
Early Inactivation of Membrane Estrogen Receptor Alpha (ERα) Recapitulates the Endothelial Dysfunction of Aged Mouse Resistance Arteries

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Mice Age, Heart Weight, Body Weight, Blood Pressure and Heart Rate
2.2. FMD in Mouse Mesenteric Arteries
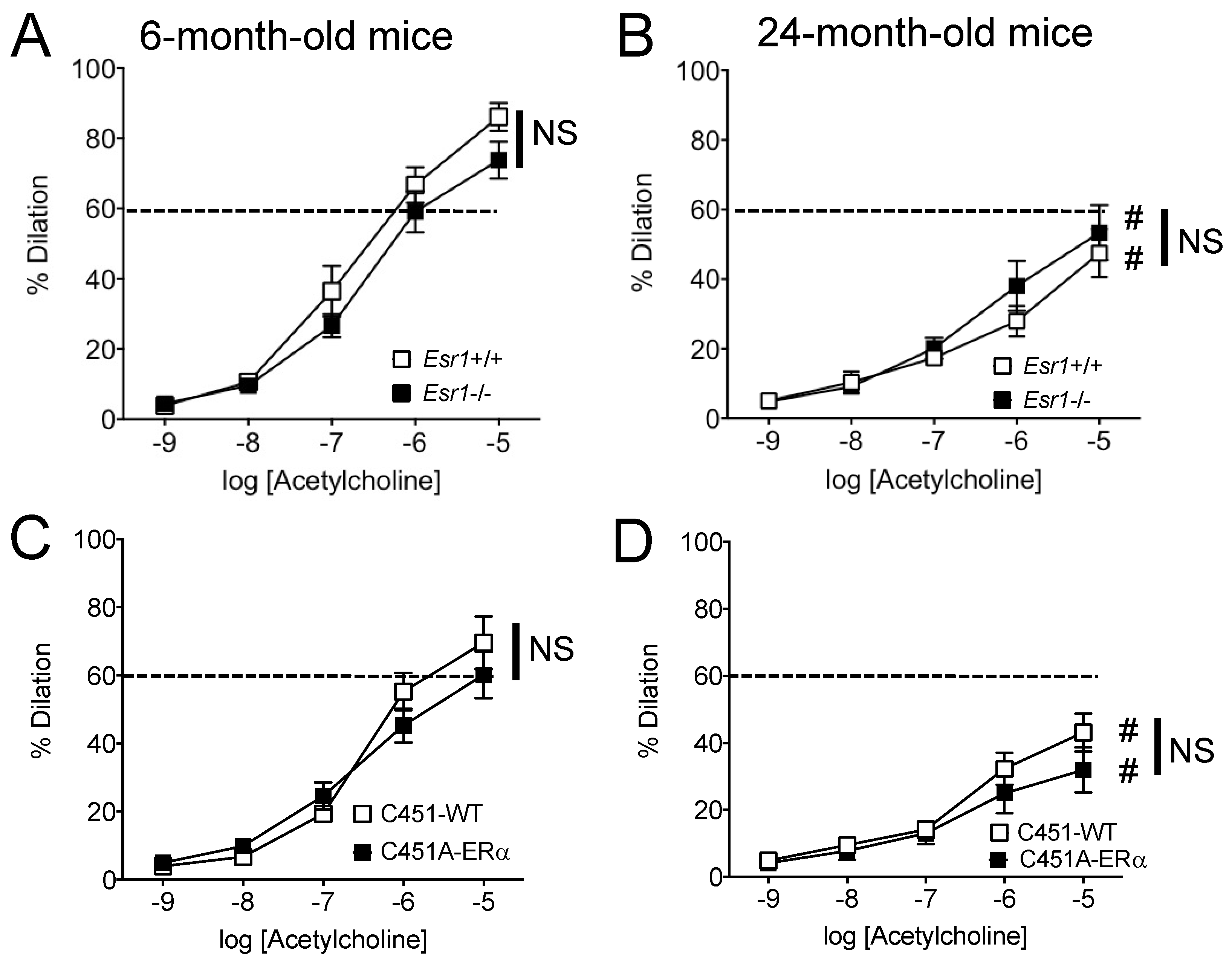
2.3. Agonist-Mediated Endothelium-Dependent Dilation in Mouse Mesenteric Arteries
2.4. Smooth Muscle-Dependent Contraction in Mouse Mesenteric Arteries
2.5. Wall Structure and Properties of Mouse Mesenteric Arteries
2.6. Gene Expression Analysis of the Main Pathways Involved in FMD
2.7. Effect of the Reduction in Oxidative Stress on FMD in Mouse Mesenteric Arteries
2.8. Effect of Age on Estradiol (E2)-Mediated Dilation in Mouse Mesenteric Arteries
3. Discussion
4. Materials and Methods
4.1. Animal Protocol
4.2. Flow-Mediated Dilation in Mesenteric Arteries In Vitro
4.3. Quantitative Real-Time PCR
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rizzoni, D.; Rosei, E.A. Small artery remodeling in hypertension and diabetes. Curr. Hypertens. Rep. 2006, 8, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Stoner, L.; Sabatier, M.J. Use of ultrasound for non-invasive assessment of flow-mediated dilation. J. Atheroscler. Thromb. 2012, 19, 407–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chehaitly, A.; Vessieres, E.; Guihot, A.-L.; Henrion, D. Flow-mediated outward arterial remodeling in aging. Mech. Ageing Dev. 2020, 194, 111416. [Google Scholar] [CrossRef] [PubMed]
- Königstein, K.; Wagner, J.; Frei, M.; Knaier, R.; Klenk, C.; Carrard, J.; Schwarz, A.; Hinrichs, T.; Schmidt-Trucksäss, A. Endothelial function of healthy adults from 20 to 91 years of age: Prediction of cardiovascular risk by vasoactive range. J. Hypertens. 2021, 39, 1361–1369. [Google Scholar] [CrossRef]
- Majerczak, J.; Grandys, M.; Frołow, M.; Szkutnik, Z.; Zakrzewska, A.; Niżankowski, R.; Duda, K.; Chlopicki, S.; Zoladz, J.A. Age-Dependent Impairment in Endothelial Function and Arterial Stiffness in Former High Class Male Athletes Is No Different to That in Men With No History of Physical Training. J. Am. Heart Assoc. 2019, 8, e012670. [Google Scholar] [CrossRef] [PubMed]
- Lane-Cordova, A.D.; Ranadive, S.M.; Kappus, R.M.; Cook, M.D.; Phillips, S.A.; Woods, J.A.; Wilund, K.R.; Baynard, T.; Fernhall, B. Aging, not age-associated inflammation, determines blood pressure and endothelial responses to acute inflammation. J. Hypertens. 2016, 34, 2402–2409. [Google Scholar] [CrossRef]
- Black, M.A.; Cable, N.; Thijssen, D.H.J.; Green, D.J. Impact of age, sex, and exercise on brachial artery flow-mediated dilatation. Am. J. Physiol. Circ. Physiol. 2009, 297, H1109–H1116. [Google Scholar] [CrossRef]
