
Nanoparticle Delivery Platforms for RNAi Therapeutics Targeting COVID-19 Disease in the Respiratory Tract

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. siRNA Therapy—A Potential and Promising Antiviral Therapeutic
2.1. Mechanisms of siRNA Therapy
2.2. siRNA Therapeutics for SARS-CoV-2, SARS-CoV-1, and MERS-CoV
3. NP Delivery—A Platform Ensuring the Efficacy and Stability of siRNAs
3.1. Lipid NPs (LNPs) in siRNA Therapeutics
3.2. Polymer NPs (PNPs) in siRNA Therapeutics
3.3. Glycogen NPs (GNPs) in siRNA Therapeutics
3.4. Other NP Platforms in siRNA Therapeutics
4. Challenges of Local Delivery of NP-siRNAs—The Importance of Administration Route
5. Methods to Perform Respiratory Delivery of NP-siRNAs
6. Methods to Assess Lung Delivery
7. Animal Models to Assess Pulmonary Delivery
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Counoupas, C.; Johansen, M.D.; Stella, A.O.; Nguyen, D.H.; Ferguson, A.L.; Aggarwal, A.; Bhattacharyya, N.D.; Grey, A.; Hutchings, O.; Patel, K.; et al. A single dose, BCG-adjuvanted COVID-19 vaccine provides sterilising immunity against SARS-CoV-2 infection. NPJ Vaccines 2021, 6, 143. [Google Scholar] [CrossRef] [PubMed]
- Seow, J.; Graham, C.; Merrick, B.; Acors, S.; Pickering, S.; Steel, K.J.A.; Hemmings, O.; O’Byrne, A.; Kouphou, N.; Galao, R.P.; et al. Longitudinal observation and decline of neutralizing antibody responses in the three months following SARS-CoV-2 infection in humans. Nat. Microbiol. 2020, 5, 1598–1607. [Google Scholar] [CrossRef] [PubMed]
- Anna, F.; Goyard, S.; Lalanne, A.I.; Nevo, F.; Gransagne, M.; Souque, P.; Louis, D.; Gillon, V.; Turbiez, I.; Bidard, F.C.; et al. High seroprevalence but short-lived immune response to SARS-CoV-2 infection in Paris. Eur. J. Immunol. 2021, 51, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Naaber, P.; Tserel, L.; Kangro, K.; Sepp, E.; Jurjenson, V.; Adamson, A.; Haljasmagi, L.; Rumm, A.P.; Maruste, R.; Karner, J.; et al. Dynamics of antibody response to BNT162b2 vaccine after six months: A longitudinal prospective study. Lancet Reg. Health Eur. 2021, 10, 100208. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.S.; Read, S.W.; Fauci, A.S. Therapy for Early COVID-19: A Critical Need. JAMA 2020, 324, 2149–2150. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. COVID-19: Pfizer’s paxlovid is 89% effective in patients at risk of serious illness, company reports. BMJ 2021, 375, n2713. [Google Scholar] [CrossRef]
- Fischer, W.; Eron, J.J.; Holman, W.; Cohen, M.S.; Fang, L.; Szewczyk, L.J.; Sheahan, T.P.; Baric, R.; Mollan, K.R.; Wolfe, C.R.; et al. Molnupiravir, an Oral Antiviral Treatment for COVID-19. medRxiv 2021. [Google Scholar] [CrossRef]
- Mahase, E. COVID-19: UK becomes first country to authorise antiviral molnupiravir. BMJ 2021, 375, n2697. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, L. In the age of Omicron variant: Paxlovid raises new hopes of COVID-19 recovery. J. Med. Virol. 2021, 27, 540. [Google Scholar] [CrossRef]
- Zhao, H.; Lu, L.; Peng, Z.; Chen, L.L.; Meng, X.; Zhang, C.; Ip, J.D.; Chan, W.M.; Chu, A.W.; Chan, K.H.; et al. SARS-CoV-2 Omicron variant shows less efficient replication and fusion activity when compared with delta variant in TMPRSS2-expressed cells. Emerg. Microbes Infect. 2021, 15, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Ferreira, I.A.T.M.; Abdullahi, A.; Goonawardane, N.; Saito, A.; Kimura, I.; Yamasoba, D.; Gerba, P.P.; Fatihi, S.; Rathore, S.; et al. SARS-CoV-2 Omicron spike mediated immune escape and tropism shift. bioRxiv 2022, 17, 473248. [Google Scholar]
- Huntington, K.E.; Carlsen, L.; So, E.Y.; Piesche, M.; Liang, O.; El-Deiry, W.S. Integrin/TGF-beta1 inhibitor GLPG-0187 blocks SARS-CoV-2 Delta and Omicron pseudovirus infection of airway epithelial cells which could attenuate disease severity. medRxiv 2022. [Google Scholar] [CrossRef]
- Diamond, M.; Halfmann, P.; Maemura, T.; Iwatsuki-Horimoto, K.; Iida, S.; Kiso, M.; Scheaffer, S.; Darling, T.; Joshi, A.; Loeber, S.; et al. The SARS-CoV-2 B.1.1.529 Omicron virus causes attenuated infection and disease in mice and hamsters. Res. Sq. 2021, 21, 1792. [Google Scholar] [CrossRef]
- Kozlov, M. Omicron’s feeble attack on the lungs could make it less dangerous. Nature 2022, 601, 177. [Google Scholar] [CrossRef] [PubMed]
- Bentley, E.G.; Kirby, A.; Sharma, P.; Kipar, A.; Mega, D.F.; Bramwell, C.; Penrice-Randal, R.; Prince, T.; Brown, J.C.; Zhou, J.; et al. SARS-CoV-2 Omicron-B.1.1.529 Variant leads to less severe disease than Pango B and Delta variants strains in a mouse model of severe COVID-19. bioRxiv 2021. [Google Scholar] [CrossRef]
- McMahan, K.; Giffin, V.; Tostanoski, L.H.; Chung, B.; Siamatu, M.; Suthar, M.S.; Halfmann, P.; Kawaoka, Y.; Piedra-Mora, C.; Martinot, A.J.; et al. Reduced Pathogenicity of the SARS-CoV-2 Omicron Variant in Hamsters. bioRxiv 2022. [Google Scholar] [CrossRef]
- Dowarah, J.; Marak, B.N.; Yadav, U.C.S.; Singh, V.P. Potential drug development and therapeutic approaches for clinical intervention in COVID-19. Bioorg. Chem. 2021, 114, 105016. [Google Scholar] [CrossRef]
- Zhao, B.; Yang, T.F.; Zheng, R. Theory and reality of antivirals against SARS-CoV-2. World J. Clin. Cases 2021, 9, 6663–6673. [Google Scholar] [CrossRef]
- Burnett, D.L.; Jackson, K.J.L.; Langley, D.B.; Aggrawal, A.; Stella, A.O.; Johansen, M.D.; Balachandran, H.; Lenthall, H.; Rouet, R.; Walker, G.; et al. Immunizations with diverse sarbecovirus receptor-binding domains elicit SARS-CoV-2 neutralizing antibodies against a conserved site of vulnerability. Immunity 2021, 54, 2908–2921.e6. [Google Scholar] [CrossRef]
- Torrente-Lopez, A.; Hermosilla, J.; Navas, N.; Cuadros-Rodriguez, L.; Cabeza, J.; Salmeron-Garcia, A. The Relevance of Monoclonal Antibodies in the Treatment of COVID-19. Vaccines 2021, 9, 557. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Siomi, H.; Siomi, M.C. On the road to reading the RNA-interference code. Nature 2009, 457, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Uludag, H.; Parent, K.; Aliabadi, H.M.; Haddadi, A. Prospects for RNAi Therapy of COVID-19. Front. Bioeng. Biotechnol. 2020, 8, 916. [Google Scholar] [CrossRef] [PubMed]
- Berber, B.; Aydin, C.; Kocabas, F.; Guney-Esken, G.; Yilancioglu, K.; Karadag-Alpaslan, M.; Caliseki, M.; Yuce, M.; Demir, S.; Tastan, C. Gene editing and RNAi approaches for COVID-19 diagnostics and therapeutics. Gene Ther. 2021, 28, 290–305. [Google Scholar] [CrossRef] [PubMed]
- Tay, H.L.; Kaiko, G.E.; Plank, M.; Li, J.; Maltby, S.; Essilfie, A.T.; Jarnicki, A.; Yang, M.; Mattes, J.; Hansbro, P.M.; et al. Antagonism of miR-328 increases the antimicrobial function of macrophages and neutrophils and rapid clearance of non-typeable Haemophilus influenzae (NTHi) from infected lung. PLoS Pathog. 2015, 11, e1004549. [Google Scholar]
- Foster, P.S.; Plank, M.; Collison, A.; Tay, H.L.; Kaiko, G.E.; Li, J.; Johnston, S.L.; Hansbro, P.M.; Kumar, R.K.; Yang, M.; et al. The emerging role of microRNAs in regulating immune and inflammatory responses in the lung. Immunol. Rev. 2013, 253, 198–215. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, C.C.; Choy, K.W.; Du, Q.; Chen, J.; Wang, Q.; Li, L.; Chung, T.K.; Tang, T. Therapeutic potentials of gene silencing by RNA interference: Principles, challenges, and new strategies. Gene 2014, 538, 217–227. [Google Scholar] [CrossRef]
- Hasan, M.; Ashik, A.I.; Chowdhury, M.B.; Tasnim, A.T.; Nishat, Z.S.; Hossain, T.; Ahmed, S. Computational prediction of potential siRNA and human miRNA sequences to silence orf1ab associated genes for future therapeutics against SARS-CoV-2. Inform. Med. Unlocked 2021, 24, 100569. [Google Scholar] [CrossRef]
- Idris, A.; Davis, A.; Supramaniam, A.; Acharya, D.; Kelly, G.; Tayyar, Y.; West, N.; Zhang, P.; McMillan, C.L.D.; Soemardy, C.; et al. A SARS-CoV-2 targeted siRNA-nanoparticle therapy for COVID-19. Mol. Ther. 2021, 29, 2219–2226. [Google Scholar] [CrossRef]
- McCaskill, J.; Singhania, R.; Burgess, M.; Allavena, R.; Wu, S.; Blumenthal, A.; McMillan, N.A. Efficient Biodistribution and Gene Silencing in the Lung epithelium via Intravenous Liposomal Delivery of siRNA. Mol. Ther. Nucleic Acids 2013, 2, e96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanotto, D.; Rossi, J.J. The promises and pitfalls of RNA-interference-based therapeutics. Nature 2009, 457, 426–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalhori, M.R.; Saadatpour, F.; Arefian, E.; Soleimani, M.; Farzaei, M.H.; Aneva, I.Y.; Echeverria, J. The Potential Therapeutic Effect of RNA Interference and Natural Products on COVID-19: A Review of the Coronaviruses Infection. Front. Pharmacol. 2021, 12, 616993. [Google Scholar] [CrossRef] [PubMed]
- Tolksdorf, B.; Nie, C.; Niemeyer, D.; Rohrs, V.; Berg, J.; Lauster, D.; Adler, J.M.; Haag, R.; Trimpert, J.; Kaufer, B.; et al. Inhibition of SARS-CoV-2 Replication by a Small Interfering RNA Targeting the Leader Sequence. Viruses 2021, 13, 2030. [Google Scholar] [CrossRef]
- Ghosh, S.; Firdous, S.M.; Nath, A. siRNA could be a potential therapy for COVID-19. EXCLI J. 2020, 19, 528–531. [Google Scholar]
- Tai, W. Current Aspects of siRNA Bioconjugate for In Vitro and In Vivo Delivery. Molecules 2019, 24, 2211. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Reske, S.N. Adenovirus-delivered siRNA. Methods Mol. Biol. 2004, 252, 523–532. [Google Scholar]
- Lundstrom, K. Viral Vectors in Gene Therapy. Diseases 2018, 6, 42. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Sonwane, A.A.; Singh, N.K.; Meshram, C.D.; Dahiya, S.S.; Pawar, S.S.; Gupta, S.P.; Chaturvedi, V.K.; Saini, M. Intracerebral delivery of small interfering RNAs (siRNAs) using adenoviral vector protects mice against lethal peripheral rabies challenge. Virus Res. 2012, 163, 11–18. [Google Scholar] [CrossRef]
- Ruigrok, M.J.R.; Frijlink, H.W.; Hinrichs, W.L.J. Pulmonary administration of small interfering RNA: The route to go? J. Control. Release 2016, 235, 14–23. [Google Scholar] [CrossRef]
- Yi, Y.; Noh, M.J.; Lee, K.H. Current advances in retroviral gene therapy. Curr. Gene Ther. 2011, 11, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Canver, M.C. Evaluation of the Clinical Success of Ex Vivo and In Vivo Gene Therapy. J. Young Investig. 2009, 7, 19. [Google Scholar]
- Nayerossadat, N.; Maedeh, T.; Ali, P.A. Viral and nonviral delivery systems for gene delivery. Adv. Biomed. Res. 2012, 1, 27. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.G.; Trobridge, G.D.; Petek, L.M.; Jacobs, M.A.; Kaul, R.; Russell, D.W. Large-scale analysis of adeno-associated virus vector integration sites in normal human cells. J. Virol. 2005, 79, 11434–11442. [Google Scholar] [CrossRef] [Green Version]
- Karda, R.; Counsell, J.R.; Karbowniczek, K.; Caproni, L.J.; Tite, J.P.; Waddington, S.N. Production of lentiviral vectors using novel, enzymatically produced, linear DNA. Gene Ther. 2019, 26, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.; Mehta, M.; Paudel, K.R.; Madheswaran, T.; Panneerselvam, J.; Gupta, G.; Su, Q.P.; Hansbro, P.M.; MacLoughlin, R.; Dua, K.; et al. Versatility of liquid crystalline nanoparticles in inflammatory lung diseases. Nanomedicine 2021, 16, 1545–1548. [Google Scholar] [CrossRef]
