
Skeletal Muscle Regeneration in Cardiotoxin-Induced Muscle Injury Models

School of Kinesiology, Shanghai University of Sport, Shanghai 200438, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(21), 13380; https://doi.org/10.3390/ijms232113380
Submission received: 6 September 2022
/
Revised: 27 October 2022
/
Accepted: 28 October 2022
/
Published: 2 November 2022
(This article belongs to the Section Molecular Biology)
Abstract
:Skeletal muscle injuries occur frequently in daily life and exercise. Understanding the mechanisms of regeneration is critical for accelerating the repair and regeneration of muscle. Therefore, this article reviews knowledge on the mechanisms of skeletal muscle regeneration after cardiotoxin-induced injury. The process of regeneration is similar in different mouse strains and is inhibited by aging, obesity, and diabetes. Exercise, microcurrent electrical neuromuscular stimulation, and mechanical loading improve regeneration. The mechanisms of regeneration are complex and strain-dependent, and changes in functional proteins involved in the processes of necrotic fiber debris clearance, M1 to M2 macrophage conversion, SC activation, myoblast proliferation, differentiation and fusion, and fibrosis and calcification influence the final outcome of the regenerative activity.
1. Introduction
Skeletal muscle, the main organ of systemic metabolism in the body, is composed of differentiated fibers and displays a strong ability to regenerate after injury. Skeletal muscle injuries occur frequently in daily life and exercise, and the capacity of regeneration is critical for the repair and functional maintenance of skeletal muscle. The regeneration of adult muscle is based on the activation of satellite cells (SCs), which are mononuclear progenitors of skeletal muscle and are located between the sarcolemma and basal lamina [1]. After injury, the regeneration of muscle occurs in three overlapping stages: in the first stage, inflammatory cells infiltrate into damaged sites, and necrotic fiber fragments are removed; in the second stage, SCs are activated and proliferate into myoblasts, thereafter differentiating and fusing to form new muscle cells and replace damaged fibers; the last stage involves the maturation of newly formed fibers and the remodeling of damaged muscle [2,3]. The processes of regeneration are highly coordinated, and the expression of genes involved in regeneration are spatially and temporally regulated [4,5]. Numerous studies have been conducted to investigate the molecular mechanisms underlying muscle regeneration. A comprehensive understanding of the events involved in muscle regeneration will facilitate the treatment of skeletal muscle diseases.
In order to achieve a better understanding of muscle regeneration following physiological injury, the innervation, tendons, vascularization, and SCs should not be injured in mouse models because they contribute to myogenesis following injury. Cardiotoxin (CTX), derived from Naja pallida, induces a transient and reproducible acute injury without affecting the vasculature or nerves, and then produces a consistent injury in the whole muscle followed by synchronized regeneration [6,7,8]. Its application also has the advantages of allowing molecular and biochemical analyses to be performed on the whole muscle in contrast to physiological injury models induced by exercise [9,10]. Additionally, CTX injury models have relatively low harmfulness for animals compared with other non-physiological models such as crushing models [11]. Due to these characteristics, the CTX-induced skeletal muscle injury model is a suitable model for exploring the mechanisms of skeletal muscle regeneration.
In this review, we summarize the effects and mechanisms of different mouse models for obesity, diabetes, exercise training, and nutrition on regeneration after CTX-induced muscle injuries. The results may provide therapeutic targets for the repair of damaged muscle in addition to new ideas for further studies.
2. The Characteristics and Positions of Injury in CTX-Induced Skeletal Muscle Injury Models
CTX, a natural amphiphilic peptide derived from Naja pallida, can affect membrane calcium binding sites, and lower the threshold of calcium-modulated calcium ion release from the sarcoplasmic reticulum, thereafter inducing the destruction of skeletal muscle [12,13]. Muscle injury occurs at days 1 to 2 after CTX injection, where inflammatory cells infiltrate and SCs are activated to proliferate; at days 3–5, the myoblasts are induced to differentiate; at days 5–7, the new fibers with a central nucleus begin to form; and at days 10–14, the major muscle structures are restored; at day 28, the damaged muscles have almost completely recovered [14,15,16]. Due to its characteristics of transience and reproducibility, the CTX-induced injury model has been widely used to explore the mechanisms of skeletal muscle regeneration.
In CTX-induced injury models, the damaged sites are hindlimb muscles, where injuries also often occur in humans. In the related literature, the tibialis anterior is the most widely studied site in CTX-induced injury models. This is because of its obvious location and the characteristics of having a mixture of fiber types. Additionally, as a highly heterogenous muscle, tibialis anterior has only one belly, which results in uniform injury. Gastrocnemius consists mainly of fast-twitch fibers and is a bicep muscle, which may result in nonuniform injury despite the obvious location. Furthermore, other hindlimb muscles were also used in the studies such as the extensor digitorum longus, soleus, and quadriceps. The characteristics of only one type of muscle fiber and the muscle group may lead to a preference for position.
Notably, there are still some limitations in CTX-induced skeletal muscle injury models. First, the skeletal muscles include antigravity (e.g., gastrocnemius, quadriceps) and non-antigravity muscles (e.g., tibialis anterior, biceps brachii) [17]. The mechanisms identified in CTX-induced non-antigravity muscle injury models may not apply directly to the CTX-induced antigravity muscles. Second, in CTX-induced injury models, it always does not affect the vasculature or nerves in muscles [8]. In contrast, the vasculature or nerve damage often occurs during the pathogenesis of human muscle injuries [18]. This discrepancy limits the exploration of the contribution of vasculature or nerves in muscle regeneration using CTX-induced muscle injury models. Third, CTX may induce a complete necrosis of the small muscles such as EDL when examined in cross-section 48 h after injection [19]. This may make it impossible to explore the mechanisms involved in the early stages of these muscles.
3. Skeletal Muscle Regeneration in Different Mouse Models after CTX-Induced Skeletal Muscle Injury
CTX has been used to induce skeletal muscle injury in many mouse models (Table 1) including that of diabetes, obesity, aging, exercise training, mechanical loading, and nutrition intervention, among others. Studies have shown that streptozocin and gene mutation-induced diabetes [20,21,22,23], high fat diet-induced obesity and ob/ob mice [22,24,25], cancer cachexia [26], aging [27,28], irradiation [29], elevated carbon dioxide (CO2) level [30], and hindlimb suspension [31,32] lead to impaired regeneration, whereas exercise training [33,34], microcurrent electrical neuromuscular stimulation [35], microelement zinc [36], and overloading [37,38] improve the regeneration of CTX-induced damaged muscle. The accumulation of mitochondrial DNA alterations activates muscle regeneration in myofibers during aging, but leads to reduced muscle mass [39].
Gender and sex hormone levels also influence the regeneration processes in CTX-induced muscle injuries. Males exhibit larger newly formed fibers than females at the same age after injury, whereas females show higher fat deposition than males during regeneration [40,44] and also remove necrotic tissue more rapidly [44]. Castration of males increases the cross-sectional areas (CSAs) of the newly formed fibers and fat accumulation, whereas ovariectomized mice exhibit inhibited regeneration and decreased adipocyte accumulation, and estrogen supplementation rescues regeneration in ovariectomized mice [41,44]. Lack of estrogen-related receptor α also impairs the recovery of mitochondrial energetic capacity and decreases the activity of adenosine 5′-moophpsphate (AMP)–activated protein kinase (AMPK), which then also leads to delayed regeneration [52].
Additionally, the studies also revealed that different mouse strains have similar regeneration processes with no significant morphological and functional differences. However, the mechanisms of skeletal muscle regeneration may be strain-dependent. For instance, toll-like receptor 4 (TLR4) plays distinct roles in the injured muscle of C57BL/6 and C3H/HeJ [49,53].
It was also reported that the regeneration of skeletal muscle is position-specific: after CTX injury in tibialis anterior and the masseter, head muscles recover slowly and eventually return to the base level, whereas limb muscles show quicker recovery and eventually excessive growth [50].
4. Mechanisms of Regeneration in CTX-Induced Injury Models
It has reported that the trajectories of skeletal muscle regeneration vary considerably despite achieving complete regeneration in different injury models [54], wherein the mechanisms of regeneration in damaged skeletal muscle depend on the injury models [54]. In the following section, we summarize the mechanisms of regeneration based on CTX-induced skeletal muscle injury.
4.1. Inflammatory Response in CTX-Induced Injury Models
Inflammatory response could play an important role in timely skeletal muscle regeneration after CTX-induced injury. In this section, we summarize the mechanisms of this process and its three stages: immune cell infiltration, M1 to M2 macrophage polarization, and the clearance of necrotic fiber debris.
4.1.1. The Mechanisms of Inflammatory Response in CTX-Induced Skeletal Muscle Injury
Upon injury, immune cells residing in the skeletal muscle are rapidly activated and then release tissue destruction factors to accelerate muscle injury [55]. Additionally, the immune cells also secrete cytokines such as tumor necrosis factor alpha (TNFα) and interleukin 6 (IL-6), recruiting neutrophils into damaged areas, which in turn stimulates the secretion of chemokines including monocyte chemoattractant protein 1 (MCP-1), macrophage inflammatory protein 1 alpha (MIP-1α), MIP-1β, and promotes the invasion of circulating monocytes [56]. Studies have shown that the mechanisms of inflammatory response involved in CTX-induced skeletal muscle regeneration are complex (Figure 1, Table 2).
It is reported that the lack of interleukin (IL-1) [77], CC chemokine receptor 2 (CCR2) [81], toll-like receptor 2 (TLR2) [79], and heat shock protein (Hsp70) [78] and the inactivation of IL-6/signal transducer and activator of transcription 3 (STAT3) signaling [82] and complement C3a-C3a receptor (C3aR)/CCL5 signaling [86] lead to reduced/delayed monocyte/macrophage infiltration, which then reduces the clearance of necrotic fiber debris and impairs myoblast proliferation, attenuating/delaying muscle regeneration. The endogenous conversion of n-6 to n-3 polyunsaturated fatty acids [76] and pretreatment with thymol [75] reduce macrophage infiltration and cell apoptosis, leading to increased SC migration and proliferation and improved muscle regeneration. Loss of Kruppel-like factor 2 (KLF2) [65], and the lack of plasminogen activator inhibitor (PAI-1) [2,23] and signal transducer and activator of transcription 1 (STAT1) in bone marrow-derived cells [66] and the inhibition of activin A [69] stimulate monocyte/macrophage recruitment, accelerate damaged muscle degradation, and promote myoblast proliferation, thereby improving muscle regeneration. Additionally, the accumulation of interleukin 17A (IL-17A)-producing T cells can also promote muscle regeneration in a microbiota-dependent way [97]. Lack of neuraminidase-1 (Neu1) [58] and estrogen signaling [59] and increased activation of calcium/calmodulin-dependent protein kinase IV (CaMKIV) [57] increase the inflammatory response but inhibit muscle regeneration. This may be because the lack of Neu1 leads to delayed myoblast differentiation and myofiber maturation [58]; however, the activation of CaMKIV and the lack of estrogen signaling increase the infiltration of pro-inflammatory macrophages, impair the transition of macrophages from M1 to M2, reduce the phagocytosis of macrophages, resulting in impaired muscle regeneration [57].
In addition, muscle cells are also involved in the immune response. Studies have shown that the inflammatory environment induced by interferon gamma (IFN-γ) stimulates the expression of major histocompatibility complex (MHC) and some co-stimulatory molecules from regenerated myofibers or cultured myoblasts and myotubes, which then contribute to the immune response [98]. Myofibers also mediate the inflammatory response through the activation of transforming growth factor beta (TGF-β)/IL-6 signaling, and direct Th17 and Treg cell responses [99]. Moreover, oxidative stress during the inflammatory response can also change the structure and function of proteins, which then regulates muscle regeneration in CTX-induced injury [100].
4.1.2. The Mechanism of Macrophage Polarization in CTX-Induced Skeletal Muscle Injury
Macrophages are the main inflammatory cells, and macrophage polarization is involved in the regulation of regeneration [79]. Infiltrated monocytes differentiate into pro-inflammatory M1 macrophages, secreting proinflammatory cytokines, cleaning up necrotic fiber debris, and maintaining an inflammatory environment. Upon the removal of necrotic fiber debris, M1 macrophages switch to M2 macrophages, secreting anti-inflammatory factors and stimulating regeneration [101]. Research shows that (Figure 1, Table 2) a preexisting inflammatory environment [91], irradiation [29], transglutaminase 2 (TG2) deficiency [102], and the excessive activation of calmodulin-dependent signaling [64] delay or impair the M1 to M2 macrophage conversion, which then delays or impairs muscle regeneration. On the other hand, estrogen signaling [59], extracellular vesicles (EVs) derived from mesenchymal stem cells (MSCs) [73], peroxisome proliferator-activated receptor γ coactivator 1α (PGC-1α) [56], and scavenger receptor class B1 (SRB1) [90] stimulate macrophage polarization, eliminate necrotic fibers, reduce fibrosis, and then induce muscle regeneration. In addition, the lack of progranulin prolongs the existence of M2 macrophages and increases the size of newly formed fibers [72]. Increased activation of peroxisome proliferator-activated receptor beta (PPARβ) promotes the recruitment of M2 macrophages and accelerates the regeneration processes [70].
4.1.3. The Mechanism of Necrotic Fiber Debris Clearance in CTX-Induced Skeletal Muscle Injury
The phagocytotic ability of macrophages plays an important role in the elimination of necrotic fiber debris (Figure 1). The lack of retinol saturase (RetSat) in macrophages results in less milk fat globule-epidermal growth factor-factor-8 (MFG-E8) produced and impaired efferocytosis [103]. However, skeletal muscle regeneration in RetSat-null mice is normal following CTX-injury. This is because other cell types participating in muscle regeneration such as myoblasts compensate for the impaired macrophage function, which leads to normal muscle regeneration in RetSat-null mice [93]. Supplementation with balenine promotes the infiltration of immune cells into damaged muscle, and increases the phagocytotic ability of macrophages, leading to improved regeneration [74].
Additionally, the genes involved in the clearance of necrotic muscle fiber debris are highly expressed in immune cells (Table 2). Disintergrin and metalloprotease (ADAM) 8 expression in neutrophils reduces the expression of P-selectin glycoprotein ligand-1 (PSGL-1) on the surface of neutrophils, which then increases the ability of neutrophils to infiltrate into damaged areas and contribute to the removal of fiber debris [88]. Tyro3/Axl/Mer (TAM) kinase signaling mediated by Axl/Mer (AM) receptor Mer expressed in CD45+ cells and SRB1 expressed in macrophages could also facilitate the elimination of necrotic fibers and stimulate macrophage transition from M1 to M2 [89,90].
4.2. SC Activation and Myoblast Proliferation, Differentiation, and Fusion in CTX-Induced Injury Models
Upon injury, the regeneration capacity of the skeletal muscle is due to the SCs, and the critical steps such as SC activation and myoblast proliferation, differentiation, and fusion determine the extent of regeneration. Additionally, myotube maturation and the self-renewal of SCs also influence regeneration.
4.2.1. Mechanisms of SCs Activation and Myoblast Proliferation in CTX-Induced Injury Models
In normal adult muscle, SCs are in a quiescent state (Figure 2) and express paired box 7 (Pax7); once injury occurs, SCs begin to express myogenic differentiation 1 (MyoD) and myogenic factor 5 (Myf5) and are activated and then enter the cell cycle [104,105,106]. During this process, the basement membrane plays an important role in triggering the activation of SCs. After injury, the components of the basement membrane that mediate the contacts of the basal lamina to SCs and myofibers are degraded, and key components of the basement membrane (collagen IV alpha 1, laminin gamma-1, nidogen-2, and heparane sulfate proteoglycan-2) are downregulated, which further leads to a release of growth factors from the dismantling basement membrane and increased the elasticity of the SC niche, thus providing a suitable environment for SC proliferation [107]. As shown in Table 3, Xin and insulin-like 6 (Insl6) are involved in the activation of SCs. Insl6 overexpression in muscle facilitates SC activation and proliferation through the reduction in cell apoptosis upon CTX injury [108]. Xin, which increased in SCs within 12 h following the CTX-induced injury, maintains the activation of SCs, and the downregulation of endogenous Xin leads to the increased proliferation and migration of myoblasts [109]. Other research has shown that Xin deficiency reduces the activation and proliferation of SCs, and muscle regeneration is then impaired through the reduction in primary myoblasts and increased apoptosis of SCs [110,111]. Additionally, studies have also reported that the over-activation of myostatin/TGF-β receptor/pSmad3 signaling in diabetic mice [43] inhibits the activation of SCs. Nevertheless, the inhibition of TGF-β signaling by simultaneous knockout of TGF-β type I receptor (Tgfbr1) and activin receptor type 2B (Acvr1b) accelerates the myogenic process and improves skeletal muscle regeneration [112], whereas knockout of TGF-β receptor II (TGF-βr2) increases the inflammatory response by affecting T-cell function and the withdrawal at the later stage of muscle regeneration. The lack of nuclear factor (erythroid-derived) like 2 (Nrf2) [113] and lipocalin-2 (LCN2) [114] also inhibits the activation of SCs. These may be associated with the pro-oxidation state, reactive oxygen species (ROS) accumulation, and reduced matrix metalloproteinase-9 (MMP-9) expression, which then lead to delayed or impaired regeneration [113,114].
The capacity for myoblast proliferation is influenced by numerous cytokines. Pretreatment with acetylated myostatin 1 (Ac-MIF1) and acetylated and amidated myostatin 2 (Ac-MIF2-NH2) stimulates muscle regeneration by increasing the capacity of myoblasts for proliferation and differentiation [242]. Chemokines including MCP-1, MIP-1α, or MIP-1β (CCL4) induce extracellular regulated protein kinase (ERK) 1/2 phosphorylation through a Gαi subunit-dependent manner, which promotes myoblast proliferation [170]. Gαi2 also promotes myoblast differentiation through the protein kinase C (PKC)/glycogen synthase kinase 3β(GSK3β)/miR-1 pathway or the histone deacetylase (HDAC) inhibition [160]. Nitric oxide (NO) stimulates the proliferation of SCs via a cyclic guanosine 3′,5′-monophosphate (GMP)-dependent pathway [145]. Double homeobox gene (Duxbl) [151] and factor for adipocyte differentiation 24 (fad24) [172] promote myoblast proliferation via increasing the cell cycle. The increased expression of anti-oxidant superoxide dismutase 1 (Sod1) and catalase (Cat) genes also facilitates the potential of proliferation and differentiation [153]. Additionally, the lack of H19 [169], hippo inhibition [117], and lack of heme oxygenase-1 (Hmox1) [67], fibroblast growth factor 6 (FGF6) [157], p38α, and p38γ [148] have also been found to promote myoblast proliferation, although the mechanisms remain unknown. Tweak/Fn14 also contributes to myoblast proliferation but inhibits their differentiation and delays their regeneration [168]. In contrast, increased inflammation, cell cycle inhibition, the destruction of membrane integrity, and iron accumulation all lead to attenuated myoblast proliferation. The results show that the lack of mitogen-activated protein kinase phosphatase-1 (MKP-1) [63] and heat shock transcription factor 1 (HSF1) [128] increased the inflammation and secretion of proinflammatory cytokines, the lack of α7 integrin destroyed the sarcolemmal integrity [165], and impaired fibroblast growth factor (FGF) responsiveness induced by deficiency of Cdon or fibroblast growth factor receptor 1 (FGFR1) led to the impairment of myoblast proliferation [164,174]. Lack of early growth response3 (Egr3) and the overexpression of calcium/calmodulin-dependent protein kinase kinase 2 (CaMKK2) might induce cell cycle arrest by the inactivation of nuclear factor kappa-B (NF-κB) and the activation of AMPK/p-cdc2-Tyr15 signaling, respectively [159,171]. Peroxisome proliferator-activated receptor δ (PPARδ) deficiency reduced forkhead box protein O1 (FOXO1) expression, which then impaired proliferation; FOXO1 overexpression also induced the expression of cell cycle inhibitors p57 and Gadd45α, which decreased the capacity for myoblast proliferation [127,167]. Iron accumulation and ROS production induced by transferrin receptor 1 (Tfr1) deletion also led to defective myoblast proliferation via the Tfr1–Slc39a14–iron axis [162]. Additionally, the deletion of Notch1 and/or Notch2 [141] and lack of nuclear T3 receptor TRα1 (p43) [158] also inhibited myoblast proliferation. However, the lack of Nur77 did not impair muscle regeneration even though it inhibited myoblast proliferation [150].
