
Reduction in Phosphoribulokinase Amount and Re-Routing Metabolism in Chlamydomonas reinhardtii CP12 Mutants

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
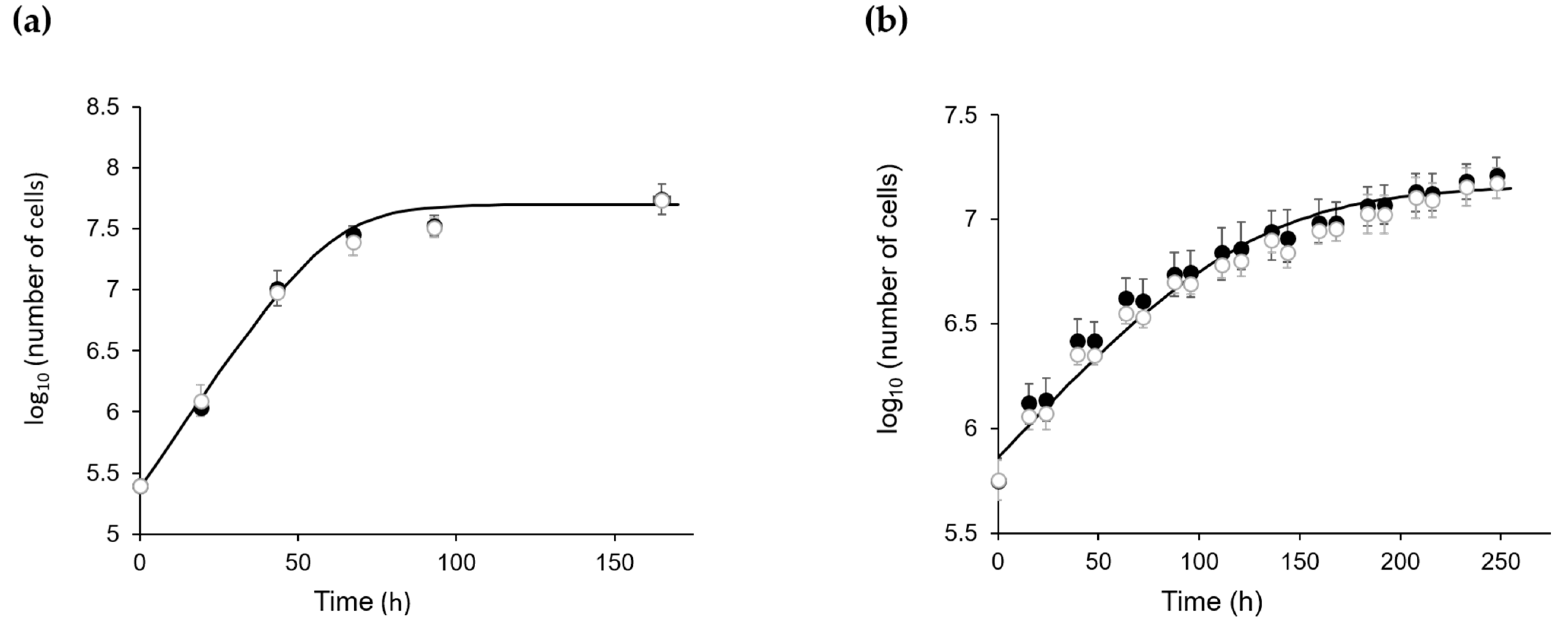
2.1. The Absence of CP12 Protein Has No Effect on C. reinhardtii Growth
2.2. Absence of CP12 Abolishes GAPDH Regulation
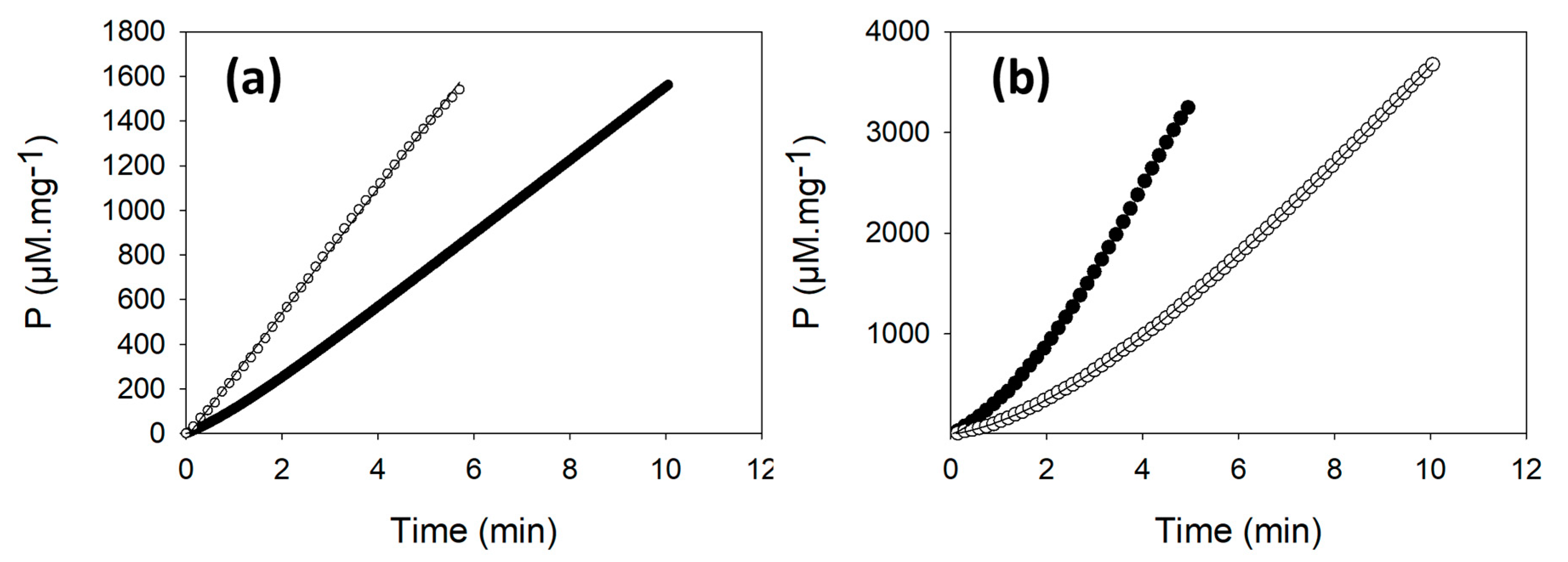
2.3. PRK Activity Is Lower in ΔCP12-Cr Compared to WT-Cr
2.4. Lower PRK Abundance in ΔCP12-Cr Compared to WT-Cr
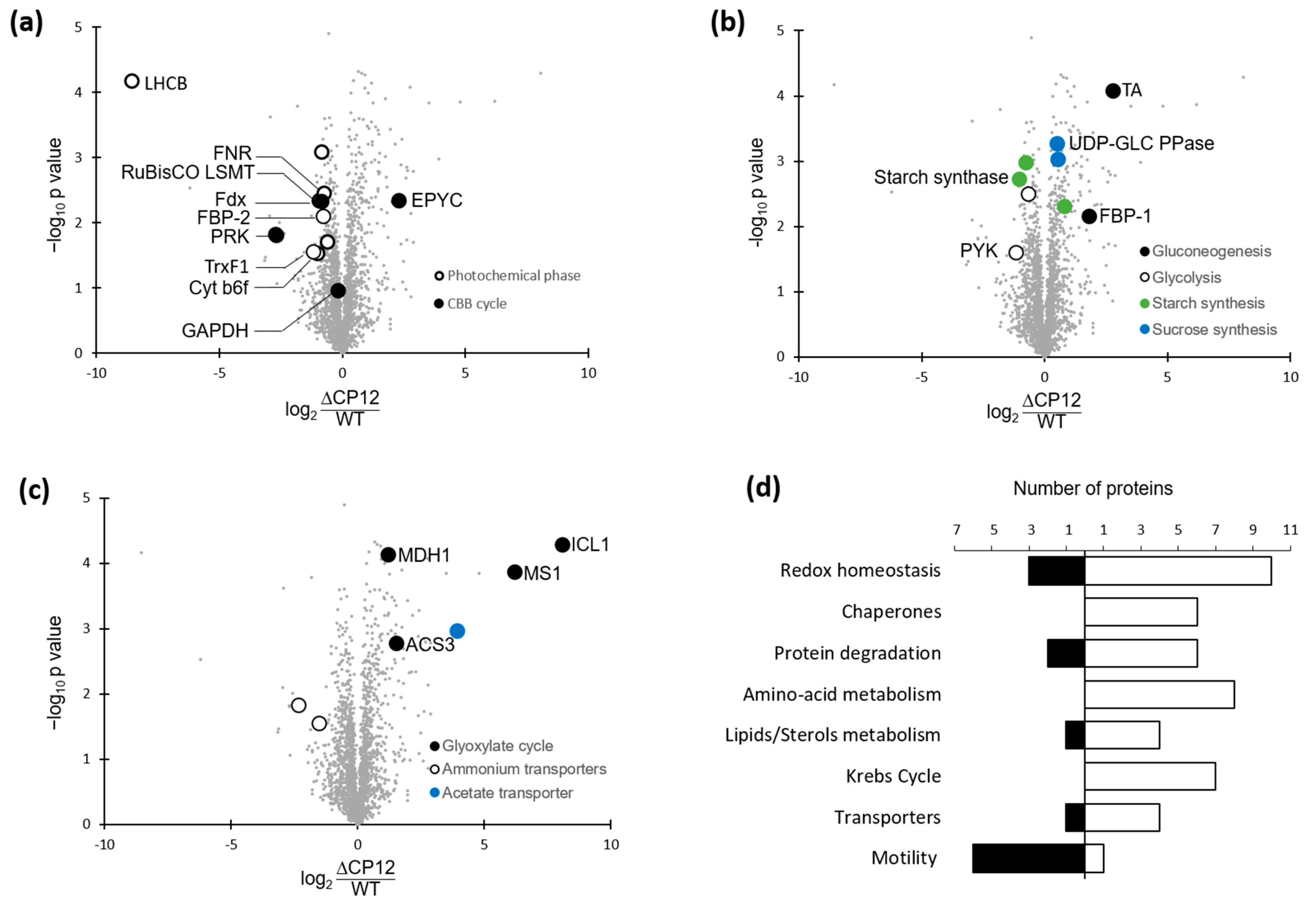
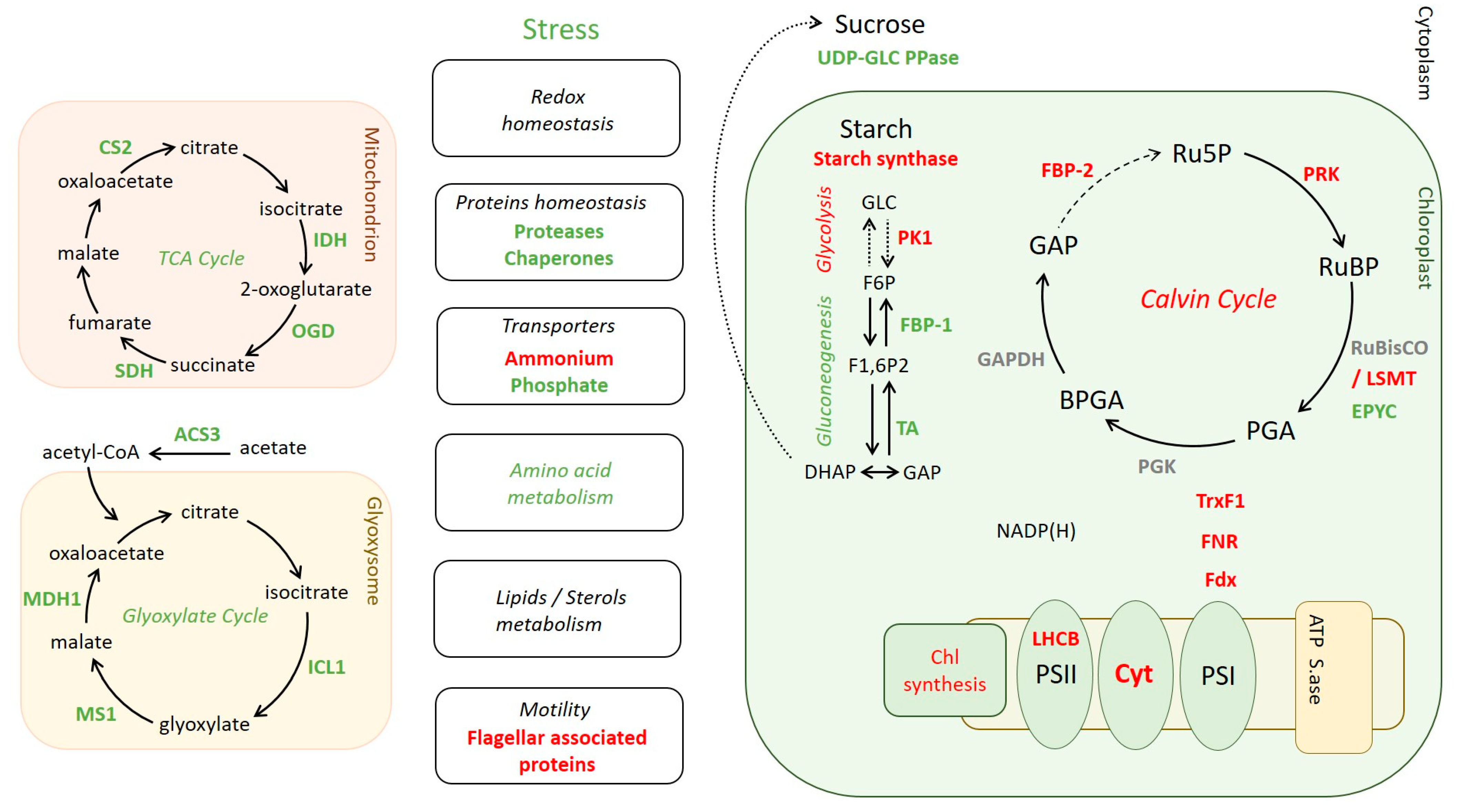
2.5. Differential Levels of Proteins Associated with Various Pathways in ΔCP12-Cr vs. WT-Cr
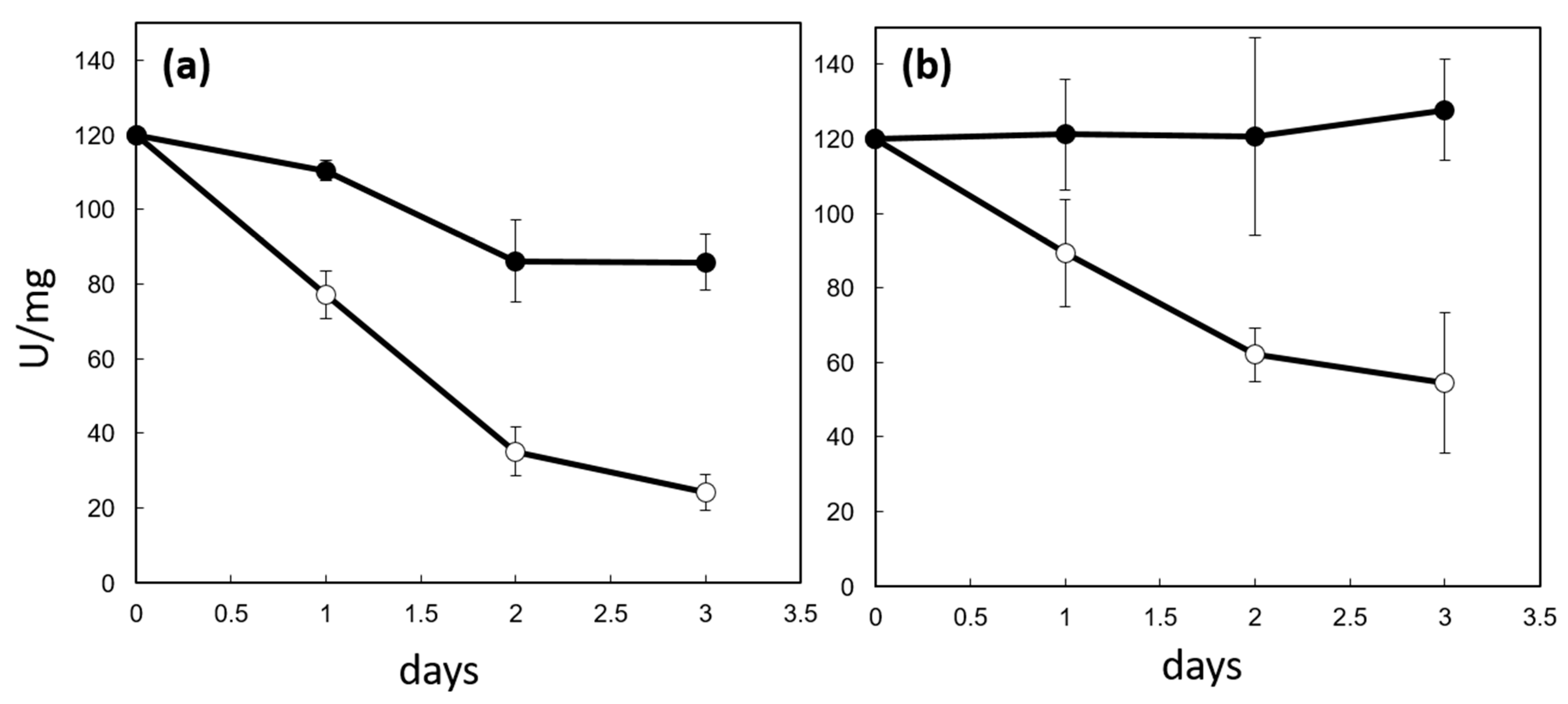
2.6. CP12 Protein Protects PRK from Irreversible Inactivation In Vitro
2.7. Identification of CP12 Residues Involved in PRK Protection
3. Discussion
4. Materials and Methods
4.1. Algal Strains and Culture Conditions
4.2. Growth Measurements
4.3. Sample Preparation for Relative Quantitative Proteomic Experiments
4.4. Relative Quantitative Proteomic Data Analysis
4.5. Activity Measurements
4.6. Progress Curve Analysis
4.7. Assay of PRK Protection against Irreversible Inhibition by CP12
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gurrieri, L.; Fermani, S.; Zaffagnini, M.; Sparla, F.; Trost, P. Calvin–Benson Cycle Regulation Is Getting Complex. Trends Plant Sci. 2021, 26, 898–912. [Google Scholar] [CrossRef] [PubMed]
- Cejudo, F.J.; Ojeda, V.; Delgado-Requerey, V.; González, M.; Pérez-Ruiz, J.M. Chloroplast Redox Regulatory Mechanisms in Plant Adaptation to Light and Darkness. Front. Plant Sci. 2019, 10, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, K.; Hisabori, T. Biochemical Basis for Redox Regulation of Chloroplast-Localized Phosphofructokinase from Arabidopsis thaliana. Plant Cell Physiol. 2021, 62, 401–410. [Google Scholar] [CrossRef]
- Fridlyand, L.E.; Scheibe, R. Regulation of the Calvin Cycle for CO2 Fixation as an Example for General Control Mechanisms in Metabolic Cycles. Biosystems 1999, 51, 79–93. [Google Scholar] [CrossRef]
- Gontero, B.; Salvucci, M.E. Regulation of Photosynthetic Carbon Metabolism in Aquatic and Terrestrial Organisms by RuBisCO Activase, Redox-Modulation and CP12. Aquat. Bot. 2014, 118, 14–23. [Google Scholar] [CrossRef]
