yoo, J.K.; Lee, J.; Lee, D.R.; Ko, J.J.; Oh, S.H.; Choo, Y.K.; Kim, J.K. The hsa-miR-5787 represses cellular growth by targeting eukaryotic translation initiation factor 5 (eIF5) in fibroblasts. Biochem. Biophys. Res. Commun. 2011, 415, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liu, H.; Zhang, Z.; Wei, R.; Zhou, X.; Wang, Z.; Zhao, L.; Guo, Q.; Zhang, Y.; Chu, C.; et al. MiR-103 protects from recurrent spontaneous abortion via inhibiting STAT1 mediated M1 macrophage polarization. Int. J. Biol. Sci. 2020, 16, 2248–2264. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhang, Y.; Cai, X.; Zhang, Y.; Yan, S.; Wang, J.; Zhang, S.; Yin, T.; Yang, C.; Yang, J. Extracellular vesicles derived from M1 macrophages deliver miR-146a-5p and miR-146b-5p to suppress trophoblast migration and invasion by targeting TRAF6 in recurrent spontaneous abortion. Theranostics 2021, 11, 5813–5830. [Google Scholar] [CrossRef]
- Cui, S.; Zhang, J.; Li, J.; Wu, H.; Zhang, H.; Yu, Q.; Zhou, Y.; Lv, X.; Zhong, Y.; Luo, S.; et al. Circulating microRNAs from serum exosomes as potential biomarkers in patients with spontaneous abortion. Am. J. Transl. Res. 2021, 13, 4197–4210. [Google Scholar]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Xiao, X.; Zhao, Y.; Yin, L.; Fu, M.; Zhang, X.; Jiang, P. The functional roles of long noncoding RNA DANCR in Human Cancers. J. Cancer 2020, 11, 6970–6981. [Google Scholar] [CrossRef]
- Cao, J. The functional role of long non-coding RNAs and epigenetics. Biol. Proced. Online 2014, 16, 11. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, E.L.; Belhocine, M.; Dao, L.T.; Puthier, D.; Spicuglia, S. Functions of lncRNA in development and diseases. Med. Sci. 2014, 30, 790–796. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, S.; Aiello, D.; Atianand, M.K.; Ricci, E.P.; Gandhi, P.; Hall, L.L.; Byron, M.; Monks, B.; Henry-Bezy, M.; Lawrence, J.B.; et al. A long noncoding RNA mediates both activation and repression of immune response genes. Science 2013, 341, 789–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S. Expression of Concern: The functional role of long non-coding RNAs and epigenetics. Biol. Proced. Online 2016, 18, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Cao, Q.; Ge, J.; Liu, C.; Ma, Y.; Meng, Y.; Wang, Y.; Zhao, X.; Liu, R.; Li, C.; et al. LncRNA-regulated infection and inflammation pathways associated with pregnancy loss: Genome wide differential expression of lncRNAs in early spontaneous abortion. Am. J. Reprod. Immunol. 2014, 72, 359–375. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, X.; Zhu, L.; Zhu, M.; Deng, K.; Nie, X.; Mo, H.; Du, T.; Huang, B.; Hu, L.; et al. Long Noncoding RNA RP11-115N4.1 Promotes Inflammatory Responses by Interacting With HNRNPH3 and Enhancing the Transcription of HSP70 in Unexplained Recurrent Spontaneous Abortion. Front. Immunol. 2021, 12, 717785. [Google Scholar] [CrossRef]
- Wang, L.; Tang, H.; Xiong, Y.; Tang, L. Differential expression profile of long noncoding RNAs in human chorionic villi of early recurrent miscarriage. Clin. Chim. Acta 2017, 464, 17–23. [Google Scholar] [CrossRef]
- Li, T.; Li, X.; Guo, Y.; Zheng, G.; Yu, T.; Zeng, W.; Qiu, L.; He, X.; Yang, Y.; Zheng, X.; et al. Distinct mRNA and long non-coding RNA expression profiles of decidual natural killer cells in patients with early missed abortion. FASEB J. 2020, 34, 14264–14286. [Google Scholar] [CrossRef]
- Brannan, C.I.; Dees, E.C.; Ingram, R.S.; Tilghman, S.M. The product of the H19 gene may function as an RNA. Mol. Cell. Biol. 1990, 10, 28–36. [Google Scholar] [CrossRef]