MicroRNAs are important regulators in SC activation and myoblast proliferation. The overexpression of miR-378 attenuates the activation and differentiation of SCs in an insulin-like growth factor 1 receptor (IGF1R)-dependent manner, which then delays the regeneration [118]. The expression of miR-29a, induced by fibroblast growth factor 2 (FGF2), reduces the expression of basement membrane members, which results in dismantling of the basement membrane, further providing a suitable environment for myoblast proliferation [107].
Epigenetic regulation is also involved in muscle regeneration. DNA methyltransferases 3a (Dnmt3a) ablation in SCs leads to hypomethylation of p57Kip2, which further induces a higher expression of p57Kip2, and then impairs the SC proliferation and attenuates the regeneration of damaged skeletal muscle [163]. Mixed lineage leukemia protein-1 (MLL1) facilitates the proliferation of myoblasts and Pax7+ SCs by epigenetically increasing the expression of Myf5 by mediating the trimethylation of lysine 4 on the histone H3 protein subunit (H3K4me3) enriched on the Myf5 promoter [123].
4.2.2. Mechanisms of Self-Renewal of SC Pool in CTX-Induced Injury Models
During regeneration, the self-renewal of SCs is also essential for the repair of damaged muscle (Figure 2). The activated SCs downregulate MyoD expression, and then replenish the SC pool through both symmetric cell division and asymmetric cell division [104,143]. In this process, primary cilium, harbored on the surface of quiescent SCs, has been shown to be an intrinsic factor controlling the self-renewal of SCs. Upon SC activation, primary cilia disassemble, and SCs enter the cell cycle. Upon exit from the cell cycle, the primary cilia reassemble again at the surface of a minority of SCs that are committed to self-renewal [143]. Disruption of the cilia reassembly impairs the self-renewal of SCs [143]. Additionally, the lack of selenoprotein N (SelN) [140], angiotensin II/Ang II AT1 receptor (AT1R) [137], and thyroid hormone receptor alpha (TRa) deficiency [139] resulted in a reduced SC pool and impaired regeneration of damaged muscle. Mammalian target of rapamycin complex 2 (mTORC2) depletion does not affect the myogenic function of SCs but impairs the replenishment of the SC pool upon repeated CTX injury [136]. Lack of collagen VI reduces the self-renewal capacity of SCs and impairs muscle regeneration [146].
In contrast, using the CRISPR/Cas9 mutagenesis technique, Sincennes et al. abolished Pax7 acetylation in mice and demonstrated that the lack of Pax7 acetylation led to reduced numbers of asymmetric stem cell divisions, expansion of the SC pool, and increased numbers of oxidative II A myofibers [124]. PKCθ deficiency upregulates Pax7 expression and activates Notch signaling for the maintenance of the self-renewal capacity of SCs in CTX-injured mdx mice [121]. Retinoblastoma (Rb) ablation in SCs increases the cell cycle re-entry of quiescent SCs and promotes the expansion of SCs. However, sustained retinoblastoma 1 (Rb1) loss impairs muscle fiber formation [135]. In addition, Klotho rejuvenates aged SCs and maintains the function of SCs by inhibiting the Wnt signaling pathway [129].
4.2.3. Mechanisms of Myoblast Differentiation in CTX-Induced Injury Models
The mechanisms of myoblast differentiation in Figure 2 and Table 3 show that the upregulation of myogenic regulatory factors (MRFs) such as MyoD and myogenin is associated with the increased capacity of myoblast differentiation, which further contributes to skeletal muscle regeneration in CTX-induced injury models. Research suggests that A-kinase anchoring protein 6 (AKAP6) [195], andrographolide [218], mouse double minute 2 homolog (Mdm2)/CCAAT/enhancer-binding protein β (C/EBPβ) [214], apolipoprotein B mRNA editing enzyme catalytic polypeptide 2 (APOBEC2) knockout [8] induces the expression of MyoD, myogenin, MyoG, and desmin. Leucine-rich repeats and transmembrane domains 1 (LRTM1) inhibit the recruitment of p52Shc to FGFR1 and inhibit the activation of ERK, further reducing the inhibition of cyclin dependent kinase 4 (CDK4) on the transcriptional activity of MyoD [196]. The increased MyoD interacts with its targets transcription elongation factor A-like 7 (Tceal7) and R3h domain containing-like (R3hdml) and promotes myoblast differentiation [1]. Additionally, cyclin D-type binding-protein 1 (Ccndbp1) can bind to MyoD and regulate muscle differentiation [3,186]. The energy metabolism in cells also influences myoblast differentiation. Micropeptide in mitochondria (MPM) increases oxygen consumption and adenosine triphosphate (ATP) synthesis and promotes myoblast differentiation [197]. The expression of the type 1 canonical subfamily of transient receptor potential channels (Trpc1) promotes the influx of calcium in myoblasts during differentiation and activates the phosphatidylinositol-3-kinase (PI3K)/AKT/mTOR/p70S6K pathway [173]. The decrease in chondroitin sulfate (CS) also stimulates the activation of PI3K/AKT signaling [181], which leads to faster regeneration [243]. Additionally, trimetazidine modulates the metabolic shift from free fatty acid β-oxidation to glucose oxidation by stimulating AMPK/PGC1α, and inducing autophagy, both of which contribute to myoblast differentiation [179]. Furthermore, the lack of signal transducer and activator of transcription 6 (STAT6) increases myoblast differentiation in an IL-4-independent way [219]. Angiotensin type 2 receptor (AT2R) inhibits the activation of ERK1/2 signaling to promote myoblast differentiation and fusion [185]. Inositol requiring enzyme 1 (IRE1) suppresses the expression of myostatin through its RNase-dependent RIDD activity, which then promotes the differentiation of myoblasts [188].
In contrast, the increased oxidation state impairs myoblast differentiation. Nonalcoholic fatty liver disease (NAFLD) reduces the SC pool and impairs SC differentiation, leading to attenuated skeletal muscle regeneration. This may be associated with TNF-α upregulation and increased levels of oxidative stress marker nicotinamide adenine dinucleotide phosphate (NADPH) oxidase-2 (NOX 2) [47]. The inhibition of carbonyl reductase1 (CBR1) leads to increased ROS levels and diminishes myoblast differentiation [216]. Iron overload impairs myoblast differentiation through oxidative stress-induced inactivation of the mitogen-activated protein kinase (MAPK) signaling pathway [45]. Lack of Hsp70 [16] and TNF-α receptors p55 and p75 [187,209] also downregulate p38MAPK activation, which further impairs myoblast differentiation. Moreover, decreased expression of MRFs such as MyoD and myogenin also impairs differentiation. Overexpression of ladybird homeobox 1 (Lbx1) [213] and the tripartite motif domain of myospryn [203] inhibit the expression of MyoD and myogenin. High levels of cardiotrophin-1 (CT-1) repress the expression of the MRFs such as MyoD through the activation of mitogen-activated protein kinase kinase (MEK)-MAPK signaling [199]. Protein-activated kinase 1 (PAK1) inhibitor IPA-3 decreases the expression of myogenin and reduces p38 phosphorylation [193]. Teashirt-3 (Tshz3) cooperates with BRG1-associated factor 57 (BAF57) and inhibits the MYOD-dependent activation of Myog [201]. Additionally, HS 6-O-endosulfatases (Sulfs) mutation and lack of tensin lead to reduced withdrawal from the cell cycle and delayed myoblast differentiation [191,211]. The deletion of RNA binding motif protein 24 (Rbm24) also regulates the alternative splicing of myogenic associated genes such as myocyte enhancer factor 2d (Mef2d), Rho-associated protein kinase 2 (Rock2), further inhibiting myoblast differentiation [106].
MicroRNAs are involved in the regulation of myoblast differentiation. The overexpression of miR-351 protects differentiating myoblasts from apoptosis by regulating the target gene E2f3 and contributes to myoblast differentiation [152]. In a normoxic state, miR-210 induces myoblast differentiation in a hypoxia-inducible factor 1-α (Hif1a)-dependent manner [239]. Overexpression of Linc-smad7 increases the expression of smad7 and insulin-like growth factor 2 (IGF2), which then induces myoblast differentiation [178]. In addition, miR-431 directly interacts with the 3′ untranslated region of Smad4. Ectopic miR-431 injection greatly reduces Smad4 levels and improves muscle regeneration in CTX-induced skeletal muscle injury models, whereas the inhibition of miR-431 significantly represses myoblast differentiation [192]. The inhibition of miR-188 reduces the expression of myogenic regulator factor 4 (MRF4) and Mef2c and impairs myoblast differentiation, whereas the overexpression of miR-188 has the opposite effect [155]. Knockdown of transactivating response RNA-binding protein (Trbp) downregulates the expression of miR-1a and miR-133a and reduces myotube formation [244]. Intriguingly, MyoR, a muscle-restricted basic helix–loop–helix transcription factor that antagonizes the actions of MyoD, is found to be anticorrelated with miR-378 during CTX-induced muscle regeneration. MyoD binds to the miR-378 gene and causes both transactivation and chromatin remodeling, thus upregulating miR-378 during myogenic differentiation. The 3′ untranslated region of MyoR contains a direct binding site for miR-378. The presence of this binding site significantly reduces the ability of MyoR and prevents the MyoD-driven transdifferentiation of fibroblasts [180].
Epigenetic regulation is also involved in myoblast differentiation. Histone- and protein arginine methyl transferases 5 (PRMT5)-associated protein COPR5 is required for cell cycle exit and myoblast differentiation. The silencing of COPR5 reduces PRMT5 recruitment to the promoters of p21 and MYOG by hindering interaction with the Runt-related transcription factor 1 (RUNX1)-core binding factor-β (CBFβ), which then inhibits the expression of p21 and MYOG and further impairs myoblast differentiation [200]. IGF-1 induces the phosphorylation and activation of ATP citrate lyase (ACL) through the PI3K/AKT pathway. The activated ACL catalyzes the conversion of citrate into oxaloacetate and acetyl-CoA, and acetyl-CoA can be further utilized by histone acetylases to acetylate H3 (K9/14) and H3 (K27) at the MyoD locus to increase MyoD expression, thereby promoting myoblast differentiation [184].
4.2.4. Mechanisms of Myoblast Fusion in CTX-Induced Injury Models
Differentiated myoblasts fuse with damaged fibers or new myotubes by cell–cell recognition, adhesion, migration, and membrane fusion, subsequently forming multinucleated myotubes [106,230]. This is a dynamic and coordinated process involving many proteins (Figure 2, Table 3).
In terms of stimulating fusion, anoctamin 5 (ANO5) stimulates the repair of the sarcolemmal membrane and facilitates myoblast fusion [224]. Stabilin-2 activates the G-protein coupled receptor (GPCR) activity of BAI3 and then recruits Elmo to the membrane to stimulate myoblast fusion [226]. NADPH oxidase 4 (Nox4) induces the expression of myomarker fusion protein (Tmeme8c) via Nox4-mediated ROS production and then contributes to myoblast fusion [229]. The activation of phospholipase D1 (PLD1) on the plasma membrane facilitates mononucleated myoblast fusion with nascent myotubes [230]. Inhibition of the hierarchical non-clustered miRNA network including highly active (miR-29a), moderately active (let-7), and mildly active (miR-125b, miR-199a, miR-221) networks, stimulates the activation of focal adhesion kinase and AKT and MAPK signaling, and leads to the formation of myotubes [189]. Transient receptor potential cation channel vanilloid I (TRPV I) can be activated by IL-4 and calcium signaling, which then facilitates myoblast fusion instead of proliferation [232]. Syncytin contributes to myoblast fusion, and this effect is male-specific [222].
The mechanism of inhibition of myoblast fusion involves the upregulation of TGF-β via calpain-3 (CAPN3) deficiency, thus leading to defective myoblast fusion [234]. C1q-like 1-4 interacts with BAI3 to repress myoblast fusion [226]. Lack of IGF-1 receptor (IGF-1R) signaling leads to reduced fiber fusion via growth hormone receptor-independent signaling [220]. The expression of (Pro)renin receptor ((P)RR) activates the Wnt/β-catenin and Yes-associated protein (YAP) signaling pathways, and decreases myoblast fusion [228]. In addition, transglutaminase 2 (TG2) [102], constitutive expression of c-Myb lacking its 3′ untranslated region (3′ UTR) [223], inhibition of 3-hydroxy 3-methylglutaryl coenzyme A reductase (HMGR) [225], myasthenia gravis [46], and the lack of ste20-like kinase (SLK) [221] also impair the capacity of myoblast fusion and decrease the fusion index.
4.2.5. Mechanisms of Myotube Maturation in CTX-Induced Injury Models
Fused multinucleated myotubes undergo terminal differentiation and eventually become mature myofibers. Kruppel-like factor (Klf5) (Table 3) is shown to interact with MyoD and Mef2 to regulate terminal differentiation [3]. Doublecortin (Dcx) facilitates myofiber maturation [235]. Sema4C stimulates the phosphorylation of p38 and activates the p38/MAPK signaling pathway to promote terminal differentiation [176]. The expression of clathrin heavy chain like 1 (CHC22) in CTX-induced injury muscle, however, diminishes glucose transporter 4 (Glut4) response and further impairs fiber maturation [237].
4.3. Fibrosis in CTX-Induced Injury Models
Fibrosis is an important stage for regeneration (Figure 3). In this process, the temporary extracellular matrix (ECM) components serve as a scaffold for new fibers and stabilize muscle tissue [89]. In damaged skeletal muscle, fibro/adipogenic progenitors (FAPs) are considered as the main source of fibroblasts [245]. After injury, FAPs are activated and begin to proliferate. This increases FAPs in the necrotic area, which need to be removed in time. Failure to clear FAPs will result in their differentiation into fibroblasts and adipocytes [246]. Fibroblasts secrete extracellular matrix proteins and growth factors and then differentiate into myofibroblasts to increase α-smooth muscle actin (α-SMA) expression and ECM synthesis, finally resulting in fibrosis [247]. Studies on the regulation of FAPs show that (Table 4) IL-4 secreted by infiltrated eosinophils stimulates the activation of FAPs in an IL-4-dependent way. IL-4/IL-13 signaling in FAPs contributes to proliferation and adipogenic differentiation of FAPs is inhibited to facilitate regeneration [55]. IL-1α and IL-1β inhibit the adipogenic differentiation of FAPs, and epidermal growth factor (EGF) and betacellulin (BTC) stimulate the proliferation of FAPs [248]. Lack of TGF-β1 in macrophages inhibits FAP proliferation and reduces fibrosis [249]. Inactivation of retinoic acid (RA) signaling in FAPs leads to adipogenic differentiation, which then impairs regeneration [246].
Additionally, studies have also shown that increases in miR-199a-5p [247], growth differentiation factor 11 (GDF11) [254], and platelet-derived growth factor receptor beta (PDGFRβ) [245], together with the lack of GDF-associated serum protein-1 (Gasp1) and/or Gasp2 [256] and a prior burst of double homeobox 4 (DUX4) [251], induced the deposition of collagen and contributed to fibrosis. In contrast, laminin-111 reduced fibrosis and facilitated skeletal muscle regeneration [165]. Losartam therapy also reduced fibrosis by inhibiting the TGF-β signaling pathway [250].
4.4. Calcification in CTX-Induced Injury Models
Calcification occurs after muscle injury. Under normal conditions, calcification can be resorbed. While in a pathological state, continuous calcification can induce chronic inflammation and/or loss of muscle function [264]. Studies have revealed that (Table 4) after CTX injection, Tie2-expressing endothelial precursors are the main contributor to calcification in a mouse model of dysregulated bone morphogenetic protein (BMP) signaling [258]. Moreover, the inflammatory microenvironment induced by CTX injection is also necessary for calcification in injured muscle [258]. At the early stage after injury, calcific nodules are present in mitochondria, which are mediated by cell death and can be cleared by infiltrated macrophages [257]. Additionally, calcification in damaged muscle may also occur in connection with reduced plasmin, and this is independent of its canonical fibrinolytic function [255]. The hypoxia state induced by CTX injury can induce osteogenic differentiation and mineralization of muscle resident stromal cells and further stimulate the formation of myofiber calcification [259].
4.5. Angiopoiesis and Neurogenesis in CTX-Induced Injury Models
In CTX-induced injury models, the capillaries are destroyed, and endothelial cells are activated to repair the skeletal muscle endothelium. It is reported that (Table 5) macrophage cells derived from bone marrow can express endothelium-related markers such as Tie2 and CD31 to promote angiogenesis [265]. Angiotensin II derived from differentiated muscle myoblasts stimulates the migration of endothelial cells, which also further facilitates angiopoiesis [266]. In contrast, CCR2 deficiency leads to decreased vascular endothelial growth factor (VEGF) production and delayed angiogenesis in injured muscle, which then impairs regeneration [81].
In terms of neurogenesis, M2 macrophages infiltrate damaged muscle, produce hepatocyte growth factor (HGF), and then stimulate the expression of semaphorin 3A (Sema 3A) in myoblasts to regulate the regeneration of motor innervation in injured muscle [270,271]. Pre-activation of satellite cells delays the maturation of the neuromuscular junction by reducing the expression of semaphoring (Sema) 3A and S100B [269]. Lack of desmin leads to disrupted neuromuscular connections [238].
4.6. Other Regeneration-Related Genes in CTX-Induced Injury Models
In addition to the mechanism described above, there are a large number of genes involved in skeletal muscle regeneration in CTX-induced injury models such as Tsukushi, Dicer, mesoderm specific transcript (Mest), filamin C, LYVE-1, and so on (Table 6). However, in these studies, the special role of these genes has not been explored. Further experiments are needed to elucidate their function.
4.7. Non-SC Stem Cells Regulate Regeneration in CTX-Induced Injury Muscle
Non-SC stem cells are also involved in the regulation of regeneration (Figure 4). The results (Table 7) show that bone marrow-derived cells [308,309], pulp cells [310], bone marrow-derived human MSCs [311], hematopoietic stem cells [312], muscle precursor cells [313,314], capillary stem cells [315], adipose-derived mesenchymal stem cells [316], and human amniotic fluid stem cells [317,318] settle in the injured sites and differentiate into myogenic cells to stimulate the skeletal muscle regeneration in CTX-induced injury models. Additionally, mobilization of bone marrow stem cells also accelerates the muscle regeneration [319].