- Chan, Y.; Ng, S.W.; Chellappan, D.K.; Madheswaran, T.; Zeeshan, F.; Kumar, P.; Pillay, V.; Gupta, G.; Wadhwa, R.; Mehta, M.; et al. Celastrol-loaded liquid crystalline nanoparticles as an anti-inflammatory intervention for the treatment of asthma. Int. J. Polym. Mater. Polym. Biomater. 2021, 70, 754–763. [Google Scholar] [CrossRef]
- Khatak, S.; Mehta, M.; Awasthi, R.; Paudel, K.R.; Singh, S.K.; Gulati, M.; Hansbro, N.G.; Hansbro, P.M.; Dua, K.; Dureja, H. Solid lipid nanoparticles containing anti-tubercular drugs attenuate the Mycobacterium marinum infection. Tuberculosis 2020, 125, 102008. [Google Scholar] [CrossRef]
- Solanki, N.; Mehta, M.; Chellappan, D.K.; Gupta, G.; Hansbro, N.G.; Tambuwala, M.M.; Aa Aljabali, A.; Paudel, K.R.; Liu, G.; Satija, S.; et al. Antiproliferative effects of boswellic acid-loaded chitosan nanoparticles on human lung cancer cell line A549. Future Med. Chem. 2020, 12, 2019–2034. [Google Scholar] [CrossRef]
- Mehta, M.; Paudel, K.R.; Shukla, S.D.; Shastri, M.D.; Satija, S.; Singh, S.K.; Gulati, M.; Dureja, H.; Zacconi, F.C.; Hansbro, P.M.; et al. Rutin-loaded liquid crystalline nanoparticles attenuate oxidative stress in bronchial epithelial cells: A PCR validation. Future Med. Chem. 2021, 13, 543–549. [Google Scholar] [CrossRef]
- Aigner, A.; Kogel, D. Nanoparticle/siRNA-based therapy strategies in glioma: Which nanoparticles, which siRNAs? Nanomedicine 2018, 13, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Yetisgin, A.A.; Cetinel, S.; Zuvin, M.; Kosar, A.; Kutlu, O. Therapeutic Nanoparticles and Their Targeted Delivery Applications. Molecules 2020, 25, 2193. [Google Scholar] [CrossRef]
- Yao, Y.; Zhou, Y.; Liu, L.; Xu, Y.; Chen, Q.; Wang, Y.; Wu, S.; Deng, Y.; Zhang, J.; Shao, A. Nanoparticle-Based Drug Delivery in Cancer Therapy and Its Role in Overcoming Drug Resistance. Front. Mol. Biosci. 2020, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; Rodriguez-Torres, M.D.P.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the clinic: An update. Bioeng. Transl. Med. 2019, 4, e10143. [Google Scholar] [CrossRef] [Green Version]
- Thi, T.T.H.; Suys, E.J.A.; Lee, J.S.; Nguyen, D.H.; Park, K.D.; Truong, N.P. Lipid-Based Nanoparticles in the Clinic and Clinical Trials: From Cancer Nanomedicine to COVID-19 Vaccines. Vaccines 2021, 9, 359. [Google Scholar] [CrossRef]
- Hoy, S.M. Patisiran: First Global Approval. Drugs 2018, 78, 1625–1631. [Google Scholar] [CrossRef]
- Synowiec, A.; Szczepanski, A.; Barreto-Duran, E.; Lie, L.K.; Pyrc, K. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): A Systemic Infection. Clin. Microbiol. Rev. 2021, 34, e00133-20. [Google Scholar] [CrossRef]
- Conickx, G.; Mestdagh, P.; Avila Cobos, F.; Verhamme, F.M.; Maes, T.; Vanaudenaerde, B.M.; Seys, L.J.; Lahousse, L.; Kim, R.Y.; Hsu, A.C.; et al. MicroRNA Profiling Reveals a Role for MicroRNA-218-5p in the Pathogenesis of Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2017, 195, 43–56. [Google Scholar] [CrossRef]
- Awasthi, R.; Roseblade, A.; Hansbro, P.M.; Rathbone, M.J.; Dua, K.; Bebawy, M. Nanoparticles in Cancer Treatment: Opportunities and Obstacles. Curr. Drug Targets 2018, 19, 1696–1709. [Google Scholar] [CrossRef]
- Prelli Bozzo, C.; Nchioua, R.; Volcic, M.; Koepke, L.; Kruger, J.; Schutz, D.; Heller, S.; Sturzel, C.M.; Kmiec, D.; Conzelmann, C.; et al. IFITM proteins promote SARS-CoV-2 infection and are targets for virus inhibition in vitro. Nat. Commun. 2021, 12, 4584. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J. Givosiran: First Approval. Drugs 2020, 80, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J.; Keam, S.J. Lumasiran: First Approval. Drugs 2021, 81, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Lamb, Y.N. Inclisiran: First Approval. Drugs 2021, 81, 389–395. [Google Scholar] [CrossRef]
- Mahmoodi Chalbatani, G.; Dana, H.; Gharagouzloo, E.; Grijalvo, S.; Eritja, R.; Logsdon, C.D.; Memari, F.; Miri, S.R.; Rad, M.R.; Marmari, V. Small interfering RNAs (siRNAs) in cancer therapy: A nano-based approach. Int. J. Nanomed. 2019, 14, 3111–3128. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.Q.; Lu, S.; Zhang, L.X.; Ji, M.; Liu, S.Y.; Wang, S.W.; Liu, R.T. An indoleamine 2, 3-dioxygenase siRNA nanoparticle-coated and Trp2-displayed recombinant yeast vaccine inhibits melanoma tumor growth in mice. J. Control. Release 2018, 273, 1–12. [Google Scholar] [CrossRef]
- Tabernero, J.; Shapiro, G.I.; LoRusso, P.M.; Cervantes, A.; Schwartz, G.K.; Weiss, G.J.; Paz-Ares, L.; Cho, D.C.; Infante, J.R.; Alsina, M.; et al. First-in-humans trial of an RNA interference therapeutic targeting VEGF and KSP in cancer patients with liver involvement. Cancer Discov. 2013, 3, 406–417. [Google Scholar] [CrossRef] [Green Version]
- Wagner, M.J.; Mitra, R.; McArthur, M.J.; Baze, W.; Barnhart, K.; Wu, S.Y.; Rodriguez-Aguayo, C.; Zhang, X.; Coleman, R.L.; Lopez-Berestein, G.; et al. Preclinical Mammalian Safety Studies of EPHARNA (DOPC Nanoliposomal EphA2-Targeted siRNA). Mol. Cancer Ther. 2017, 16, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Tolcher, A.W.; Papadopoulos, K.P.; Patnaik, A.; Rasco, D.W.; Martinez, D.; Wood, D.L.; Fielman, B.; Sharma, M.; Janisch, L.A.; Brown, B.D. Safety and Activity of DCR-MYC, a First-in-Class Dicer-Substrate Small Interfering RNA (DsiRNA) Targeting MYC, in a Phase I Study in Patients with Advanced Solid Tumors; American Society of Clinical Oncology: Alexandria, VA, USA, 2015. [Google Scholar]
- Schultheis, B.; Strumberg, D.; Santel, A.; Vank, C.; Gebhardt, F.; Keil, O.; Lange, C.; Giese, K.; Kaufmann, J.; Khan, M.; et al. First-in-human phase I study of the liposomal RNA interference therapeutic Atu027 in patients with advanced solid tumors. J. Clin. Oncol. 2014, 32, 4141–4148. [Google Scholar] [CrossRef]
- Zuckerman, J.E.; Gritli, I.; Tolcher, A.; Heidel, J.D.; Lim, D.; Morgan, R.; Chmielowski, B.; Ribas, A.; Davis, M.E.; Yen, Y. Correlating animal and human phase Ia/Ib clinical data with CALAA-01, a targeted, polymer-based nanoparticle containing siRNA. Proc. Natl. Acad. Sci. USA 2014, 111, 11449–11454. [Google Scholar] [CrossRef] [Green Version]
- Varghese, A.M.; Ang, C.; Dimaio, C.J.; Javle, M.M.; Gutierrez, M.; Yarom, N.; Stemmer, S.M.; Golan, T.; Geva, R.; Semenisty, V. A Phase II Study of siG12D-LODER in Combination with Chemotherapy in Patients with Locally Advanced Pancreatic Cancer (PROTACT); American Society of Clinical Oncology: Alexandria, VA, USA, 2020. [Google Scholar]
- Sajid, M.I.; Moazzam, M.; Cho, Y.; Kato, S.; Xu, A.; Way, J.J.; Lohan, S.; Tiwari, R.K. siRNA Therapeutics for the Therapy of COVID-19 and Other Coronaviruses. Mol. Pharm. 2021, 18, 2105–2121. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, Y.; Fu, L.; Yu, C.; Li, X.; Li, Y.; Zhang, X.; Rong, Z.; Wang, Y.; Ning, H.; et al. siRNA targeting the leader sequence of SARS-CoV inhibits virus replication. Gene Ther. 2005, 12, 751–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbashir, S.M.; Harborth, J.; Lendeckel, W.; Yalcin, A.; Weber, K.; Tuschl, T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001, 411, 494–498. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Doudna, J.A. A three-dimensional view of the molecular machinery of RNA interference. Nature 2009, 457, 405–412. [Google Scholar] [CrossRef]
- Klemm, V.; Mitchell, J.; Cortez-Jugo, C.; Cavalieri, F.; Symonds, G.; Caruso, F.; Kelleher, A.D.; Ahlenstiel, C. Achieving HIV-1 Control through RNA-Directed Gene Regulation. Genes 2016, 7, 119. [Google Scholar] [CrossRef] [Green Version]
- Morris, K.V.; Chan, S.W.; Jacobsen, S.E.; Looney, D.J. Small interfering RNA-induced transcriptional gene silencing in human cells. Science 2004, 305, 1289–1292. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, H.; Taira, K.; Morris, K.V. siRNA induced transcriptional gene silencing in mammalian cells. Cell Cycle 2005, 4, 442–448. [Google Scholar] [CrossRef]
- Callinan, P.A.; Feinberg, A.P. The emerging science of epigenomics. Hum. Mol. Genet. 2006, 15, R95–R101. [Google Scholar] [CrossRef]
- Corman, V.M.; Lienau, J.; Witzenrath, M. [Coronaviruses as the cause of respiratory infections]. Internist 2019, 60, 1136–1145. [Google Scholar] [CrossRef] [Green Version]
- Araf, Y.; Faruqui, N.A.; Anwar, S.; Hosen, M.J. SARS-CoV-2: A new dimension to our understanding of coronaviruses. Int. Microbiol. 2021, 24, 19–24. [Google Scholar] [CrossRef]
- Forgie, S.; Marrie, T.J. Healthcare-associated atypical pneumonia. Semin. Respir. Crit. Care Med. 2009, 30, 67–85. [Google Scholar] [CrossRef]
- Menachery, V.D.; Mitchell, H.D.; Cockrell, A.S.; Gralinski, L.E.; Yount, B.L., Jr.; Graham, R.L.; McAnarney, E.T.; Douglas, M.G.; Scobey, T.; Beall, A.; et al. MERS-CoV Accessory ORFs Play Key Role for Infection and Pathogenesis. mBio 2017, 8, e00665-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhong, L. Genomics functional analysis and drug screening of SARS-CoV-2. Genes Dis. 2020, 7, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar] [PubMed] [Green Version]
- Zhu, Z.; Lian, X.; Su, X.; Wu, W.; Marraro, G.A.; Zeng, Y. From SARS and MERS to COVID-19: A brief summary and comparison of severe acute respiratory infections caused by three highly pathogenic human coronaviruses. Respir. Res. 2020, 21, 224. [Google Scholar] [CrossRef]
- Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; Peters, B.; Sette, A. A Sequence Homology and Bioinformatic Approach Can Predict Candidate Targets for Immune Responses to SARS-CoV-2. Cell Host Microbe 2020, 27, 671–680.e2. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Kandeel, M.; Ibrahim, A.; Fayez, M.; Al-Nazawi, M. From SARS and MERS CoVs to SARS-CoV-2: Moving toward more biased codon usage in viral structural and nonstructural genes. J. Med. Virol. 2020, 92, 660–666. [Google Scholar] [CrossRef]
- Niktab, I.; Haghparast, M.; Beigi, M.H.; Megraw, T.L.; Kiani, A.; Ghaedi, K. Design of advanced siRNA therapeutics for the treatment of COVID-19. Meta Gene 2021, 29, 100910. [Google Scholar] [CrossRef]
- Bestle, D.; Heindl, M.R.; Limburg, H.; Van Lam van, T.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; et al. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 3, 786. [Google Scholar] [CrossRef]
- Aljaberi, A.; Migdadi, E.M.; Khadra, K.M.A.; Samak, M.A.; Basheti, I.A.; Al-Zoubi, N. siRNA as a potential therapy for COVID-19. Curr. Drug Deliv. 2021, 19, 528. [Google Scholar] [CrossRef] [PubMed]
- Fakhr, E.; Zare, F.; Teimoori-Toolabi, L. Precise and efficient siRNA design: A key point in competent gene silencing. Cancer Gene Ther. 2016, 23, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Bing, S.U.N.; Bojian, Z.; Wei, L.U.; Ke, X.U. SARS Coronavirus Disturbance RNA and Its Uses. Patent No. CN101085986, 8 June 2006. [Google Scholar]