- Deanfield, J.E.; Halcox, J.P.; Rabelink, T.J. Endothelial function and dysfunction: Testing and clinical relevance. Circulation 2007, 115, 1285–1295. [Google Scholar] [CrossRef]
- Beyer, A.M.; Zinkevich, N.; Miller, B.; Liu, Y.; Wittenburg, A.L.; Mitchell, M.; Galdieri, R.; Sorokin, A.; Gutterman, D.D. Transition in the mechanism of flow-mediated dilation with aging and development of coronary artery disease. Basic Res. Cardiol. 2017, 112, 5. [Google Scholar] [CrossRef]
- Favre, J.; Vessieres, E.; Guihot, A.-L.; Proux, C.; Grimaud, L.; Rivron, J.; Garcia, M.C.; Réthoré, L.; Zahreddine, R.; Davezac, M.; et al. Membrane estrogen receptor alpha (ERα) participates in flow-mediated dilation in a ligand-independent manner. eLife 2021, 10, e68695. [Google Scholar] [CrossRef]
- Smith, E.P.; Boyd, J.; Frank, G.R.; Takahashi, H.; Cohen, R.M.; Specker, B.; Williams, T.C.; Lubahn, D.B.; Korach, K. Estrogen Resistance Caused by a Mutation in the Estrogen-Receptor Gene in a Man. N. Engl. J. Med. 1994, 331, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- Sudhir, K.; Chou, T.M.; Messina, L.M.; Hutchison, S.J.; Korach, K.; Chatterjee, K.; Rubanyi, G.M. Endothelial dysfunction in a man with disruptive mutation in oestrogen-receptor gene. Lancet 1997, 349, 1146–1147. [Google Scholar] [CrossRef]
- Arnal, J.F.; Lenfant, F.; Metivier, R.; Flouriot, G.; Henrion, D.; Adlanmerini, M.; Fontaine, C.; Gourdy, P.; Chambon, P.; Katzenellenbogen, B.; et al. Membrane and Nuclear Estrogen Receptor Alpha Actions: From Tissue Specificity to Medical Implications. Physiol. Rev. 2017, 97, 1045–1087. [Google Scholar] [CrossRef] [PubMed]
- Adlanmerini, M.; Solinhac, R.; Abot, A.; Fabre, A.; Letron, I.R.; Guihot, A.-L.; Boudou, F.; Sautier, L.; Vessières, E.; Kim, S.H.; et al. Mutation of the palmitoylation site of estrogen receptor α in vivo reveals tissue-specific roles for membrane versus nuclear actions. Proc. Natl. Acad. Sci. USA 2014, 111, E283–E290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guivarc’H, E.; Favre, J.; Guihot, A.; Vessières, E.; Grimaud, L.; Proux, C.; Rivron, J.; Barbelivien, A.; Fassot, C.; Briet, M.; et al. Nuclear Activation Function 2 Estrogen Receptor α Attenuates Arterial and Renal Alterations Due to Aging and Hypertension in Female Mice. J. Am. Heart Assoc. 2020, 9, e013895. [Google Scholar] [CrossRef]
- Davezac, M.; Buscato, M.; Zahreddine, R.; Lacolley, P.; Henrion, D.; Lenfant, F.; Arnal, J.-F.; Fontaine, C. Estrogen Receptor and Vascular Aging. Front. Aging 2021, 2, 727380. [Google Scholar] [CrossRef]
- Levy, B.I.; Schiffrin, E.; Mourad, J.-J.; Agostini, D.; Vicaut, E.; Safar, M.E.; Struijker-Boudier, H.A. Impaired tissue perfusion: A pathology common to hypertension, obesity, and diabetes mellitus. Circulation 2008, 118, 968–976. [Google Scholar] [CrossRef]