5. Conclusions
Skeletal muscle has a tremendous capacity for regeneration after injury. This is largely due to muscle SCs. In order to learn about the mechanisms of regeneration, skeletal muscle regeneration has been studied for decades in numerous injury models. However, differences in injury exist among the different models, which makes their comparison difficult. In the CTX-induced injury model, a transient and reproducible acute injury is induced without affecting the vasculature or nerves, and this allows for the possibility of performing molecular and biochemical analyses of the whole muscle. Additionally, CTX injury models have a relatively low level of harm for animals in contrast to crushing models, which are invasive and associated with the risk of infection. This explains why CTX-induced injury models have been widely used in exploring the mechanisms of muscle regeneration. To understand the regeneration mechanisms in CTX-induced injury models, we explored all the studies and summarized the characteristics and injury positions, different models of CTX injury, and functional factors involved in the process of regeneration. The results show that the process of regeneration is similar in different mouse strains but that differences exist between gender. Regeneration is impaired in obese, diabetic, and aging mice, whereas exercise, electrical stimulation, and overloading facilitate the regeneration of damaged muscle. Non-SCs transplanted in damaged muscle following CTX injury can also differentiate into myogenic cells and facilitate myogenesis. The emphasis throughout was on the process of regeneration, the changes in the functional proteins involved in the processes of clearance of necrotic fiber debris, M1 to M2 macrophage conversion, SC activation, myoblast proliferation, differentiation and fusion, and fibrosis and calcification, which influence the final outcome of the regenerative activity. However, the inflammatory process in muscle injury and repair is complex, with different effects on muscle regeneration observed in various studies. Additionally, angiopoiesis and neurogenesis also influence the outcome of regeneration, which are easily ignored. Thus, further experiments are needed to explore the mechanisms of inflammatory response during muscle regeneration.
Author Contributions
Writing—original draft preparation, Y.W., J.L. and Y.L.; writing—review and editing, Y.W., J.L. and Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Natural Science Foundation of China (NO.31971102, NO.32271180, NO.31971101) and Shanghai Frontiers Science Research Base of Exercise and Metabolic Health.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Shi, X.; Garry, D.J. Myogenic regulatory factors transactivate the Tceal7 gene and modulate muscle differentiation. Biochem. J. 2010, 428, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, T.J.; Bryer, S.C.; Pucci, A.M.; Sisson, T.H. Mice deficient in plasminogen activator inhibitor-1 have improved skeletal muscle regeneration. Am. J. Physiol. Cell Physiol. 2005, 289, C217–C223. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Manabe, I.; Suzuki, Y.; Relaix, F.; Oishi, Y. Klf5 regulates muscle differentiation by directly targeting muscle-specific genes in cooperation with MyoD in mice. eLife 2016, 5, e17462. [Google Scholar] [CrossRef] [PubMed]
- Alves, J.M.; Martins, A.H.; Lameu, C.; Glaser, T.; Boukli, N.M.; Bassaneze, V.; Dariolli, R.; Nascimento, I.C.; Martins, P.C.M.; de Souza, H.D.N.; et al. Kinin-B2 Receptor Activity in Skeletal Muscle Regeneration and Myoblast Differentiation. Stem Cell Rev. Rep. 2019, 15, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Choi, S.; Liu, X.; Zhang, M.; Schageman, J.J.; Lee, S.Y.; Hart, R.; Lin, L.; Thurmond, F.A.; Williams, R.S. Highly coordinated gene regulation in mouse skeletal muscle regeneration. J. Biol. Chem. 2003, 278, 8826–8836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsumi, R.; Suzuki, T.; Do, M.Q.; Ohya, Y.; Anderson, J.E.; Shibata, A.; Kawaguchi, M.; Ohya, S.; Ohtsubo, H.; Mizunoya, W.; et al. Slow-Myofiber Commitment by Semaphorin 3A Secreted from Myogenic Stem Cells. Stem Cells 2017, 35, 1815–1834. [Google Scholar] [CrossRef] [Green Version]
- Ramadasan-Nair, R.; Gayathri, N.; Mishra, S.; Sunitha, B.; Mythri, R.B.; Nalini, A.; Subbannayya, Y.; Harsha, H.C.; Kolthur-Seetharam, U.; Srinivas Bharath, M.M. Mitochondrial alterations and oxidative stress in an acute transient mouse model of muscle degeneration: Implications for muscular dystrophy and related muscle pathologies. J. Biol. Chem. 2014, 289, 485–509. [Google Scholar] [CrossRef] [Green Version]
- Ohtsubo, H.; Sato, Y.; Suzuki, T.; Mizunoya, W.; Nakamura, M.; Tatsumi, R.; Ikeuchi, Y. APOBEC2 negatively regulates myoblast differentiation in muscle regeneration. Int. J. Biochem. Cell Biol. 2017, 85, 91–101. [Google Scholar] [CrossRef]
- Parise, G.; McKinnell, I.W.; Rudnicki, M.A. Muscle satellite cell and atypical myogenic progenitor response following exercise. Muscle Nerve 2008, 37, 611–619. [Google Scholar] [CrossRef]
- Armand, A.S.; Launay, T.; Gaspera, B.D.; Charbonnier, F.; Gallien, C.L.; Chanoine, C. Effects of eccentric treadmill running on mouse soleus: Degeneration/regeneration studied with Myf-5 and MyoD probes. Acta Physiol. Scand. 2003, 179, 75–84. [Google Scholar] [CrossRef]
- Czerwinska, A.M.; Streminska, W.; Ciemerych, M.A.; Grabowska, I. Mouse gastrocnemius muscle regeneration after mechanical or cardiotoxin injury. Folia Histochem. Cytobiol. 2012, 50, 144–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, J.E.; Jiang, M.S.; Gong, Q.H.; Yudkowsky, M.L.; Wieland, S.J. Effects of a cardiotoxin from Naja naja kaouthia venom on skeletal muscle: Involvement of calcium-induced calcium release, sodium ion currents and phospholipases A2 and C. Toxicon 1991, 29, 1489–1500. [Google Scholar] [CrossRef]
- Lin Shiau, S.Y.; Huang, M.C.; Lee, C.Y. Mechanism of action of cobra cardiotoxin in the skeletal muscle. J. Pharmacol. Exp. Ther. 1976, 196, 758–770. [Google Scholar] [PubMed]
- Aoki, Y.; Nagata, T.; Yokota, T.; Nakamura, A.; Wood, M.J.; Partridge, T.; Takeda, S. Highly efficient in vivo delivery of PMO into regenerating myotubes and rescue in laminin-alpha2 chain-null congenital muscular dystrophy mice. Hum. Mol. Genet. 2013, 22, 4914–4928. [Google Scholar] [CrossRef]
- Randazzo, D.; Khalique, U.; Belanto, J.J.; Kenea, A.; Talsness, D.M.; Olthoff, J.T.; Tran, M.D.; Zaal, K.J.; Pak, K.; Pinal-Fernandez, I.; et al. Persistent upregulation of the beta-tubulin tubb6, linked to muscle regeneration, is a source of microtubule disorganization in dystrophic muscle. Hum. Mol. Genet. 2019, 28, 1117–1135. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Gao, X.K.; Rao, X.S.; Shi, Y.P.; Liu, X.C.; Wang, F.Y.; Liu, Y.F.; Cong, X.X.; He, M.Y.; Xu, S.B.; et al. Hsp70 Interacts with Mitogen-Activated Protein Kinase (MAPK)-Activated Protein Kinase 2 To Regulate p38MAPK Stability and Myoblast Differentiation during Skeletal Muscle Regeneration. Mol. Cell. Biol. 2018, 38, e00211-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Boer, M.D.; Seynnes, O.R.; di Prampero, P.E.; Pisot, R.; Mekjavic, I.B.; Biolo, G.; Narici, M.V. Effect of 5 weeks horizontal bed rest on human muscle thickness and architecture of weight bearing and non-weight bearing muscles. Eur. J. Appl. Physiol. 2008, 104, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Morton, A.B.; Jacobsen, N.L.; Segal, S.S. Functionalizing biomaterials to promote neurovascular regeneration following skeletal muscle injury. Am. J. Physiol. Cell Physiol. 2021, 320, C1099–C1111. [Google Scholar] [CrossRef]
- Markert, C.; Petroski, G.F.; Childers, C.K.; McDonald, K.S.; Childers, M.K. Stretch-induced force deficits in murine extensor digitorum longus muscles after cardiotoxin injection. Muscle Nerve 2006, 34, 485–488. [Google Scholar] [CrossRef]
- Takahashi, Y.; Shimizu, T.; Kato, S.; Nara, M.; Suganuma, Y.; Sato, T.; Morii, T.; Yamada, Y.; Fujita, H. Reduction of Superoxide Dismutase 1 Delays Regeneration of Cardiotoxin-Injured Skeletal Muscle in KK/Ta-Ins2(Akita) Mice with Progressive Diabetic Nephropathy. Int. J. Mol. Sci. 2021, 22, 5491. [Google Scholar] [CrossRef]
- Vignaud, A.; Ramond, F.; Hourde, C.; Keller, A.; Butler-Browne, G.; Ferry, A. Diabetes provides an unfavorable environment for muscle mass and function after muscle injury in mice. Pathobiology 2007, 74, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.H.; Cheng, M.; Koh, T.J. Impaired muscle regeneration in ob/ob and db/db mice. Sci. World J. 2011, 11, 1525–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, M.P.; Al-Sajee, D.; D’Souza, D.M.; Rebalka, I.A.; Moradi, J.; Riddell, M.C.; Hawke, T.J. Impaired macrophage and satellite cell infiltration occurs in a muscle-specific fashion following injury in diabetic skeletal muscle. PLoS ONE 2013, 8, e70971. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, D.M.; Trajcevski, K.E.; Al-Sajee, D.; Wang, D.C.; Thomas, M.; Anderson, J.E.; Hawke, T.J. Diet-induced obesity impairs muscle satellite cell activation and muscle repair through alterations in hepatocyte growth factor signaling. Physiol. Rep. 2015, 3, e12506. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wang, H.; Lee, I.H.; Modi, S.; Wang, X.; Du, J.; Mitch, W.E. PTEN inhibition improves muscle regeneration in mice fed a high-fat diet. Diabetes 2010, 59, 1312–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inaba, S.; Hinohara, A.; Tachibana, M.; Tsujikawa, K.; Fukada, S.I. Muscle regeneration is disrupted by cancer cachexia without loss of muscle stem cell potential. PLoS ONE 2018, 13, e0205467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.P.; Shin, Y.J.; Kwon, K.S. microRNA for determining the age-related myogenic capabilities of skeletal muscle. BMB Rep. 2015, 48, 595–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouisel, E.; Vignaud, A.; Hourde, C.; Butler-Browne, G.; Ferry, A. Muscle weakness and atrophy are associated with decreased regenerative capacity and changes in mTOR signaling in skeletal muscles of venerable (18-24-month-old) dystrophic mdx mice. Muscle Nerve 2010, 41, 809–818. [Google Scholar] [CrossRef]
- Patsalos, A.; Pap, A.; Varga, T.; Trencsenyi, G.; Contreras, G.A.; Garai, I.; Papp, Z.; Dezso, B.; Pintye, E.; Nagy, L. In situ macrophage phenotypic transition is affected by altered cellular composition prior to acute sterile muscle injury. J. Physiol. 2017, 595, 5815–5842. [Google Scholar] [CrossRef] [Green Version]
- Ceco, E.; Celli, D.; Weinberg, S.; Shigemura, M.; Welch, L.C.; Volpe, L.; Chandel, N.S.; Bharat, A.; Lecuona, E.; Sznajder, J.I. Elevated CO2 Levels Delay Skeletal Muscle Repair by Increasing Fatty Acid Oxidation. Front. Physiol. 2020, 11, 630910. [Google Scholar] [CrossRef]
- Ohno, Y.; Matsuba, Y.; Hashimoto, N.; Sugiura, T.; Ohira, Y.; Yoshioka, T.; Goto, K. Suppression of Myostatin Stimulates Regenerative Potential of Injured Antigravitational Soleus Muscle in Mice under Unloading Condition. Int. J. Med. Sci. 2016, 13, 680–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuba, Y.; Goto, K.; Morioka, S.; Naito, T.; Akema, T.; Hashimoto, N.; Sugiura, T.; Ohira, Y.; Beppu, M.; Yoshioka, T. Gravitational unloading inhibits the regenerative potential of atrophied soleus muscle in mice. Acta Physiol. 2009, 196, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Joanisse, S.; Nederveen, J.P.; Baker, J.M.; Snijders, T.; Iacono, C.; Parise, G. Exercise conditioning in old mice improves skeletal muscle regeneration. FASEB J. 2016, 30, 3256–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horii, N.; Uchida, M.; Hasegawa, N.; Fujie, S.; Oyanagi, E.; Yano, H.; Hashimoto, T.; Iemitsu, M. Resistance training prevents muscle fibrosis and atrophy via down-regulation of C1q-induced Wnt signaling in senescent mice. FASEB J. 2018, 32, 3547–3559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiya, H.; Ogura, Y.; Ohno, Y.; Goto, A.; Nakamura, A.; Ohashi, K.; Uematsu, D.; Aoki, H.; Musha, H.; Goto, K. Microcurrent electrical neuromuscular stimulation facilitates regeneration of injured skeletal muscle in mice. J. Sports Sci. Med. 2015, 14, 297–303. [Google Scholar]
- Jinno, N.; Nagata, M.; Takahashi, T. Marginal zinc deficiency negatively affects recovery from muscle injury in mice. Biol. Trace Elem. Res. 2014, 158, 65–72. [Google Scholar] [CrossRef]
- Morioka, S.; Goto, K.; Kojima, A.; Naito, T.; Matsuba, Y.; Akema, T.; Fujiya, H.; Sugiura, T.; Ohira, Y.; Beppu, M.; et al. Functional overloading facilitates the regeneration of injured soleus muscles in mice. J. Physiol. Sci. 2008, 58, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Kohno, S.; Yamashita, Y.; Abe, T.; Hirasaka, K.; Oarada, M.; Ohno, A.; Teshima-Kondo, S.; Higashibata, A.; Choi, I.; Mills, E.M.; et al. Unloading stress disturbs muscle regeneration through perturbed recruitment and function of macrophages. J. Appl. Physiol. 2012, 112, 1773–1782. [Google Scholar] [CrossRef] [Green Version]
- Kimoloi, S.; Sen, A.; Guenther, S.; Braun, T.; Brugmann, T.; Sasse, P.; Wiesner, R.J.; Pla-Martin, D.; Baris, O.R. Combined fibre atrophy and decreased muscle regeneration capacity driven by mitochondrial DNA alterations underlie the development of sarcopenia. J. Cachexia Sarcopenia Muscle 2022, 13, 2132–2145. [Google Scholar] [CrossRef]
- Fearing, C.M.; Melton, D.W.; Lei, X.; Hancock, H.; Wang, H.; Sarwar, Z.U.; Porter, L.; McHale, M.; McManus, L.M.; Shireman, P.K. Increased Adipocyte Area in Injured Muscle With Aging and Impaired Remodeling in Female Mice. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 992–1004. [Google Scholar] [CrossRef] [Green Version]
- Chaiyasing, R.; Sugiura, A.; Ishikawa, T.; Ojima, K.; Warita, K.; Hosaka, Y.Z. Estrogen modulates the skeletal muscle regeneration process and myotube morphogenesis: Morphological analysis in mice with a low estrogen status. J. Vet. Med. Sci. 2021, 83, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- Rebalka, I.A.; Cao, A.W.; Raleigh, M.J.; Henriksbo, B.D.; Coleman, S.K.; Schertzer, J.D.; Hawke, T.J. Statin Therapy Negatively Impacts Skeletal Muscle Regeneration and Cutaneous Wound Repair in Type 1 Diabetic Mice. Front. Physiol. 2017, 8, 1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.; Conboy, M.J.; Conboy, I.M. Pharmacological inhibition of myostatin/TGF-beta receptor/pSmad3 signaling rescues muscle regenerative responses in mouse model of type 1 diabetes. Acta Pharmacol. Sin. 2013, 34, 1052–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHale, M.J.; Sarwar, Z.U.; Cardenas, D.P.; Porter, L.; Salinas, A.S.; Michalek, J.E.; McManus, L.M.; Shireman, P.K. Increased fat deposition in injured skeletal muscle is regulated by sex-specific hormones. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R331–R339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, Y.; Satoh, A.; Horinouchi, Y.; Hamano, H.; Watanabe, H.; Imao, M.; Imanishi, M.; Zamami, Y.; Takechi, K.; Izawa-Ishizawa, Y.; et al. Iron accumulation causes impaired myogenesis correlated with MAPK signaling pathway inhibition by oxidative stress. FASEB J. 2019, 33, 9551–9564. [Google Scholar] [CrossRef] [Green Version]
- Attia, M.; Maurer, M.; Robinet, M.; Le Grand, F.; Fadel, E.; Le Panse, R.; Butler-Browne, G.; Berrih-Aknin, S. Muscle satellite cells are functionally impaired in myasthenia gravis: Consequences on muscle regeneration. Acta Neuropathol. 2017, 134, 869–888. [Google Scholar] [CrossRef] [PubMed]
- Saliu, T.P.; Kumrungsee, T.; Mitsumoto, K.; Chen, S.; Yanaka, N. Satellite cell content and muscle regeneration in a mouse model of NAFLD. Nutrition 2022, 96, 111570. [Google Scholar] [CrossRef]
- Rahman, F.A.; Angus, S.A.; Stokes, K.; Karpowicz, P.; Krause, M.P. Impaired ECM Remodeling and Macrophage Activity Define Necrosis and Regeneration Following Damage in Aged Skeletal Muscle. Int. J. Mol. Sci. 2020, 21, 4575. [Google Scholar] [CrossRef]
- Paiva-Oliveira, E.L.; da Silva, R.F.; Bellio, M.; Quirico-Santos, T.; Lagrota-Candido, J. Pattern of cardiotoxin-induced muscle remodeling in distinct TLR-4 deficient mouse strains. Histochem. Cell Biol. 2017, 148, 49–60. [Google Scholar] [CrossRef]
- Yoshioka, K.; Kitajima, Y.; Seko, D.; Tsuchiya, Y.; Ono, Y. The body region specificity in murine models of muscle regeneration and atrophy. Acta Physiol. 2021, 231, e13553. [Google Scholar] [CrossRef]