- Kallen, A.N.; Zhou, X.B.; Xu, J.; Qiao, C.; Ma, J.; Yan, L.; Lu, L.; Liu, C.; Yi, J.S.; Zhang, H.; et al. The imprinted H19 lncRNA antagonizes let-7 microRNAs. Mol. Cell 2013, 52, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Zhou, J.; Gao, Y.; Ghazal, S.; Lu, L.; Bellone, S.; Yang, Y.; Liu, N.; Zhao, X.; Santin, A.D.; et al. Regulation of tumor cell migration and invasion by the H19/let-7 axis is antagonized by metformin-induced DNA methylation. Oncogene 2015, 34, 3076–3084. [Google Scholar] [CrossRef]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of Ferroptotic Cancer Cell Death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, L.; Munir, S.; Fu, G.; Dunk, C.; Baczyk, D.; Caniggia, I.; Lye, S.; Peng, C. Nodal signals through activin receptor-like kinase 7 to inhibit trophoblast migration and invasion: Implication in the pathogenesis of preeclampsia. Am. J. Pathol. 2011, 178, 1177–1189. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.J.; Cheng, Y.X.; Li, X.C.; Wang, F.; Qin, C.M.; Ma, X.L.; Yang, J.; Lin, Y. The YY1/MMP2 axis promotes trophoblast invasion at the maternal-fetal interface. J. Pathol. 2016, 239, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Ding, J.; Wang, Y.; Yuan, M.; Xian, S.; Zhang, L.; Liu, S.; Dai, F.; Wang, F.; Zheng, Y.; et al. YY1-PVT1 affects trophoblast invasion and adhesion by regulating mTOR pathway-mediated autophagy. J. Cell. Physiol. 2020, 235, 6637–6646. [Google Scholar] [CrossRef]
- Zhang, Y.; Jin, F.; Li, X.C.; Shen, F.J.; Ma, X.L.; Wu, F.; Zhang, S.M.; Zeng, W.H.; Liu, X.R.; Fan, J.X.; et al. The YY1-HOTAIR-MMP2 Signaling Axis Controls Trophoblast Invasion at the Maternal-Fetal Interface. Mol. Ther. 2017, 25, 2394–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldassarre, A.; Masotti, A. Long non-coding RNAs and p53 regulation. Int. J. Mol. Sci. 2012, 13, 16708–16717. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.H.; Helfand, S.L. New tricks of an old molecule: Lifespan regulation by p53. Aging Cell 2006, 5, 437–440. [Google Scholar] [CrossRef] [Green Version]
- Kaare, M.; Bützow, R.; Ulander, V.M.; Kaaja, R.; Aittomäki, K.; Painter, J.N. Study of p53 gene mutations and placental expression in recurrent miscarriage cases. Reprod. Biomed. Online 2009, 18, 430–435. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.Z.; Liu, Y.; Wang, H.J.; Pang, W.W.; Zhang, J.J. Disordered p53-MALAT1 pathway is associated with recurrent miscarriage. Kaohsiung J. Med. Sci. 2019, 35, 87–94. [Google Scholar] [CrossRef]
- Wu, L.; Liu, Q.; Fan, C.; Yi, X.; Cheng, B. MALAT1 recruited the E3 ubiquitin ligase FBXW7 to induce CRY2 ubiquitin-mediated degradation and participated in trophoblast migration and invasion. J. Cell. Physiol. 2021, 236, 2169–2177. [Google Scholar] [CrossRef]
- Li, H.; Hou, Y.; Zhang, S.; Zhou, Y.; Wang, D.; Tao, S.; Ni, F. CD49a regulates the function of human decidual natural killer cells. Am. J. Reprod. Immunol. 2019, 81, e13101. [Google Scholar] [CrossRef]
- Sheng, F.; Sun, N.; Ji, Y.; Ma, Y.; Ding, H.; Zhang, Q.; Yang, F.; Li, W. Aberrant expression of imprinted lncRNA MEG8 causes trophoblast dysfunction and abortion. J. Cell. Biochem. 2019, 120, 17378–17390. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Du, G.; Huang, X.; Han, L.; Han, X.; Xu, B.; Zhang, Y.; Yu, M.; Qin, Y.; Xia, Y.; et al. The enhancer RNA lnc-SLC4A1-1 epigenetically regulates unexplained recurrent pregnancy loss (URPL) by activating CXCL8 and NF-kB pathway. EBioMedicine 2018, 38, 162–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, D.; Zeng, H.; Chen, J.; Xiao, L.; Zhao, Y.; Liu, N. H19 regulates trophoblastic spheroid adhesion by competitively binding to let-7. Reproduction 2019, 157, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Yan, H.; Sun, B.; Feng, F.; Chen, P. Decreased expression of long non-coding RNA SNHG7 cause recurrent spontaneous abortion through suppression proliferation and invasion of trophoblast cells via miR-34a. Am. J. Transl. Res. 2019, 11, 463–472. [Google Scholar]