- Hou, W.; Shen, L.I.; Lu, A. Medicine for Preventing and Treatig SARS Coronavirus. Patent No. CN1548054, 5 May 2003. [Google Scholar]
- Ying, W.; Li, L.I.U.; Shuhui, W.; Yun, Z. Small Interfering RNA for Restraining SARS Corona Virus M Protein Gene Expression, Encoding Gene and Application Thereof. Patent No. CN20061114168, 31 October 2006. [Google Scholar]
- Yizheng, Z.; Gang, W.; Min, L.I.; Haiyan, W.; Hong, F. siRNA Disturbing RdRp Gene Function of SARS Virus. Patent CN101113158, 18 December 2006. [Google Scholar]
- Nur, S.M.; Hasan, M.A.; Amin, M.A.; Hossain, M.; Sharmin, T. Design of Potential RNAi (miRNA and siRNA) Molecules for Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Gene Silencing by Computational Method. Interdiscip. Sci. 2015, 7, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S.; Berindan-Neagoe, I.; Cismaru, C.A.; Schaafsma, D.; Nabavi, S.F.; Ghavami, S.; Banach, M.; Nabavi, S.M. Lessons from SARS and MERS remind us of the possible therapeutic effects of implementing a siRNA strategy to target COVID-19: Shoot the messenger! J. Cell Mol. Med. 2020, 24, 10267–10269. [Google Scholar] [CrossRef]
- Shawan, M.; Sharma, A.R.; Bhattacharya, M.; Mallik, B.; Akhter, F.; Shakil, M.S.; Hossain, M.M.; Banik, S.; Lee, S.S.; Hasan, M.A.; et al. Designing an effective therapeutic siRNA to silence RdRp gene of SARS-CoV-2. Infect. Genet. Evol. 2021, 93, 104951. [Google Scholar] [CrossRef]
- Chen, W.; Feng, P.; Liu, K.; Wu, M.; Lin, H. Computational Identification of Small Interfering RNA Targets in SARS-CoV-2. Virol. Sin. 2020, 35, 359–361. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.K.; Verma, S. An in silico analysis of effective siRNAs against COVID-19 by targeting the leader sequence of SARS-CoV-2. Adv. Cell Gene Ther. 2021, 15, e107. [Google Scholar] [CrossRef]
- Wu, R.; Luo, K.Q. Developing effective siRNAs to reduce the expression of key viral genes of COVID-19. Int. J. Biol. Sci. 2021, 17, 1521–1529. [Google Scholar] [CrossRef]
- Sohrab, S.S.; Aly El-Kafrawy, S.; Abbas, A.T.; Bajrai, L.H.; Azhar, E.I. In silico Prediction and Designing of Potential siRNAs to be Used as Antivirals Against SARS-CoV-2. Curr. Pharm. Des. 2021, 27, 3490–3500. [Google Scholar] [CrossRef]
- Zhang, D.; Lu, J. In Silico Design of siRNAs Targeting Existing and Future Respiratory Viruses with VirusSi. bioRxiv 2020. [Google Scholar] [CrossRef]
- Rizkita, L.D.; Astuti, I. The potential of miRNA-based therapeutics in severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection: A review. J. Pharm. Anal. 2021, 11, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Sohrab, S.S.; El-Kafrawy, S.A.; Mirza, Z.; Hassan, A.M.; Alsaqaf, F.; Azhar, E.I. Designing and evaluation of MERS-CoV siRNAs in HEK-293 cell line. J. Infect. Public Health 2021, 14, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Sohrab, S.S.; Aly El-Kafrawy, S.; Mirza, Z.; Hassan, A.M.; Alsaqaf, F.; Azhar, E.I. In silico prediction and experimental validation of siRNAs targeting ORF1ab of MERS-CoV in Vero cell line. Saudi J. Biol. Sci. 2021, 28, 1348–1355. [Google Scholar] [CrossRef] [PubMed]
- El-Kafrawy, S.A.; Sohrab, S.S.; Mirza, Z.; Hassan, A.M.; Alsaqaf, F.; Azhar, E.I. In Vitro Inhibitory Analysis of Rationally Designed siRNAs against MERS-CoV Replication in Huh7 Cells. Molecules 2021, 26, 2610. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.J.; Huang, H.W.; Liu, C.Y.; Hong, C.F.; Chan, Y.L. Inhibition of SARS-CoV replication by siRNA. Antivir. Res. 2005, 65, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.J.; Guan, Y.; Tang, Q.; Du, C.; Xie, F.Y.; He, M.L.; Chan, K.W.; Wong, K.L.; Lader, E.; Woodle, M.C.; et al. Prophylactic and therapeutic effects of small interfering RNA targeting SARS-coronavirus. Antivir. Ther. 2004, 9, 365–374. [Google Scholar] [PubMed]
- Li, B.J.; Tang, Q.; Cheng, D.; Qin, C.; Xie, F.Y.; Wei, Q.; Xu, J.; Liu, Y.; Zheng, B.J.; Woodle, M.C.; et al. Using siRNA in prophylactic and therapeutic regimens against SARS coronavirus in Rhesus macaque. Nat. Med. 2005, 11, 944–951. [Google Scholar] [CrossRef]
- He, M.L.; Zheng, B.; Peng, Y.; Peiris, J.S.; Poon, L.L.; Yuen, K.Y.; Lin, M.C.; Kung, H.F.; Guan, Y. Inhibition of SARS-associated coronavirus infection and replication by RNA interference. JAMA 2003, 290, 2665–2666. [Google Scholar] [CrossRef] [Green Version]
- He, M.L.; Zheng, B.J.; Chen, Y.; Wong, K.L.; Huang, J.D.; Lin, M.C.; Yuen, K.Y.; Sung, J.J.; Kung, H.F. Development of interfering RNA agents to inhibit SARS-associated coronavirus infection and replication. Hong Kong Med. J. 2009, 15 (Suppl. 4), 28–31. [Google Scholar]
- Shi, Y.; Yang, D.H.; Xiong, J.; Jia, J.; Huang, B.; Jin, Y.X. Inhibition of genes expression of SARS coronavirus by synthetic small interfering RNAs. Cell Res. 2005, 15, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ren, L.; Zhao, X.; Hung, T.; Meng, A.; Wang, J.; Chen, Y.G. Inhibition of severe acute respiratory syndrome virus replication by small interfering RNAs in mammalian cells. J. Virol. 2004, 78, 7523–7527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaitov, M.; Nikonova, A.; Shilovskiy, I.; Kozhikhova, K.; Kofiadi, I.; Vishnyakova, L.; Nikolskii, A.; Gattinger, P.; Kovchina, V.; Barvinskaia, E.; et al. Silencing of SARS-CoV-2 with modified siRNA-peptide dendrimer formulation. Allergy 2021, 76, 2840–2854. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, U.F.; Sharif Shohan, M.U.; Hoque, K.I.; Beg, M.A.; Sharif Siam, M.K.; Moni, M.A. A computational approach to design potential siRNA molecules as a prospective tool for silencing nucleocapsid phosphoprotein and surface glycoprotein gene of SARS-CoV-2. Genomics 2021, 113 Pt 1, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Bappy, S.S.; Shibly, A.Z.; Sultana, S.; Mohiuddin, A.K.M.; Kabir, Y. Designing potential siRNA molecule for the nucleocapsid(N) gene silencing of different SARS-CoV-2 strains of Bangladesh: Computational approach. Comput. Biol. Chem. 2021, 92, 107486. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Liu, R.; Wang, Y.; Wang, F.; Xia, G.; Zhang, H.; Li, X.; Yin, H.; Chen, B. PLGA-PLL-PEG-Tf-based targeted nanoparticles drug delivery system enhance antitumor efficacy via intrinsic apoptosis pathway. Int. J. Nanomed. 2015, 10, 557–566. [Google Scholar] [CrossRef]
- Mintzer, M.A.; Simanek, E.E. Nonviral vectors for gene delivery. Chem. Rev. 2009, 109, 259–302. [Google Scholar] [CrossRef] [PubMed]
- Itani, R.; Tobaiqy, M.; Al Faraj, A. Optimizing use of theranostic nanoparticles as a life-saving strategy for treating COVID-19 patients. Theranostics 2020, 10, 5932–5942. [Google Scholar] [CrossRef]
- Faria, M.J.; Lopes, C.M.; das Neves, J.; Lucio, M. Lipid Nanocarriers for Anti-HIV Therapeutics: A Focus on Physicochemical Properties and Biotechnological Advances. Pharmaceutics 2021, 13, 1294. [Google Scholar] [CrossRef]
- Suzuki, Y.; Ishihara, H. Difference in the lipid nanoparticle technology employed in three approved siRNA (Patisiran) and mRNA (COVID-19 vaccine) drugs. Drug Metab. Pharm. 2021, 41, 100424. [Google Scholar] [CrossRef]
- Ortega-Berlanga, B.; Gonzalez, C.; Navarro-Tovar, G. Recent Advances in the Use of Lipid-Based Nanoparticles Against Glioblastoma Multiforme. Arch. Immunol. Ther. Exp. 2021, 69, 8. [Google Scholar] [CrossRef]
- Kulkarni, J.A.; Witzigmann, D.; Chen, S.; Cullis, P.R.; van der Meel, R. Lipid Nanoparticle Technology for Clinical Translation of siRNA Therapeutics. Acc. Chem. Res. 2019, 52, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.K.; Nougarede, A.; Clement, F.; Fournier, C.; Jouvin-Marche, E.; Escude, M.; Jary, D.; Navarro, F.P.; Marche, P.N. Tuning the Immunostimulation Properties of Cationic Lipid Nanocarriers for Nucleic Acid Delivery. Front. Immunol. 2021, 12, 722411. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, J.A.; Darjuan, M.M.; Mercer, J.E.; Chen, S.; van der Meel, R.; Thewalt, J.L.; Tam, Y.Y.C.; Cullis, P.R. On the Formation and Morphology of Lipid Nanoparticles Containing Ionizable Cationic Lipids and siRNA. ACS Nano 2018, 12, 4787–4795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Choi, Y.K.; Kim, S.; Lee, J.; Im, W. CHARMM-GUI Membrane Builder for Lipid Nanoparticles with Ionizable Cationic Lipids and PEGylated Lipids. bioRxiv 2021, 61, 5192–5202. [Google Scholar] [CrossRef]
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D.J.A. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int. J. Pharm. 2021, 601, 120586. [Google Scholar] [CrossRef]
- Aldosari, B.N.; Alfagih, I.M.; Almurshedi, A.S. Lipid Nanoparticles as Delivery Systems for RNA-Based Vaccines. Pharmaceutics 2021, 13, 206. [Google Scholar] [CrossRef]
- Tao, W.; Davide, J.P.; Cai, M.; Zhang, G.J.; South, V.J.; Matter, A.; Ng, B.; Zhang, Y.; Sepp-Lorenzino, L. Noninvasive imaging of lipid nanoparticle-mediated systemic delivery of small-interfering RNA to the liver. Mol. Ther. 2010, 18, 1657–1666. [Google Scholar] [CrossRef]
- Basha, G.; Novobrantseva, T.I.; Rosin, N.; Tam, Y.Y.; Hafez, I.M.; Wong, M.K.; Sugo, T.; Ruda, V.M.; Qin, J.; Klebanov, B.; et al. Influence of cationic lipid composition on gene silencing properties of lipid nanoparticle formulations of siRNA in antigen-presenting cells. Mol. Ther. 2011, 19, 2186–2200. [Google Scholar] [CrossRef]
- Gilleron, J.; Querbes, W.; Zeigerer, A.; Borodovsky, A.; Marsico, G.; Schubert, U.; Manygoats, K.; Seifert, S.; Andree, C.; Stoter, M.; et al. Image-based analysis of lipid nanoparticle-mediated siRNA delivery, intracellular trafficking and endosomal escape. Nat. Biotechnol. 2013, 31, 638–646. [Google Scholar] [CrossRef]
- McLendon, J.M.; Joshi, S.R.; Sparks, J.; Matar, M.; Fewell, J.G.; Abe, K.; Oka, M.; McMurtry, I.F.; Gerthoffer, W.T. Lipid nanoparticle delivery of a microRNA-145 inhibitor improves experimental pulmonary hypertension. J. Control. Release 2015, 210, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Basha, G.; Ordobadi, M.; Scott, W.R.; Cottle, A.; Liu, Y.; Wang, H.; Cullis, P.R. Lipid Nanoparticle Delivery of siRNA to Osteocytes Leads to Effective Silencing of SOST and Inhibition of Sclerostin In Vivo. Mol. Ther. Nucleic Acids 2016, 5, e363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wei, X.; Sun, X.; Chen, C.; Zhou, J.; Zhang, G.; Wu, H.; Guo, B.; Wei, L. A novel therapeutic strategy for cartilage diseases based on lipid nanoparticle-RNAi delivery system. Int. J. Nanomed. 2018, 13, 617–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhao, Y.; Zhang, E.; Jiang, M.; Zhi, D.; Chen, H.; Cui, S.; Zhen, Y.; Cui, J.; Zhang, S. Co-delivery of paclitaxel and anti-VEGF siRNA by tripeptide lipid nanoparticle to enhance the anti-tumor activity for lung cancer therapy. Drug Deliv. 2020, 27, 1397–1411. [Google Scholar] [CrossRef] [PubMed]