- Nagata, K.; Nakamura, T.; Fujihara, S.; Tanaka, E. Ultrasound modulates the inflammatory response and promotes muscle regeneration in injured muscles. Ann. Biomed. Eng. 2013, 41, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- LaBarge, S.; McDonald, M.; Smith-Powell, L.; Auwerx, J.; Huss, J.M. Estrogen-related receptor-alpha (ERRalpha) deficiency in skeletal muscle impairs regeneration in response to injury. FASEB J. 2014, 28, 1082–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.A.; Ryan, T.E.; Lin, C.T.; Inigo, M.M.R.; Green, T.D.; Brault, J.J.; Spangenburg, E.E.; McClung, J.M. Diminished force production and mitochondrial respiratory deficits are strain-dependent myopathies of subacute limb ischemia. J. Vasc. Surg. 2017, 65, 1504–1514.e1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, D.; Besnard, A.; Latil, M.; Jouvion, G.; Briand, D.; Thepenier, C.; Pascal, Q.; Guguin, A.; Gayraud-Morel, B.; Cavaillon, J.M.; et al. Comparative Study of Injury Models for Studying Muscle Regeneration in Mice. PLoS ONE 2016, 11, e0147198. [Google Scholar] [CrossRef] [Green Version]
- Heredia, J.E.; Mukundan, L.; Chen, F.M.; Mueller, A.A.; Deo, R.C.; Locksley, R.M.; Rando, T.A.; Chawla, A. Type 2 innate signals stimulate fibro/adipogenic progenitors to facilitate muscle regeneration. Cell 2013, 153, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Dinulovic, I.; Furrer, R.; Di Fulvio, S.; Ferry, A.; Beer, M.; Handschin, C. PGC-1alpha modulates necrosis, inflammatory response, and fibrotic tissue formation in injured skeletal muscle. Skelet. Muscle 2016, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Gu, R.; Song, Y.; Ding, M.; Huang, T.; Guo, M.; Xiao, J.; Huang, W.; Liao, H. Calcium/Calmodulin-Dependent Protein Kinase IV (CaMKIV) Mediates Acute Skeletal Muscle Inflammatory Response. Inflammation 2018, 41, 199–212. [Google Scholar] [CrossRef]
- Neves Jde, C.; Rizzato, V.R.; Fappi, A.; Garcia, M.M.; Chadi, G.; van de Vlekkert, D.; d’Azzo, A.; Zanoteli, E. Neuraminidase-1 mediates skeletal muscle regeneration. Biochim. Biophys. Acta 2015, 1852, 1755–1764. [Google Scholar] [CrossRef] [Green Version]
- Liao, Z.H.; Huang, T.; Xiao, J.W.; Gu, R.C.; Ouyang, J.; Wu, G.; Liao, H. Estrogen signaling effects on muscle-specific immune responses through controlling the recruitment and function of macrophages and T cells. Skelet. Muscle 2019, 9, 20. [Google Scholar] [CrossRef]
- Kohno, S.; Ueji, T.; Abe, T.; Nakao, R.; Hirasaka, K.; Oarada, M.; Harada-Sukeno, A.; Ohno, A.; Higashibata, A.; Mukai, R.; et al. Rantes secreted from macrophages disturbs skeletal muscle regeneration after cardiotoxin injection in Cbl-b-deficient mice. Muscle Nerve 2011, 43, 223–229. [Google Scholar] [CrossRef]
- Wang, H.; Melton, D.W.; Porter, L.; Sarwar, Z.U.; McManus, L.M.; Shireman, P.K. Altered macrophage phenotype transition impairs skeletal muscle regeneration. Am. J. Pathol. 2014, 184, 1167–1184. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Pierce, S.A.; von Drehle, M.; Ivey, K.N.; Morgan, J.A.; Blau, H.M.; Srivastava, D. skNAC, a Smyd1-interacting transcription factor, is involved in cardiac development and skeletal muscle growth and regeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 20750–20755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Boadu, E.; Mercan, F.; Le, A.M.; Flach, R.J.; Zhang, L.; Tyner, K.J.; Olwin, B.B.; Bennett, A.M. MAP kinase phosphatase-1 deficiency impairs skeletal muscle regeneration and exacerbates muscular dystrophy. FASEB J. 2010, 24, 2985–2997. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Shi, D.; Ding, M.; Huang, T.; Gu, R.; Xiao, J.; Xian, C.J.; Dong, J.; Wang, L.; Liao, H. Calmodulin-dependent signalling pathways are activated and mediate the acute inflammatory response of injured skeletal muscle. J. Physiol. 2019, 597, 5161–5177. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, P.; Song, T.; Radzyukevich, T.L.; Sadayappan, S.; Lingrel, J.B.; Heiny, J.A. KLF2 in Myeloid Lineage Cells Regulates the Innate Immune Response during Skeletal Muscle Injury and Regeneration. iScience 2019, 17, 334–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Li, Y.; Guo, X.; Wu, Z.; Zhang, W. Loss of STAT1 in bone marrow-derived cells accelerates skeletal muscle regeneration. PLoS ONE 2012, 7, e37656. [Google Scholar] [CrossRef] [PubMed]
- Kozakowska, M.; Pietraszek-Gremplewicz, K.; Ciesla, M.; Seczynska, M.; Bronisz-Budzynska, I.; Podkalicka, P.; Bukowska-Strakova, K.; Loboda, A.; Jozkowicz, A.; Dulak, J. Lack of Heme Oxygenase-1 Induces Inflammatory Reaction and Proliferation of Muscle Satellite Cells after Cardiotoxin-Induced Skeletal Muscle Injury. Am. J. Pathol. 2018, 188, 491–506. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Cheng, N.; Qiao, B.; Zhang, F.; Wu, J.; Liu, C.; Li, Y.; Du, J. Age-related decline of interferon-gamma responses in macrophage impairs satellite cell proliferation and regeneration. J. Cachexia Sarcopenia Muscle 2020, 11, 1291–1305. [Google Scholar] [CrossRef]
- Yaden, B.C.; Wang, Y.X.; Wilson, J.M.; Culver, A.E.; Milner, A.; Datta-Mannan, A.; Shetler, P.; Croy, J.E.; Dai, G.; Krishnan, V. Inhibition of activin A ameliorates skeletal muscle injury and rescues contractile properties by inducing efficient remodeling in female mice. Am. J. Pathol. 2014, 184, 1152–1166. [Google Scholar] [CrossRef]
- Mothe-Satney, I.; Piquet, J.; Murdaca, J.; Sibille, B.; Grimaldi, P.A.; Neels, J.G.; Rousseau, A.S. Peroxisome Proliferator Activated Receptor Beta (PPARbeta) activity increases the immune response and shortens the early phases of skeletal muscle regeneration. Biochimie 2017, 136, 33–41. [Google Scholar] [CrossRef]
- Tanaka, Y.; Kita, S.; Nishizawa, H.; Fukuda, S.; Fujishima, Y.; Obata, Y.; Nagao, H.; Masuda, S.; Nakamura, Y.; Shimizu, Y.; et al. Adiponectin promotes muscle regeneration through binding to T-cadherin. Sci. Rep. 2019, 9, 16. [Google Scholar] [CrossRef]
- Sugihara, H.; Miyaji, K.; Yamanouchi, K.; Matsuwaki, T.; Nishihara, M. Progranulin deficiency leads to prolonged persistence of macrophages, accompanied with myofiber hypertrophy in regenerating muscle. J. Vet. Med. Sci. 2018, 80, 346–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Sicco, C.; Reverberi, D.; Balbi, C.; Ulivi, V.; Principi, E.; Pascucci, L.; Becherini, P.; Bosco, M.C.; Varesio, L.; Franzin, C.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles as Mediators of Anti-Inflammatory Effects: Endorsement of Macrophage Polarization. Stem Cells Transl. Med. 2017, 6, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Sun, L.; Kawabata, Y.; Murayama, F.; Maegawa, T.; Nikawa, T.; Hirasaka, K. Balenine, Imidazole Dipeptide Promotes Skeletal Muscle Regeneration by Regulating Phagocytosis Properties of Immune Cells. Mar. Drugs 2022, 20, 313. [Google Scholar] [CrossRef]
- Cardoso, E.S.; Santana, T.A.; Diniz, P.B.; Montalvao, M.M.; Bani, C.C.; Thomazzi, S.M. Thymol accelerates the recovery of the skeletal muscle of mice injured with cardiotoxin. J. Pharm. Pharmacol. 2016, 68, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.G.; Zhu, Z.Q.; He, Z.Y.; Cheng, P.; Liang, S.; Chen, A.M.; Yang, Q. Endogenous conversion of n-6 to n-3 polyunsaturated fatty acids facilitates the repair of cardiotoxin-induced skeletal muscle injury in fat-1 mice. Aging 2021, 13, 8454–8466. [Google Scholar] [CrossRef] [PubMed]
- Chaweewannakorn, C.; Tsuchiya, M.; Koide, M.; Hatakeyama, H.; Tanaka, Y.; Yoshida, S.; Sugawara, S.; Hagiwara, Y.; Sasaki, K.; Kanzaki, M. Roles of IL-1alpha/beta in regeneration of cardiotoxin-injured muscle and satellite cell function. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R90–R103. [Google Scholar] [CrossRef] [Green Version]
- Nikolaidis, N.; Senf, S.M.; Howard, T.M.; Ahn, B.; Ferreira, L.F.; Judge, A.R. Loss of the Inducible Hsp70 Delays the Inflammatory Response to Skeletal Muscle Injury and Severely Impairs Muscle Regeneration. PLoS ONE 2013, 8, e62687. [Google Scholar] [CrossRef] [Green Version]
- Mojumdar, K.; Giordano, C.; Lemaire, C.; Liang, F.; Divangahi, M.; Qureshi, S.T.; Petrof, B.J. Divergent impact of Toll-like receptor 2 deficiency on repair mechanisms in healthy muscle versus Duchenne muscular dystrophy. J. Pathol. 2016, 239, 10–22. [Google Scholar] [CrossRef]
- Varga, T.; Mounier, R.; Gogolak, P.; Poliska, S.; Chazaud, B.; Nagy, L. Tissue LyC6- macrophages are generated in the absence of circulating LyC6- monocytes and Nur77 in a model of muscle regeneration. J. Immunol. 2013, 191, 5695–5701. [Google Scholar] [CrossRef] [Green Version]
- Ochoa, O.; Sun, D.; Reyes-Reyna, S.M.; Waite, L.L.; Michalek, J.E.; McManus, L.M.; Shireman, P.K. Delayed angiogenesis and VEGF production in CCR2-/- mice during impaired skeletal muscle regeneration. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R651–R661. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, Y.; Wu, Y.; Wang, L.; Wang, X.; Du, J. Interleukin-6/signal transducer and activator of transcription 3 (STAT3) pathway is essential for macrophage infiltration and myoblast proliferation during muscle regeneration. J. Biol. Chem. 2013, 288, 1489–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Xiao, Z.; Qu, C.; Cui, W.; Wang, X.; Du, J. CD8 T cells are involved in skeletal muscle regeneration through facilitating MCP-1 secretion and Gr1(high) macrophage infiltration. J. Immunol. 2014, 193, 5149–5160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, C.O.; McHale, M.J.; Wells, J.T.; Ochoa, O.; Michalek, J.E.; McManus, L.M.; Shireman, P.K. Regulation of skeletal muscle regeneration by CCR2-activating chemokines is directly related to macrophage recruitment. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R832–R842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.; Nguyen, M.H.; Fantuzzi, G.; Koh, T.J. Endogenous interferon-gamma is required for efficient skeletal muscle regeneration. Am. J. Physiol. Cell Physiol. 2008, 294, C1183–C1191. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, C.; Li, Y.; Miwa, T.; Liu, C.; Cui, W.; Song, W.C.; Du, J. Complement C3a signaling facilitates skeletal muscle regeneration by regulating monocyte function and trafficking. Nat. Commun. 2017, 8, 2078. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Martinez, C.O.; Ochoa, O.; Ruiz-Willhite, L.; Bonilla, J.R.; Centonze, V.E.; Waite, L.L.; Michalek, J.E.; McManus, L.M.; Shireman, P.K. Bone marrow-derived cell regulation of skeletal muscle regeneration. FASEB J. 2009, 23, 382–395. [Google Scholar] [CrossRef]
- Nishimura, D.; Sakai, H.; Sato, T.; Sato, F.; Nishimura, S.; Toyama-Sorimachi, N.; Bartsch, J.W.; Sehara-Fujisawa, A. Roles of ADAM8 in elimination of injured muscle fibers prior to skeletal muscle regeneration. Mech. Dev. 2015, 135, 58–67. [Google Scholar] [CrossRef]
- Al-Zaeed, N.; Budai, Z.; Szondy, Z.; Sarang, Z. TAM kinase signaling is indispensable for proper skeletal muscle regeneration in mice. Cell Death Dis. 2021, 12, 611. [Google Scholar] [CrossRef]
- Zhang, J.; Qu, C.; Li, T.; Cui, W.; Wang, X.; Du, J. Phagocytosis mediated by scavenger receptor class BI promotes macrophage transition during skeletal muscle regeneration. J. Biol. Chem. 2019, 294, 15672–15685. [Google Scholar] [CrossRef]
- Jin, R.M.; Warunek, J.; Wohlfert, E.A. Chronic infection stunts macrophage heterogeneity and disrupts immune-mediated myogenesis. JCI Insight 2018, 3, e121549. [Google Scholar] [CrossRef] [PubMed]
- Bronisz-Budzynska, I.; Kozakowska, M.; Podkalicka, P.; Kachamakova-Trojanowska, N.; Loboda, A.; Dulak, J. The role of Nrf2 in acute and chronic muscle injury. Skelet. Muscle 2020, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Tarban, N.; Halász, H.; Gogolák, P.; Garabuczi, É.; Moise, A.R.; Palczewski, K.; Sarang, Z.; Szondy, Z. Regenerating Skeletal Muscle Compensates for the Impaired Macrophage Functions Leading to Normal Muscle Repair in Retinol Saturase Null Mice. Cells 2022, 11, 1333. [Google Scholar] [CrossRef] [PubMed]
- Dalle, S.; Poffe, C.; Hiroux, C.; Suhr, F.; Deldicque, L.; Koppo, K. Ibuprofen does not impair skeletal muscle regeneration upon cardiotoxin-induced injury. Physiol. Res. 2020, 69, 847–859. [Google Scholar] [CrossRef]
- Shen, W.; Li, Y.; Zhu, J.; Schwendener, R.; Huard, J. Interaction between macrophages, TGF-beta1, and the COX-2 pathway during the inflammatory phase of skeletal muscle healing after injury. J. Cell. Physiol. 2008, 214, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, A.S.; Murdaca, J.; Le Menn, G.; Sibille, B.; Wahli, W.; Le Garf, S.; Chinetti, G.; Neels, J.G.; Mothe-Satney, I. Invalidation of the Transcriptional Modulator of Lipid Metabolism PPARbeta/delta in T Cells Prevents Age-Related Alteration of Body Composition and Loss of Endurance Capacity. Front. Physiol. 2021, 12, 587753. [Google Scholar] [CrossRef] [PubMed]
- Mann, A.O.; Hanna, B.S.; Munoz-Rojas, A.R.; Sandrock, I.; Prinz, I.; Benoist, C.; Mathis, D. IL-17A-producing gammadeltaT cells promote muscle regeneration in a microbiota-dependent manner. J. Exp. Med. 2022, 219, e20211504. [Google Scholar] [CrossRef]
- Ding, M.; Huang, T.; Zhu, R.; Gu, R.; Shi, D.; Xiao, J.; Guo, M.; Li, J.; Hu, J.; Liao, H. Immunological Behavior Analysis of Muscle Cells under IFN-gamma Stimulation in Vitro and in Vivo. Anat. Rec. 2018, 301, 1551–1563. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Huang, J.; Liao, Z.; Lan, H.; Jian, X.; Gu, R.; Ouyang, J.; Hu, J.; Liao, H. Regenerating myofiber directs Tregs and Th17 responses in inflamed muscle through the intrinsic TGF-beta signaling-mediated IL-6 production. Am J. Physiol. Endocrinol. Metab. 2022, 323, E92–E106. [Google Scholar] [CrossRef]
- Pierce, A.P.; de Waal, E.; McManus, L.M.; Shireman, P.K.; Chaudhuri, A.R. Oxidation and structural perturbation of redox-sensitive enzymes in injured skeletal muscle. Free Radic. Biol. Med. 2007, 43, 1584–1593. [Google Scholar] [CrossRef]
- Sciorati, C.; Rigamonti, E.; Manfredi, A.A.; Rovere-Querini, P. Cell death, clearance and immunity in the skeletal muscle. Cell Death Differ. 2016, 23, 927–937. [Google Scholar] [CrossRef]
- Budai, Z.; Al-Zaeed, N.; Szentesi, P.; Halasz, H.; Csernoch, L.; Szondy, Z.; Sarang, Z. Impaired Skeletal Muscle Development and Regeneration in Transglutaminase 2 Knockout Mice. Cells 2021, 10, 3089. [Google Scholar] [CrossRef] [PubMed]
- Sarang, Z.; Saghy, T.; Budai, Z.; Ujlaky-Nagy, L.; Bedekovics, J.; Beke, L.; Mehes, G.; Nagy, G.; Ruhl, R.; Moise, A.R.; et al. Retinol Saturase Knock-Out Mice are Characterized by Impaired Clearance of Apoptotic Cells and Develop Mild Autoimmunity. Biomolecules 2019, 9, 737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Zou, X.; Wu, R.; Zhong, R.; Zhu, D.; Zhang, Y. Accelerated regeneration of the skeletal muscle in RNF13-knockout mice is mediated by macrophage-secreted IL-4/IL-6. Protein Cell 2014, 5, 235–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, R.N.; Tajbakhsh, S.; Mouly, V.; Cossu, G.; Buckingham, M.; Butler-Browne, G.S. In vivo satellite cell activation via Myf5 and MyoD in regenerating mouse skeletal muscle. J. Cell Sci. 1999, 112 Pt 17, 2895–2901. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Han, Y.; Liu, J.; Liu, L.; Zheng, L.; Chen, Y.; Xia, R.; Yao, D.; Cai, X.; Xu, X. Rbm24 modulates adult skeletal muscle regeneration via regulation of alternative splicing. Theranostics 2020, 10, 11159–11177. [Google Scholar] [CrossRef]
- Galimov, A.; Merry, T.L.; Luca, E.; Rushing, E.J.; Mizbani, A.; Turcekova, K.; Hartung, A.; Croce, C.M.; Ristow, M.; Krutzfeldt, J. MicroRNA-29a in Adult Muscle Stem Cells Controls Skeletal Muscle Regeneration During Injury and Exercise Downstream of Fibroblast Growth Factor-2. Stem Cells 2016, 34, 768–780. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Akasaki, Y.; Sato, K.; Ouchi, N.; Izumiya, Y.; Walsh, K. Insulin-like 6 is induced by muscle injury and functions as a regenerative factor. J. Biol. Chem. 2010, 285, 36060–36069. [Google Scholar] [CrossRef] [Green Version]
- Hawke, T.J.; Atkinson, D.J.; Kanatous, S.B.; Van der Ven, P.F.; Goetsch, S.C.; Garry, D.J. Xin, an actin binding protein, is expressed within muscle satellite cells and newly regenerated skeletal muscle fibers. Am. J. Physiol. Cell Physiol. 2007, 293, C1636–C1644. [Google Scholar] [CrossRef] [Green Version]
- Al-Sajee, D.; Nissar, A.A.; Coleman, S.K.; Rebalka, I.A.; Chiang, A.; Wathra, R.; van der Ven, P.F.; Orfanos, Z.; Hawke, T.J. Xin-deficient mice display myopathy, impaired contractility, attenuated muscle repair and altered satellite cell functionality. Acta Physiol. 2015, 214, 248–260. [Google Scholar] [CrossRef]
- Nissar, A.A.; Zemanek, B.; Labatia, R.; Atkinson, D.J.; van der Ven, P.F.; Furst, D.O.; Hawke, T.J. Skeletal muscle regeneration is delayed by reduction in Xin expression: Consequence of impaired satellite cell activation? Am. J. Physiol. Cell Physiol. 2012, 302, C220–C227. [Google Scholar] [CrossRef] [PubMed]
- Hillege, M.M.G.; Shi, A.; Galli, R.A.; Wu, G.; Bertolino, P.; Hoogaars, W.M.H.; Jaspers, R.T. Lack of Tgfbr1 and Acvr1b synergistically stimulates myofibre hypertrophy and accelerates muscle regeneration. Elife 2022, 11, e77610. [Google Scholar] [CrossRef] [PubMed]