- Li, Z.; Zhou, G.; Tao, F.; Cao, Y.; Han, W.; Li, Q. circ-ZUFSP regulates trophoblasts migration and invasion through sponging miR-203 to regulate STOX1 expression. Biochem. Biophys. Res. Commun. 2020, 531, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.T.; Coca-Prados, M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature 1979, 280, 339–340. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Lasda, E.; Parker, R. Circular RNAs: Diversity of form and function. RNA 2014, 20, 1829–1842. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Huang, C.; Wang, X.; Shan, G. Circular RNAs in Eukaryotic Cells. Curr. Genomics 2015, 16, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Hanan, M.; Soreq, H.; Kadener, S. CircRNAs in the brain. RNA Biol. 2017, 14, 1028–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piwecka, M.; Glazar, P.; Hernandez-Miranda, L.R.; Memczak, S.; Wolf, S.A.; Rybak-Wolf, A.; Filipchyk, A.; Klironomos, F.; Cerda Jara, C.A.; Fenske, P.; et al. Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science 2017, 357, eaam8526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.H.; Xiao, Z.; Tong, J.H.; To, K.F.; Fang, X.; Cheng, A.S.; Chen, Y. EZH2 coupled with HOTAIR to silence MicroRNA-34a by the induction of heterochromatin formation in human pancreatic ductal adenocarcinoma. Int. J. Cancer 2017, 140, 120–129. [Google Scholar] [CrossRef]
- Du, W.W.; Zhang, C.; Yang, W.; Yong, T.; Awan, F.M.; Yang, B.B. Identifying and Characterizing circRNA-Protein Interaction. Theranostics 2017, 7, 4183–4191. [Google Scholar] [CrossRef]
- Dong, W.; Dai, Z.H.; Liu, F.C.; Guo, X.G.; Ge, C.M.; Ding, J.; Liu, H.; Yang, F. The RNA-binding protein RBM3 promotes cell proliferation in hepatocellular carcinoma by regulating circular RNA SCD-circRNA 2 production. EBioMedicine 2019, 45, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell 2017, 66, 9–21.e7. [Google Scholar] [CrossRef] [Green Version]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Gao, Y.; Shang, R.; Sun, W.; Dou, K.; Li, H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, I.; Karedath, T.; Andrews, S.S.; Al-Azwani, I.K.; Mohamoud, Y.A.; Querleu, D.; Rafii, A.; Malek, J.A. Altered expression pattern of circular RNAs in primary and metastatic sites of epithelial ovarian carcinoma. Oncotarget 2016, 7, 36366–36381. [Google Scholar] [CrossRef] [Green Version]
- Xin, Z.; Ma, Q.; Ren, S.; Wang, G.; Li, F. The understanding of circular RNAs as special triggers in carcinogenesis. Brief Funct. Genom. 2017, 16, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, X.; Liu, X.; Liu, X.; He, J.; Ding, Y.; Tong, C.; Peng, C.; Wang, Y.; Gao, R. CircRNA expression profiles in decidual tissue of patients with early recurrent miscarriage. Genes Dis. 2020, 7, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Wang, X.; Ruan, H.; Rui, C.; Mao, P.; Cheng, Q.; Jia, R. Circular RNAs expressed in chorionic villi are probably involved in the occurrence of recurrent spontaneous abortion. Biomed. Pharmacother. 2017, 88, 1154–1162. [Google Scholar] [CrossRef]
- Wu, X.; Sui, Z.; Zhang, H.; Wang, Y.; Yu, Z. Integrated Analysis of lncRNA-Mediated ceRNA Network in Lung Adenocarcinoma. Front. Oncol. 2020, 10, 554759. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.S.; Zhang, E.X.; Sun, Q.F.; Ye, Z.J.; Liu, J.W.; Zhou, D.H.; Tang, Y. Integrated analysis of lncRNA-miRNA-mRNA ceRNA network in squamous cell carcinoma of tongue. BMC Cancer 2019, 19, 779. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Bai, Y.; Yang, X.; Lin, J.; Yang, X.; Wang, D.; He, L.; Zheng, Y.; Zhao, H. Construction and comprehensive analysis of a ceRNA network to reveal potential prognostic biomarkers for hepatocellular carcinoma. Cancer Cell Int. 2019, 19, 90. [Google Scholar] [CrossRef]