- Graciet, E.; Lebreton, S.; Gontero, B. Emergence of New Regulatory Mechanisms in the Benson-Calvin Pathway via Protein-Protein Interactions: A Glyceraldehyde-3-Phosphate Dehydrogenase/CP12/Phosphoribulokinase Complex. J. Exp. Bot. 2004, 55, 1245–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebreton, S.; Graciet, E.; Gontero, B. Modulation, via Protein-Protein Interactions, of Glyceraldehyde-3-Phosphate Dehydrogenase Activity through Redox Phosphoribulokinase Regulation. J. Biol. Chem. 2003, 278, 12078–12084. [Google Scholar] [CrossRef] [Green Version]
- Wedel, N.; Soll, J. Evolutionary Conserved Light Regulation of Calvin Cycle Activity by NADPH-Mediated Reversible Phosphoribulokinase/CP12/ Glyceraldehyde-3-Phosphate Dehydrogenase Complex Dissociation. Proc. Natl. Acad. Sci. USA 1998, 95, 9699–9704. [Google Scholar] [CrossRef] [Green Version]
- Groben, R.; Kaloudas, D.; Raines, C.A.; Offmann, B.; Maberly, S.C.; Gontero, B. Comparative Sequence Analysis of CP12, a Small Protein Involved in the Formation of a Calvin Cycle Complex in Photosynthetic Organisms. Photosynth. Res. 2010, 103, 183–194. [Google Scholar] [CrossRef]
- Launay, H.; Barré, P.; Puppo, C.; Manneville, S.; Gontero, B.; Receveur-Bréchot, V. Absence of Residual Structure in the Intrinsically Disordered Regulatory Protein CP12 in Its Reduced State. BBRC 2016, 477, 20–26. [Google Scholar] [CrossRef]
- Launay, H.; Barré, P.; Puppo, C.; Zhang, Y.; Maneville, S.; Gontero, B.; Receveur-Bréchot, V. Cryptic Disorder out of Disorder: Encounter between Conditionally Disordered CP12 and Glyceraldehyde-3-Phosphate Dehydrogenase. J. Mol. Biol. 2018, 430, 1218–1234. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, D.; Jakob, U. The Roles of Conditional Disorder in Redox Proteins. Curr. Opin. Struct. Biol. 2013, 23, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gontero, B.; Maberly, S.C. An Intrinsically Disordered Protein, CP12: Jack of All Trades and Master of the Calvin Cycle. Biochem. Soc. Trans. 2012, 40, 995–999. [Google Scholar] [CrossRef] [Green Version]
- Tamoi, M.; Miyazaki, T.; Fukamizo, T.; Shigeoka, S. The Calvin Cycle in Cyanobacteria Is Regulated by CP12 via the NAD(H)/NADP(H) Ratio under Light/Dark Conditions. Plant J. 2005, 42, 504–513. [Google Scholar] [CrossRef]