- Shelar, S.B.; Narasimhan, M.; Shanmugam, G.; Litovsky, S.H.; Gounder, S.S.; Karan, G.; Arulvasu, C.; Kensler, T.W.; Hoidal, J.R.; Darley-Usmar, V.M.; et al. Disruption of nuclear factor (erythroid-derived-2)-like 2 antioxidant signaling: A mechanism for impaired activation of stem cells and delayed regeneration of skeletal muscle. FASEB J. 2016, 30, 1865–1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebalka, I.A.; Monaco, C.M.F.; Varah, N.E.; Berger, T.; D’Souza, D.M.; Zhou, S.; Mak, T.W.; Hawke, T.J. Loss of the adipokine lipocalin-2 impairs satellite cell activation and skeletal muscle regeneration. Am. J. Physiol. Cell Physiol. 2018, 315, C714–C721. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, G.J.; van Gestel, T.J.M.; In‘t Groen, S.L.M.; de Jong, B.; Boomaars, B.; Tarallo, A.; Cardone, M.; Parenti, G.; van der Ploeg, A.T.; Pijnappel, W. Satellite cells maintain regenerative capacity but fail to repair disease-associated muscle damage in mice with Pompe disease. Acta Neuropathol. Commun. 2018, 6, 119. [Google Scholar] [CrossRef] [Green Version]
- Serra, C.; Tangherlini, F.; Rudy, S.; Lee, D.; Toraldo, G.; Sandor, N.L.; Zhang, A.; Jasuja, R.; Bhasin, S. Testosterone improves the regeneration of old and young mouse skeletal muscle. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Pan, S.; Liu, S.; Zhang, S.; Willerson, J.T.; Martin, J.F.; Dixon, R.A.F. Suppressing Hippo signaling in the stem cell niche promotes skeletal muscle regeneration. Stem Cells 2021, 39, 737–749. [Google Scholar] [CrossRef]
- Zeng, P.; Han, W.; Li, C.; Li, H.; Zhu, D.; Zhang, Y.; Liu, X. miR-378 attenuates muscle regeneration by delaying satellite cell activation and differentiation in mice. Acta Biochim. Biophys. Sin. 2016, 48, 833–839. [Google Scholar] [CrossRef] [Green Version]
- Lagalice, L.; Pichon, J.; Gougeon, E.; Soussi, S.; Deniaud, J.; Ledevin, M.; Maurier, V.; Leroux, I.; Durand, S.; Ciron, C.; et al. Satellite cells fail to contribute to muscle repair but are functional in Pompe disease (glycogenosis type II). Acta Neuropathol. Commun. 2018, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Mizbani, A.; Luca, E.; Rushing, E.J.; Krutzfeldt, J. MicroRNA deep sequencing in two adult stem cell populations identifies miR-501 as a novel regulator of myosin heavy chain during muscle regeneration. Development 2016, 143, 4137–4148. [Google Scholar] [CrossRef] [Green Version]
- Fiore, P.F.; Benedetti, A.; Sandona, M.; Madaro, L.; De Bardi, M.; Saccone, V.; Puri, P.L.; Gargioli, C.; Lozanoska-Ochser, B.; Bouche, M. Lack of PKCtheta Promotes Regenerative Ability of Muscle Stem Cells in Chronic Muscle Injury. Int. J. Mol. Sci. 2020, 21, 932. [Google Scholar] [CrossRef] [PubMed]
- Fortier, M.; Figeac, N.; White, R.B.; Knopp, P.; Zammit, P.S. Sphingosine-1-phosphate receptor 3 influences cell cycle progression in muscle satellite cells. Dev. Biol. 2013, 382, 504–516. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Zhu, Q.; Guo, C.; Yuan, R.; Zhang, X.; Nie, Y.; Chen, L.; Fang, Y.; Chen, K.; Zhang, J.; et al. MLL1 promotes myogenesis by epigenetically regulating Myf5. Cell Prolif. 2020, 53, e12744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sincennes, M.C.; Brun, C.E.; Lin, A.Y.T.; Rosembert, T.; Datzkiw, D.; Saber, J.; Ming, H.; Kawabe, Y.I.; Rudnicki, M.A. Acetylation of PAX7 controls muscle stem cell self-renewal and differentiation potential in mice. Nat. Commun. 2021, 12, 3253. [Google Scholar] [CrossRef] [PubMed]
- Naito, T.; Goto, K.; Morioka, S.; Matsuba, Y.; Akema, T.; Sugiura, T.; Ohira, Y.; Beppu, M.; Yoshioka, T. Administration of granulocyte colony-stimulating factor facilitates the regenerative process of injured mice skeletal muscle via the activation of Akt/GSK3alphabeta signals. Eur. J. Appl. Physiol. 2009, 105, 643–651. [Google Scholar] [CrossRef]
- Price, F.D.; von Maltzahn, J.; Bentzinger, C.F.; Dumont, N.A.; Yin, H.; Chang, N.C.; Wilson, D.H.; Frenette, J.; Rudnicki, M.A. Inhibition of JAK-STAT signaling stimulates adult satellite cell function. Nat. Med. 2014, 20, 1174–1181. [Google Scholar] [CrossRef] [Green Version]
- Angione, A.R.; Jiang, C.; Pan, D.; Wang, Y.X.; Kuang, S. PPARdelta regulates satellite cell proliferation and skeletal muscle regeneration. Skelet. Muscle 2011, 1, 33. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, S.; Koya, T.; Ohno, Y.; Goto, A.; Ikuita, A.; Suzuki, M.; Ohira, T.; Egawa, T.; Nakai, A.; Sugiura, T.; et al. Regeneration of injured skeletal muscle in heat shock transcription factor 1-null mice. Physiol. Rep. 2013, 1, e00071. [Google Scholar] [CrossRef]
- Ahrens, H.E.; Huettemeister, J.; Schmidt, M.; Kaether, C.; von Maltzahn, J. Klotho expression is a prerequisite for proper muscle stem cell function and regeneration of skeletal muscle. Skelet. Muscle 2018, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, K.; Furuichi, Y.; Yamamoto, M.; Takahashi, M.; Akimoto, Y.; Ishikawa, T.; Shimizu, T.; Fujimoto, M.; Takada-Watanabe, A.; Hayashi, A.; et al. R3hdml regulates satellite cell proliferation and differentiation. EMBO Rep. 2019, 20, e47957. [Google Scholar] [CrossRef]
- Bye, A.J.H.; Pugazhendhi, D.; Woodhouse, S.; Brien, P.; Watson, R.; Turner, M.; Pell, J. The RNA-binding proteins Zfp36l1 and Zfp36l2 act redundantly in myogenesis. Skelet. Muscle 2018, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Tonami, K.; Hata, S.; Ojima, K.; Ono, Y.; Kurihara, Y.; Amano, T.; Sato, T.; Kawamura, Y.; Kurihara, H.; Sorimachi, H. Calpain-6 deficiency promotes skeletal muscle development and regeneration. PLoS Genet. 2013, 9, e1003668. [Google Scholar] [CrossRef] [PubMed]
- Accornero, F.; Kanisicak, O.; Tjondrokoesoemo, A.; Attia, A.C.; McNally, E.M.; Molkentin, J.D. Myofiber-specific inhibition of TGFbeta signaling protects skeletal muscle from injury and dystrophic disease in mice. Hum. Mol. Genet. 2014, 23, 6903–6915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ry, P.M.; Minogue, P.; Hodges, B.L.; Burkin, D.J. Laminin-111 improves muscle repair in a mouse model of merosin-deficient congenital muscular dystrophy. Hum. Mol. Genet. 2014, 23, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosoyama, T.; Nishijo, K.; Prajapati, S.I.; Li, G.; Keller, C. Rb1 gene inactivation expands satellite cell and postnatal myoblast pools. J. Biol. Chem. 2011, 286, 19556–19564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rion, N.; Castets, P.; Lin, S.; Enderle, L.; Reinhard, J.R.; Ruegg, M.A. mTORC2 affects the maintenance of the muscle stem cell pool. Skelet. Muscle 2019, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Galvez, S.; Tiwari, S.; Rezk, B.M.; Semprun-Prieto, L.; Higashi, Y.; Sukhanov, S.; Yablonka-Reuveni, Z.; Delafontaine, P. Angiotensin II inhibits satellite cell proliferation and prevents skeletal muscle regeneration. J. Biol. Chem. 2013, 288, 23823–23832. [Google Scholar] [CrossRef] [Green Version]
- Armand, A.S.; Laziz, I.; Djeghloul, D.; Lecolle, S.; Bertrand, A.T.; Biondi, O.; De Windt, L.J.; Chanoine, C. Apoptosis-inducing factor regulates skeletal muscle progenitor cell number and muscle phenotype. PLoS ONE 2011, 6, e27283. [Google Scholar] [CrossRef]
- Milanesi, A.; Lee, J.W.; Yang, A.; Liu, Y.Y.; Sedrakyan, S.; Cheng, S.Y.; Perin, L.; Brent, G.A. Thyroid Hormone Receptor Alpha is Essential to Maintain the Satellite Cell Niche During Skeletal Muscle Injury and Sarcopenia of Aging. Thyroid 2017, 27, 1316–1322. [Google Scholar] [CrossRef]
- Castets, P.; Bertrand, A.T.; Beuvin, M.; Ferry, A.; Le Grand, F.; Castets, M.; Chazot, G.; Rederstorff, M.; Krol, A.; Lescure, A.; et al. Satellite cell loss and impaired muscle regeneration in selenoprotein N deficiency. Hum. Mol. Genet. 2011, 20, 694–704. [Google Scholar] [CrossRef] [Green Version]
- Fujimaki, S.; Seko, D.; Kitajima, Y.; Yoshioka, K.; Tsuchiya, Y.; Masuda, S.; Ono, Y. Notch1 and Notch2 Coordinately Regulate Stem Cell Function in the Quiescent and Activated States of Muscle Satellite Cells. Stem Cells 2018, 36, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.P.; Bellamy, L.M.; Lisio, M.D.; Parise, G. Captopril treatment induces hyperplasia but inhibits myonuclear accretion following severe myotrauma in murine skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R363–R369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaafar Marican, N.H.; Cruz-Migoni, S.B.; Borycki, A.G. Asymmetric Distribution of Primary Cilia Allocates Satellite Cells for Self-Renewal. Stem Cell Rep. 2016, 6, 798–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, C.; Murad, K.B.A.; Tan, A.L.T.; Yada, S.; Sagiraju, S.; Bode, P.K.; Barker, N. Lgr5 Marks Adult Progenitor Cells Contributing to Skeletal Muscle Regeneration and Sarcoma Formation. Cell Rep. 2020, 33, 108535. [Google Scholar] [CrossRef] [PubMed]
- Buono, R.; Vantaggiato, C.; Pisa, V.; Azzoni, E.; Bassi, M.T.; Brunelli, S.; Sciorati, C.; Clementi, E. Nitric oxide sustains long-term skeletal muscle regeneration by regulating fate of satellite cells via signaling pathways requiring Vangl2 and cyclic GMP. Stem Cells 2012, 30, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Urciuolo, A.; Quarta, M.; Morbidoni, V.; Gattazzo, F.; Molon, S.; Grumati, P.; Montemurro, F.; Tedesco, F.S.; Blaauw, B.; Cossu, G.; et al. Collagen VI regulates satellite cell self-renewal and muscle regeneration. Nat. Commun. 2013, 4, 1964. [Google Scholar] [CrossRef] [Green Version]
- Alexeev, V.; Arita, M.; Donahue, A.; Bonaldo, P.; Chu, M.L.; Igoucheva, O. Human adipose-derived stem cell transplantation as a potential therapy for collagen VI-related congenital muscular dystrophy. Stem Cell Res. Ther. 2014, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Brien, P.; Pugazhendhi, D.; Woodhouse, S.; Oxley, D.; Pell, J.M. p38alpha MAPK regulates adult muscle stem cell fate by restricting progenitor proliferation during postnatal growth and repair. Stem Cells 2013, 31, 1597–1610. [Google Scholar] [CrossRef]
- Hawke, T.J.; Meeson, A.P.; Jiang, N.; Graham, S.; Hutcheson, K.; DiMaio, J.M.; Garry, D.J. p21 is essential for normal myogenic progenitor cell function in regenerating skeletal muscle. Am. J. Physiol. Cell Physiol. 2003, 285, C1019–C1027. [Google Scholar] [CrossRef]
- Cortez-Toledo, O.; Schnair, C.; Sangngern, P.; Metzger, D.; Chao, L.C. Nur77 deletion impairs muscle growth during developmental myogenesis and muscle regeneration in mice. PLoS ONE 2017, 12, e0171268. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.L.; Li, G.Z.; Chou, C.Y.; Tsai, M.S.; Chen, Y.P.; Li, C.J.; Liou, G.G.; Chang, W.W.; Chen, S.L.; Wang, S.H. Double homeobox gene, Duxbl, promotes myoblast proliferation and abolishes myoblast differentiation by blocking MyoD transactivation. Cell Tissue Res. 2014, 358, 551–566. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Melton, D.W.; Gelfond, J.A.; McManus, L.M.; Shireman, P.K. MiR-351 transiently increases during muscle regeneration and promotes progenitor cell proliferation and survival upon differentiation. Physiol. Genom. 2012, 44, 1042–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, C.; Sinha, K.; Pan, H.; Cui, Y.; Guo, P.; Lin, C.Y.; Yang, F.; Deng, Z.; Eltzschig, H.K.; Lu, A.; et al. Markers of Accelerated Skeletal Muscle Regenerative Response in Murphy Roths Large Mice: Characteristics of Muscle Progenitor Cells and Circulating Factors. Stem Cells 2019, 37, 357–367. [Google Scholar] [CrossRef]
- Jia, Y.; Suzuki, N.; Yamamoto, M.; Gassmann, M.; Noguchi, C.T. Endogenous erythropoietin signaling facilitates skeletal muscle repair and recovery following pharmacologically induced damage. FASEB J. 2012, 26, 2847–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibasaki, H.; Imamura, M.; Arima, S.; Tanihata, J.; Kuraoka, M.; Matsuzaka, Y.; Uchiumi, F.; Tanuma, S.I.; Takeda, S. Characterization of a novel microRNA, miR-188, elevated in serum of muscular dystrophy dog model. PLoS ONE 2019, 14, e0211597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.J.; Shaikh, S.; Ahmad, K.; Ahmad, S.S.; Lim, J.H.; Park, S.; Yang, H.J.; Cho, W.K.; Park, S.J.; Lee, Y.H.; et al. Isolation and Characterization of Compounds from Glycyrrhiza uralensis as Therapeutic Agents for the Muscle Disorders. Int. J. Mol. Sci. 2021, 22, 876. [Google Scholar] [CrossRef] [PubMed]
- Armand, A.-S.; Launay, T.; Pariset, C.; Della Gaspera, B.; Charbonnier, F.; Chanoine, C. Injection of FGF6 accelerates regeneration of the soleus muscle in adult mice. Biochim. Biophys. Acta Mol. Cell Res. 2003, 1642, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Pessemesse, L.; Tintignac, L.; Blanchet, E.; Cortade, F.; Jublanc, E.; Demangel, R.; Py, G.; Sar, C.; Cabello, G.; Wrutniak-Cabello, C.; et al. Regulation of mitochondrial activity controls the duration of skeletal muscle regeneration in response to injury. Sci. Rep. 2019, 9, 12249. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Zhang, D.; Zhao, L.; Li, Y.; Yao, X.; Wang, H.; Zhang, S.; Liu, W.; Cao, H.; Yu, S.; et al. CaMKK2 Suppresses Muscle Regeneration through the Inhibition of Myoblast Proliferation and Differentiation. Int. J. Mol. Sci. 2016, 17, 1695. [Google Scholar] [CrossRef] [Green Version]
- Minetti, G.C.; Feige, J.N.; Bombard, F.; Heier, A.; Morvan, F.; Nurnberg, B.; Leiss, V.; Birnbaumer, L.; Glass, D.J.; Fornaro, M. Galphai2 signaling is required for skeletal muscle growth, regeneration, and satellite cell proliferation and differentiation. Mol. Cell. Biol. 2014, 34, 619–630. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.C.; Han, Y.C.; Cheng, N.X.; DU, J. Effect of arachidonic acid cytochrome P450ω hydroxylase Cyp4a14 gene knockout on skeletal muscle regeneration after injury. Sheng Li Xue Bao 2021, 73, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Chen, S.; Pan, X.; Dai, X.; Pan, G.; Li, Z.; Mai, X.; Tian, Y.; Zhang, S.; Liu, B.; et al. Transferrin receptor 1 ablation in satellite cells impedes skeletal muscle regeneration through activation of ferroptosis. J. Cachexia Sarcopenia Muscle 2021, 12, 746–768. [Google Scholar] [CrossRef] [PubMed]
- Naito, M.; Mori, M.; Inagawa, M.; Miyata, K.; Hashimoto, N.; Tanaka, S.; Asahara, H. Dnmt3a Regulates Proliferation of Muscle Satellite Cells via p57Kip2. PLoS Genet. 2016, 12, e1006167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.H.; Hong, M.; Jeong, H.J.; Kim, H.; Lee, S.J.; Ryu, D.; Bae, G.U.; Cho, S.C.; Lee, Y.S.; Krauss, R.S.; et al. Satellite cell-specific ablation of Cdon impairs integrin activation, FGF signalling, and muscle regeneration. J. Cachexia Sarcopenia Muscle 2020, 11, 1089–1103. [Google Scholar] [CrossRef] [Green Version]
- Rooney, J.E.; Gurpur, P.B.; Yablonka-Reuveni, Z.; Burkin, D.J. Laminin-111 restores regenerative capacity in a mouse model for alpha7 integrin congenital myopathy. Am. J. Pathol. 2009, 174, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Matsumoto, M.; Katoh, Y.; Liu, L.; Ochiai, K.; Aizawa, Y.; Nagatomi, R.; Okuno, H.; Itoi, E.; Igarashi, K. Bach1 promotes muscle regeneration through repressing Smad-mediated inhibition of myoblast differentiation. PLoS ONE 2020, 15, e0236781. [Google Scholar] [CrossRef]
- Yamashita, A.; Hatazawa, Y.; Hirose, Y.; Ono, Y.; Kamei, Y. FOXO1 delays skeletal muscle regeneration and suppresses myoblast proliferation. Biosci. Biotechnol. Biochem. 2016, 80, 1531–1535. [Google Scholar] [CrossRef] [Green Version]
- Girgenrath, M.; Weng, S.; Kostek, C.A.; Browning, B.; Wang, M.; Brown, S.A.; Winkles, J.A.; Michaelson, J.S.; Allaire, N.; Schneider, P.; et al. TWEAK, via its receptor Fn14, is a novel regulator of mesenchymal progenitor cells and skeletal muscle regeneration. EMBO J. 2006, 25, 5826–5839. [Google Scholar] [CrossRef] [Green Version]
- Martinet, C.; Monnier, P.; Louault, Y.; Benard, M.; Gabory, A.; Dandolo, L. H19 controls reactivation of the imprinted gene network during muscle regeneration. Development 2016, 143, 962–971. [Google Scholar] [CrossRef] [Green Version]
- Yahiaoui, L.; Gvozdic, D.; Danialou, G.; Mack, M.; Petrof, B.J. CC family chemokines directly regulate myoblast responses to skeletal muscle injury. J. Physiol. 2008, 586, 3991–4004. [Google Scholar] [CrossRef]