- Patel, A.; Honoré, E. Polycystins and renovascular mechanosensory transduction. Nat. Rev. Nephrol. 2010, 6, 530–538. [Google Scholar] [CrossRef]
- Dunn, K.M.; Nelson, M.T. Neurovascular signaling in the brain and the pathological consequences of hypertension. Am. J. Physiol. Circ. Physiol. 2014, 306, H1–H14. [Google Scholar] [CrossRef] [Green Version]
- Struijker-Boudier, H.A.; Rosei, A.E.; Bruneval, P.; Camici, P.G.; Christ, F.; Henrion, D.; Lévy, B.I.; Pries, A.; Vanoverschelde, J.-L. Evaluation of the microcirculation in hypertension and cardiovascular disease. Eur. Heart J. 2007, 28, 2834–2840. [Google Scholar] [CrossRef]
- Celermajer, D.S.; Sorensen, K.E.; Spiegelhalter, D.J.; Georgakopoulos, D.; Robinson, J.; Deanfield, J.E. Aging is associated with endothelial dysfunction in healthy men years before the age-related decline in women. J. Am. Coll. Cardiol. 1994, 24, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Csiszar, A.; Ungvari, Z.; Edwards, J.G.; Kaminski, P.; Wolin, M.S.; Koller, A.; Kaley, G. Aging-Induced Phenotypic Changes and Oxidative Stress Impair Coronary Arteriolar Function. Circ. Res. 2002, 90, 1159–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungvari, Z.; Tarantini, S.; Kiss, T.; Wren, J.D.; Giles, C.B.; Griffin, C.T.; Murfee, W.L.; Pacher, P.; Csiszar, A. Endothelial dysfunction and angiogenesis impairment in the ageing vasculature. Nat. Rev. Cardiol. 2018, 15, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Moreau, K.L.; Hildreth, K.L.; Klawitter, J.; Blatchford, P.; Kohrt, W.M. Decline in endothelial function across the menopause transition in healthy women is related to decreased estradiol and increased oxidative stress. GeroScience 2020, 42, 1699–1714. [Google Scholar] [CrossRef]
- Gros, R.; Van Wert, R.; You, X.; Thorin, E.; Husain, M. Effects of age, gender, and blood pressure on myogenic responses of mesenteric arteries from C57BL/6 mice. Am. J. Physiol. Circ. Physiol. 2002, 282, H380–H388. [Google Scholar] [CrossRef]
- Shipley, R.D.; Muller-Delp, J.M. Aging decreases vasoconstrictor responses of coronary resistance arterioles through endothelium-dependent mechanisms. Cardiovasc. Res. 2005, 66, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.S.; Kim, S.; Dominguez, J.M., 2nd; Sindler, A.L.; Dick, G.M.; Muller-Delp, J.M. Aging and muscle fiber type alter K+ channel contributions to the myogenic response in skeletal muscle arterioles. J. Appl. Physiol. 2009, 107, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Jelinic, M.; Tare, M.; Conrad, K.P.; Parry, L.J. Differential effects of relaxin deficiency on vascular aging in arteries of male mice. Age 2015, 37, 66. [Google Scholar] [CrossRef] [Green Version]