- Knapp, C.M.; He, J.; Lister, J.; Whitehead, K.A. Lipid nanoparticle siRNA cocktails for the treatment of mantle cell lymphoma. Bioeng. Transl. Med. 2018, 3, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Jyotsana, N.; Sharma, A.; Chaturvedi, A.; Budida, R.; Scherr, M.; Kuchenbauer, F.; Lindner, R.; Noyan, F.; Suhs, K.W.; Stangel, M.; et al. Lipid nanoparticle-mediated siRNA delivery for safe targeting of human CML in vivo. Ann. Hematol. 2019, 98, 1905–1918. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, J.A.; Witzigmann, D.; Leung, J.; Tam, Y.Y.C.; Cullis, P.R. On the role of helper lipids in lipid nanoparticle formulations of siRNA. Nanoscale 2019, 11, 21733–21739. [Google Scholar] [CrossRef] [PubMed]
- Schlich, M.; Palomba, R.; Costabile, G.; Mizrahy, S.; Pannuzzo, M.; Peer, D.; Decuzzi, P. Cytosolic delivery of nucleic acids: The case of ionizable lipid nanoparticles. Bioeng. Transl. Med. 2021, 6, e10213. [Google Scholar] [CrossRef]
- Law, S.L.; Huang, K.J.; Chou, V.H.; Cherng, J.Y. Enhancement of nasal absorption of calcitonin loaded in liposomes. J. Liposome Res. 2001, 11, 165–174. [Google Scholar] [CrossRef]
- Seyfoori, A.; Shokrollahi Barough, M.; Mokarram, P.; Ahmadi, M.; Mehrbod, P.; Sheidary, A.; Madrakian, T.; Kiumarsi, M.; Walsh, T.; McAlinden, K.D.; et al. Emerging Advances of Nanotechnology in Drug and Vaccine Delivery against Viral Associated Respiratory Infectious Diseases (VARID). Int J. Mol. Sci 2021, 22, 6937. [Google Scholar] [CrossRef]
- Kulkarni, J.A.; Thomson, S.B.; Zaifman, J.; Leung, J.; Wagner, P.K.; Hill, A.; Tam, Y.Y.C.; Cullis, P.R.; Petkau, T.L.; Leavitt, B.R. Spontaneous, solvent-free entrapment of siRNA within lipid nanoparticles. Nanoscale 2020, 12, 23959–23966. [Google Scholar] [CrossRef]
- Dobrowolski, C.; Paunovska, K.; Hatit, M.Z.C.; Lokugamage, M.P.; Dahlman, J.E. Therapeutic RNA Delivery for COVID and Other Diseases. Adv. Healthc. Mater. 2021, 10, e2002022. [Google Scholar] [CrossRef] [PubMed]
- Jorge, A.; Pais, A.; Vitorino, C. Targeted siRNA Delivery Using Lipid Nanoparticles. Methods Mol. Biol 2020, 2059, 259–283. [Google Scholar] [PubMed]
- Taratula, O.; Kuzmov, A.; Shah, M.; Garbuzenko, O.B.; Minko, T. Nanostructured lipid carriers as multifunctional nanomedicine platform for pulmonary co-delivery of anticancer drugs and siRNA. J. Control. Release 2013, 171, 349–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajik-Ahmadabad, B.; Mechler, A.; Muir, B.W.; McLean, K.; Hinton, T.M.; Separovic, F.; Polyzos, A. A QCM-D and SAXS Study of the Interaction of Functionalised Lyotropic Liquid Crystalline Lipid Nanoparticles with siRNA. Chembiochem 2017, 18, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zakrewsky, M.; Gupta, V.; Anselmo, A.C.; Slee, D.H.; Muraski, J.A.; Mitragotri, S. Topical delivery of siRNA into skin using SPACE-peptide carriers. J. Control. Release 2014, 179, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Lin, W.; Li, M.; Yang, Y.; Deng, J.; Liu, H.; Chen, Y.; Fu, X.; Liu, H.; Yang, Y. Efficient siRNA Delivery Using Novel Cell-Penetrating Peptide-siRNA Conjugate-Loaded Nanobubbles and Ultrasound. Ultrasound Med. Biol. 2016, 42, 1362–1374. [Google Scholar] [CrossRef]
- Connerty, P.; Moles, E.; de Bock, C.E.; Jayatilleke, N.; Smith, J.L.; Meshinchi, S.; Mayoh, C.; Kavallaris, M.; Lock, R.B. Development of siRNA-Loaded Lipid Nanoparticles Targeting Long Non-Coding RNA LINC01257 as a Novel and Safe Therapeutic Approach for t(8;21) Pediatric Acute Myeloid Leukemia. Pharmaceutics 2021, 13, 1681. [Google Scholar] [CrossRef]
- Kawase, W.; Kurotaki, D.; Suzuki, Y.; Ishihara, H.; Ban, T.; Sato, G.R.; Ichikawa, J.; Yanai, H.; Taniguchi, T.; Tsukahara, K.; et al. Irf5 siRNA-loaded biodegradable lipid nanoparticles ameliorate concanavalin A-induced liver injury. Mol. Ther. Nucleic Acids 2021, 25, 708–715. [Google Scholar] [CrossRef]
- Sanghani, A.; Kafetzis, K.N.; Sato, Y.; Elboraie, S.; Fajardo-Sanchez, J.; Harashima, H.; Tagalakis, A.D.; Yu-Wai-Man, C. Novel PEGylated Lipid Nanoparticles Have a High Encapsulation Efficiency and Effectively Deliver MRTF-B siRNA in Conjunctival Fibroblasts. Pharmaceutics 2021, 13, 382. [Google Scholar] [CrossRef]
- Chen, D.; Ganesh, S.; Wang, W.; Lupieri, A.; Amiji, M. Role of vitronectin-rich protein corona on tumor-specific siRNA delivery and transfection with lipid nanoparticles. Nanomedicine 2021, 16, 535–551. [Google Scholar] [CrossRef]
- Huang, X.; Chau, Y. Enhanced Delivery of siRNA to Retinal Ganglion Cells by Intravitreal Lipid Nanoparticles of Positive Charge. Mol. Pharm. 2021, 18, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Shobaki, N.; Sato, Y.; Suzuki, Y.; Okabe, N.; Harashima, H. Manipulating the function of tumor-associated macrophages by siRNA-loaded lipid nanoparticles for cancer immunotherapy. J. Control. Release 2020, 325, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Younis, M.A.; Khalil, I.A.; Elewa, Y.H.A.; Kon, Y.; Harashima, H. Ultra-small lipid nanoparticles encapsulating sorafenib and midkine-siRNA selectively-eradicate sorafenib-resistant hepatocellular carcinoma in vivo. J. Control. Release 2021, 331, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, M.; Woloshun, R.R.; Yu, Y.; Lee, J.K.; Flores, S.R.L.; Merlin, D.; Collins, J.F. Oral Administration of Ginger-Derived Lipid Nanoparticles and Dmt1 siRNA Potentiates the Effect of Dietary Iron Restriction and Mitigates Pre-Existing Iron Overload in Hamp KO Mice. Nutrients 2021, 13, 1686. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, M.S.; Hufnagel, S.; Trementozzi, A.N.; Sakran, W.; Stachowiak, J.C.; Koleng, J.J.; Cui, Z. PD-1 siRNA-Encapsulated Solid Lipid Nanoparticles Downregulate PD-1 Expression by Macrophages and Inhibit Tumor Growth: PD-1 siRNA-Encapsulated Solid Lipid Nanoparticles. AAPS PharmSciTech 2021, 22, 60. [Google Scholar] [CrossRef]
- Wang, J.L.; Hanafy, M.S.; Xu, H.; Leal, J.; Zhai, Y.; Ghosh, D.; Williams Iii, R.O.; David Charles Smyth, H.; Cui, Z. Aerosolizable siRNA-encapsulated solid lipid nanoparticles prepared by thin-film freeze-drying for potential pulmonary delivery. Int. J. Pharm. 2021, 596, 120215. [Google Scholar] [CrossRef]
- Buyukkoroglu, G.; Senel, B.; Yenilmez, E. Vaginal Suppositories with siRNA and Paclitaxel-Incorporated Solid Lipid Nanoparticles for Cervical Cancer: Preparation and In Vitro Evaluation. Methods Mol. Biol. 2019, 1974, 303–328. [Google Scholar]
- Rassu, G.; Soddu, E.; Posadino, A.M.; Pintus, G.; Sarmento, B.; Giunchedi, P.; Gavini, E. Nose-to-brain delivery of BACE1 siRNA loaded in solid lipid nanoparticles for Alzheimer’s therapy. Colloids Surf. B Biointerfaces 2017, 152, 296–301. [Google Scholar] [CrossRef]
- Huang, R.; Yao, X.; Chen, Y.; Sun, X.; Lin, Y. [Cytological Study in vitro on Co-delivery of siRNA and Paclitaxel within Solid Lipid Nanoparticles to Overcome Multidrug Resistance in Tumors]. Sheng Wu Yi Xue Gong Cheng Xue Za Zhi 2016, 33, 108–114. [Google Scholar]
- Ying, B.; Campbell, R.B. Delivery of kinesin spindle protein targeting siRNA in solid lipid nanoparticles to cellular models of tumor vasculature. Biochem. Biophys. Res. Commun. 2014, 446, 441–447. [Google Scholar] [CrossRef]
- Bae, K.H.; Lee, J.Y.; Lee, S.H.; Park, T.G.; Nam, Y.S. Optically traceable solid lipid nanoparticles loaded with siRNA and paclitaxel for synergistic chemotherapy with in situ imaging. Adv. Healthc. Mater. 2013, 2, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.H.; Kim, E.; Park, D.E.; Shim, G.; Lee, S.; Kim, Y.B.; Kim, C.W.; Oh, Y.K. Cationic solid lipid nanoparticles for co-delivery of paclitaxel and siRNA. Eur. J. Pharm. Biopharm. 2012, 80, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Bae, K.H.; Yang, H.; Lee, S.J.; Kim, H.; Kim, Y.; Joo, K.M.; Seo, S.W.; Park, T.G.; Nam, D.H. In vivo specific delivery of c-Met siRNA to glioblastoma using cationic solid lipid nanoparticles. Bioconjug. Chem. 2011, 22, 2568–2572. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Kim, I.K.; Bae, K.H.; Lee, S.H.; Lee, Y.; Park, T.G. Cationic solid lipid nanoparticles reconstituted from low density lipoprotein components for delivery of siRNA. Mol. Pharm. 2008, 5, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Oner, E.; Kotmakci, M.; Baird, A.M.; Gray, S.G.; Debelec Butuner, B.; Bozkurt, E.; Kantarci, A.G.; Finn, S.P. Development of EphA2 siRNA-loaded lipid nanoparticles and combination with a small-molecule histone demethylase inhibitor in prostate cancer cells and tumor spheroids. J. Nanobiotechnol. 2021, 19, 71. [Google Scholar] [CrossRef]
- Chenthamara, D.; Subramaniam, S.; Ramakrishnan, S.G.; Krishnaswamy, S.; Essa, M.M.; Lin, F.H.; Qoronfleh, M.W. Therapeutic efficacy of nanoparticles and routes of administration. Biomater. Res. 2019, 23, 20. [Google Scholar] [CrossRef]
- Zielinska, A.; Carreiro, F.; Oliveira, A.M.; Neves, A.; Pires, B.; Venkatesh, D.N.; Durazzo, A.; Lucarini, M.; Eder, P.; Silva, A.M.; et al. Polymeric Nanoparticles: Production, Characterization, Toxicology and Ecotoxicology. Molecules 2020, 25, 3731. [Google Scholar] [CrossRef]
- Silva, A.T.; Nguyen, A.; Ye, C.; Verchot, J.; Moon, J.H. Conjugated polymer nanoparticles for effective siRNA delivery to tobacco BY-2 protoplasts. BMC Plant. Biol. 2010, 10, 291. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.H.; Mendez, E.; Kim, Y.; Kaur, A. Conjugated polymer nanoparticles for small interfering RNA delivery. Chem. Commun. 2011, 47, 8370–8372. [Google Scholar] [CrossRef]
- Devulapally, R.; Paulmurugan, R. Polymer nanoparticles for drug and small silencing RNA delivery to treat cancers of different phenotypes. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2014, 6, 40–60. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, S.; Ye, W.H.; Yoon, H.S.; Yang, Y.Y. Co-delivery of drugs and DNA from cationic core-shell nanoparticles self-assembled from a biodegradable copolymer. Nat. Mater. 2006, 5, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Beh, C.W.; Seow, W.Y.; Wang, Y.; Zhang, Y.; Ong, Z.Y.; Ee, P.L.; Yang, Y.Y. Efficient delivery of Bcl-2-targeted siRNA using cationic polymer nanoparticles: Downregulating mRNA expression level and sensitizing cancer cells to anticancer drug. Biomacromolecules 2009, 10, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Heo, M.B.; Cho, M.Y.; Lim, Y.T. Polymer nanoparticles for enhanced immune response: Combined delivery of tumor antigen and small interference RNA for immunosuppressive gene to dendritic cells. Acta Biomater. 2014, 10, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Tian, H.; Wang, P.; Wang, Y.; Chen, X. The suppression of metastatic lung cancer by pulmonary administration of polymer nanoparticles for co-delivery of doxorubicin and Survivin siRNA. Biomater. Sci. 2016, 4, 1646–1654. [Google Scholar] [CrossRef]
- Boyer, C.; Teo, J.; Phillips, P.; Erlich, R.B.; Sagnella, S.; Sharbeen, G.; Dwarte, T.; Duong, H.T.; Goldstein, D.; Davis, T.P.; et al. Effective delivery of siRNA into cancer cells and tumors using well-defined biodegradable cationic star polymers. Mol. Pharm. 2013, 10, 2435–2444. [Google Scholar] [CrossRef] [Green Version]