- Erales, J.; Lignon, S.; Gontero, B. CP12 from Chlamydomonas reinhardtii, a Permanent Specific “Chaperone-like” Protein of Glyceraldehyde-3-Phosphate Dehydrogenase. J. Biol. Chem. 2009, 284, 12735–12744. [Google Scholar] [CrossRef] [Green Version]
- Delobel, A.; Graciet, E.; Andreescu, S.; Gontero, B.; Halgand, F.; Laprévote, O. Mass Spectrometric Analysis of the Interactions between CP12, a Chloroplast Protein, and Metal Ions: A Possible Regulatory Role within a PRK/GAPDH/CP12 Complex. Rapid Commun. Mass Spectrom. 2005, 19, 3379–3388. [Google Scholar] [CrossRef] [Green Version]
- Erales, J.; Gontero, B.; Whitelegge, J.; Halgand, F. Mapping of a Copper-Binding Site on the Small CP12 Chloroplastic Protein of Chlamydomonas reinhardtii Using Top-down Mass Spectrometry and Site-Directed Mutagenesis. Biochem. J. 2009, 419, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.G.; Vothknecht, U.C. Identification of CP12 as a Novel Calcium-Binding Protein in Chloroplasts. Plants 2013, 2, 530–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erales, J.; Avilan, L.; Lebreton, S.; Gontero, B. Exploring CP12 Binding Proteins Revealed Aldolase as a New Partner for the Phosphoribulokinase/Glyceraldehyde 3-Phosphate Dehydrogenase/CP12 Complex—Purification and Kinetic Characterization of This Enzyme from Chlamydomonas reinhardtii. FEBS J. 2008, 275, 1248–1259. [Google Scholar] [CrossRef]
- Howard, T.P.; Fryer, M.J.; Singh, P.; Metodiev, M.; Lytovchenko, A.; Obata, T.; Fernie, A.R.; Kruger, N.J.; Quick, W.P.; Lloyd, J.C.; et al. Antisense Suppression of the Small Chloroplast Protein CP12 in Tobacco Alters Carbon Partitioning and Severely Restricts Growth. Plant Physiol. 2011, 157, 620–631. [Google Scholar] [CrossRef] [Green Version]
- Howard, T.P.; Upton, G.J.G.; Lloyd, J.C.; Raines, C.A. Antisense Suppression of the Small Chloroplast Protein CP12 in Tobacco: A Transcriptional Viewpoint. Plant Signal Behav. 2011, 6, 2026–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Calcagno, P.E.; Abuzaid, O.; Lawson, T.; Raines, C.A. Arabidopsis CP12 Mutants Have Reduced Levels of Phosphoribulokinase and Impaired Function of the Calvin–Benson Cycle. J. Exp. Bot. 2017, 68, 2285–2298. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Qiu, H.; Zhou, M.; Lin, Y.; Guo, Z.; Lu, S. Chloroplast Protein 12 Expression Alters Growth and Chilling Tolerance in Tropical Forage Stylosanthes guianensis (Aublet) Sw. Front. Plant Sci. 2018, 9, 1319. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; van ’t Riet, K. Modeling of the Bacterial Growth Curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [Green Version]
- Plancke, C.; Vigeolas, H.; Höhner, R.; Roberty, S.; Emonds-Alt, B.; Larosa, V.; Willamme, R.; Duby, F.; Onga Dhali, D.; Thonart, P.; et al. Lack of Isocitrate Lyase in Chlamydomonas Leads to Changes in Carbon Metabolism and in the Response to Oxidative Stress under Mixotrophic Growth. Plant J. 2014, 77, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Lauersen, K.J.; Willamme, R.; Coosemans, N.; Joris, M.; Kruse, O.; Remacle, C. Peroxisomal Microbodies Are at the Crossroads of Acetate Assimilation in the Green Microalga Chlamydomonas reinhardtii. Algal. Res. 2016, 16, 266–274. [Google Scholar] [CrossRef]
- Marri, L.; Zaffagnini, M.; Collin, V.; Issakidis-Bourguet, E.; Lemaire, S.D.; Pupillo, P.; Sparla, F.; Miginiac-Maslow, M.; Trost, P. Prompt and Easy Activation by Specific Thioredoxins of Calvin Cycle Enzymes of Arabidopsis thaliana Associated in the GAPDH/CP12/PRK Supramolecular Complex. Mol. Plant 2009, 2, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Thieulin-Pardo, G.; Remy, T.; Lignon, S.; Lebrun, R.; Gontero, B. Phosphoribulokinase from Chlamydomonas reinhardtii: A Benson-Calvin Cycle Enzyme Enslaved to Its Cysteine Residues. Mol. Biosyst. 2015, 11, 1134–1145. [Google Scholar] [CrossRef]
- Yu, A.; Xie, Y.; Pan, X.; Zhang, H.; Cao, P.; Su, X.; Chang, W.; Li, M. Photosynthetic Phosphoribulokinase Structures: Enzymatic Mechanisms and the Redox Regulation of the Calvin-Benson-Bassham Cycle. Plant Cell 2020, 32, 1556–1573. [Google Scholar] [CrossRef] [Green Version]
- Gurrieri, L.; Del Giudice, A.; Demitri, N.; Falini, G.; Pavel, N.V.; Zaffagnini, M.; Polentarutti, M.; Crozet, P.; Marchand, C.H.; Henri, J.; et al. Arabidopsis and Chlamydomonas Phosphoribulokinase Crystal Structures Complete the Redox Structural Proteome of the Calvin-Benson Cycle. Proc. Natl. Acad. Sci. USA 2019, 116, 8048–8053. [Google Scholar] [CrossRef] [Green Version]