- Lacolley, P.; Regnault, V.; Segers, P.; Laurent, S. Vascular Smooth Muscle Cells and Arterial Stiffening: Relevance in Development, Aging, and Disease. Physiol. Rev. 2017, 97, 1555–1617. [Google Scholar] [CrossRef]
- Avolio, A.P.; Kuznetsova, T.; Heyndrickx, G.R.; Kerkhof, P.L.M.; Li, J.K.-J. Arterial Flow, Pulse Pressure and Pulse Wave Velocity in Men and Women at Various Ages. Adv. Exp. Med. Biol. 2018, 1065, 153–168. [Google Scholar] [CrossRef]
- Tarhouni, K.; Guihot, A.L.; Freidja, M.L.; Toutain, B.; Henrion, B.; Baufreton, C.; Pinaud, F.; Procaccio, V.; Grimaud, L.; Ayer, A.; et al. Key Role of Estrogens and Endothelial Estrogen Receptor α in Blood Flow–Mediated Remodeling of Resistance Arteries. Arter. Thromb. Vasc. Biol. 2013, 33, 605–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarhouni, K.; Freidja, M.L.; Guihot, A.L.; Vessieres, E.; Grimaud, L.; Toutain, B.; Lenfant, F.; Arnal, J.F.; Loufrani, L.; Henrion, D. Role of estrogens and age in flow-mediated outward remodeling of rat mesenteric resistance arteries. Am. J. Physiol. Circ. Physiol. 2014, 307, H504–H514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarhouni, K.; Guihot, A.-L.; Vessières, E.; Toutain, B.; Procaccio, V.; Grimaud, L.; Loufrani, L.; Lenfant, F.; Arnal, J.-F.; Henrion, D. Determinants of Flow-Mediated Outward Remodeling in Female Rodents: Respective roles of age, estrogens, and timing. Arter. Thromb. Vasc. Biol. 2014, 34, 1281–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guivarc’H, E.; Buscato, M.; Guihot, A.; Favre, J.; Vessières, E.; Grimaud, L.; Wakim, J.; Melhem, N.; Zahreddine, R.; Adlanmerini, M.; et al. Predominant Role of Nuclear Versus Membrane Estrogen Receptor α in Arterial Protection: Implications for Estrogen Receptor α Modulation in Cardiovascular Prevention/Safety. J. Am. Heart Assoc. 2018, 7, e008950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvestre, J.-S.; Bergaya, S.; Tamarat, R.; Duriez, M.; Boulanger, C.M.; Levy, B.I. Proangiogenic Effect of Angiotensin-Converting Enzyme Inhibition Is Mediated by the Bradykinin B 2 Receptor Pathway. Circ. Res. 2001, 89, 678–683. [Google Scholar] [CrossRef] [Green Version]
- Adlanmerini, M.; Fébrissy, C.; Zahreddine, R.; Vessières, E.; Buscato, M.; Solinhac, R.; Favre, J.; Anquetil, T.; Guihot, A.-L.; Boudou, F.; et al. Mutation of Arginine 264 on ERα (Estrogen Receptor Alpha) Selectively Abrogates the Rapid Signaling of Estradiol in the Endothelium Without Altering Fertility. Arter. Thromb. Vasc. Biol. 2020, 40, 2143–2158. [Google Scholar] [CrossRef]
- Kublickiene, K.; Svedas, E.; Landgren, B.-M.; Crisby, M.; Nahar, N.; Nisell, H.; Poston, L. Small Artery Endothelial Dysfunction in Postmenopausal Women:In VitroFunction, Morphology, and Modification by Estrogen and Selective Estrogen Receptor Modulators. J. Clin. Endocrinol. Metab. 2005, 90, 6113–6122. [Google Scholar] [CrossRef] [Green Version]