- Xu, H.; Zhou, M.; Cao, Y.; Zhang, D.; Han, M.; Gao, X.; Xu, B.; Zhang, A. Genome-wide analysis of long noncoding RNAs, microRNAs, and mRNAs forming a competing endogenous RNA network in repeated implantation failure. Gene 2019, 720, 144056. [Google Scholar] [CrossRef]
- Huang, Y.; Hao, J.; Liao, Y.; Zhou, L.; Wang, K.; Zou, H.; Hu, Y.; Li, J. Transcriptome sequencing identified the ceRNA network associated with recurrent spontaneous abortion. BMC Med. Genomics 2021, 14, 278. [Google Scholar] [CrossRef]
- Zang, X.; Gu, T.; Wang, W.; Zhou, C.; Ding, Y.; Gu, S.; Xu, Z.; Xie, Y.; Li, Z.; Cai, G.; et al. Integrated Insight into the Molecular Mechanisms of Spontaneous Abortion during Early Pregnancy in Pigs. Int. J. Mol. Sci. 2021, 22, 6644. [Google Scholar] [CrossRef]
- Wang, M.M.; Zhong, J.X.; Xiang, Y.Y. LncRNA-GAS5 related to the processes of recurrent pregnancy loss by regulating Th1/Th2 balance. Kaohsiung J. Med. Sci. 2021, 37, 479–486. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, Y.; Li, J.; Mao, B. Urothelial cancer associated 1 (UCA1) regulates trophoblast viability, proliferation, and migration via modulating the UCA1/miR-455/RUNX2 signaling pathway. Acta Biochim. Biophys. Sin. 2020, 52, 1120–1130. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Hansen, T.B.; Veno, M.T.; Kjems, J. Circular RNAs in cancer: Opportunities and challenges in the field. Oncogene 2018, 37, 555–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Li, Y.; Liu, W.; He, J.; Zhang, L.; Li, H.; Li, P.; Lv, L. Comprehensive circRNA expression profile and construction of circRNA-associated ceRNA network in fur skin. Exp. Dermatol. 2018, 27, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Gu, T.; He, Y.; Zhou, C.; Hu, Q.; Wang, X.; Zheng, E.; Huang, S.; Xu, Z.; Yang, J.; et al. Genome-Wide Analysis of Circular RNAs Mediated ceRNA Regulation in Porcine Embryonic Muscle Development. Front. Cell Dev. Biol. 2019, 7, 289. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Chen, L.; Shan, Y.; Chen, G.; Chu, Y.; Dai, H.; Liu, X.; Bao, H. Hsa_circ_0038383-mediated competitive endogenous RNA network in recurrent implantation failure. Aging 2021, 13, 6076–6090. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Zhao, H.; Gong, L.; Xiao, X.; Zhou, Q.; Lu, H.; Cui, Y.; Xu, H.; Wu, S.; Tang, Y.; et al. Differentially expressed circular RNAs and the competing endogenous RNA network associated with preeclampsia. Placenta 2021, 103, 232–241. [Google Scholar] [CrossRef]
- Jin, D.; Wu, X.; Yu, H.; Jiang, L.; Zhou, P.; Yao, X.; Meng, J.; Wang, L.; Zhang, M.; Zhang, Y. Systematic analysis of lncRNAs, mRNAs, circRNAs and miRNAs in patients with postmenopausal osteoporosis. Am. J. Transl. Res. 2018, 10, 1498–1510. [Google Scholar]
- Chen, D.; He, B.; Zheng, P.; Wang, S.; Zhao, X.; Liu, J.; Yang, X.; Cheng, W. Identification of mRNA-, circRNA- and lncRNA- Associated ceRNA Networks and Potential Biomarkers for Preeclampsia From Umbilical Vein Endothelial Cells. Front. Mol. Biosci. 2021, 8, 652250. [Google Scholar] [CrossRef]
- Prather, R.S.; Lorson, M.; Ross, J.W.; Whyte, J.J.; Walters, E. Genetically engineered pig models for human diseases. Annu Rev Anim. Biosci. 2013, 1, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Jia, N.; Li, J. Noncoding RNAs in Unexplained Recurrent Spontaneous Abortions and Their Diagnostic Potential. Dis. Markers 2019, 2019, 7090767. [Google Scholar] [CrossRef] [Green Version]
- Viereck, J.; Thum, T. Circulating Noncoding RNAs as Biomarkers of Cardiovascular Disease and Injury. Circ. Res. 2017, 120, 381–399. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Etiology of SA. It is currently known that the factors affecting SA are genetic factors, physiological factors, environmental factors, and immune factors. Among them, genetic factors are the most crucial etiology of SA. EMT, epithelial-mesenchymal transformation.