- Teo, J.; McCarroll, J.A.; Boyer, C.; Youkhana, J.; Sagnella, S.M.; Duong, H.T.; Liu, J.; Sharbeen, G.; Goldstein, D.; Davis, T.P.; et al. A Rationally Optimized Nanoparticle System for the Delivery of RNA Interference Therapeutics into Pancreatic Tumors in Vivo. Biomacromolecules 2016, 17, 2337–2351. [Google Scholar] [CrossRef] [Green Version]
- McCarroll, J.A.; Sharbeen, G.; Kavallaris, M.; Phillips, P.A. The Use of Star Polymer Nanoparticles for the Delivery of siRNA to Mouse Orthotopic Pancreatic Tumor Models. Methods Mol. Biol. 2019, 1974, 329–353. [Google Scholar]
- Bayat, N.; McOrist, N.; Ariotti, N.; Lai, M.; Sia, K.C.; Li, Y.; Grace, J.L.; Quinn, J.F.; Whittaker, M.R.; Kavallaris, M.; et al. Thiol-Reactive Star Polymers Functionalized with Short Ethoxy-Containing Moieties Exhibit Enhanced Uptake in Acute Lymphoblastic Leukemia Cells. Int. J. Nanomed. 2019, 14, 9795–9808. [Google Scholar] [CrossRef] [Green Version]
- Dutta, K.; Bochicchio, D.; Ribbe, A.E.; Alfandari, D.; Mager, J.; Pavan, G.M.; Thayumanavan, S. Symbiotic Self-Assembly Strategy toward Lipid-Encased Cross-Linked Polymer Nanoparticles for Efficient Gene Silencing. ACS Appl. Mater. Interfaces 2019, 11, 24971–24983. [Google Scholar] [CrossRef]
- Shi, J.; Xu, Y.; Xu, X.; Zhu, X.; Pridgen, E.; Wu, J.; Votruba, A.R.; Swami, A.; Zetter, B.R.; Farokhzad, O.C. Hybrid lipid-polymer nanoparticles for sustained siRNA delivery and gene silencing. Nanomedicine 2014, 10, 897–900. [Google Scholar] [CrossRef]
- Xu, C.-F.; Liu, Y.; Shen, S.; Zhu, Y.-H.; Wang, J. Targeting glucose uptake of glioma cells by siRNA delivery with polymer nanoparticle. J. Control. Release 2015, 213, e23–e24. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, K.A.; Cu, Y.; Booth, C.J.; Saucier-Sawyer, J.K.; Wood, M.J.; Saltzman, W.M. Intravaginal gene silencing using biodegradable polymer nanoparticles densely loaded with small-interfering RNA. Nat. Mater. 2009, 8, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Luo, H.C.; Yang, C.; Deng, J.J.; Ren, M.; Xie, X.Y.; Lin, D.Z.; Yan, L.; Zhang, L.M. Cationic star-shaped polymer as an siRNA carrier for reducing MMP-9 expression in skin fibroblast cells and promoting wound healing in diabetic rats. Int. J. Nanomed. 2014, 9, 3377–3387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbach, J.M.; Weller, C.E.; Booth, C.J.; Saltzman, W.M. Polymer nanoparticles encapsulating siRNA for treatment of HSV-2 genital infection. J. Control. Release 2012, 162, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- d’Angelo, I.; Costabile, G.; Durantie, E.; Brocca, P.; Rondelli, V.; Russo, A.; Russo, G.; Miro, A.; Quaglia, F.; Petri-Fink, A.; et al. Hybrid Lipid/Polymer Nanoparticles for Pulmonary Delivery of siRNA: Development and Fate Upon In Vitro Deposition on the Human Epithelial Airway Barrier. J. Aerosol. Med. Pulm. Drug Deliv. 2018, 31, 170–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koide, H.; Fukuta, T.; Okishim, A.; Ariizumi, S.; Kiyokawa, C.; Tsuchida, H.; Nakamoto, M.; Yoshimatsu, K.; Ando, H.; Dewa, T.; et al. Engineering the Binding Kinetics of Synthetic Polymer Nanoparticles for siRNA Delivery. Biomacromolecules 2019, 20, 3648–3657. [Google Scholar] [CrossRef]
- Sarett, S.M.; Kilchrist, K.V.; Miteva, M.; Duvall, C.L. Conjugation of palmitic acid improves potency and longevity of siRNA delivered via endosomolytic polymer nanoparticles. J. Biomed. Mater. Res. A 2015, 103, 3107–3116. [Google Scholar] [CrossRef] [Green Version]
- Dearnley, M.; Reynolds, N.P.; Cass, P.; Wei, X.; Shi, S.; Mohammed, A.A.; Le, T.; Gunatillake, P.; Tizard, M.L.; Thang, S.H.; et al. Comparing Gene Silencing and Physiochemical Properties in siRNA Bound Cationic Star-Polymer Complexes. Biomacromolecules 2016, 17, 3532–3546. [Google Scholar] [CrossRef]
- Dragoni, L.; Ferrari, R.; Lupi, M.; Cesana, A.; Falcetta, F.; Ubezio, P.; D’Incalci, M.; Morbidelli, M.; Moscatelli, D. Small interfering RNA delivery through positively charged polymer nanoparticles. Nanotechnology 2016, 27, 125102. [Google Scholar] [CrossRef]
- Ullah, A.; Qazi, J.; Rahman, L.; Kanaras, A.G.; Khan, W.S.; Hussain, I.; Rehman, A. Nanoparticles-assisted delivery of antiviral-siRNA as inhalable treatment for human respiratory viruses: A candidate approach against SARS-CoV-2. Nano Sel. 2020, 1, 612–621. [Google Scholar] [CrossRef]
- Wojnilowicz, M.; Glab, A.; Bertucci, A.; Caruso, F.; Cavalieri, F. Super-resolution Imaging of Proton Sponge-Triggered Rupture of Endosomes and Cytosolic Release of Small Interfering RNA. ACS Nano 2019, 13, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Besford, Q.A.; Cavalieri, F.; Caruso, F. Glycogen as a Building Block for Advanced Biological Materials. Adv. Mater. 2020, 32, e1904625. [Google Scholar] [CrossRef] [PubMed]
- Pacchin Tomanin, P.; Zhou, J.; Amodio, A.; Cimino, R.; Glab, A.; Cavalieri, F.; Caruso, F. Nanoengineering multifunctional hybrid interfaces using adhesive glycogen nanoparticles. J. Mater. Chem. B 2020, 8, 4851–4858. [Google Scholar] [CrossRef] [PubMed]
- Besford, Q.A.; Weiss, A.C.G.; Schubert, J.; Ryan, T.M.; Maitz, M.F.; Tomanin, P.P.; Savioli, M.; Werner, C.; Fery, A.; Caruso, F.; et al. Protein Component of Oyster Glycogen Nanoparticles: An Anchor Point for Functionalization. ACS Appl. Mater. Interfaces 2020, 12, 38976–38988. [Google Scholar] [CrossRef]
- Sonaje, K.; Chuang, E.Y.; Lin, K.J.; Yen, T.C.; Su, F.Y.; Tseng, M.T.; Sung, H.W. Opening of epithelial tight junctions and enhancement of paracellular permeation by chitosan: Microscopic, ultrastructural, and computed-tomographic observations. Mol. Pharm. 2012, 9, 1271–1279. [Google Scholar] [CrossRef]
- Besford, Q.A.; Wojnilowicz, M.; Suma, T.; Bertleff-Zieschang, N.; Caruso, F.; Cavalieri, F. Lactosylated Glycogen Nanoparticles for Targeting Prostate Cancer Cells. ACS Appl. Mater. Interfaces 2017, 9, 16869–16879. [Google Scholar] [CrossRef]
- Wojnilowicz, M.; Besford, Q.A.; Wu, Y.L.; Loh, X.J.; Braunger, J.A.; Glab, A.; Cortez-Jugo, C.; Caruso, F.; Cavalieri, F. Glycogen-nucleic acid constructs for gene silencing in multicellular tumor spheroids. Biomaterials 2018, 176, 34–49. [Google Scholar] [CrossRef]
- Cimino, R.; Bhangu, S.K.; Baral, A.; Ashokkumar, M.; Cavalieri, F. Ultrasound-Assisted Microencapsulation of Soybean Oil and Vitamin D Using Bare Glycogen Nanoparticles. Molecules 2021, 26, 5157. [Google Scholar] [CrossRef]
- Conde, J.; Tian, F.; Hernandez, Y.; Bao, C.; Baptista, P.V.; Cui, D.; Stoeger, T.; de la Fuente, J.M. RNAi-based glyconanoparticles trigger apoptotic pathways for in vitro and in vivo enhanced cancer-cell killing. Nanoscale 2015, 7, 9083–9091. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Zhang, X.; Li, J.; Li, Y.; Wang, Q.; Teng, W. Comparison of osteogenic differentiation induced by siNoggin and pBMP-2 delivered by lipopolysaccharide-amine nanopolymersomes and underlying molecular mechanisms. Int. J. Nanomed. 2019, 14, 4229–4245. [Google Scholar] [CrossRef] [Green Version]
- Engelberth, S.A.; Hempel, N.; Bergkvist, M. Cationic dendritic starch as a vehicle for photodynamic therapy and siRNA co-delivery. J. Photochem. Photobiol. B 2017, 168, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Gong, H.; Zeng, R.; Liang, X.; Zhang, L.M.; Yang, L.; Lan, Y. Efficient delivery of NF-kappaB siRNA to human retinal pigment epithelial cells with hyperbranched cationic polysaccharide derivative-based nanoparticles. Int. J. Nanomed. 2015, 10, 2735–2749. [Google Scholar]
- Gálisová, A.; Jirátová, M.; Rabyk, M.; Sticová, E.; Hájek, M.; Hrubý, M.; Jirák, D. Glycogen as an advantageous polymer carrier in cancer theranostics: Straightforward in vivo evidence. Sci. Rep. 2020, 10, 10411. [Google Scholar] [CrossRef] [PubMed]
- Kirkland-York, S.; Zhang, Y.; Smith, A.E.; York, A.W.; Huang, F.; McCormick, C.L. Tailored design of Au nanoparticle-siRNA carriers utilizing reversible addition-fragmentation chain transfer polymers. Biomacromolecules 2010, 11, 1052–1059. [Google Scholar] [CrossRef]
- Marques Neto, L.M.; Kipnis, A.; Junqueira-Kipnis, A.P. Role of Metallic Nanoparticles in Vaccinology: Implications for Infectious Disease Vaccine Development. Front. Immunol. 2017, 8, 239. [Google Scholar] [CrossRef] [Green Version]
- Lytton-Jean, A.K.; Langer, R.; Anderson, D.G. Five years of siRNA delivery: Spotlight on gold nanoparticles. Small 2011, 7, 1932–1937. [Google Scholar] [CrossRef]
- Behrouzi, K.; Lin, L. Gold nanoparticle based plasmonic sensing for the detection of SARS-CoV-2 nucleocapsid proteins. Biosens. Bioelectron. 2021, 195, 113669. [Google Scholar] [CrossRef]
- Aithal, S.; Mishriki, S.; Gupta, R.; Sahu, R.P.; Botos, G.; Tanvir, S.; Hanson, R.W.; Puri, I.K. SARS-CoV-2 detection with aptamer-functionalized gold nanoparticles. Talanta 2022, 236, 122841. [Google Scholar] [CrossRef]
- El-Dakdouki, M.H.; El-Boubbou, K.; Kamat, M.; Huang, R.; Abela, G.S.; Kiupel, M.; Zhu, D.C.; Huang, X. CD44 targeting magnetic glyconanoparticles for atherosclerotic plaque imaging. Pharm. Res. 2014, 31, 1426–1437. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; Rösch, E.L.; Viereck, T.; Schilling, M.; Ludwig, F. Toward Rapid and Sensitive Detection of SARS-CoV-2 with Functionalized Magnetic Nanoparticles. ACS Sens. 2021, 6, 976–984. [Google Scholar] [CrossRef]
- Acharya, R. The recent progresses in shRNA-nanoparticle conjugate as a therapeutic approach. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 104, 109928. [Google Scholar] [CrossRef] [PubMed]
- Acharya, R. Prospective vaccination of COVID-19 using shRNA-plasmid-LDH nanoconjugate. Med. Hypotheses 2020, 143, 110084. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Xiong, S. Tagged extracellular vesicles with the RBD of the viral spike protein for delivery of antiviral agents against SARS-CoV-2 infection. J. Control. Release 2021, 335, 584–595. [Google Scholar] [CrossRef] [PubMed]
- El-Megharbel, S.M.; Alsawat, M.; Al-Salmi, F.A.; Hamza, R.Z. Utilizing of (Zinc Oxide Nano-Spray) for Disinfection against “SARS-CoV-2” and Testing Its Biological Effectiveness on Some Biochemical Parameters during (COVID-19 Pandemic)—”ZnO Nanoparticles Have Antiviral Activity against (SARS-CoV-2)”. Coatings 2021, 11, 388. [Google Scholar] [CrossRef]
- Hamza, R.Z.; Gobouri, A.A.; Al-Yasi, H.M.; Al-Talhi, T.A.; El-Megharbel, S.M. A New Sterilization Strategy Using TiO2 Nanotubes for Production of Free Radicals that Eliminate Viruses and Application of a Treatment Strategy to Combat Infections Caused by Emerging SARS-CoV-2 during the COVID-19 Pandemic. Coatings 2021, 11, 680. [Google Scholar] [CrossRef]
- Haasnoot, J.; Westerhout, E.M.; Berkhout, B. RNA interference against viruses: Strike and counterstrike. Nat. Biotechnol. 2007, 25, 1435–1443. [Google Scholar] [CrossRef]
- Ketzinel-Gilad, M.; Shaul, Y.; Galun, E. RNA interference for antiviral therapy. J. Gene Med. 2006, 8, 933–950. [Google Scholar] [CrossRef]