- Kublickiene, K.; Fu, X.-D.; Svedas, E.; Landgren, B.-M.; Genazzani, A.R.; Simoncini, T. Effects in Postmenopausal Women of Estradiol and Medroxyprogesterone Alone and Combined on Resistance Artery Function and Endothelial Morphology and Movement. J. Clin. Endocrinol. Metab. 2008, 93, 1874–1883. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, A.J.; Reyes, R.; Kang, L.S.; Dailey, R.A.; Stallone, J.N.; Moningka, N.C.; Muller-Delp, J.M. Estrogen replacement restores flow-induced vasodilation in coronary arterioles of aged and ovariectomized rats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2009, 297, R1713–R1723. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, J.C.; Rodriguez-Miguelez, P.; Zimmerman, M.A.; Harris, R.A. Differences in angiotensin (1–7) between men and women. Am. J. Physiol. Circ. Physiol. 2015, 308, H1171–H1176. [Google Scholar] [CrossRef]
- Sudhir, K.; Chou, T.M.; Chatterjee, K.; Smith, E.P.; Williams, T.C.; Kane, J.P.; Malloy, M.J.; Korach, K.S.; Rubanyi, G.M. Premature coronary artery disease associated with a disruptive mutation in the estrogen receptor gene in a man. Circulation 1997, 96, 3774–3777. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.; Filardo, E.J.; Lolait, S.J.; Thomas, P.; Maggiolini, M.; Prossnitz, E.R. Twenty years of the G protein-coupled estrogen receptor GPER: Historical and personal perspectives. J. Steroid Biochem. Mol. Biol. 2018, 176, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredette, N.C.; Meyer, M.R.; Prossnitz, E.R. Role of GPER in estrogen-dependent nitric oxide formation and vasodilation. J. Steroid Biochem. Mol. Biol. 2017, 176, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prossnitz, E.R. GPER modulators: Opportunity Nox on the heels of a class Akt. J. Steroid Biochem. Mol. Biol. 2018, 176, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.R.; Fredette, N.C.; Daniel, C.; Sharma, G.; Amann, K.; Arterburn, J.B.; Barton, M.; Prossnitz, E.R. Obligatory role for GPER in cardiovascular aging and disease. Sci. Signal. 2016, 9, ra105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.R.; Rosemann, T.; Barton, M.; Prossnitz, E.R. GPER Mediates Functional Endothelial Aging in Renal Arteries. Pharmacology 2017, 100, 188–193. [Google Scholar] [CrossRef]
- Valera, M.-C.; Gourdy, P.; Trémollières, F.; Arnal, J.-F. From the Women’s Health Initiative to the combination of estrogen and selective estrogen receptor modulators to avoid progestin addition. Maturitas 2015, 82, 274–277. [Google Scholar] [CrossRef]
- Gu, L.; Robinson, R.A.S. Proteomic approaches to quantify cysteine reversible modifications in aging and neurodegenerative diseases. Proteom.-Clin. Appl. 2016, 10, 1159–1177. [Google Scholar] [CrossRef] [Green Version]