Figure 1.
Etiology of SA. It is currently known that the factors affecting SA are genetic factors, physiological factors, environmental factors, and immune factors. Among them, genetic factors are the most crucial etiology of SA. EMT, epithelial-mesenchymal transformation.

Figure 2.
The mechanism for lncRNA to participate in SA. There are two main ways lncRNAs participate in the occurrence and development of SA: (1) directly regulate the expression of target mRNA; (2) regulate the transcription and expression of downstream target mRNA by targeting miRNAs. “?” represents our hypothesis for this regulatory pathway according to existing research.
Figure 2.
The mechanism for lncRNA to participate in SA. There are two main ways lncRNAs participate in the occurrence and development of SA: (1) directly regulate the expression of target mRNA; (2) regulate the transcription and expression of downstream target mRNA by targeting miRNAs. “?” represents our hypothesis for this regulatory pathway according to existing research.
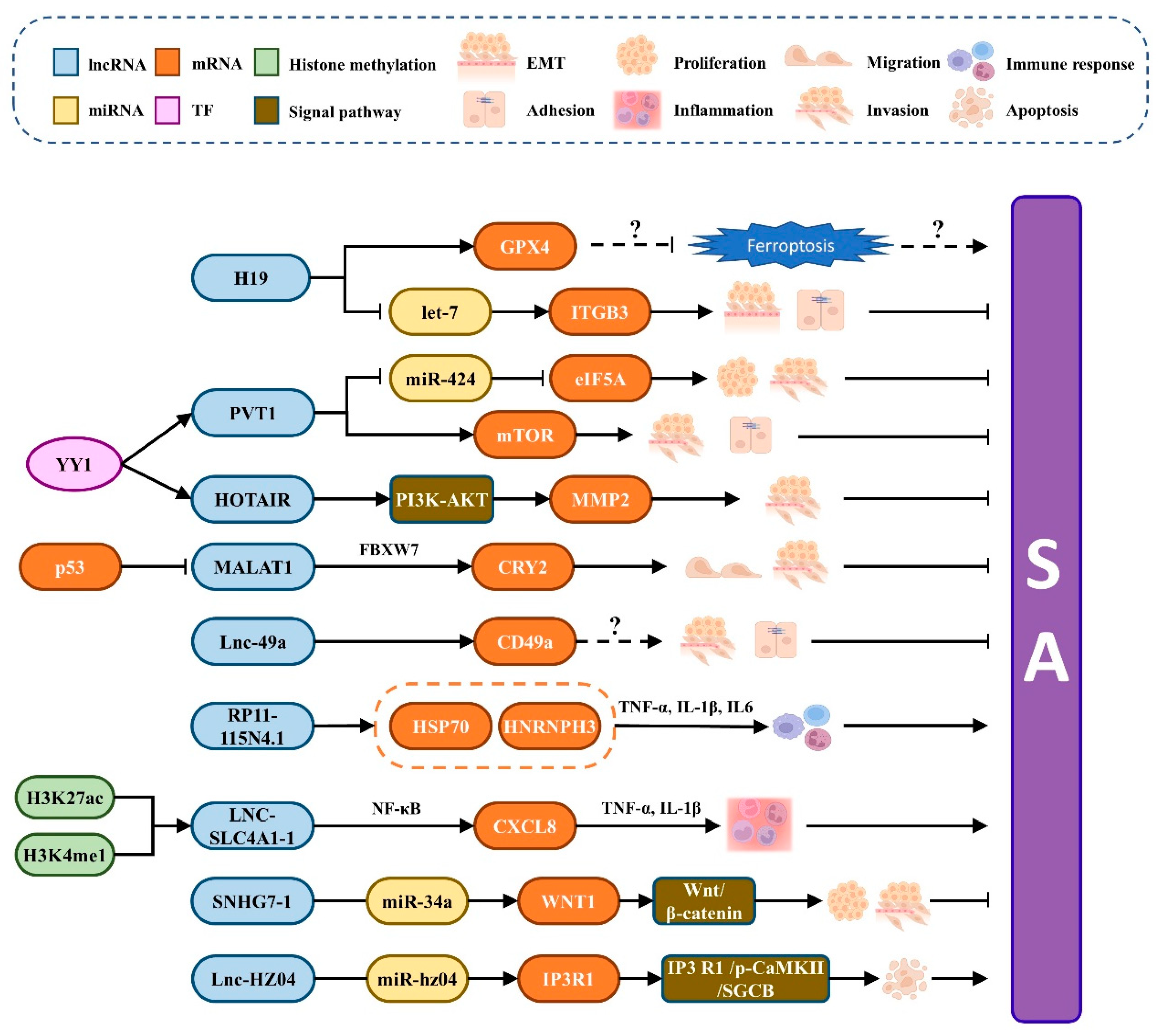
Figure 3.
Regulatory mechanism of ceRNA network. (A) Relationship of mRNA, miRNA, circRNA, and lncRNA in ceRNA network hypothesis; (B) specific regulation mechanism of lncRNA-mediated ceRNA network. LncRNA regulates the expression of mRNA by competing for the opportunity of binding to miRNA; (C) specific regulation mechanism when lncRNA and circRNA are as ceRNA simultaneously. lncRNA and circRNA simultaneously compete with mRNA for the opportunity to bind to miRNA to regulate mRNA expression. LncRNA, circRNA, and mRNA compete with each other.
Figure 3.
Regulatory mechanism of ceRNA network. (A) Relationship of mRNA, miRNA, circRNA, and lncRNA in ceRNA network hypothesis; (B) specific regulation mechanism of lncRNA-mediated ceRNA network. LncRNA regulates the expression of mRNA by competing for the opportunity of binding to miRNA; (C) specific regulation mechanism when lncRNA and circRNA are as ceRNA simultaneously. lncRNA and circRNA simultaneously compete with mRNA for the opportunity to bind to miRNA to regulate mRNA expression. LncRNA, circRNA, and mRNA compete with each other.