- Mendez, C.; Ledger, S.; Petoumenos, K.; Ahlenstiel, C.; Kelleher, A.D. RNA-induced epigenetic silencing inhibits HIV-1 reactivation from latency. Retrovirology 2018, 15, 67. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Ou, M.; Keough, E.; Roberts, J.; Koeplinger, K.; Lyman, M.; Fauty, S.; Carlini, E.; Stern, M.; Zhang, R.; et al. Quantitation of physiological and biochemical barriers to siRNA liver delivery via lipid nanoparticle platform. Mol. Pharm. 2014, 11, 1424–1434. [Google Scholar] [CrossRef]
- Keil, T.W.; Merkel, O.M. Dry powder inhalation of siRNA. Ther. Deliv. 2019, 10, 265–267. [Google Scholar] [CrossRef] [Green Version]
- Mehta, A.; Michler, T.; Merkel, O.M. siRNA Therapeutics against Respiratory Viral Infections-What Have We Learned for Potential COVID-19 Therapies? Adv. Healthc. Mater. 2021, 10, e2001650. [Google Scholar] [CrossRef] [PubMed]
- Merkel, O.M.; Kissel, T. Nonviral pulmonary delivery of siRNA. Acc. Chem. Res. 2012, 45, 961–970. [Google Scholar] [CrossRef]
- Ou, C.; Hang, J.; Deng, Q. Particle Deposition in Human Lung Airways: Effects of Airflow, Particle Size, and Mechanisms. Aerosol Air Qual. Res. 2020, 20, 2846–2858. [Google Scholar] [CrossRef]
- Thomas, R.J. Particle size and pathogenicity in the respiratory tract. Virulence 2013, 4, 847–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsson, J.K.F.; Aaltonen, H.L.; Nicklasson, H.; Gudmundsson, A.; Rissler, J.; Wollmer, P.; Londahl, J. Altered deposition of inhaled nanoparticles in subjects with chronic obstructive pulmonary disease. BMC Pulm. Med. 2018, 18, 129. [Google Scholar] [CrossRef] [PubMed]
- Londahl, J.; Moller, W.; Pagels, J.H.; Kreyling, W.G.; Swietlicki, E.; Schmid, O. Measurement techniques for respiratory tract deposition of airborne nanoparticles: A critical review. J. Aerosol Med. Pulm. Drug Deliv. 2014, 27, 229–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Seth, A.; Wibowo, N.; Zhao, C.X.; Mitter, N.; Yu, C.; Middelberg, A.P. Nanoparticle vaccines. Vaccine 2014, 32, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Alshweiat, A.; Ambrus, R.; Csoka, I. Intranasal Nanoparticulate Systems as Alternative Route of Drug Delivery. Curr. Med. Chem. 2019, 26, 6459–6492. [Google Scholar] [CrossRef] [Green Version]
- Frede, A.; Neuhaus, B.; Knuschke, T.; Wadwa, M.; Kollenda, S.; Klopfleisch, R.; Hansen, W.; Buer, J.; Bruder, D.; Epple, M.; et al. Local delivery of siRNA-loaded calcium phosphate nanoparticles abates pulmonary inflammation. Nanomedicine 2017, 13, 2395–2403. [Google Scholar] [CrossRef]
- Conte, C.; Monteiro, P.F.; Gurnani, P.; Stolnik, S.; Ungaro, F.; Quaglia, F.; Clarke, P.; Grabowska, A.; Kavallaris, M.; Alexander, C. Multi-component bioresponsive nanoparticles for synchronous delivery of docetaxel and TUBB3 siRNA to lung cancer cells. Nanoscale 2021, 13, 11414–11426. [Google Scholar] [CrossRef]
- Hibbitts, A.J.; Ramsey, J.M.; Barlow, J.; MacLoughlin, R.; Cryan, S.A. In Vitro and In Vivo Assessment of PEGylated PEI for Anti-IL-8/CxCL-1 siRNA Delivery to the Lungs. Nanomaterials 2020, 10, 1248. [Google Scholar] [CrossRef]
- Bohr, A.; Tsapis, N.; Foged, C.; Andreana, I.; Yang, M.; Fattal, E. Treatment of acute lung inflammation by pulmonary delivery of anti-TNF-alpha siRNA with PAMAM dendrimers in a murine model. Eur. J. Pharm. Biopharm. 2020, 156, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Li, H.; Cui, H.; Bi, Q.; Wang, M. Solid lipid nanoparticle delivery of rhynchophylline enhanced the efficiency of allergic asthma treatment via the upregulation of suppressor of cytokine signaling 1 by repressing the p38 signaling pathway. Bioengineered 2021, 12, 8635–8649. [Google Scholar] [CrossRef] [PubMed]
- Falciani, C.; Zevolini, F.; Brunetti, J.; Riolo, G.; Gracia, R.; Marradi, M.; Loinaz, I.; Ziemann, C.; Cossio, U.; Llop, J.; et al. Antimicrobial Peptide-Loaded Nanoparticles as Inhalation Therapy for Pseudomonas aeruginosa Infections. Int. J. Nanomed. 2020, 15, 1117–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merckx, P.; De Backer, L.; Van Hoecke, L.; Guagliardo, R.; Echaide, M.; Baatsen, P.; Olmeda, B.; Saelens, X.; Perez-Gil, J.; De Smedt, S.C.; et al. Surfactant protein B (SP-B) enhances the cellular siRNA delivery of proteolipid coated nanogels for inhalation therapy. Acta Biomater. 2018, 78, 236–246. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, S.I.; El-Gendy, N.; Bowen, L.E.; Berkland, C.; Bailey, M.M. Formulation and Characterization of Nanocluster Ceftazidime for the Treatment of Acute Pulmonary Melioidosis. J. Pharm. Sci. 2016, 105, 3399–3408. [Google Scholar] [CrossRef]
- Huang, X.; Chisholm, J.; Zhuang, J.; Xiao, Y.; Duncan, G.; Chen, X.; Suk, J.S.; Hanes, J. Protein nanocages that penetrate airway mucus and tumor tissue. Proc. Natl. Acad. Sci. USA 2017, 114, E6595–E6602. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.S.; Xu, Q.; Boylan, N.J.; Chisholm, J.; Tang, B.C.; Schuster, B.S.; Henning, A.; Ensign, L.M.; Lee, E.; Adstamongkonkul, P.; et al. Nanoparticles that do not adhere to mucus provide uniform and long-lasting drug delivery to airways following inhalation. Sci. Adv. 2017, 3, e1601556. [Google Scholar] [CrossRef] [Green Version]
- Gulin-Sarfraz, T.; Jonasson, S.; Wigenstam, E.; von Haartman, E.; Bucht, A.; Rosenholm, J.M. Feasibility Study of Mesoporous Silica Particles for Pulmonary Drug Delivery: Therapeutic Treatment with Dexamethasone in a Mouse Model of Airway Inflammation. Pharmaceutics 2019, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Popov, A.; Schopf, L.; Bourassa, J.; Chen, H. Enhanced pulmonary delivery of fluticasone propionate in rodents by mucus-penetrating nanoparticles. Int. J. Pharm. 2016, 502, 188–197. [Google Scholar] [CrossRef]
- Li, X.; Xue, M.; Raabe, O.G.; Aaron, H.L.; Eisen, E.A.; Evans, J.E.; Hayes, F.A.; Inaga, S.; Tagmount, A.; Takeuchi, M.; et al. Aerosol droplet delivery of mesoporous silica nanoparticles: A strategy for respiratory-based therapeutics. Nanomedicine 2015, 11, 1377–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Zhou, K.; Xiong, H.; Miller, J.B.; Motea, E.A.; Boothman, D.A.; Liu, L.; Siegwart, D.J. Aerosol delivery of stabilized polyester-siRNA nanoparticles to silence gene expression in orthotopic lung tumors. Biomaterials 2017, 118, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.; Bardoliwala, D.; Lalani, R.; Patil, S.; Ghosh, S.; Javia, A.; Misra, A. Development of a dry powder for inhalation of nanoparticles codelivering cisplatin and ABCC3 siRNA in lung cancer. Ther. Deliv. 2021, 12, 651–670. [Google Scholar] [CrossRef] [PubMed]
- Merckx, P.; Lammens, J.; Nuytten, G.; Bogaert, B.; Guagliardo, R.; Maes, T.; Vervaet, C.; De Beer, T.; De Smedt, S.C.; Raemdonck, K. Lyophilization and nebulization of pulmonary surfactant-coated nanogels for siRNA inhalation therapy. Eur. J. Pharm. Biopharm. 2020, 157, 191–199. [Google Scholar] [CrossRef]
- De Backer, L.; Naessens, T.; De Koker, S.; Zagato, E.; Demeester, J.; Grooten, J.; De Smedt, S.C.; Raemdonck, K. Hybrid pulmonary surfactant-coated nanogels mediate efficient in vivo delivery of siRNA to murine alveolar macrophages. J. Control. Release 2015, 217, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Bielski, E.; Zhong, Q.; Mirza, H.; Brown, M.; Molla, A.; Carvajal, T.; da Rocha, S.R.P. TPP-dendrimer nanocarriers for siRNA delivery to the pulmonary epithelium and their dry powder and metered-dose inhaler formulations. Int. J. Pharm. 2017, 527, 171–183. [Google Scholar] [CrossRef]
- Bardoliwala, D.; Patel, V.; Javia, A.; Ghosh, S.; Patel, A.; Misra, A. Nanocarriers in effective pulmonary delivery of siRNA: Current approaches and challenges. Ther. Deliv. 2019, 10, 311–332. [Google Scholar] [CrossRef]
- Bade, B.C.; Dela Cruz, C.S. Lung Cancer 2020: Epidemiology, Etiology, and Prevention. Clin. Chest. Med. 2020, 41, 1–24. [Google Scholar] [CrossRef]
- Schaefer, I.M.; Padera, R.F.; Solomon, I.H.; Kanjilal, S.; Hammer, M.M.; Hornick, J.L.; Sholl, L.M. In situ detection of SARS-CoV-2 in lungs and airways of patients with COVID-19. Mod. Pathol. 2020, 33, 2104–2114. [Google Scholar] [CrossRef]
- Lam, J.K.; Liang, W.; Chan, H.K. Pulmonary delivery of therapeutic siRNA. Adv. Drug Deliv. Rev. 2012, 64, 1–15. [Google Scholar] [CrossRef]
- Knowles, M.R.; Boucher, R.C. Mucus clearance as a primary innate defense mechanism for mammalian airways. J. Clin. Investig. 2002, 109, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Roy, I.; Vij, N. Nanodelivery in airway diseases: Challenges and therapeutic applications. Nanomedicine 2010, 6, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, A.T.; Veerati, P.C.; Gosens, R.; Bartlett, N.W.; Wark, P.A.; Grainge, C.L.; Stick, S.M.; Kicic, A.; Moheimani, F.; Hansbro, P.M.; et al. Persistent induction of goblet cell differentiation in the airways: Therapeutic approaches. Pharmacol. Ther. 2018, 185, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Kubczak, M.; Michlewska, S.; Bryszewska, M.; Aigner, A.; Ionov, M. Nanoparticles for local delivery of siRNA in lung therapy. Adv. Drug Deliv. Rev. 2021, 179, 114038. [Google Scholar] [CrossRef]
- Gencer, A.; Duraloglu, C.; Ozbay, S.; Ciftci, T.T.; Yabanoglu-Ciftci, S.; Arica, B. Recent Advances in Treatment of Lung Cancer: Nanoparticle-based Drug and siRNA Delivery Systems. Curr. Drug Deliv. 2021, 18, 103–120. [Google Scholar] [CrossRef]
- Gu, Y.; Zhang, R.; Jiang, B.; Xu, X.; Guan, J.J.; Jiang, X.J.; Zhou, Y.; Zhou, Y.L.; Chen, X. Repair of Spinal Cord Injury by Inhibition of PLK4 Expression Through Local Delivery of siRNA-Loaded Nanoparticles. J. Mol. Neurosci. 2021, 9, 1–11. [Google Scholar] [CrossRef]
- Ahmed, Z.; Kalinski, H.; Berry, M.; Almasieh, M.; Ashush, H.; Slager, N.; Brafman, A.; Spivak, I.; Prasad, N.; Mett, I.; et al. Ocular neuroprotection by siRNA targeting caspase-2. Cell Death Dis. 2011, 2, e173. [Google Scholar] [CrossRef] [Green Version]
- Longest, P.W.; McLeskey, J.T., Jr.; Hindle, M. Characterization of Nanoaerosol Size Change During Enhanced Condensational Growth. Aerosol Sci. Technol. 2010, 44, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Worth Longest, P.; Hindle, M. Evaluation of the Respimat Soft Mist Inhaler using a concurrent CFD and in vitro approach. J. Aerosol Med. Pulm. Drug Deliv. 2009, 22, 99–112. [Google Scholar] [CrossRef]
- Dolovich, M.B.; Dhand, R. Aerosol drug delivery: Developments in device design and clinical use. Lancet 2011, 377, 1032–1045. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, Y.; Ma, C.; Ma, X.; Zhang, X.; Lin, L.; Zhao, Z.; Pan, X.; Wu, C. Endotracheal Aerosolization Device for Laboratory Investigation of Pulmonary Delivery of Nanoparticle Suspensions: In Vitro and in Vivo Validation. Mol. Pharm. 2018, 15, 5521–5533. [Google Scholar] [CrossRef] [PubMed]
- Geller, D.E. Comparing clinical features of the nebulizer, metered-dose inhaler, and dry powder inhaler. Respir. Care 2005, 50, 1313–1321; discussion 1321–1322. [Google Scholar] [PubMed]
- Rangaraj, N.; Pailla, S.R.; Sampathi, S. Insight into pulmonary drug delivery: Mechanism of drug deposition to device characterization and regulatory requirements. Pulm. Pharmacol. Ther. 2019, 54, 1–21. [Google Scholar] [CrossRef]