- Novella, S.; Dantas, A.P.; Segarra, G.; Medina, P.; Hermenegildo, C. Vascular Aging in Women: Is Estrogen the Fountain of Youth? Front. Physiol. 2012, 3, 165. [Google Scholar] [CrossRef] [Green Version]
- Brandes, R.P.; Fleming, I.; Busse, R. Endothelial aging. Cardiovasc. Res. 2005, 66, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Li, Y.-S.; Chien, S. Shear Stress–Initiated Signaling and Its Regulation of Endothelial Function. Arter. Thromb. Vasc. Biol. 2014, 34, 2191–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souilhol, C.; Serbanovic-Canic, J.; Fragiadaki, M.; Chico, T.J.; Ridger, V.; Roddie, H.; Evans, P.C. Endothelial responses to shear stress in atherosclerosis: A novel role for developmental genes. Nat. Rev. Cardiol. 2020, 17, 52–63. [Google Scholar] [CrossRef] [PubMed]
- López-Lluch, G.; Irusta, P.M.; Navas, P.; de Cabo, R. Mitochondrial biogenesis and healthy aging. Exp. Gerontol. 2008, 43, 813–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretón-Romero, R.; Acín-Perez, R.; Rodríguez-Pascual, F.; Martínez-Molledo, M.; Brandes, R.P.; Rial, E.; Enríquez, J.A.; Lamas, S. Laminar shear stress regulates mitochondrial dynamics, bioenergetics responses and PRX3 activation in endothelial cells. Biochim. Biophys. Acta 2014, 1843, 2403–2413. [Google Scholar] [CrossRef] [Green Version]
- Scheitlin, C.G.; Julian, J.A.; Shanmughapriya, S.; Madesh, M.; Tsoukias, N.M.; Alevriadou, B.R. Endothelial mitochondria regulate the intracellular Ca2+ response to fluid shear stress. Am. J. Physiol.-Cell Physiol. 2016, 310, C479–C490. [Google Scholar] [CrossRef] [Green Version]
- Razmara, A.; Sunday, L.; Stirone, C.; Wang, X.B.; Krause, D.N.; Duckles, S.P.; Procaccio, V. Mitochondrial Effects of Estrogen Are Mediated by Estrogen Receptor α in Brain Endothelial Cells. J. Pharmacol. Exp. Ther. 2008, 325, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Antal, M.C.; Krust, A.; Chambon, P.; Mark, M. Sterility and absence of histopathological defects in nonreproductive organs of a mouse ERbeta-null mutant. Proc. Natl. Acad. Sci. USA 2008, 105, 2433–2438. [Google Scholar] [CrossRef] [Green Version]
- Iglarz, M.; Matrougui, K.; Lévy, B.I.; Henrion, D. Chronic blockade of endothelin ETA receptors improves flow dependent dilation in resistance arteries of hypertensive rats. Cardiovasc. Res. 1998, 39, 657–664. [Google Scholar] [CrossRef] [Green Version]
- Henrion, D.; Terzi, F.; Matrougui, K.; Duriez, M.; Boulanger, C.M.; Colucci-Guyon, E.; Babinet, C.; Briand, P.; Friedlander, G.; Poitevin, P.; et al. Impaired flow-induced dilation in mesenteric resistance arteries from mice lacking vimentin. J. Clin. Investig. 1997, 100, 2909–2914. [Google Scholar] [CrossRef]