{kind=link}
{kind=link}
{kind=link}
Table 1.
miRNAs involved in regulating SA genesis and development.
| miRNA Names | Expression in SA | Target | Tissue | Function | Reference |
|---|---|---|---|---|---|
| MiR-93 | Upregulation | BCL2L2 | Chorionic villi, trophoblast | Promotes proliferation, migration, invasion, and apoptosis | [40] |
| MiR-19b | Upregulation | PTEN | Placental villi | Regulates abnormal cellular invasion and apoptosis | [41] |
| MiR-494 | Downregulation | PTEN | Placental villi | Regulates abnormal cellular invasion and apoptosis | [41] |
| MiR-365 | Upregulation | SGK1 | Decidua | Regulates cellular apoptosis | [43] |
| MiR-199B-5p | Upregulation | SGK1 | Decidua | Regulates cellular apoptosis | [44] |
| MiR-520 | Upregulation | PARP1 | Trophoblast | Promotes DNA damage-induced apoptosis | [45] |
| MiR-27A-3p | Upregulation | USP25 | Trophoblast | Regulates the process of EMT, cell invasion, and migration | [48] |
| MiR-16 | Upregulation | VEGF | Placenta | Regulates angiogenesis and development | [49] |
| MiR-219a | Downregulation | VEGFR2 | Trophoblast | Negative regulation of cellular proliferation and invasion | [26] |
| MiR-155-5p | Downregulation | NF-kB pathway | Decidual stroma | Promotes cellular growth and proliferation and inhibits apoptosis | [25] |
| MiR-34a-3p/5p | Upregulation | P53 | NK cells | Regulates cellular apoptosis and proliferation | [57,58] |
| MiR-141-3p/5p | Upregulation | PTEN | NK cells | Regulates cellular apoptosis and proliferation | [57,61] |
| MiR-24 | Downregulation | IFNG | NK cells | Regulates cellular apoptosis and proliferation | [57,59,60] |
| Hsa-miR-5787 | Upregulation | EIF5 | NK cells | Inhibits cellular growth | [62,63] |
| MiR-103 | Downregulation | STAT1/IRF1 pathway | M1 macrophages | Negative regulation of cellular polarization | [64] |
| MiR-146a/b-5p | Upregulation | TRAF6 | M1 macrophages | Inhibits EMT and maintains cellular interaction | [65] |
Table 2.