- Taki, M.; Marriott, C.; Zeng, X.M.; Martin, G.P. Aerodynamic deposition of combination dry powder inhaler formulations in vitro: A comparison of three impactors. Int. J. Pharm. 2010, 388, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Marple Virgil, A.O.B.A. Sampling and Measurement Using Inertial, Gravitational, Centrifugal, and Thermal Techniques. Aerosol Meas. 2011, 15, 129–151. [Google Scholar]
- Loo, B.; Jaklevic, J. Fine Particles: Aerosol Generation, Measurement, Sampling, and Analysis; Academic Press: New York, NY, USA, 1976. [Google Scholar]
- Dechraksa, J.; Suwandecha, T.; Maliwan, K.; Srichana, T. The comparison of fluid dynamics parameters in an Andersen cascade impactor equipped with and without a preseparator. AAPS PharmSciTech 2014, 15, 792–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marple, V.A.; Roberts, D.L.; Romay, F.J.; Miller, N.C.; Truman, K.G.; Van Oort, M.; Olsson, B.; Holroyd, M.J.; Mitchell, J.P.; Hochrainer, D. Next generation pharmaceutical impactor (a new impactor for pharmaceutical inhaler testing). Part I: Design. J. Aerosol Med. 2003, 16, 283–299. [Google Scholar] [CrossRef] [Green Version]
- Nichols, S.C.; Mitchell, J.P.; Shelton, C.M.; Roberts, D.L. Good Cascade Impactor Practice (GCIP) and considerations for “in-use” specifications. AAPS PharmSciTech 2013, 14, 375–390. [Google Scholar] [CrossRef] [Green Version]
- Hallworth, G.W.; Westmoreland, D.G. The twin impinger: A simple device for assessing the delivery of drugs from metered dose pressurized aerosol inhalers. J. Pharm. Pharmacol. 1987, 39, 966–972. [Google Scholar] [CrossRef]
- Mohan, M.; Lee, S.; Guo, C.; Peri, S.P.; Doub, W.H. Evaluation of Abbreviated Impactor Measurements (AIM) and Efficient Data Analysis (EDA) for Dry Powder Inhalers (DPIs) Against the Full-Resolution Next Generation Impactor (NGI). AAPS PharmSciTech 2017, 18, 1585–1594. [Google Scholar] [CrossRef]
- Youngren-Ortiz, S.R.; Gandhi, N.S.; Espana-Serrano, L.; Chougule, M.B. Aerosol Delivery of siRNA to the Lungs. Part 1: Rationale for Gene Delivery Systems. Kona 2016, 33, 63–85. [Google Scholar] [CrossRef] [Green Version]
- Cidem, A.; Bradbury, P.; Traini, D.; Ong, H.X. Modifying and Integrating in vitro and ex vivo Respiratory Models for Inhalation Drug Screening. Front. Bioeng. Biotechnol. 2020, 8, 581995. [Google Scholar] [CrossRef] [PubMed]
- Hsu, A.C.; Starkey, M.R.; Hanish, I.; Parsons, K.; Haw, T.J.; Howland, L.J.; Barr, I.; Mahony, J.B.; Foster, P.S.; Knight, D.A.; et al. Targeting PI3K-p110alpha Suppresses Influenza Virus Infection in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit Care Med. 2015, 191, 1012–1023. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.X.; Traini, D.; Young, P.M. Pharmaceutical applications of the Calu-3 lung epithelia cell line. Expert Opin. Drug Deliv. 2013, 10, 1287–1302. [Google Scholar] [CrossRef]
- Wark, P.; Hsu, A.; Starkey, M.; Hansbro, P. Micro-RNA-125a/b target A20 and MAVS to promote inflammatory and impair antiviral responses in chronic obstructive pulmonary disease. Eur. Respir. J. 2016, 48 (Suppl. 60), PA4658. [Google Scholar]
- Kedzierski, L.; Tate, M.D.; Hsu, A.C.; Kolesnik, T.B.; Linossi, E.M.; Dagley, L.; Dong, Z.; Freeman, S.; Infusini, G.; Starkey, M.R.; et al. Suppressor of cytokine signaling (SOCS)5 ameliorates influenza infection via inhibition of EGFR signaling. Elife 2017, 6, e20444. [Google Scholar] [CrossRef] [PubMed]
- Hayman, T.J.; Hsu, A.C.; Kolesnik, T.B.; Dagley, L.F.; Willemsen, J.; Tate, M.D.; Baker, P.J.; Kershaw, N.J.; Kedzierski, L.; Webb, A.I.; et al. RIPLET, and not TRIM25, is required for endogenous RIG-I-dependent antiviral responses. Immunol. Cell Biol. 2019, 97, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, Z.; Bradbury, P.; Pozzoli, M.; Young, P.M.; Ong, H.X.; Traini, D. An in vitro model for assessing drug transport in cystic fibrosis treatment: Characterisation of the CuFi-1 cell line. Eur. J. Pharm. Biopharm. 2020, 156, 121–130. [Google Scholar] [CrossRef]
- Fallacara, A.; Busato, L.; Pozzoli, M.; Ghadiri, M.; Ong, H.X.; Young, P.M.; Manfredini, S.; Traini, D. Combination of urea-crosslinked hyaluronic acid and sodium ascorbyl phosphate for the treatment of inflammatory lung diseases: An in vitro study. Eur. J. Pharm. Sci. 2018, 120, 96–106. [Google Scholar] [CrossRef]
- Haghi, M.; Traini, D.; Young, P. In vitro cell integrated impactor deposition methodology for the study of aerodynamically relevant size fractions from commercial pressurised metered dose inhalers. Pharm. Res. 2014, 31, 1779–1787. [Google Scholar] [CrossRef]
- Ong, H.X.; Traini, D.; Loo, C.Y.; Sarkissian, L.; Lauretani, G.; Scalia, S.; Young, P.M. Is the cellular uptake of respiratory aerosols delivered from different devices equivalent? Eur. J. Pharm. Biopharm. 2015, 93, 320–327. [Google Scholar] [CrossRef] [PubMed]
- van Rensburg, L.; van Zyl, J.M.; Smith, J. Deposition and transport of linezolid mediated by a synthetic surfactant Synsurf((R)) within a pressurized metered dose inhaler: A Calu-3 model. Drug Des. Devel. Ther. 2018, 12, 1107–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Bariwal, J.; Narang, A.S.; Tso, J.; Cheong, J.; Mahato, R.I. Functional similarity of modified cascade impactor to deposit drug particles on cells. Int. J. Pharm. 2020, 583, 119404. [Google Scholar] [CrossRef] [PubMed]
- Johansen, M.D.; Irving, A.; Montagutelli, X.; Tate, M.D.; Rudloff, I.; Nold, M.F.; Hansbro, N.G.; Kim, R.Y.; Donovan, C.; Liu, G.; et al. Animal and translational models of SARS-CoV-2 infection and COVID-19. Mucosal Immunol. 2020, 13, 877–891. [Google Scholar] [CrossRef] [PubMed]
- Enkirch, T.; von Messling, V. Ferret models of viral pathogenesis. Virology 2015, 479–480, 259–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. U.S. Product Development under the Animal Rule Guidance for Industry; U.S. Department of Health and Human Services; Food and Drug Administration; Center for Drug Evaluation and Research; Center for Biologics Evaluation and Research: New York, NY, USA, 2015.
- Heyder, J.; Gebhart, J.; Rudolf, G.; Schiller, C.F.; Stahlhofen, W. Deposition of particles in the human respiratory tract in the size range 0.005–15 μm. J. Aerosol Sci. 1986, 17, 811–825. [Google Scholar] [CrossRef]
- Sakagami, M. In vivo, in vitro and ex vivo models to assess pulmonary absorption and disposition of inhaled therapeutics for systemic delivery. Adv. Drug Deliv. Rev. 2006, 58, 1030–1060. [Google Scholar] [CrossRef]
- Cryan, S.A.; Sivadas, N.; Garcia-Contreras, L. In vivo animal models for drug delivery across the lung mucosal barrier. Adv. Drug Deliv. Rev. 2007, 59, 1133–1151. [Google Scholar] [CrossRef]
- Zafra, M.P.; Mazzeo, C.; Gamez, C.; Rodriguez Marco, A.; de Zulueta, A.; Sanz, V.; Bilbao, I.; Ruiz-Cabello, J.; Zubeldia, J.M.; del Pozo, V. Gene silencing of SOCS3 by siRNA intranasal delivery inhibits asthma phenotype in mice. PLoS ONE 2014, 9, e91996. [Google Scholar]
- Kim, R.Y.; Horvat, J.C.; Pinkerton, J.W.; Starkey, M.R.; Essilfie, A.T.; Mayall, J.R.; Nair, P.M.; Hansbro, N.G.; Jones, B.; Haw, T.J.; et al. MicroRNA-21 drives severe, steroid-insensitive experimental asthma by amplifying phosphoinositide 3-kinase-mediated suppression of histone deacetylase 2. J. Allergy Clin. Immunol. 2017, 139, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Morales, L.; Oliveros, J.C.; Fernandez-Delgado, R.; tenOever, B.R.; Enjuanes, L.; Sola, I. SARS-CoV-Encoded Small RNAs Contribute to Infection-Associated Lung Pathology. Cell Host Microbe 2017, 21, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.T.; Lu, C.Y.; Shao, P.L.; Chang, L.Y.; Wang, J.Y.; Chang, Y.H.; Lai, M.J.; Chi, Y.H.; Huang, L.M. In vivo inhibition of influenza A virus replication by RNA interference targeting the PB2 subunit via intratracheal delivery. PLoS ONE 2017, 12, e0174523. [Google Scholar] [CrossRef] [PubMed]
- Miwata, K.; Okamoto, H.; Nakashima, T.; Ihara, D.; Horimasu, Y.; Masuda, T.; Miyamoto, S.; Iwamoto, H.; Fujitaka, K.; Hamada, H.; et al. Intratracheal Administration of siRNA Dry Powder Targeting Vascular Endothelial Growth Factor Inhibits Lung Tumor Growth in Mice. Mol. Ther. Nucleic Acids 2018, 12, 698–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, B.; Cash-Mason, T.; Wang, Y.; Seitzer, J.; Burchard, J.; Brown, D.; Dudkin, V.; Davide, J.; Jadhav, V.; Sepp-Lorenzino, L.; et al. Intratracheal Administration of siRNA Triggers mRNA Silencing in the Lung to Modulate T Cell Immune Response and Lung Inflammation. Mol. Ther. Nucleic Acids 2019, 16, 194–205. [Google Scholar] [CrossRef] [Green Version]
- Dua, K.; Wadhwa, R.; Singhvi, G.; Rapalli, V.; Shukla, S.D.; Shastri, M.D.; Gupta, G.; Satija, S.; Mehta, M.; Khurana, N.; et al. The potential of siRNA based drug delivery in respiratory disorders: Recent advances and progress. Drug Dev. Res. 2019, 80, 714–730. [Google Scholar] [CrossRef]
- Hsu, A.C.; Dua, K.; Starkey, M.R.; Haw, T.J.; Nair, P.M.; Nichol, K.; Zammit, N.; Grey, S.T.; Baines, K.J.; Foster, P.S.; et al. MicroRNA-125a and -b inhibit A20 and MAVS to promote inflammation and impair antiviral response in COPD. JCI Insight 2017, 2, e90443. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Cooley, M.A.; Jarnicki, A.G.; Hsu, A.C.; Nair, P.M.; Haw, T.J.; Fricker, M.; Gellatly, S.L.; Kim, R.Y.; Inman, M.D.; et al. Fibulin-1 regulates the pathogenesis of tissue remodeling in respiratory diseases. JCI Insight 2016, 1, e86380. [Google Scholar] [CrossRef]
- Seth, S.; Templin, M.V.; Severson, G.; Baturevych, O. A potential therapeutic for pandemic influenza using RNA interference. Methods Mol. Biol. 2010, 623, 397–422. [Google Scholar]
- Li, T.; Koshy, S.; Folkesson, H.G. RNA interference for CFTR attenuates lung fluid absorption at birth in rats. Respir. Res. 2008, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Yang, J.; Liu, T.; Xuan, P.; Bu, B.; Xu, X.; Wu, R. Effect of Src tyrosine kinase on a rat model of asthma. Exp. Ther. Med. 2022, 23, 172. [Google Scholar] [CrossRef]
- Tang, Q.; Li, B.; Woodle, M.; Lu, P.Y. Application of siRNA against SARS in the rhesus macaque model. Methods Mol. Biol 2008, 442, 139–158. [Google Scholar] [PubMed]
- Geisbert, T.W.; Lee, A.C.; Robbins, M.; Geisbert, J.B.; Honko, A.N.; Sood, V.; Johnson, J.C.; de Jong, S.; Tavakoli, I.; Judge, A.; et al. Postexposure protection of non-human primates against a lethal Ebola virus challenge with RNA interference: A proof-of-concept study. Lancet 2010, 375, 1896–1905. [Google Scholar] [CrossRef]
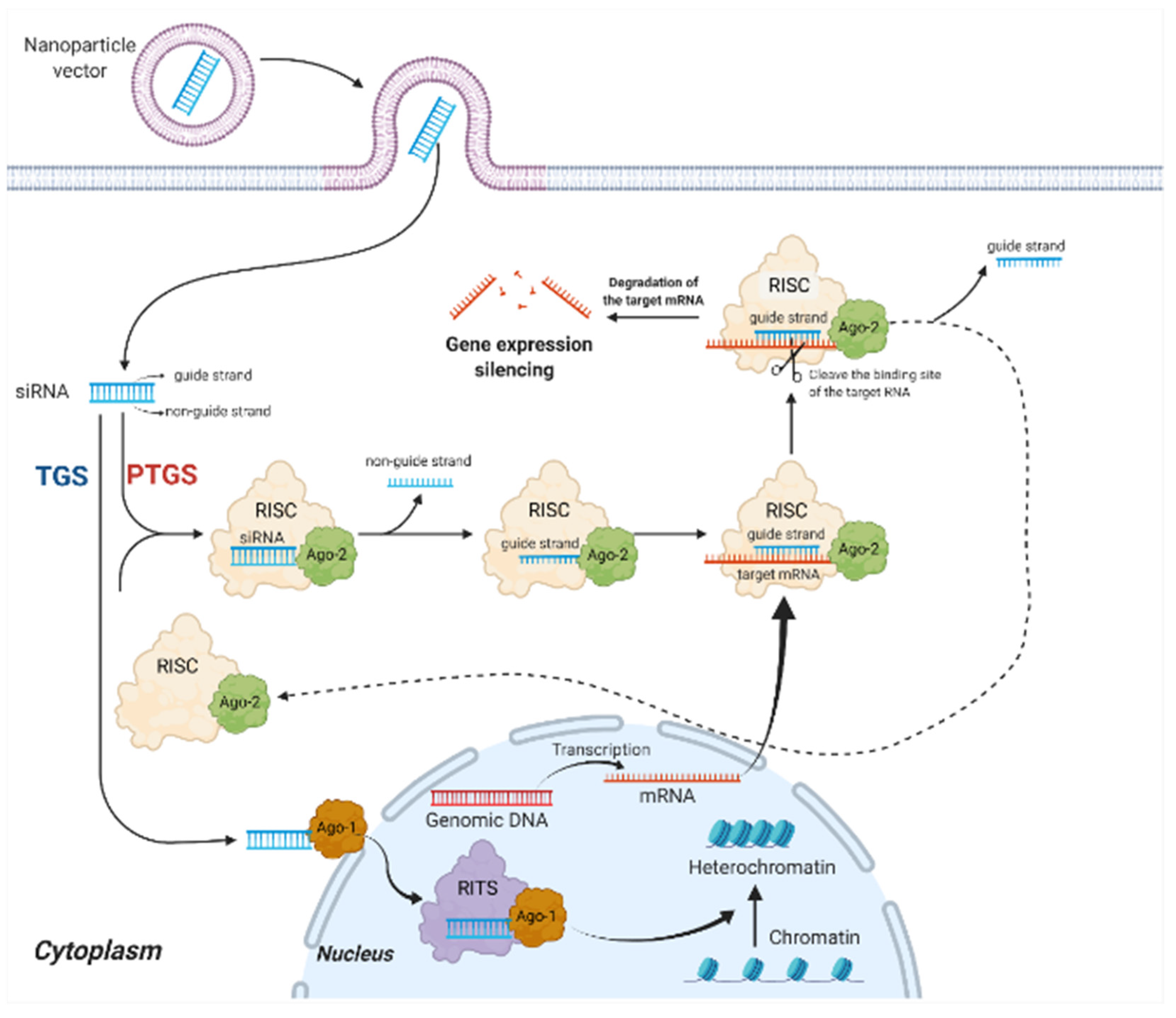
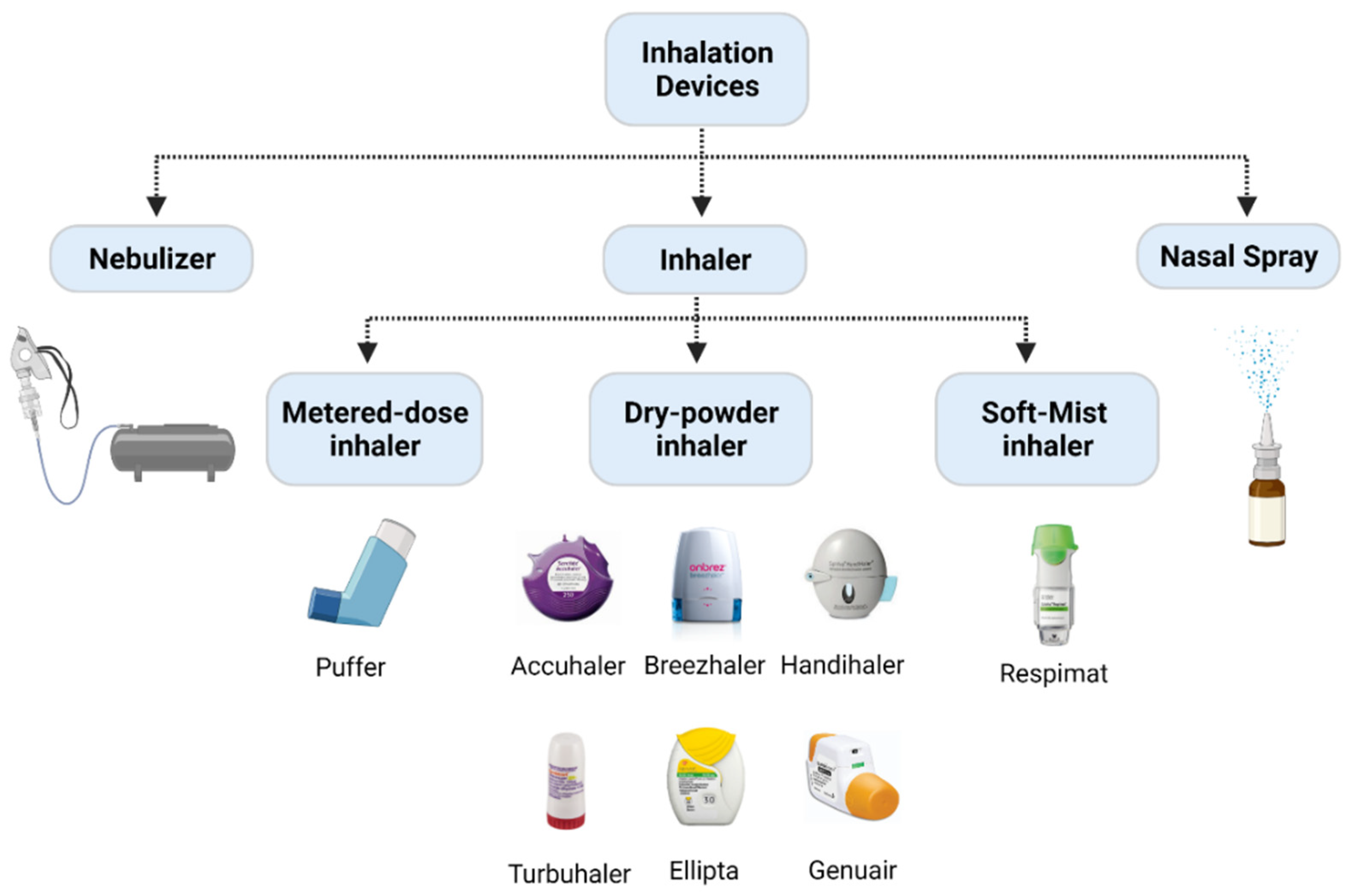

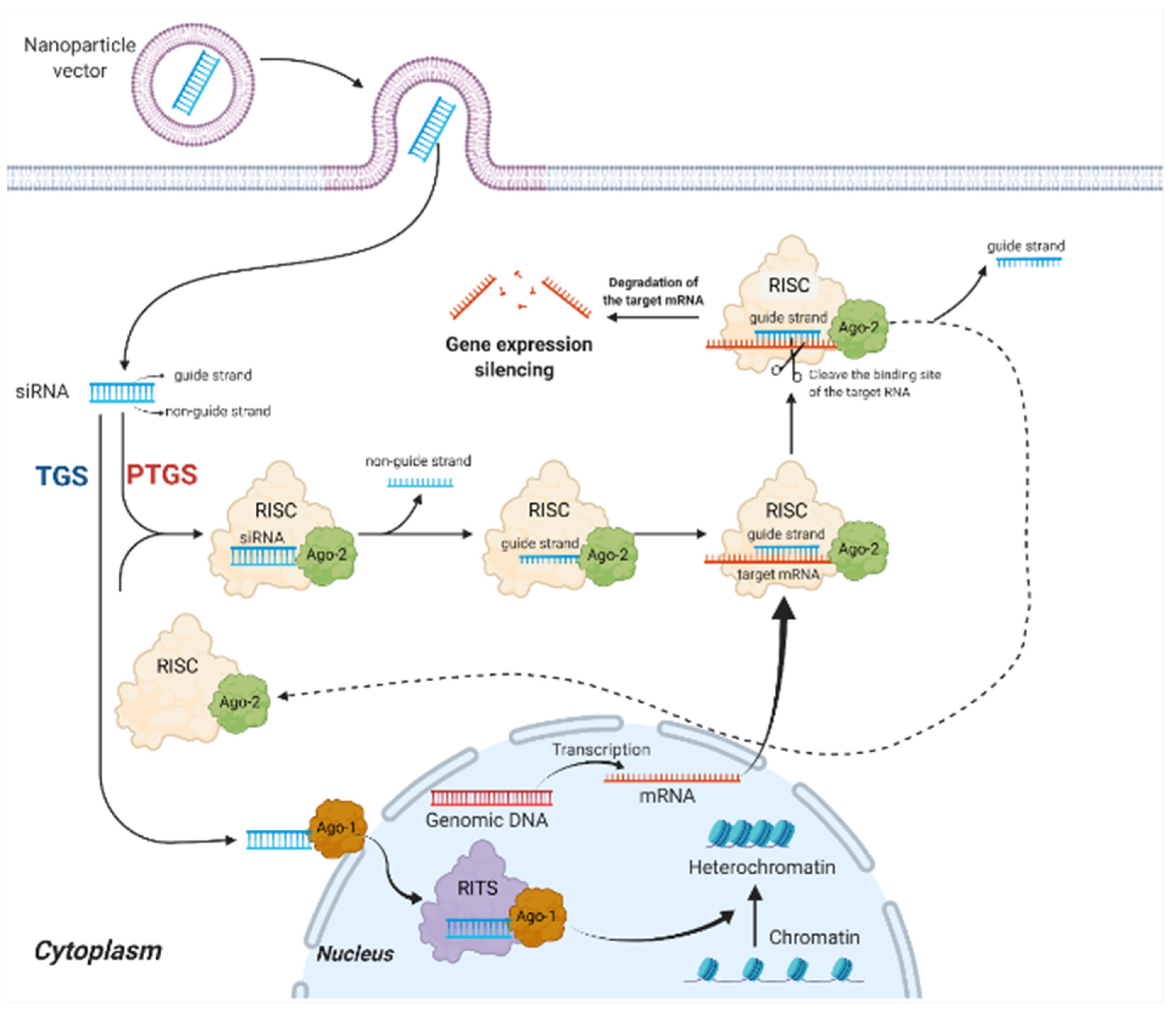{kind=link}
{kind=link}
{kind=link}
{kind=link}
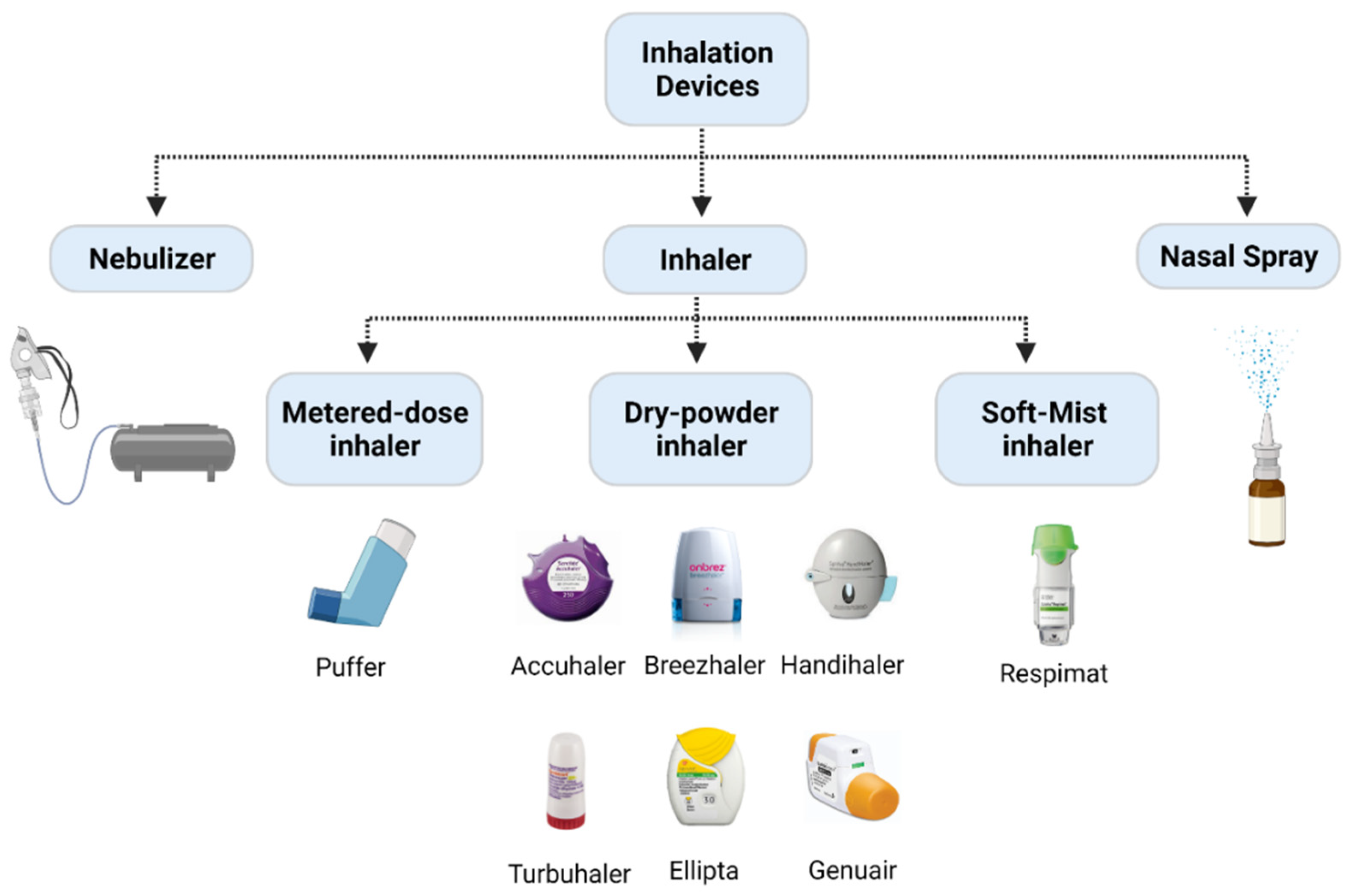{kind=link}
{kind=link}
| Virus | Sequence (5′-3′) | Region | Stage | Refs |
|---|---|---|---|---|
| MERS-CoV | UAGAAGAACAGCUAUCACCCU | ORF1ab | Computational approach | [99] |