- Bouvet, C.; de Chantemele, E.B.; Guihot, A.-L.; Vessières, E.; Bocquet, A.; Dumont, O.; Jardel, A.; Loufrani, L.; Moreau, P.; Henrion, D. Flow-Induced Remodeling in Resistance Arteries from Obese Zucker Rats Is Associated with Endothelial Dysfunction. Hypertension 2007, 50, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Freed, J.K.; Beyer, A.M.; Logiudice, J.A.; Hockenberry, J.C.; Gutterman, D.D. Ceramide Changes the Mediator of Flow-Induced Vasodilation from Nitric Oxide to Hydrogen Peroxide in the Human Microcirculation. Circ. Res. 2014, 115, 525–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauffenstein, G.; Tamareille, S.; Prunier, F.; Roy, C.; Ayer, A.; Toutain, B.; Billaud, M.; Isakson, B.E.; Grimaud, L.; Loufrani, L.; et al. Central Role of P2Y 6 UDP Receptor in Arteriolar Myogenic Tone. Arter. Thromb. Vasc. Biol. 2016, 36, 1598–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Protein | NCBI Reference Sequence | Forward Sequence (5’–3’) | Reverse Sequence (5’–3’) |
|---|---|---|---|---|
| Ace | Ace | NM_009598.2 | GGAACAAGTCGATGTTAGAGAAGC | ACAGAGGTACACTGCTTGATCCT |
| ActB | Beta-actin | NM_007393.1 | GCCGGGACCTGACTGACTAC | TTCTCCTTAATGTCACGCACGAT |
| Agtr1b | AT1R (b) | NM_175086.3 | GTGACATGATCCCCTGACAGT | AGTGAGTGAACTGTCTAGCTAAATGC |
| Ahr | Transcription factor cyp1b1 | NM_001314027.1 | GATGCCAAAGGGCAGCTTATTC | CCACCTCCAGCGACTGTGTTT |
| Ar | Androgen receptor | NM_013476.4 | CCAGTCCCAATTGTGTCAAA | TCCCTGGTACTGTCCAAACG |
| Cav1 | Caveolin1 | NM_007616.2 | AACGACGACGTGGTCAAGA | CACAGTGAAGGTGGTGAAGC |
| Comt | Catechol O-methyltransferase | NM_001111062.1 | CCGCTACCTTCCAGACACAC | GTTCCCGGGACAATGACA |
| Cyp1b1 | Cytochrome P450 1b1 | NM_009994.1 | AGCCAGGACACCCTTTCC | CCTGAACATCCGGGTATCTG |
| Dnm1l | Drp1 | NM_001276340.1 | AGATCGTCGTAGTGGGAACG | CCACTAGGCTTTCCAGCACT |
| Entpd1 | NtPDase1 | NM_009848.3 | CTCCTGCAAGGCTATAACTTCAC | GCGTTGCTGTCTTTGATCTTG |
| Esr1 | ER alpha | MN_007956.4 | GCTCCTAACTTGCTCCTGGAC | CAGCAACATGTCAAAGATCTCC |
| Gja4 | Cx37 | NM_008120.3 | TCCTGGGAAAAAGCACTGAT | CTGTGTCTGTCCAGGTGACG |
| Gja5 | Cx40 | NM_008121.2 | CAGTGTGATCCTCCTTTTAGGG | TTTCCTGCCTCACACTCCTT |
| Gpx1 | gPx-1 | NM_008160.6 | TTTCCCGTGCAATCAGTTC | TCGGACGTACTTGAGGGAAT |
| Gusb | GUSB | NM_010368.1 | CTCTGGTGGCCTTACCTGAT | CAGTTGTTGTCACCTTCACCTC |
| Hprt ter | HPRT | NM_013556.2 | TGATAGATCCATTCCTATGACTGTAGA | AAGACATTCTTTCCAGTTAAAGTTGAG |
| Icam1 | ICAM | NM_010493.2 | GCTACCATCACCGTGTATTCG | AGGTCCTTGCCTACTTGCTG |
| Kcnma1 | Bkca alpha1 | NM_001253358.1 | GTACCTGTGGACCGTTTGCT | CGTCCACTGGCTTGAGAGTA |
| Klf2 | KLF | NM_008452.2 | CTAAAGGCGCATCTGCGTA | TAGTGGCGGGTAAGCTCGT |
| Nos3 | eNOS | NM_008713.4 | CCAGTGCCCTGCTTCATC | GCAGGGCAAGTTAGGATCAG |
| Nr2c3 | Nuclear receptor subfamily 3, group C, member 2 | NM_001083906.1 | TTCGGAGAAAGAACTGTCCTG | CCCAGCTTCTTTGACTTTCG |