lncRNAs and circRNAs involved in the occurrence of SA.
| RNA Names | Expression in SA | Target | Tissue | Function | Reference |
|---|---|---|---|---|---|
| H19 | Downregulation | GPX4 | Placental villi | Regulates invasion and migration, and may be related to ferroptosis. | [80] |
| Let-7 | Competes with miRNA let-7 to control the target gene ITGB3 | [93,94] | |||
| PVT1 | Downregulation | MTOR | Trophoblast | Control apoptosis, adhesion, and invasion through the mTOR pathway. | [82,83] |
| MiR-424 | Regulate the function of trophoblast cells through the PVT1/miR-424/eIF5A pathway | [83] | |||
| HOTAIR | Downregulation | PI3K-AKT | Trophoblast | Promotes the production of matrix metalloproteinase 2 and enhances cell invasion | [84,85] |
| MALAT1 | Downregulation | FBXW7 | Trophoblast | Inhibits SA by playing a key role in the P53-MALAT1-FBXW7-CRY2 axis. | [88,89] |
| Lnc-49a | Downregulation | CD49a | DNK cells | Impedes SA by regulating the expression of CD49a | [90] |
| RP11-115N4.1 | Upregulation | PARP1 | Peripheral blood | Inhibits the migration of trophoblast cells by increasing HSP70 and HNRNPH3. | [74] |
| MEG8 | Upregulation | Unknown | Chorion villi | Inhibits cell proliferation and invasion in trophoblast cell lines | [91] |
| LNC-SLC4A1-1 | Upregulation | CXCL8 | URSA Villi | Intensifies trophoblast cell inflammation by activating CXCL8 | [92] |
| SNHG7-1 | Upregulation | MiR-34a | Placental villi | Promotes the occur-rence of RSA via the Wnt/β-catenin signaling pathway | [95] |
| Lnc-HZ04 | Upregulation | MiR-hz04 | Villi | Weakens the decreasing effect of miR-hz04 on inositol IP3R1 expression and stability | [28] |
| CIRC-ZUFSP | Upregulation | MiR-203 | Trophoblast | Inhibits migration and invasion of trophoblast cells through the CIRC-ZUFSP/miR-203/STOX1 pathway | [96] |
| CIRCPUM1 | Downregulation | MiR-30a-5p | Placenta | Promotes trophoblast cell processes and anti-inflammatory effects through the miR-30a-5p/JunB axis | [31] |
| CircFOXP1 | Downregulation | MiR-143-3p | Trophoblast | Regulates the function of trophoblast cells via the miR-143-3p/S100A11 pathway | [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gan, J.; Gu, T.; Yang, H.; Ao, Z.; Cai, G.; Hong, L.; Wu, Z. Non-Coding RNAs Regulate Spontaneous Abortion: A Global Network and System Perspective. Int. J. Mol. Sci. 2022, 23, 4214. https://doi.org/10.3390/ijms23084214
AMA Style
Gan J, Gu T, Yang H, Ao Z, Cai G, Hong L, Wu Z. Non-Coding RNAs Regulate Spontaneous Abortion: A Global Network and System Perspective. International Journal of Molecular Sciences. 2022; 23(8):4214. https://doi.org/10.3390/ijms23084214
Chicago/Turabian StyleGan, Jianyu, Ting Gu, Huaqiang Yang, Zheng Ao, Gengyuan Cai, Linjun Hong, and Zhenfang Wu. 2022. "Non-Coding RNAs Regulate Spontaneous Abortion: A Global Network and System Perspective" International Journal of Molecular Sciences 23, no. 8: 4214. https://doi.org/10.3390/ijms23084214
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.