- Kurosaka, M.; Ogura, Y.; Funabashi, T.; Akema, T. Early Growth Response 3 (Egr3) Contributes a Maintenance of C2C12 Myoblast Proliferation. J. Cell. Physiol. 2017, 232, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, N.; Nishizuka, M.; Osada, S.; Imagawa, M. Fad24, a Positive Regulator of Adipogenesis, Is Required for S Phase Re-entry of C2C12 Myoblasts Arrested in G0 Phase and Involved in p27(Kip1) Expression at the Protein Level. Biol. Pharm. Bull. 2016, 39, 807–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanou, N.; Schakman, O.; Louis, P.; Ruegg, U.T.; Dietrich, A.; Birnbaumer, L.; Gailly, P. Trpc1 ion channel modulates phosphatidylinositol 3-kinase/Akt pathway during myoblast differentiation and muscle regeneration. J. Biol. Chem. 2012, 287, 14524–14534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yablonka-Reuveni, Z.; Danoviz, M.E.; Phelps, M.; Stuelsatz, P. Myogenic-specific ablation of Fgfr1 impairs FGF2-mediated proliferation of satellite cells at the myofiber niche but does not abolish the capacity for muscle regeneration. Front. Aging Neurosci. 2015, 7, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Tian, X.; Yan, C.; Liu, D.; Wang, S.; Han, Y. Nicotine promotes the differentiation of C2C12 myoblasts and improves skeletal muscle regeneration in obese mice. Biochem. Biophys. Res. Commun. 2019, 511, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, X.; Liu, S.; Wu, Y.; Zhao, T.; Chen, X.; Zhu, L.; Wu, Y.; Ding, X.; Peng, X.; et al. Sema4C participates in myogenic differentiation in vivo and in vitro through the p38 MAPK pathway. Eur. J. Cell Biol. 2007, 86, 331–344. [Google Scholar] [CrossRef]
- Liu, N.; Williams, A.H.; Maxeiner, J.M.; Bezprozvannaya, S.; Shelton, J.M.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. microRNA-206 promotes skeletal muscle regeneration and delays progression of Duchenne muscular dystrophy in mice. J. Clin. Invest. 2012, 122, 2054–2065. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Wang, J.; Ma, Y.; Yang, Z.; Dong, D.; Li, H.; Yang, J.; Huang, Y.; Plath, M.; Ma, Y.; et al. Linc-smad7 promotes myoblast differentiation and muscle regeneration via sponging miR-125b. Epigenetics 2018, 13, 591–604. [Google Scholar] [CrossRef] [Green Version]
- Gatta, L.; Vitiello, L.; Gorini, S.; Chiandotto, S.; Costelli, P.; Giammarioli, A.M.; Malorni, W.; Rosano, G.; Ferraro, E. Modulating the metabolism by trimetazidine enhances myoblast differentiation and promotes myogenesis in cachectic tumor-bearing c26 mice. Oncotarget 2017, 8, 113938–113956. [Google Scholar] [CrossRef] [Green Version]
- Gagan, J.; Dey, B.K.; Layer, R.; Yan, Z.; Dutta, A. MicroRNA-378 targets the myogenic repressor MyoR during myoblast differentiation. J. Biol. Chem. 2011, 286, 19431–19438. [Google Scholar] [CrossRef] [Green Version]
- Mikami, T.; Koyama, S.; Yabuta, Y.; Kitagawa, H. Chondroitin sulfate is a crucial determinant for skeletal muscle development/regeneration and improvement of muscular dystrophies. J. Biol. Chem. 2012, 287, 38531–38542. [Google Scholar] [CrossRef] [PubMed]
- Storbeck, C.J.; Al-Zahrani, K.N.; Sriram, R.; Kawesa, S.; O’Reilly, P.; Daniel, K.; McKay, M.; Kothary, R.; Tsilfidis, C.; Sabourin, L.A. Distinct roles for Ste20-like kinase SLK in muscle function and regeneration. Skelet. Muscle 2013, 3, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, H.; Leong, D.; Smith, L.R.; Barton, E.R. Matrix metalloproteinase 13 is a new contributor to skeletal muscle regeneration and critical for myoblast migration. Am. J. Physiol. Cell Physiol. 2013, 305, C529–C538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Morvan, F.; Morozzi, G.; Jourde, B.; Minetti, G.C.; Kahle, P.; Rivet, H.; Brebbia, P.; Toussaint, G.; Glass, D.J.; et al. ATP Citrate Lyase Regulates Myofiber Differentiation and Increases Regeneration by Altering Histone Acetylation. Cell Rep. 2017, 21, 3003–3011. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Huq, T.S.; Delafontaine, P. Angiotensin type 2 receptor signaling in satellite cells potentiates skeletal muscle regeneration. J. Biol. Chem. 2014, 289, 26239–26248. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Chen, B.; Ye, M.; Liang, P.; Zhangfang, Y.; Huang, J.; Liu, M.; Songyang, Z.; Ma, W. Ccndbp1 is a new positive regulator of skeletal myogenesis. J. Cell Sci. 2016, 129, 2767–2777. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.E.; Jin, B.; Li, Y.P. TNF-alpha regulates myogenesis and muscle regeneration by activating p38 MAPK. Am. J. Physiol. Cell Physiol. 2007, 292, C1660–C1671. [Google Scholar] [CrossRef]
- He, S.; Fu, T.; Yu, Y.; Liang, Q.; Li, L.; Liu, J.; Zhang, X.; Zhou, Q.; Guo, Q.; Xu, D.; et al. IRE1alpha regulates skeletal muscle regeneration through Myostatin mRNA decay. J. Clin. Invest. 2021, 131, e143737. [Google Scholar] [CrossRef]
- Luca, E.; Turcekova, K.; Hartung, A.; Mathes, S.; Rehrauer, H.; Krutzfeldt, J. Genetic deletion of microRNA biogenesis in muscle cells reveals a hierarchical non-clustered network that controls focal adhesion signaling during muscle regeneration. Mol. Metab. 2020, 36, 100967. [Google Scholar] [CrossRef]
- Esteca, M.V.; Severino, M.B.; Silvestre, J.G.; Palmeira Dos Santos, G.; Tamborlin, L.; Luchessi, A.D.; Moriscot, A.S.; Gustafsson, A.B.; Baptista, I.L. Loss of Parkin Results in Altered Muscle Stem Cell Differentiation during Regeneration. Int. J. Mol. Sci. 2020, 21, 8007. [Google Scholar] [CrossRef]
- Ishii, A.; Lo, S.H. A role of tensin in skeletal-muscle regeneration. Biochem. J. 2001, 356, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Shin, Y.J.; Panda, A.C.; Abdelmohsen, K.; Kim, J.Y.; Lee, S.M.; Bahn, Y.J.; Choi, J.Y.; Kwon, E.S.; Baek, S.J.; et al. miR-431 promotes differentiation and regeneration of old skeletal muscle by targeting Smad4. Genes Dev. 2015, 29, 1605–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerquone Perpetuini, A.; Re Cecconi, A.D.; Chiappa, M.; Martinelli, G.B.; Fuoco, C.; Desiderio, G.; Castagnoli, L.; Gargioli, C.; Piccirillo, R.; Cesareni, G. Group I Paks support muscle regeneration and counteract cancer-associated muscle atrophy. J. Cachexia Sarcopenia Muscle 2018, 9, 727–746. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Hong, Y.; Kong, G.; Lee, D.H.; Kim, M.; Tran, Q.; Cho, H.; Kim, C.; Park, S.; Kim, S.H.; et al. Yin Yang 1 is required for PHD finger protein 20-mediated myogenic differentiation in vitro and in vivo. Cell Death Differ. 2020, 27, 3321–3336. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Won, J.Y.; Yang, J.; Lee, J.; Kim, S.Y.; Lee, E.J.; Kim, H.S. AKAP6 inhibition impairs myoblast differentiation and muscle regeneration: Positive loop between AKAP6 and myogenin. Sci. Rep. 2015, 5, 16523. [Google Scholar] [CrossRef] [Green Version]
- Li, H.K.; Zhou, Y.; Ding, J.; Xiong, L.; Shi, Y.X.; He, Y.J.; Yang, D.; Deng, Z.L.; Nie, M.; Fei Gao, Y. LRTM1 promotes the differentiation of myoblast cells by negatively regulating the FGFR1 signaling pathway. Exp. Cell Res. 2020, 396, 112237. [Google Scholar] [CrossRef]
- Lin, Y.F.; Xiao, M.H.; Chen, H.X.; Meng, Y.; Zhao, N.; Yang, L.; Tang, H.; Wang, J.L.; Liu, X.; Zhu, Y.; et al. A novel mitochondrial micropeptide MPM enhances mitochondrial respiratory activity and promotes myogenic differentiation. Cell Death Dis. 2019, 10, 528. [Google Scholar] [CrossRef] [Green Version]
- Harada, A.; Maehara, K.; Ono, Y.; Taguchi, H.; Yoshioka, K.; Kitajima, Y.; Xie, Y.; Sato, Y.; Iwasaki, T.; Nogami, J.; et al. Histone H3.3 sub-variant H3mm7 is required for normal skeletal muscle regeneration. Nat. Commun. 2018, 9, 1400. [Google Scholar] [CrossRef] [Green Version]
- Miyake, T.; Alli, N.S.; Aziz, A.; Knudson, J.; Fernando, P.; Megeney, L.A.; McDermott, J.C. Cardiotrophin-1 maintains the undifferentiated state in skeletal myoblasts. J. Biol. Chem. 2009, 284, 19679–19693. [Google Scholar] [CrossRef] [Green Version]
- Paul, C.; Sardet, C.; Fabbrizio, E. The histone- and PRMT5-associated protein COPR5 is required for myogenic differentiation. Cell Death Differ. 2012, 19, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Faralli, H.; Martin, E.; Core, N.; Liu, Q.C.; Filippi, P.; Dilworth, F.J.; Caubit, X.; Fasano, L. Teashirt-3, a novel regulator of muscle differentiation, associates with BRG1-associated factor 57 (BAF57) to inhibit myogenin gene expression. J. Biol. Chem. 2011, 286, 23498–23510. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Nelson, B.R.; Bezprozvannaya, S.; Shelton, J.M.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. Requirement of MEF2A, C, and D for skeletal muscle regeneration. Proc. Natl. Acad. Sci. USA 2014, 111, 4109–4114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielbasa, O.M.; Reynolds, J.G.; Wu, C.L.; Snyder, C.M.; Cho, M.Y.; Weiler, H.; Kandarian, S.; Naya, F.J. Myospryn is a calcineurin-interacting protein that negatively modulates slow-fiber-type transformation and skeletal muscle regeneration. FASEB J. 2011, 25, 2276–2286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verpoorten, S.; Sfyri, P.; Scully, D.; Mitchell, R.; Tzimou, A.; Mougios, V.; Patel, K.; Matsakas, A. Loss of CD36 protects against diet-induced obesity but results in impaired muscle stem cell function, delayed muscle regeneration and hepatic steatosis. Acta Physiol. 2020, 228, e13395. [Google Scholar] [CrossRef] [PubMed]
- Andree, B.; Fleige, A.; Arnold, H.H.; Brand, T. Mouse Pop1 is required for muscle regeneration in adult skeletal muscle. Mol. Cell. Biol. 2002, 22, 1504–1512. [Google Scholar] [CrossRef] [Green Version]
- Paolini, A.; Omairi, S.; Mitchell, R.; Vaughan, D.; Matsakas, A.; Vaiyapuri, S.; Ricketts, T.; Rubinsztein, D.C.; Patel, K. Attenuation of autophagy impacts on muscle fibre development, starvation induced stress and fibre regeneration following acute injury. Sci. Rep. 2018, 8, 9062. [Google Scholar] [CrossRef] [Green Version]
- Clow, C.; Jasmin, B.J. Brain-derived neurotrophic factor regulates satellite cell differentiation and skeltal muscle regeneration. Mol. Biol. Cell 2010, 21, 2182–2190. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.L.; Holmberg, J.; Chou, E.; Ocampo, A.C.; Oh, J.; Lee, J.; Peter, A.K.; Martin, P.T.; Crosbie-Watson, R.H. Sarcospan-dependent Akt activation is required for utrophin expression and muscle regeneration. J. Cell Biol. 2012, 197, 1009–1027. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.E.; Gerken, E.; Zhang, Y.; Zhan, M.; Mohan, R.K.; Li, A.S.; Reid, M.B.; Li, Y.P. Role of TNF-{alpha} signaling in regeneration of cardiotoxin-injured muscle. Am. J. Physiol. Cell Physiol. 2005, 289, C1179–C1187. [Google Scholar] [CrossRef]
- Ravel-Chapuis, A.; Crawford, T.E.; Blais-Crepeau, M.L.; Belanger, G.; Richer, C.T.; Jasmin, B.J. The RNA-binding protein Staufen1 impairs myogenic differentiation via a c-myc-dependent mechanism. Mol. Biol. Cell 2014, 25, 3765–3778. [Google Scholar] [CrossRef] [Green Version]
- Langsdorf, A.; Do, A.T.; Kusche-Gullberg, M.; Emerson, C.P., Jr.; Ai, X. Sulfs are regulators of growth factor signaling for satellite cell differentiation and muscle regeneration. Dev. Biol. 2007, 311, 464–477. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Niu, A.; Chen, S.E.; Li, Y.P. Beta3-integrin mediates satellite cell differentiation in regenerating mouse muscle. FASEB J. 2011, 25, 1914–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, S.; Kondo, S.; Hayasaka, M.; Hanaoka, K. Functional analysis of homeodomain-containing transcription factor Lbx1 in satellite cells of mouse skeletal muscle. J. Cell Sci. 2007, 120, 4178–4187. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Lala-Tabbert, N.; Lee, H.; Wiper-Bergeron, N. Mdm2 promotes myogenesis through the ubiquitination and degradation of CCAAT/enhancer-binding protein beta. J. Biol. Chem. 2015, 290, 10200–10207. [Google Scholar] [CrossRef] [Green Version]
- Schroer, A.B.; Mohamed, J.S.; Willard, M.D.; Setola, V.; Oestreich, E.; Siderovski, D.P. A role for Regulator of G protein Signaling-12 (RGS12) in the balance between myoblast proliferation and differentiation. PLoS ONE 2019, 14, e0216167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.; Shin, J.Y.; Jo, A.; Jyothi, K.R.; Nguyen, M.N.; Choi, T.G.; Kim, J.; Park, J.H.; Eun, Y.G.; Yoon, K.S.; et al. Carbonyl reductase 1 is an essential regulator of skeletal muscle differentiation and regeneration. Int. J. Biochem. Cell Biol. 2013, 45, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Mammen, A.L.; Casciola-Rosen, L.A.; Hall, J.C.; Christopher-Stine, L.; Corse, A.M.; Rosen, A. Expression of the dermatomyositis autoantigen Mi-2 in regenerating muscle. Arthritis Rheum. 2009, 60, 3784–3793. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Xu, H.; Xu, Y.; Fan, W.; Yao, H.; Wang, Y.; Hu, W.; Lou, G.; Shi, Y.; Chen, X.; et al. Andrographolide promotes skeletal muscle regeneration after acute injury through epigenetic modulation. Eur. J. Pharmacol. 2020, 888, 173470. [Google Scholar] [CrossRef]
- Kurosaka, M.; Ogura, Y.; Sato, S.; Kohda, K.; Funabashi, T. Transcription factor signal transducer and activator of transcription 6 (STAT6) is an inhibitory factor for adult myogenesis. Skelet. Muscle 2021, 11, 14. [Google Scholar] [CrossRef]
- Vijayakumar, A.; Buffin, N.J.; Gallagher, E.J.; Blank, J.; Wu, Y.; Yakar, S.; LeRoith, D. Deletion of growth hormone receptors in postnatal skeletal muscle of male mice does not alter muscle mass and response to pathological injury. Endocrinology 2013, 154, 3776–3783. [Google Scholar] [CrossRef] [Green Version]
- Pryce, B.R.; Al-Zahrani, K.N.; Dufresne, S.; Belkina, N.; Labreche, C.; Patino-Lopez, G.; Frenette, J.; Shaw, S.; Sabourin, L.A. Deletion of the Ste20-like kinase SLK in skeletal muscle results in a progressive myopathy and muscle weakness. Skelet. Muscle 2017, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Redelsperger, F.; Raddi, N.; Bacquin, A.; Vernochet, C.; Mariot, V.; Gache, V.; Blanchard-Gutton, N.; Charrin, S.; Tiret, L.; Dumonceaux, J.; et al. Genetic Evidence That Captured Retroviral Envelope syncytins Contribute to Myoblast Fusion and Muscle Sexual Dimorphism in Mice. PLoS Genet. 2016, 12, e1006289. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, P.; Ilencikova, K.; Zikova, M.; Horvath, O.; Cermak, V.; Bartunek, P.; Strnad, H. c-Myb inhibits myoblast fusion. PLoS ONE 2013, 8, e76742. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.A.; Johnson, R.W.; Whitlock, J.M.; Pozsgai, E.R.; Heller, K.N.; Grose, W.E.; Arnold, W.D.; Sahenk, Z.; Hartzell, H.C.; Rodino-Klapac, L.R. Defective membrane fusion and repair in Anoctamin5-deficient muscular dystrophy. Hum. Mol. Genet. 2016, 25, 1900–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapani, L.; Segatto, M.; La Rosa, P.; Fanelli, F.; Moreno, S.; Marino, M.; Pallottini, V. 3-hydroxy 3-methylglutaryl coenzyme A reductase inhibition impairs muscle regeneration. J. Cell. Biochem. 2012, 113, 2057–2063. [Google Scholar] [CrossRef] [PubMed]
- Hamoud, N.; Tran, V.; Aimi, T.; Kakegawa, W.; Lahaie, S.; Thibault, M.P.; Pelletier, A.; Wong, G.W.; Kim, I.S.; Kania, A.; et al. Spatiotemporal regulation of the GPCR activity of BAI3 by C1qL4 and Stabilin-2 controls myoblast fusion. Nat. Commun. 2018, 9, 4470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamilarasan, K.P.; Temmel, H.; Das, S.K.; Al Zoughbi, W.; Schauer, S.; Vesely, P.W.; Hoefler, G. Skeletal muscle damage and impaired regeneration due to LPL-mediated lipotoxicity. Cell Death Dis. 2012, 3, e354. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Endo, J.; Kinouchi, K.; Kitakata, H.; Moriyama, H.; Kataoka, M.; Yamamoto, T.; Shirakawa, K.; Morimoto, S.; Nishiyama, A.; et al. (Pro)renin receptor accelerates development of sarcopenia via activation of Wnt/YAP signaling axis. Aging Cell 2019, 18, e12991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youm, T.H.; Woo, S.H.; Kwon, E.S.; Park, S.S. NADPH Oxidase 4 Contributes to Myoblast Fusion and Skeletal Muscle Regeneration. Oxid. Med. Cell. Longev. 2019, 2019, 3585390. [Google Scholar] [CrossRef] [Green Version]
- Teng, S.; Stegner, D.; Chen, Q.; Hongu, T.; Hasegawa, H.; Chen, L.; Kanaho, Y.; Nieswandt, B.; Frohman, M.A.; Huang, P. Phospholipase D1 facilitates second-phase myoblast fusion and skeletal muscle regeneration. Mol. Biol. Cell 2015, 26, 506–517. [Google Scholar] [CrossRef]
- Krause, M.P.; Moradi, J.; Coleman, S.K.; D’Souza, D.M.; Liu, C.; Kronenberg, M.S.; Rowe, D.W.; Hawke, T.J.; Hadjiargyrou, M. A novel GFP reporter mouse reveals Mustn1 expression in adult regenerating skeletal muscle, activated satellite cells and differentiating myoblasts. Acta Physiol. 2013, 208, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Kurosaka, M.; Ogura, Y.; Funabashi, T.; Akema, T. Involvement of Transient Receptor Potential Cation Channel Vanilloid 1 (TRPV1) in Myoblast Fusion. J. Cell. Physiol. 2016, 231, 2275–2285. [Google Scholar] [CrossRef] [PubMed]
- Singhal, N.; Martin, P.T. A role for Galgt1 in skeletal muscle regeneration. Skelet. Muscle 2015, 5, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yalvac, M.E.; Amornvit, J.; Braganza, C.; Chen, L.; Hussain, S.A.; Shontz, K.M.; Montgomery, C.L.; Flanigan, K.M.; Lewis, S.; Sahenk, Z. Impaired regeneration in calpain-3 null muscle is associated with perturbations in mTORC1 signaling and defective mitochondrial biogenesis. Skelet. Muscle 2017, 7, 27. [Google Scholar] [CrossRef]