- Mcfarlane, C.; Shah, N.; Kabasakal, B.V.; Cotton, C.A.R.; Bubeck, D.; Murray, J.W. Structural Basis of Light-Induced Redox Regulation in the Calvin Cycle. Proc. Natl. Acad. Sci. USA 2019, 116, 20984–20990. [Google Scholar] [CrossRef] [Green Version]
- Gardebien, F.; Thangudu, R.R.; Gontero, B.; Offmann, B. Construction of a 3D Model of CP12, a Protein Linker. J. Mol. Graph. Model. 2006, 25, 186–195. [Google Scholar] [CrossRef]
- Howard, T.P.; Lloyd, J.C.; Raines, C.A. Inter-Species Variation in the Oligomeric States of the Higher Plant Calvin Cycle Enzymes Glyceraldehyde-3-Phosphate Dehydrogenase and Phosphoribulokinase. J. Exp. Bot. 2011, 62, 3799–3805. [Google Scholar] [CrossRef]
- Tamoi, M.; Shigeoka, S. CP12 Is Involved in Protection against High Light Intensity by Suppressing the ROS Generation in Synechococcus elongatus PCC7942. Plants 2021, 10, 1275. [Google Scholar] [CrossRef] [PubMed]
- Johnson, X.; Alric, J. Central Carbon Metabolism and Electron Transport in Chlamydomonas reinhardtii: Metabolic Constraints for Carbon Partitioning between Oil and Starch. Eukaryot. Cell 2013, 12, 776–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avilan, L.; Puppo, C.; Erales, J.; Woudstra, M.; Lebrun, R.; Gontero, B. CP12 Residues Involved in the Formation and Regulation of the Glyceraldehyde-3-Phosphate Dehydrogenase-CP12-Phosphoribulokinase Complex in Chlamydomonas reinhardtii. Mol. Biosyst. 2012, 8, 2994–3002. [Google Scholar] [CrossRef] [PubMed]
- Porter, M.A.; Stringer, C.D.; Hartman, F.C. Characterization of the Regulatory Thioredoxin Site of Phosphoribulokinase. J. Biol. Chem. 1988, 263, 123–129. [Google Scholar] [CrossRef]
- Avilan, L.; Lebreton, S.; Gontero, B. Thioredoxin Activation of Phosphoribulokinase in a Bi-Enzyme Complex from Chlamydomonas reinhardtii Chloroplasts. J. Biol. Chem. 2000, 275, 9447–9451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, M.J.; Knight, J.S.; Habash, D.; Parry, M.A.J.; Lawlor, D.W.; Barnes, S.A.; Loynes, A.; Gray, J.C. Reduction in Phosphoribulokinase Activity by Antisense RNA in Transgenic Tobacco: Effect on CO2 Assimilation and Growth in Low Irradiance. Plant J. 1995, 7, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Habash, D.Z.; Parry, M.A.; Parmar, S.; Paul, M.J.; Driscoll, S.; Knight, J.; Gray, J.C.; Lawlor, D.W. The Regulation of Component Processes of Photosynthesis in Transgenic Tobacco with Decreased Phosphoribulokinase Activity. Photosynth. Res. 1996, 49, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Oesterhelt, C.; Klocke, S.; Holtgrefe, S.; Linke, V.; Weber, A.P.M.; Scheibe, R. Redox Regulation of Chloroplast Enzymes in Galdieria Sulphuraria in View of Eukaryotic Evolution. Plant Cell Physiol. 2007, 48, 1359–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, M.; Sugase, K.; Dyson, H.J.; Wright, P.E. Conformational Propensities of Intrinsically Disordered Proteins Influence the Mechanism of Binding and Folding. Proc. Natl. Acad. Sci. USA 2015, 112, 9614–9619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habchi, J.; Tompa, P.; Longhi, S.; Uversky, V.N. Introducing Protein Intrinsic Disorder. Chem. Rev. 2014, 114, 6561–6588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, B.B. The Ferredoxin/Thioredoxin System: A Key Element in the Regulatory Function of Light in Photosynthesis. Bioscience 1984, 34, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Wunder, T.; Cheng, S.L.H.; Lai, S.-K.; Li, H.-Y.; Mueller-Cajar, O. The Phase Separation Underlying the Pyrenoid-Based Microalgal RuBisCO Supercharger. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mackinder, L.C.M.; Meyer, M.T.; Mettler-Altmann, T.; Chen, V.K.; Mitchell, M.C.; Caspari, O.; Freeman Rosenzweig, E.S.; Pallesen, L.; Reeves, G.; Itakura, A.; et al. A Repeat Protein Links RuBisCO to Form the Eukaryotic Carbon-Concentrating Organelle. Proc. Natl. Acad. Sci. USA 2016, 113, 5958–5963. [Google Scholar] [CrossRef] [Green Version]
- Santhanagopalan, I.; Wong, R.; Mathur, T.; Griffiths, H. Orchestral Manoeuvres in the Light: Crosstalk Needed for Regulation of the Chlamydomonas Carbon Concentration Mechanism. J. Exp. Bot. 2021, 72, 4604–4624. [Google Scholar] [CrossRef]
- Trievel, R.C.; Flynn, E.M.; Houtz, R.L.; Hurley, J.H. Mechanism of Multiple Lysine Methylation by the SET Domain Enzyme RuBisCO LSMT. Nat. Struct. Biol. 2003, 10, 545–552. [Google Scholar] [CrossRef]