| Opa1 | Opa1 | NM_001199177.1 | ACCAGGAGAAGTAGACTGTGTCAA | TCTTCAAATAAACGCAGAGGTG |
| P2rx4 | P2X4 | NM_011026.2 | CCAACACTTCTCAGCTTGGAT | TGGTCATGATGAAGAGGGAGT |
| P2ry6 | P2Y6 | NM_183168.1 | TCTTCCATCTTGCATGAGACA | GGATGGTGCCATTGTCCT |
| Pecam1 | CD31 | NM_001032378.1 | CGGTGTTCAGCGAGATCC | CGACAGGATGGAAATCACAA |
| Piezo1 | PIEZO1 | NM_001037298.1 | ATCAAGTGCAGCCGAGAGAC | TAATGAGGCCTCCCATACCA |
| Ptgis | PgI2 synthase, prostacyclin synthase | NM_008968.3 | AGGAAAAGCACGGTGACATATT | CCCACACCACTGTGTCGTAA |
| Ptgs1 | COX1 | NM_008969.3 | CCTCTTTCCAGGAGCTCACA | TCGATGTCACCGTACAGCTC |
| Ptgs2 | COX2 | NM_011198.3 | GGGAGTCTGGAACATTGTGAA | GCACATTGTAAGTAGGTGGACTGT |
| Ptpn1 | Protein tyrosine phosphatase, non-receptor type 1 | NM_011201.3 | CATCATGGAGAAAGGCTCGT | CCTGTGTCATCAAAGACCATCT |
| S1pr1 | Sphingosine-1-phosphate receptor 1 | NM_007901.5 | CGGTGTAGACCCAGAGTCCT | AGCTTTCCTTGGCTGGAG |
| Sdc4 | Syndecan4 | NM_011521.2 | GACCTCCTGGAAGGCAGATA | GCTCCTCCGTGTCATCCA |
| Shc1a | p66shc isoform a | NM_001113331 | GGACCCATTCTGCCTCCTCT | GCCAGCTTCAGGTTGCTCAT |
| Sirt1 | Sirtuin1 | NM_019812.2 | CAGTGAGAAAATGCTGGCCTA | TTGGTGGTACAAACAGGTATTGA |
| Sod1 | SOD1 | NM_011434.1 | CAGGACCTCATTTTAATCCTCAC | TGCCCAGGTCTCCAACAT |
| Sod2 | SOD2 | NM_013671.3 | GACCCATTGCAAGGAACAA | GTAGTAAGCGTGCTCCCACAC |
| Tfam | Tfam | NM_009360.4 | CAAAGGATGATTCGGCTCAG | AAGCTGAATATATGCCTGCTTTTC |
| Trpv4 | Trpv4 | NM_022017.3 | GGCAAGAGTGAAATCTACCAGTACTAT | ACCGAGGACCAACGATCC |
| Vcam1 | VCAM1 | NM_011693.2 | TGATTGGGAGAGACAAAGCA | AACAACCGAATCCCCAACTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Favre, J.; Vessieres, E.; Guihot, A.-L.; Grimaud, L.; Proux, C.; Loufrani, L.; Lenfant, F.; Fontaine, C.; Arnal, J.-F.; Henrion, D. Early Inactivation of Membrane Estrogen Receptor Alpha (ERα) Recapitulates the Endothelial Dysfunction of Aged Mouse Resistance Arteries. Int. J. Mol. Sci. 2022, 23, 2862. https://doi.org/10.3390/ijms23052862
Favre J, Vessieres E, Guihot A-L, Grimaud L, Proux C, Loufrani L, Lenfant F, Fontaine C, Arnal J-F, Henrion D. Early Inactivation of Membrane Estrogen Receptor Alpha (ERα) Recapitulates the Endothelial Dysfunction of Aged Mouse Resistance Arteries. International Journal of Molecular Sciences. 2022; 23(5):2862. https://doi.org/10.3390/ijms23052862
Chicago/Turabian StyleFavre, Julie, Emilie Vessieres, Anne-Laure Guihot, Linda Grimaud, Coralyne Proux, Laurent Loufrani, Françoise Lenfant, Coralie Fontaine, Jean-François Arnal, and Daniel Henrion. 2022. "Early Inactivation of Membrane Estrogen Receptor Alpha (ERα) Recapitulates the Endothelial Dysfunction of Aged Mouse Resistance Arteries" International Journal of Molecular Sciences 23, no. 5: 2862. https://doi.org/10.3390/ijms23052862