- Ogawa, R.; Ma, Y.; Yamaguchi, M.; Ito, T.; Watanabe, Y.; Ohtani, T.; Murakami, S.; Uchida, S.; De Gaspari, P.; Uezumi, A.; et al. Doublecortin marks a new population of transiently amplifying muscle progenitor cells and is required for myofiber maturation during skeletal muscle regeneration. Development 2015, 142, 810. [Google Scholar] [CrossRef] [Green Version]
- Ohno, Y.; Ando, K.; Ito, T.; Suda, Y.; Matsui, Y.; Oyama, A.; Kaneko, H.; Yokoyama, S.; Egawa, T.; Goto, K. Lactate Stimulates a Potential for Hypertrophy and Regeneration of Mouse Skeletal Muscle. Nutrients 2019, 11, 869. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, S.; Sakamoto, K.; Vassilopoulos, S.; Camus, S.M.; Griffin, C.A.; Esk, C.; Torres, J.A.; Ohkoshi, N.; Ishii, A.; Tamaoka, A.; et al. The CHC22 clathrin-GLUT4 transport pathway contributes to skeletal muscle regeneration. PLoS ONE 2013, 8, e77787. [Google Scholar] [CrossRef]
- Agbulut, O.; Li, Z.; Perie, S.; Ludosky, M.A.; Paulin, D.; Cartaud, J.; Butler-Browne, G. Lack of desmin results in abortive muscle regeneration and modifications in synaptic structure. Cell Motil. Cytoskelet. 2001, 49, 51–66. [Google Scholar] [CrossRef]
- Cicchillitti, L.; Di Stefano, V.; Isaia, E.; Crimaldi, L.; Fasanaro, P.; Ambrosino, V.; Antonini, A.; Capogrossi, M.C.; Gaetano, C.; Piaggio, G.; et al. Hypoxia-inducible factor 1-alpha induces miR-210 in normoxic differentiating myoblasts. J. Biol. Chem. 2012, 287, 44761–44771. [Google Scholar] [CrossRef] [Green Version]
- Piccioni, A.; Gaetani, E.; Neri, V.; Gatto, I.; Palladino, M.; Silver, M.; Smith, R.C.; Giarretta, I.; Pola, E.; Hlatky, L.; et al. Sonic hedgehog therapy in a mouse model of age-associated impairment of skeletal muscle regeneration. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Garry, G.A.; Li, S.; Bezprozvannaya, S.; Sanchez-Ortiz, E.; Chen, B.; Shelton, J.M.; Jaichander, P.; Bassel-Duby, R.; Olson, E.N. A Twist2-dependent progenitor cell contributes to adult skeletal muscle. Nat. Cell Biol. 2017, 19, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Shaikh, S.; Baig, M.H.; Park, S.Y.; Lim, J.H.; Ahmad, S.S.; Ali, S.; Ahmad, K.; Choi, I. MIF1 and MIF2 Myostatin Peptide Inhibitors as Potent Muscle Mass Regulators. Int. J. Mol. Sci. 2022, 23, 4222. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kay, D.I.; Rudra, R.T.; Chen, B.M.; Hsu, N.; Izumiya, Y.; Martinez, L.; Spencer, M.J.; Walsh, K.; Grinnell, A.D.; et al. Myogenic Akt signaling attenuates muscular degeneration, promotes myofiber regeneration and improves muscle function in dystrophin-deficient mdx mice. Hum. Mol. Genet. 2011, 20, 1324–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Nie, M.; Liu, J.; Hu, X.; Ma, L.; Deng, Z.L.; Wang, D.Z. Trbp Is Required for Differentiation of Myoblasts and Normal Regeneration of Skeletal Muscle. PLoS ONE 2016, 11, e0155349. [Google Scholar] [CrossRef] [Green Version]
- Murray, I.R.; Gonzalez, Z.N.; Baily, J.; Dobie, R.; Wallace, R.J.; Mackinnon, A.C.; Smith, J.R.; Greenhalgh, S.N.; Thompson, A.I.; Conroy, K.P.; et al. alphav integrins on mesenchymal cells regulate skeletal and cardiac muscle fibrosis. Nat. Commun. 2017, 8, 1118. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Son, J.S.; Wang, B.; Tian, Q.; Chen, Y.; Liu, X.; de Avila, J.M.; Zhu, M.J.; Du, M. Retinoic acid signalling in fibro/adipogenic progenitors robustly enhances muscle regeneration. EBioMedicine 2020, 60, 103020. [Google Scholar] [CrossRef]
- Zanotti, S.; Gibertini, S.; Blasevich, F.; Bragato, C.; Ruggieri, A.; Saredi, S.; Fabbri, M.; Bernasconi, P.; Maggi, L.; Mantegazza, R.; et al. Exosomes and exosomal miRNAs from muscle-derived fibroblasts promote skeletal muscle fibrosis. Matrix Biol. 2018, 74, 77–100. [Google Scholar] [CrossRef]
- Vumbaca, S.; Giuliani, G.; Fiorentini, V.; Tortolici, F.; Cerquone Perpetuini, A.; Riccio, F.; Sennato, S.; Gargioli, C.; Fuoco, C.; Castagnoli, L.; et al. Characterization of the Skeletal Muscle Secretome Reveals a Role for Extracellular Vesicles and IL1alpha/IL1beta in Restricting Fibro/Adipogenic Progenitor Adipogenesis. Biomolecules 2021, 11, 1171. [Google Scholar] [CrossRef]
- Stepien, D.M.; Hwang, C.; Marini, S.; Pagani, C.A.; Sorkin, M.; Visser, N.D.; Huber, A.K.; Edwards, N.J.; Loder, S.J.; Vasquez, K.; et al. Tuning Macrophage Phenotype to Mitigate Skeletal Muscle Fibrosis. J. Immunol. 2020, 204, 2203–2215. [Google Scholar] [CrossRef]
- Burks, T.N.; Andres-Mateos, E.; Marx, R.; Mejias, R.; Van Erp, C.; Simmers, J.L.; Walston, J.D.; Ward, C.W.; Cohn, R.D. Losartan restores skeletal muscle remodeling and protects against disuse atrophy in sarcopenia. Sci. Transl. Med. 2011, 3, 82ra37. [Google Scholar] [CrossRef] [Green Version]
- Bosnakovski, D.; Oyler, D.; Mitanoska, A.; Douglas, M.; Ener, E.T.; Shams, A.S.; Kyba, M. Persistent Fibroadipogenic Progenitor Expansion Following Transient DUX4 Expression Provokes a Profibrotic State in a Mouse Model for FSHD. Int. J. Mol. Sci. 2022, 23, 1983. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, S.; Cheng, X.W.; Inoue, A.; Hu, L.; Piao, L.; Yu, C.; Goto, H.; Xu, W.; Zhao, G.; Lei, Y.; et al. Cathepsin K activity controls cardiotoxin-induced skeletal muscle repair in mice. J. Cachexia Sarcopenia Muscle 2018, 9, 160–175. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lee, Y.S.; Zimmers, T.A.; Soleimani, A.; Matzuk, M.M.; Tsuchida, K.; Cohn, R.D.; Barton, E.R. Regulation of muscle mass by follistatin and activins. Mol. Endocrinol. 2010, 24, 1998–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldi, F.; Zhang, Y.; Mondragon-Gonzalez, R.; Harvey, J.; Perlingeiro, R.C.R. Treatment with rGDF11 does not improve the dystrophic muscle pathology of mdx mice. Skelet. Muscle 2016, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mignemi, N.A.; Yuasa, M.; Baker, C.E.; Moore, S.N.; Ihejirika, R.C.; Oelsner, W.K.; Wallace, C.S.; Yoshii, T.; Okawa, A.; Revenko, A.S.; et al. Plasmin Prevents Dystrophic Calcification After Muscle Injury. J. Bone Miner. Res. 2017, 32, 294–308. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, S.J. Regulation of GDF-11 and myostatin activity by GASP-1 and GASP-2. Proc. Natl. Acad. Sci. USA 2013, 110, E3713–E3722. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Urganus, A.L.; Spevak, L.; Shrestha, S.; Doty, S.B.; Boskey, A.L.; Pachman, L.M. Characterization of dystrophic calcification induced in mice by cardiotoxin. Calcif. Tissue Int. 2009, 85, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Lounev, V.Y.; Ramachandran, R.; Wosczyna, M.N.; Yamamoto, M.; Maidment, A.D.; Shore, E.M.; Glaser, D.L.; Goldhamer, D.J.; Kaplan, F.S. Identification of progenitor cells that contribute to heterotopic skeletogenesis. J. Bone Joint Surg. Am. 2009, 91, 652–663. [Google Scholar] [CrossRef] [Green Version]
- Drouin, G.; Couture, V.; Lauzon, M.A.; Balg, F.; Faucheux, N.; Grenier, G. Muscle injury-induced hypoxia alters the proliferation and differentiation potentials of muscle resident stromal cells. Skelet. Muscle 2019, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Arsic, N.; Zacchigna, S.; Zentilin, L.; Ramirez-Correa, G.; Pattarini, L.; Salvi, A.; Sinagra, G.; Giacca, M. Vascular endothelial growth factor stimulates skeletal muscle regeneration in vivo. Mol. Ther. 2004, 10, 844–854. [Google Scholar] [CrossRef]
- Sinha-Hikim, I.; Braga, M.; Shen, R.; Sinha Hikim, A.P. Involvement of c-Jun NH2-terminal kinase and nitric oxide-mediated mitochondria-dependent intrinsic pathway signaling in cardiotoxin-induced muscle cell death: Role of testosterone. Apoptosis 2007, 12, 1965–1978. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Lawan, A.; Bennett, A.M. Loss of MKP-5 promotes myofiber survival by activating STAT3/Bcl-2 signaling during regenerative myogenesis. Skelet. Muscle 2017, 7, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tjondrokoesoemo, A.; Schips, T.; Kanisicak, O.; Sargent, M.A.; Molkentin, J.D. Genetic overexpression of Serpina3n attenuates muscular dystrophy in mice. Hum. Mol. Genet. 2016, 25, 1192–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, S.N.; Hawley, G.D.; Smith, E.N.; Mignemi, N.A.; Ihejirika, R.C.; Yuasa, M.; Cates, J.M.; Liu, X.; Schoenecker, J.G. Validation of a Radiography-Based Quantification Designed to Longitudinally Monitor Soft Tissue Calcification in Skeletal Muscle. PLoS ONE 2016, 11, e0159624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ieronimakis, N.; Hays, A.; Reyes, M. Bone marrow-derived cells do not engraft into skeletal muscle microvasculature but promote angiogenesis after acute injury. Exp. Hematol. 2012, 40, 238–249.e233. [Google Scholar] [CrossRef] [Green Version]
- Bellamy, L.M.; Johnston, A.P.; De Lisio, M.; Parise, G. Skeletal muscle-endothelial cell cross talk through angiotensin II. Am. J. Physiol. Cell Physiol. 2010, 299, C1402–C1408. [Google Scholar] [CrossRef] [Green Version]
- Mellows, B.; Mitchell, R.; Antonioli, M.; Kretz, O.; Chambers, D.; Zeuner, M.T.; Denecke, B.; Musante, L.; Ramachandra, D.L.; Debacq-Chainiaux, F.; et al. Protein and Molecular Characterization of a Clinically Compliant Amniotic Fluid Stem Cell-Derived Extracellular Vesicle Fraction Capable of Accelerating Muscle Regeneration Through Enhancement of Angiogenesis. Stem Cells Dev. 2017, 26, 1316–1333. [Google Scholar] [CrossRef] [Green Version]
- Hosaka, Y.; Yokota, T.; Miyagoe-Suzuki, Y.; Yuasa, K.; Imamura, M.; Matsuda, R.; Ikemoto, T.; Kameya, S.; Takeda, S. Alpha1-syntrophin-deficient skeletal muscle exhibits hypertrophy and aberrant formation of neuromuscular junctions during regeneration. J. Cell Biol. 2002, 158, 1097–1107. [Google Scholar] [CrossRef] [Green Version]
- Daneshvar, N.; Tatsumi, R.; Peeler, J.; Anderson, J.E. Premature satellite cell activation before injury accelerates myogenesis and disrupts neuromuscular junction maturation in regenerating muscle. Am. J. Physiol. Cell Physiol. 2020, 319, C116–C128. [Google Scholar] [CrossRef]
- Sawano, S.; Suzuki, T.; Do, M.K.; Ohtsubo, H.; Mizunoya, W.; Ikeuchi, Y.; Tatsumi, R. Supplementary immunocytochemistry of hepatocyte growth factor production in activated macrophages early in muscle regeneration. Anim. Sci. J. 2014, 85, 994–1000. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Shono, J.; Suzuki, T.; Sawano, S.; Anderson, J.E.; Do, M.K.; Ohtsubo, H.; Mizunoya, W.; Sato, Y.; Nakamura, M.; et al. Implication of anti-inflammatory macrophages in regenerative moto-neuritogenesis: Promotion of myoblast migration and neural chemorepellent semaphorin 3A expression in injured muscle. Int. J. Biochem. Cell Biol. 2014, 54, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Cha, H.N.; Jo, H.J.; Song, I.H.; Baek, S.H.; Dan, J.M.; Kim, Y.W.; Kim, J.Y.; Lee, I.K.; Seo, J.S.; et al. TLR2 deficiency attenuates skeletal muscle atrophy in mice. Biochem. Biophys. Res. Commun. 2015, 459, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, S.; Lee, M.; Akimoto, T. Conditional Deletion of Dicer in Adult Mice Impairs Skeletal Muscle Regeneration. Int. J. Mol. Sci. 2019, 20, 5686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiramuki, Y.; Sato, T.; Furuta, Y.; Surani, M.A.; Sehara-Fujisawa, A. Mest but Not MiR-335 Affects Skeletal Muscle Growth and Regeneration. PLoS ONE 2015, 10, e0130436. [Google Scholar] [CrossRef] [Green Version]
- Norton, C.R.; Chen, Y.; Han, X.H.; Bradley, C.K.; Krebs, L.T.; Yoon, J.K.; Gridley, T. Absence of a major role for the snai1 and snai3 genes in regulating skeletal muscle regeneration in mice. PLoS Curr. 2013, 5. [Google Scholar] [CrossRef]
- Call, J.A.; Wilson, R.J.; Laker, R.C.; Zhang, M.; Kundu, M.; Yan, Z. Ulk1-mediated autophagy plays an essential role in mitochondrial remodeling and functional regeneration of skeletal muscle. Am. J. Physiol. Cell Physiol. 2017, 312, C724–C732. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, N.; Ahmad, K.; Yadav, B.S.; Lee, E.J.; Sonkar, S.C.; Marina, N.; Choi, I. Understanding Calcium-Dependent Conformational Changes in S100A1 Protein: A Combination of Molecular Dynamics and Gene Expression Study in Skeletal Muscle. Cells 2020, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- Parks, C.A.; Pak, K.; Pinal-Fernandez, I.; Huang, W.; Derfoul, A.; Mammen, A.L. Trim33 (Tif1gamma) is not required for skeletal muscle development or regeneration but suppresses cholecystokinin expression. Sci. Rep. 2019, 9, 18507. [Google Scholar] [CrossRef] [Green Version]
- Goetsch, S.C.; Martin, C.M.; Embree, L.J.; Garry, D.J. Myogenic progenitor cells express filamin C in developing and regenerating skeletal muscle. Stem Cells Dev. 2005, 14, 181–187. [Google Scholar] [CrossRef]
- Wardrop, K.E.; Dominov, J.A. Proinflammatory signals and the loss of lymphatic vessel hyaluronan receptor-1 (LYVE-1) in the early pathogenesis of laminin alpha2-deficient skeletal muscle. J. Histochem. Cytochem. 2011, 59, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Merkulova, T.; Dehaupas, M.; Nevers, M.C.; Creminon, C.; Alameddine, H.; Keller, A. Differential modulation of alpha, beta and gamma enolase isoforms in regenerating mouse skeletal muscle. Eur. J. Biochem. 2000, 267, 3735–3743. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, K.; Hagiwara, Y.; Ando, M.; Nakamura, A.; Takeda, S.; Hijikata, T. MicroRNA-206 is highly expressed in newly formed muscle fibers: Implications regarding potential for muscle regeneration and maturation in muscular dystrophy. Cell Struct. Funct. 2008, 33, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casciola-Rosen, L.; Hall, J.C.; Mammen, A.L.; Christopher-Stine, L.; Rosen, A. Isolated elevation of aldolase in the serum of myositis patients: A potential biomarker of damaged early regenerating muscle cells. Clin. Exp. Rheumatol. 2012, 30, 548–553. [Google Scholar]
- Mammen, A.L.; Mahoney, J.A.; St Germain, A.; Badders, N.; Taylor, J.P.; Rosen, A.; Spinette, S. A novel conserved isoform of the ubiquitin ligase UFD2a/UBE4B is expressed exclusively in mature striated muscle cells. PLoS ONE 2011, 6, e28861. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Tsukamoto, Y.; Hijiya, N.; Higuchi, Y.; Yano, S.; Yokoyama, S.; Kumamoto, T.; Moriyama, M. Induction of GNE in myofibers after muscle injury. Pathobiology 2010, 77, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Do, M.K.; Suzuki, T.; Ohtsubo, H.; Mizunoya, W.; Nakamura, M.; Furuse, M.; Ikeuchi, Y.; Tatsumi, R. Satellite cells produce neural chemorepellent semaphorin 3A upon muscle injury. Anim. Sci. J. 2013, 84, 185–189. [Google Scholar] [CrossRef]
- Garry, D.J.; Meeson, A.; Elterman, J.; Zhao, Y.; Yang, P.; Bassel-Duby, R.; Williams, R.S. Myogenic stem cell function is impaired in mice lacking the forkhead/winged helix protein MNF. Proc. Natl. Acad. Sci. USA 2000, 97, 5416–5421. [Google Scholar] [CrossRef] [Green Version]
- Kemp, M.W.; Edwards, B.; Burgess, M.; Clarke, W.T.; Nicholson, G.; Parry, D.A.; Davies, K.E. Syncoilin isoform organization and differential expression in murine striated muscle. J. Struct. Biol. 2009, 165, 196–203. [Google Scholar] [CrossRef]
- Miura, P.; Thompson, J.; Chakkalakal, J.V.; Holcik, M.; Jasmin, B.J. The utrophin A 5’-untranslated region confers internal ribosome entry site-mediated translational control during regeneration of skeletal muscle fibers. J. Biol. Chem. 2005, 280, 32997–33005. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Qiu, X.; Liu, T.; Ahn, C.; Horowitz, J.F.; Lin, J.D. The hepatokine TSK maintains myofiber integrity and exercise endurance and contributes to muscle regeneration. JCI Insight 2022, 7, e154746. [Google Scholar] [CrossRef]
- McCullagh, K.J.; Edwards, B.; Kemp, M.W.; Giles, L.C.; Burgess, M.; Davies, K.E. Analysis of skeletal muscle function in the C57BL6/SV129 syncoilin knockout mouse. Mamm. Genome 2008, 19, 339–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demonbreun, A.R.; Lapidos, K.A.; Heretis, K.; Levin, S.; Dale, R.; Pytel, P.; Svensson, E.C.; McNally, E.M. Myoferlin regulation by NFAT in muscle injury, regeneration and repair. J. Cell Sci. 2010, 123, 2413–2422. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Yonemochi, Y.; Nakajyo, Y.; Hidaka, H.; Ikeda, T.; Ando, Y. CXCL12 and osteopontin from bone marrow-derived mesenchymal stromal cells improve muscle regeneration. Sci. Rep. 2017, 7, 3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darabi, R.; Gehlbach, K.; Bachoo, R.M.; Kamath, S.; Osawa, M.; Kamm, K.E.; Kyba, M.; Perlingeiro, R.C. Functional skeletal muscle regeneration from differentiating embryonic stem cells. Nat. Med. 2008, 14, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Di Rocco, A.; Uchibe, K.; Larmour, C.; Berger, R.; Liu, M.; Barton, E.R.; Iwamoto, M. Selective Retinoic Acid Receptor gamma Agonists Promote Repair of Injured Skeletal Muscle in Mouse. Am. J. Pathol. 2015, 185, 2495–2504. [Google Scholar] [CrossRef]