- Mininno, M.; Brugière, S.; Pautre, V.; Gilgen, A.; Ma, S.; Ferro, M.; Tardif, M.; Alban, C.; Ravanel, S. Characterization of Chloroplastic Fructose 1,6-Bisphosphate Aldolases as Lysine-Methylated Proteins in Plants. J. Biol. Chem. 2012, 287, 21034–21044. [Google Scholar] [CrossRef] [Green Version]
- Küken, A.; Sommer, F.; Yaneva-Roder, L.; Mackinder, L.C.; Höhne, M.; Geimer, S.; Jonikas, M.C.; Schroda, M.; Stitt, M.; Nikoloski, Z.; et al. Effects of Microcompartmentation on Flux Distribution and Metabolic Pools in Chlamydomonas reinhardtii Chloroplasts. eLife 2018, 7, e37960. [Google Scholar] [CrossRef]
- Lejay, L.; Wirth, J.; Pervent, M.; Cross, J.M.-F.; Tillard, P.; Gojon, A. Oxidative Pentose Phosphate Pathway-Dependent Sugar Sensing as a Mechanism for Regulation of Root Ion Transporters by Photosynthesis. Plant Physiol. 2008, 146, 2036–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaput, V.; Martin, A.; Lejay, L. Redox Metabolism: The Hidden Player in Carbon and Nitrogen Signaling? J. Exp. Bot. 2020, 71, 3816–3826. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.S.; Kim, J.; Park, H.; Hong, S.-J.; Lee, C.-G.; Jin, E. Enhanced Lipid Productivity in AGP Knockout Marine Microalga Tetraselmis Sp. Using a DNA-Free CRISPR-Cas9 RNP Method. Bioresour. Technol. 2020, 303, 122932. [Google Scholar] [CrossRef]
- Song, I.; Kim, J.; Baek, K.; Choi, Y.; Shin, B.; Jin, E. The Generation of Metabolic Changes for the Production of High-Purity Zeaxanthin Mediated by CRISPR-Cas9 in Chlamydomonas reinhardtii. Microb. Cell Fact 2020, 19, 220. [Google Scholar] [CrossRef]
- Gorman, D.S.; Levine, R.P. Cytochrome f and Plastocyanin: Their Sequence in the Photosynthetic Electron Transport Chain of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1965, 54, 1665–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avilan, L.; Lebrun, R.; Puppo, C.; Citerne, S.; Cuiné, S.; Li-Beisson, Y.; Menand, B.; Field, B.; Gontero, B. ppGpp Influences Protein Protection, Growth and Photosynthesis in Phaeodactylum tricornutum. New Phytologist. 2021, 230, 1517–1532. [Google Scholar] [CrossRef]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A Peptide Search Engine Integrated into the MaxQuant Environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus Computational Platform for Comprehensive Analysis of (Prote)Omics Data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Graciet, E.; Gans, P.; Wedel, N.; Lebreton, S.; Camadro, J.-M.; Gontero, B. The Small Protein CP12: A Protein Linker for Supramolecular Complex Assembly. Biochemistry 2003, 42, 8163–8170. [Google Scholar] [CrossRef] [Green Version]