| MERS-CoV | AACAUAGAAAGCAGAUAGGUC | ORF1ab | Computational approach | |
| MERS-CoV | UCUAAGAGCUGCAUAAGUGUC | ORF1ab | Computational approach | |
| MERS-CoV | AUCAAGAAAAGCGUUAGAGGA | ORF1ab | Computational approach | |
| MERS-CoV | UUUGUAGUACCAAUGACGCAA | ORF1ab | Computational approach | |
| MERS-CoV | UAAUAGUAAAAAUAGAUUGCU | ORF1ab | In vitro (HEK-293) | [108] |
| MERS-CoV | AACAUUAAUAGCAUUAUCCAU | ORF1ab | In vitro (HEK-293) | |
| MERS-CoV | UAAGAUAUCAUCUAAAGUGUC | ORF1ab | In vitro (HEK-293) | |
| MERS-CoV | UUAAAACUCAAACUAAUAGCA | ORF1ab | In vitro (HEK-293) | |
| MERS-CoV | UAGUUAAAGAGUUUCUAAGAG | ORF1ab | In vitro (HEK-293) | |
| MERS-CoV | UAAUAGUAAAAAUAGAUUGCU | ORF1ab | In vitro (Vero cells) | [109] |
| MERS-CoV | UAAGAUAUCAUCUAAAGUGUC | ORF1ab | In vitro (Vero cells) | |
| MERS-CoV | AACAUUAAUAGCAUUAUCCAU | ORF1ab | In vitro (Huh7) | [110] |
| MERS-CoV | UUAAAACUCAAACUAAUAGCA | ORF1ab | In vitro (Huh7) | |
| MERS-CoV | AUUAAAUCUGUUAAUGUUGUU | ORF1ab | In vitro (Huh7) | |
| MERS-CoV | AAAUAGUUAUGAAUAGUUGAG | ORF1ab | In vitro (Huh7) | |
| SARS-CoV-1 | GGGCUAUCAACCUAUAGAU | Spike protein (S) | In vitro (Vero E6) | [111] |
| SARS-CoV-1 | CAAGGCGAUUAGUCAAAUU | Spike protein (S) | In vitro (Vero E6) | |
| SARS-CoV-1 | CGUAACUAAACAGCACAAG | 3′-UTR | In vitro (Vero E6) | |
| SARS-CoV-1 | GCUCCUAAUUACACUCAAC | ORF2, spike | In vitro (FRhk-4) | [112] |
| SARS-CoV-1 | GGAUGAGGAAGGCAAUUUA | ORF1b, nsp-12 | In vitro (FRhk-4) | |
| SARS-CoV-1 | GGAUAAGUCAGCUCAAUGC | ORF1b, nsp-13 | In vitro (FRhk-4) | |
| SARS-CoV-1 | CUGGCACACUACUUGUCGA | ORF1b, nsp-16 | In vitro (FRhk-4) | |
| SARS-CoV-1 | GCUCCUAAUUACACUCAAC | ORF2, spike | In vivo (Rhesus macaque) | [113] |
| SARS-CoV-1 | GGAUGAGG AAGGCAAUUUA | ORF1b, nsp-12 | In vivo (Rhesus macaque) | |
| SARS-CoV-1 | GUACCCUCUUGAUUGCAUC | Replicase 1A | In vitro (FRhk-4) | [114] |
| SARS-CoV-1 | GAGUCGAAGAGAGGUGUCU | Replicase 1A | In vitro (FRhk-4) | |
| SARS-CoV-1 | GCACUUGUCUACCUUGAUG | Replicase 1A | In vitro (FRhk-4) | |
| SARS-CoV-1 | CACUGAUUCCGUUCGAGAUC | S | In vitro (FRhk-4) | [115] |
| SARS-CoV-1 | CGUUUCGGAAGAAACAGGUAC | E | In vitro (FRhk-4) | |
| SARS-CoV-1 | UGCUUGCUGCUGUCUACAG | M | In vitro (FRhk-4) | |
| SARS-CoV-1 | GUGGCUUAGCUACUUCGUUG | M | In vitro (FRhk-4) | |
| SARS-CoV-1 | UGAAGGAGUUCCUGAUCUUCU | Small envelope protein (E) | In vitro (Vero E6) | [116] |
| SARS-CoV-1 | AGCUUAAACAACUCCUGGAAC | Small envelope protein (E) | In vitro (Vero E6) | |
| SARS-CoV-1 | GAUAAUGGACCCCAAUCAAAC | Membrane protein (M) | In vitro (Vero E6) | |
| SARS-CoV-1 | CUUACAUAGCUCGCGUCUC | Nt 14450 to 14468 | In vitro (Vero cells) | [117] |
| SARS-CoV-1 | GAAUAUUAGGCGCAGGCUG | Nt 15877 to 15895 | In vitro (Vero cells) | |
| SARS-CoV-2 | UUCGUUUAGAGAACAGAUC | 5′-UTR | In vitro (Vero E6) | [34] |
| SARS-CoV-2 | GUACUUUUUUUGAACUUCUACA | Surface glycoprotein | In vitro (Vero cells) | [91] |
| SARS-CoV-2 | CAACAAAGAUAGCACUUAA | ORF1ab polyprotein | In vitro (Vero cells) | |
| SARS-CoV-2 | UCAUACCACUUAUGUACAA | ORF1ab polyprotein | In vitro (Vero cells) | |
| SARS-CoV-2 | CCAAAAUCAUAACCCUCAAA | ORF3a protein | In vitro (Vero cells) | |
| SARS-CoV-2 | AAACCUUCUUUUUACGUUUA | Envelope protein | In vitro (Vero cells) | |
| SARS-CoV-2 | CGAACGCUUUCUUAUUACAA | Membrane glycoprotein | In vitro (Vero cells) | |
| SARS-CoV-2 | AUGAACCACAAAUCAUUACUA | S | In vitro (human HUVECs and A549 cells) | [104] |
| SARS-CoV-2 | AGAUCAACUUACUCCUACUUG | S | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | AUAUAAUUCCGCAUCAUUUUC | S | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | AAGUCAAACAAAUUUACAAAA | S | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | AAUACACCAAAAGAUCACAUU | N | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | UAUUGACGCAUACAAAACAUU | N | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | AAGGAACUGAUUACAAACAUU | N | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | GUUCCAAUUAACACCAAUAGC | N | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | AACUAUAAAUUAAACACAGAC | M | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | AAACUAACAUUCUUCUCAACG | M | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | CUAUUCCUUACAUGGAUUUGU | M | In vitro (human HUVECs and A549 cells) | |
| SARS-CoV-2 | GUUGGACUGAGACUGACCUUA | RdRp | In vitro (Vero E6) and in vivo (K18-hACE2 mice) | [30] |
| SARS-CoV-2 | UUAUACCUUCCCAGGUAACAA | 5′UTR | In vitro (Vero E6) and in vivo (K18-hACE2 mice) | |
| SARS-CoV-2 | UCACCUUAUAAUUCACAGAAU | Helicase | In vitro (Vero E6) and in vivo (K18-hACE2 mice) | |
| SARS-CoV-2 | GGAAGGAAGUUCUGUUGAA | RdRp | In vitro (Vero cells) and in vivo (Syrian hamsters) | [118] |
| SARS-CoV-2 | UUUGUAUGCGUCAAUAUGCUU | Nucleocapsid phosphoprotein | Computational approach | [119] |
| SARS-CoV-2 | UCAACGUACACUUUGUUUCUG | Surface glycoprotein genes | Computational approach | |
| SARS-CoV-2 | AAAAACUUCACCAAAAGGGCA | Surface glycoprotein genes | Computational approach | |
| SARS-CoV-2 | UUAAAAACUUCACCAAAAGGG | Surface glycoprotein genes | Computational approach | |
| SARS-CoV-2 | UUAAAGCACGGUUUAAUUGUG | Surface glycoprotein genes | Computational approach | |
| SARS-CoV-2 | AACUUCUUGGGUGUUUUUGUC | Surface glycoprotein genes | Computational approach | |
| SARS-CoV-2 | UUUGAUUGUCCAAGUACACAC | Surface glycoprotein genes | Computational approach | |
| SARS-CoV-2 | UAAUUUGACUCCUUUGAGCAC | Surface glycoprotein genes | Computational approach | |
| SARS-CoV-2 | UCCUUCUUUAGAAACUAUACA | ORF1ab | Computational approach | [102] |
| SARS-CoV-2 | UGGUUUCACUACUUUCUGUUU | ORF1ab | Computational approach | |
| SARS-CoV-2 | UUCACUACUUUCUGUUUUGCU | ORF1ab | Computational approach | |
| SARS-CoV-2 | AUGUCAUCCCUACUAUAACUCAAA | ORF1ab | Computational approach | |
| SARS-CoV-2 | UUAAAAUAUAAUGAAAAUGGA | S | Computational approach | |
| SARS-CoV-2 | CUUGAAGCCCCUUUUCUCUAUCUUU | ORF3a | Computational approach | |
| SARS-CoV-2 | UUGAAUACACCAAAAGAUCACAUU | N | Computational approach | |
| SARS-CoV-2 | AGUAGAAAUACCAUCUUGGAC | N | Computational approach | [120] |
| SARS-CoV-2 | GUUUAGAGAACAGAUCUACAA | The leader sequence | Computational approach | [103] |
| SARS-CoV-2 | UAGUACUACAGAUAGAGACAC | RdRp | Molecular dynamics (MD) simulation | [101] |
| Carrier | siRNAs’ Target | Cells/Animal Models | Refs |
|---|---|---|---|
| Ionizable LNPs | LINC01257 siRNA | Kasumi-1 cells (human acute myeloid leukemia cell line) | [153] |
| Ionizable LNPs | Irf5 siRNA | C57BL/6 Irf5−/− Apoe−/− mice | [154] |
| Ionizable LNPs | MRTF-B siRNA | Human conjunctival fibroblasts | [155] |
| Ionizable LNPs | CTNNB1 Dicer substrate RNA (DsiRNA) | HepG2 and A375.S2 cells | [156] |
| Ionizable LNPs | RBPMS siRNA | HEK293-GFP stable cells (GFP293) | [157] |
| Ionizable LNPs (CL4H6) | STAT3 HIF-1α | ICR (♀, 4–10 weeks) mice and BALB/cAjcl-nu/nu (♂, 4–5 weeks) mice | [158] |
| Ultra-small LNPs | MK siRNA | Human HCC cell line, HepG2, and male BALB/c nude mice | [159] |
| Ginger-derived NPs (GDNPs) | Dmt1 siRNA | Hamp KO mice | [160] |
| SLNs | PD-1 | J774A.1 murine macrophages and B16-F10 murine melanoma cells | [161] |
| SLNs | TNF-α | J774A.1 murine macrophages | [162] |
| SLNs | Bcl-2 | HeLa (human cervical cancer adenocarcinoma) cell line | [163] |
| SLNs | BACE1 | Caco-2 cells (human epithelial colorectal adenocarcinoma cells) | [164] |
| SLNs | MDR1 | MCF-7 cells and MCF-7/ADR cells | [165] |
| SLNs | KSP | Murine endothelial cell line MS1-VEGF and human primary umbilical vein endothelial cell line (HUVEC); human dermal microvascular endothelial cell (HMEC-1) line | [166] |
| SLNs | Bcl-2 | A549 cells (human lung carcinoma) | [167] |
| SLNs | MCL1 | KB cells (human epithelial carcinoma) and BALB/c nu/nu mice | [168] |
| SLNs | c-Met | U-87MG human GBM cells and U-87MG tumor-bearing mouse | [169] |
| SLNs | VEGF | PC-3 (human prostate cancer cells) and MDAMB435 (human breast cancer cells) | [170] |
| Cationic SLNs | EphA2 siRNA | PC-3 and DU145 (prostate cancer cell lines) | [171] |
| NLCs | BCL2 and MRP1 | A549 human lung adenocarcinoma cells | [149] |
| NLNs | GFP | Chinese hamster ovary cells (CHO-GFP) and human embryonic kidney (HEK293) cells | [150] |
| Ethosomes | GAPDH | Human adult epidermal keratinocytes and female BALB/c mice | [151] |
| Echogenic Liposomes | CPP | Human breast adenocarcinoma cells (HT-1080) | [152] |
| Mouse Models | Carriers | Drugs | Delivery | Refs |
|---|---|---|---|---|
| Female BALB/c mice | Solid lipid nanoparticles (SLNs) | Rhynchophylline (Rhy) | Aerosol administration | [239] |
| Female BALB/c mice | Dextran nanoparticles | SET-M33 (a non-natural antimicrobial peptide) | Hamilton syringe and laryngoscope | [240] |
| Female BALB/c mice | Dextran nanogels | siRNA | Tracheal aspiration | [241] |
| Female BLAB/c mice | Nanocluster | Ceftazidime | Dry powder inhalation | [242] |
| Female CF-1 mice | PEGylated human ferritin nanocages (FTn) | Doxorubicin (DOX) | Inhalation | [243] |
| Female CF-1 mice | PLGA and PLGA–PEG NP | Dexamethasone sodium phosphate (DP) | Intranasal instillation | [244] |
| Female C57BL/6OlaHsd mice | PEG–PEI copolymer | Dexamethasone (DEX) | Aerosol administration | [245] |
| Male CF-1 mice | Mucus-penetrating particles (MPP) | Fluticasone propionate (FP) | Intratracheal instillation | [246] |
| Male CD-1 mice | Mesoporous silica nanoparticles (MSNs) | Nanopure water droplets | Aerosol nose-only exposure system | [247] |
| Impactors | Devices | Flow Rates | # of Stages | Particle Size Range |
|---|---|---|---|---|
| Andersen cascade impactor | pMDIs and DPIs | 28.3 L/min (60 and 90 L/min using modifications) | 6 or 8 | 0.4–0.9 microns (28.3 L/min), 0.3–8.6 (60 L/min), 0.2–8 (90 L/min) |
| Multi-stage liquid impinger | DPIs | 30–100 L/min | 4 | 1.7–13 microns |
| Twin-stage impinger | pMDIs and nebulizers | 60 L/min | 2 | 6.4 microns |
| Fast-screening impactor | pMDIs, DPIs, and nasal spray | 30–100 L/min | 2 | 5 and 10 microns |
| Next-generation pharmaceutical impactor | pMDIs, DPIs, and nebulizers | 15–100 L/min | 7 | 0.24–14.1 microns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Almazi, J.G.; Ong, H.X.; Johansen, M.D.; Ledger, S.; Traini, D.; Hansbro, P.M.; Kelleher, A.D.; Ahlenstiel, C.L. Nanoparticle Delivery Platforms for RNAi Therapeutics Targeting COVID-19 Disease in the Respiratory Tract. Int. J. Mol. Sci. 2022, 23, 2408. https://doi.org/10.3390/ijms23052408
Zhang Y, Almazi JG, Ong HX, Johansen MD, Ledger S, Traini D, Hansbro PM, Kelleher AD, Ahlenstiel CL. Nanoparticle Delivery Platforms for RNAi Therapeutics Targeting COVID-19 Disease in the Respiratory Tract. International Journal of Molecular Sciences. 2022; 23(5):2408. https://doi.org/10.3390/ijms23052408
Chicago/Turabian StyleZhang, Yuan, Juhura G. Almazi, Hui Xin Ong, Matt D. Johansen, Scott Ledger, Daniela Traini, Philip M. Hansbro, Anthony D. Kelleher, and Chantelle L. Ahlenstiel. 2022. "Nanoparticle Delivery Platforms for RNAi Therapeutics Targeting COVID-19 Disease in the Respiratory Tract" International Journal of Molecular Sciences 23, no. 5: 2408. https://doi.org/10.3390/ijms23052408