- Gontero, B.; Cárdenas, M.L.; Ricard, J. A Functional Five-Enzyme Complex of Chloroplasts Involved in the Calvin Cycle. Eur. J. Biochem. 1988, 173, 437–443. [Google Scholar] [CrossRef]
- Neet, K.E.; Ainslie, G.R. Hysteretic Enzymes. Methods Enzymol. 1980, 64, 192–226. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, S.; Gontero, B.; Avilan, L.; Ricard, J. Information Transfer in Multienzyme Complexes–1. Thermodynamics of Conformational Constraints and Memory Effects in the Bienzyme Glyceraldehyde-3-Phosphate-Dehydrogenase-Phosphoribulokinase Complex of Chlamydomonas reinhardtii Chloroplasts. Eur. J. Biochem. 1997, 250, 286–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NADPH-Dependent GAPDH Activity (µM.min−1) | PRK Activity (µM.min−1) | |||
|---|---|---|---|---|
| Reduced Cell Extract 1 | Untreated 2 | Reduced Cell Extract 1 | Untreated 2 | |
| WT-Cr | 212 ± 73 | 168 ± 47 | 1143 ± 328 | 940 ± 309 |
| ΔCP12-Cr | 332 ± 47 | 320 ± 61 | 430 ± 94 | 395 ± 99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gérard, C.; Lebrun, R.; Lemesle, E.; Avilan, L.; Chang, K.S.; Jin, E.; Carrière, F.; Gontero, B.; Launay, H. Reduction in Phosphoribulokinase Amount and Re-Routing Metabolism in Chlamydomonas reinhardtii CP12 Mutants. Int. J. Mol. Sci. 2022, 23, 2710. https://doi.org/10.3390/ijms23052710
Gérard C, Lebrun R, Lemesle E, Avilan L, Chang KS, Jin E, Carrière F, Gontero B, Launay H. Reduction in Phosphoribulokinase Amount and Re-Routing Metabolism in Chlamydomonas reinhardtii CP12 Mutants. International Journal of Molecular Sciences. 2022; 23(5):2710. https://doi.org/10.3390/ijms23052710
Chicago/Turabian StyleGérard, Cassy, Régine Lebrun, Erwan Lemesle, Luisana Avilan, Kwang Suk Chang, EonSeon Jin, Frédéric Carrière, Brigitte Gontero, and Hélène Launay. 2022. "Reduction in Phosphoribulokinase Amount and Re-Routing Metabolism in Chlamydomonas reinhardtii CP12 Mutants" International Journal of Molecular Sciences 23, no. 5: 2710. https://doi.org/10.3390/ijms23052710