- Bryer, S.C.; Koh, T.J. The urokinase-type plasminogen activator receptor is not required for skeletal muscle inflammation or regeneration. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1152–R1158. [Google Scholar] [CrossRef]
- Bryan, B.A.; Mitchell, D.C.; Zhao, L.; Ma, W.; Stafford, L.J.; Teng, B.B.; Liu, M. Modulation of muscle regeneration, myogenesis, and adipogenesis by the Rho family guanine nucleotide exchange factor GEFT. Mol. Cell. Biol. 2005, 25, 11089–11101. [Google Scholar] [CrossRef] [Green Version]
- Mathes, A.L.; Lafyatis, R. Role for Toll-like receptor 3 in muscle regeneration after cardiotoxin injury. Muscle Nerve 2011, 43, 733–740. [Google Scholar] [CrossRef]
- Wu, G.; Sher, R.B.; Cox, G.A.; Vance, D.E. Differential expression of choline kinase isoforms in skeletal muscle explains the phenotypic variability in the rostrocaudal muscular dystrophy mouse. Biochim. Biophys. Acta 2010, 1801, 446–454. [Google Scholar] [CrossRef]
- Fujita, R.; Kawano, F.; Ohira, T.; Nakai, N.; Shibaguchi, T.; Nishimoto, N.; Ohira, Y. Anti-interleukin-6 receptor antibody (MR16-1) promotes muscle regeneration via modulation of gene expressions in infiltrated macrophages. Biochim. Biophys. Acta 2014, 1840, 3170–3180. [Google Scholar] [CrossRef]
- Wu, G.; Sher, R.B.; Cox, G.A.; Vance, D.E. Understanding the muscular dystrophy caused by deletion of choline kinase beta in mice. Biochim. Biophys. Acta 2009, 1791, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Wada, E.; Kato, M.; Yamashita, K.; Kokuba, H.; Liang, W.C.; Bonne, G.; Hayashi, Y.K. Deficiency of emerin contributes differently to the pathogenesis of skeletal and cardiac muscles in LmnaH222P/H222P mutant mice. PLoS ONE 2019, 14, e0221512. [Google Scholar] [CrossRef] [PubMed]
- Mofarrahi, M.; McClung, J.M.; Kontos, C.D.; Davis, E.C.; Tappuni, B.; Moroz, N.; Pickett, A.E.; Huck, L.; Harel, S.; Danialou, G.; et al. Angiopoietin-1 enhances skeletal muscle regeneration in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R576–R589. [Google Scholar] [CrossRef] [Green Version]
- Gattazzo, F.; Molon, S.; Morbidoni, V.; Braghetta, P.; Blaauw, B.; Urciuolo, A.; Bonaldo, P. Cyclosporin A Promotes in vivo Myogenic Response in Collagen VI-Deficient Myopathic Mice. Front. Aging Neurosci. 2014, 6, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laziz, I.; Armand, A.S.; Pariset, C.; Lecolle, S.; Della Gaspera, B.; Charbonnier, F.; Chanoine, C. Sprouty gene expression is regulated by nerve and FGF6 during regeneration of mouse muscles. Growth Factors 2007, 25, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Armand, A.S.; Della Gaspera, B.; Launay, T.; Charbonnier, F.; Gallien, C.L.; Chanoine, C. Expression and neural control of follistatin versus myostatin genes during regeneration of mouse soleus. Dev. Dyn. 2003, 227, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gao, H.L.; Shimokawa, T.; Nabeka, H.; Hamada, F.; Araki, H.; Cao, Y.M.; Kobayashi, N.; Matsuda, S. Prosaposin expression in the regenerated muscles of mdx and cardiotoxin-treated mice. Histol. Histopathol. 2013, 28, 875–892. [Google Scholar] [CrossRef]
- Cizkova, D.; Vavrova, J.; Micuda, S.; Filip, S.; Brcakova, E.; Bruckova, L.; Mokry, J. Role of transplanted bone marrow cells in response to skeletal muscle injury. Folia Biol. 2011, 57, 232–241. [Google Scholar]
- Wernig, G.; Janzen, V.; Schafer, R.; Zweyer, M.; Knauf, U.; Hoegemeier, O.; Mundegar, R.R.; Garbe, S.; Stier, S.; Franz, T.; et al. The vast majority of bone-marrow-derived cells integrated into mdx muscle fibers are silent despite long-term engraftment. Proc. Natl. Acad. Sci. USA 2005, 102, 11852–11857. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.E.; Song, M.J.; Shin, S.; Choi, Y.J.; Kim, K.H.; Chung, C.J. Local myogenic pulp-derived cell injection enhances craniofacial muscle regeneration in vivo. Orthod. Craniofac. Res. 2017, 20, 35–43. [Google Scholar] [CrossRef]
- de la Garza-Rodea, A.S.; van der Velde, I.; Boersma, H.; Goncalves, M.A.; van Bekkum, D.W.; de Vries, A.A.; Knaan-Shanzer, S. Long-term contribution of human bone marrow mesenchymal stromal cells to skeletal muscle regeneration in mice. Cell Transplant 2011, 20, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Mori, J.; Ishihara, Y.; Matsuo, K.; Nakajima, H.; Terada, N.; Kosaka, K.; Kizaki, Z.; Sugimoto, T. Hematopoietic contribution to skeletal muscle regeneration in acid alpha-glucosidase knockout mice. J. Histochem. Cytochem. 2008, 56, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, J.; Dumont, N.; Lebel, C.; Quenneville, S.P.; Cote, C.H.; Frenette, J.; Tremblay, J.P. Dystrophin expression following the transplantation of normal muscle precursor cells protects mdx muscle from contraction-induced damage. Cell Transplant 2010, 19, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Gang, E.J.; Darabi, R.; Bosnakovski, D.; Xu, Z.; Kamm, K.E.; Kyba, M.; Perlingeiro, R.C. Engraftment of mesenchymal stem cells into dystrophin-deficient mice is not accompanied by functional recovery. Exp. Cell Res. 2009, 315, 2624–2636. [Google Scholar] [CrossRef] [PubMed]
- Kano, K.; Horiuchi, K.; Yoshida, Y.; Hayasaka, T.; Kabara, M.; Tomita, Y.; Tatsukawa, T.; Matsuo, R.; Sawada, J.; Nakagawa, N.; et al. EphA7(+) perivascular cells as myogenic and angiogenic precursors improving skeletal muscle regeneration in a muscular dystrophic mouse model. Stem Cell Res. 2020, 47, 101914. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, X.; Sun, Z.; Chen, B.; Han, Q.; Li, J.; Zhao, R.C. Flk-1+ adipose-derived mesenchymal stem cells differentiate into skeletal muscle satellite cells and ameliorate muscular dystrophy in mdx mice. Stem Cells Dev. 2007, 16, 695–706. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, S.; Zhou, J.; Chen, B.; Shang, Y.; Gao, T.; Wang, X.; Xie, H.; Chen, F. Clone-derived human AF-amniotic fluid stem cells are capable of skeletal myogenic differentiation in vitro and in vivo. J. Tissue Eng. Regen. Med. 2012, 6, 598–613. [Google Scholar] [CrossRef]
- Kim, J.A.; Shon, Y.H.; Lim, J.O.; Yoo, J.J.; Shin, H.I.; Park, E.K. MYOD mediates skeletal myogenic differentiation of human amniotic fluid stem cells and regeneration of muscle injury. Stem Cell Res. Ther. 2013, 4, 147. [Google Scholar] [CrossRef] [Green Version]
- Drapeau, C.; Antarr, D.; Ma, H.; Yang, Z.; Tang, L.; Hoffman, R.M.; Schaeffer, D.J. Mobilization of bone marrow stem cells with StemEnhance improves muscle regeneration in cardiotoxin-induced muscle injury. Cell Cycle 2010, 9, 1819–1823. [Google Scholar] [CrossRef] [Green Version]
- Xuan, W.; Khan, M.; Ashraf, M. Pluripotent stem cell-induced skeletal muscle progenitor cells with givinostat promote myoangiogenesis and restore dystrophin in injured Duchenne dystrophic muscle. Stem Cell Res. Ther. 2021, 12, 131. [Google Scholar] [CrossRef]
- Naldaiz-Gastesi, N.; Goicoechea, M.; Aragon, I.M.; Perez-Lopez, V.; Fuertes-Alvarez, S.; Herrera-Imbroda, B.; Lopez de Munain, A.; de Luna-Diaz, R.; Baptista, P.M.; Fernandez, M.A.; et al. Isolation and characterization of myogenic precursor cells from human cremaster muscle. Sci. Rep. 2019, 9, 3454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abedi, M.; Foster, B.M.; Wood, K.D.; Colvin, G.A.; McLean, S.D.; Johnson, K.W.; Greer, D.A. Haematopoietic stem cells participate in muscle regeneration. Br. J. Haematol. 2007, 138, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.; Suk, S.; Shih, Y.R.; Seo, T.; Du, B.; Xie, Y.; Li, Z.; Varghese, S. WNT3A promotes myogenesis of human embryonic stem cells and enhances in vivo engraftment. Sci. Rep. 2014, 4, 5916. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, M.; Franzin, C.; Bertin, E.; Urbani, L.; Blaauw, B.; Repele, A.; Taschin, E.; Cenedese, A.; Zanon, G.F.; Andre-Schmutz, I.; et al. Amniotic fluid stem cells restore the muscle cell niche in a HSA-Cre, Smn(F7/F7) mouse model. Stem Cells 2012, 30, 1675–1684. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Chen, M.; Lee, C.H.; Yoon, R.; Lal, S.; Mao, J.J. Clones of ectopic stem cells in the regeneration of muscle defects in vivo. PLoS ONE 2010, 5, e13547. [Google Scholar] [CrossRef]
- Bossolasco, P.; Corti, S.; Strazzer, S.; Borsotti, C.; Del Bo, R.; Fortunato, F.; Salani, S.; Quirici, N.; Bertolini, F.; Gobbi, A.; et al. Skeletal muscle differentiation potential of human adult bone marrow cells. Exp. Cell Res. 2004, 295, 66–78. [Google Scholar] [CrossRef]
- Fukada, S.; Miyagoe-Suzuki, Y.; Tsukihara, H.; Yuasa, K.; Higuchi, S.; Ono, S.; Tsujikawa, K.; Takeda, S.; Yamamoto, H. Muscle regeneration by reconstitution with bone marrow or fetal liver cells from green fluorescent protein-gene transgenic mice. J. Cell Sci. 2002, 115, 1285–1293. [Google Scholar] [CrossRef]
- Luth, E.S.; Jun, S.J.; Wessen, M.K.; Liadaki, K.; Gussoni, E.; Kunkel, L.M. Bone marrow side population cells are enriched for progenitors capable of myogenic differentiation. J. Cell Sci. 2008, 121, 1426–1434. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.K.; Wang, Y.; Karandikar, A.; Wang, Q.; Gai, H.; Liu, A.L.; Peng, C.; Sheng, H.Z. Skeletal myogenesis by human embryonic stem cells. Cell Res. 2006, 16, 713–722. [Google Scholar] [CrossRef]
- Meeson, A.P.; Hawke, T.J.; Graham, S.; Jiang, N.; Elterman, J.; Hutcheson, K.; Dimaio, J.M.; Gallardo, T.D.; Garry, D.J. Cellular and molecular regulation of skeletal muscle side population cells. Stem Cells 2004, 22, 1305–1320. [Google Scholar] [CrossRef]
- Kowalski, K.; Archacki, R.; Archacka, K.; Streminska, W.; Paciorek, A.; Golabek, M.; Ciemerych, M.A.; Brzoska, E. Stromal derived factor-1 and granulocyte-colony stimulating factor treatment improves regeneration of Pax7-/- mice skeletal muscles. J. Cachexia Sarcopenia Muscle 2016, 7, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.; Mellows, B.; Sheard, J.; Antonioli, M.; Kretz, O.; Chambers, D.; Zeuner, M.T.; Tomkins, J.E.; Denecke, B.; Musante, L.; et al. Secretome of adipose-derived mesenchymal stem cells promotes skeletal muscle regeneration through synergistic action of extracellular vesicle cargo and soluble proteins. Stem Cell Res. Ther. 2019, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Tobin, S.W.; Alibhai, F.J.; Wlodarek, L.; Yeganeh, A.; Millar, S.; Wu, J.; Li, S.H.; Weisel, R.D.; Li, R.K. Delineating the relationship between immune system aging and myogenesis in muscle repair. Aging Cell 2021, 20, e13312. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The inflammatory response in CTX-induced skeletal muscle injury. (I) Infiltration of the immune cells. Upon injury, the immune cells are recruited into the damaged area, then induce inflammatory response in the damaged muscle. In this process, the lack of Klf2, STAT1, Hmox1, MKP-1 and upregulation of adiponectin (APN), activin A, and calmodulin (CaM) signaling increase, while the lack of IL-1a/β, Hsp70, CCR2, and IL-6/STAT3 and so on, inhibits the inflammatory infiltration. (II) Elimination of necrotic muscle fibers. The monocyte infiltrated into the damaged site, and become pro-inflammatory macrophages (M1 macrophages). M1 macrophages could secrete proinflammatory cytokines, maintain the inflammatory environment, then clean the necrotic debris. In this process, RetSat/MFG-E8 is required in the regulation of efferocytosis. ADAM8/PSGL-1 signaling, the TAM kinase signaling pathway, and the expression of SRB1 facilitate the elimination of necrotic fibers. Additionally, supplementation of balenine also increases the phagocytosis ability of macrophages. (III) Polarization of the macrophages. As the clearance of the muscle debris, the pro-inflammatory macrophages switch to anti-inflammatory macrophages (M2 macrophages), then secrete anti-inflammatory factors and stimulate regeneration. TG2 deficiency and the excessive calmodulin-dependent signaling delay/impair, while PGC-1α, SRB1, and so on, stimulate the polarization of the macrophages.
Figure 1.
The inflammatory response in CTX-induced skeletal muscle injury. (I) Infiltration of the immune cells. Upon injury, the immune cells are recruited into the damaged area, then induce inflammatory response in the damaged muscle. In this process, the lack of Klf2, STAT1, Hmox1, MKP-1 and upregulation of adiponectin (APN), activin A, and calmodulin (CaM) signaling increase, while the lack of IL-1a/β, Hsp70, CCR2, and IL-6/STAT3 and so on, inhibits the inflammatory infiltration. (II) Elimination of necrotic muscle fibers. The monocyte infiltrated into the damaged site, and become pro-inflammatory macrophages (M1 macrophages). M1 macrophages could secrete proinflammatory cytokines, maintain the inflammatory environment, then clean the necrotic debris. In this process, RetSat/MFG-E8 is required in the regulation of efferocytosis. ADAM8/PSGL-1 signaling, the TAM kinase signaling pathway, and the expression of SRB1 facilitate the elimination of necrotic fibers. Additionally, supplementation of balenine also increases the phagocytosis ability of macrophages. (III) Polarization of the macrophages. As the clearance of the muscle debris, the pro-inflammatory macrophages switch to anti-inflammatory macrophages (M2 macrophages), then secrete anti-inflammatory factors and stimulate regeneration. TG2 deficiency and the excessive calmodulin-dependent signaling delay/impair, while PGC-1α, SRB1, and so on, stimulate the polarization of the macrophages.

Figure 2.
SC activation, myoblast proliferation, differentiation and fusion, and the myotube maturation in CTX-induced injury models. (I) SC activation. In the damaged muscle, the quiescent SCs are activated to repair the skeletal muscle. Conversion of n-6 to n-3 polyunsaturated fatty acids (PUFAs), Salvador, testosterone, upregulation of Insl6, and so on, promote, while the lack of Nrf2, LCN2, and miRNA-501 inhibit the activation of SCs. (II) Myoblast proliferation. Activated SCs begin to proliferate and become myoblast. FGF2, RING finger protein13 (RNF13) derived from macrophages, Sod1/Cat, lack of p21, and so on facilitate, while FOXO1/p57/Gadd45α and CaMKK2/AMPK, lack of Gαi2 and H19/Igf2, etc., inhibit myoblast proliferation. (III) Myoblast differentiation and fusion. The expended myoblasts experience differentiation and fuse with other muscle fibers to form multinuclear myotubes. Lack of SLK and APOBEC2, upregulation of miR-206, miR-378, MMP-13, and Ccndbp1 enhance; Parkin, tension, Rbm24, and Hsp70 deficiency inhibit the capacity of myoblast differentiation. Lack of ANO5, HMGR, BAI3, and miR-188, and so on, inhibit, upregulation of Nox4/ROS, PLD1, Mustn1, and TRPV1 stimulate myoblast fusion. (IV) Maturation of myofibers. Lack of Dcx, PTEN, and desmin influence the myofiber maturation. (V) Self-renewal of SCs. The activated SCs proliferate to replenish the SC pool of the skeletal muscle. In this process, Angiotensin II, lack of mTORC2, AIF, SelN, and Notch1/Notch2 impair, while Lgr5, laminin-111, lack of PKC θ, Rb1, and Pax7 acetylation stimulate SC self-renewal or SC pool replenishment.
Figure 2.
SC activation, myoblast proliferation, differentiation and fusion, and the myotube maturation in CTX-induced injury models. (I) SC activation. In the damaged muscle, the quiescent SCs are activated to repair the skeletal muscle. Conversion of n-6 to n-3 polyunsaturated fatty acids (PUFAs), Salvador, testosterone, upregulation of Insl6, and so on, promote, while the lack of Nrf2, LCN2, and miRNA-501 inhibit the activation of SCs. (II) Myoblast proliferation. Activated SCs begin to proliferate and become myoblast. FGF2, RING finger protein13 (RNF13) derived from macrophages, Sod1/Cat, lack of p21, and so on facilitate, while FOXO1/p57/Gadd45α and CaMKK2/AMPK, lack of Gαi2 and H19/Igf2, etc., inhibit myoblast proliferation. (III) Myoblast differentiation and fusion. The expended myoblasts experience differentiation and fuse with other muscle fibers to form multinuclear myotubes. Lack of SLK and APOBEC2, upregulation of miR-206, miR-378, MMP-13, and Ccndbp1 enhance; Parkin, tension, Rbm24, and Hsp70 deficiency inhibit the capacity of myoblast differentiation. Lack of ANO5, HMGR, BAI3, and miR-188, and so on, inhibit, upregulation of Nox4/ROS, PLD1, Mustn1, and TRPV1 stimulate myoblast fusion. (IV) Maturation of myofibers. Lack of Dcx, PTEN, and desmin influence the myofiber maturation. (V) Self-renewal of SCs. The activated SCs proliferate to replenish the SC pool of the skeletal muscle. In this process, Angiotensin II, lack of mTORC2, AIF, SelN, and Notch1/Notch2 impair, while Lgr5, laminin-111, lack of PKC θ, Rb1, and Pax7 acetylation stimulate SC self-renewal or SC pool replenishment.

Figure 3.
Fibrosis in CTX-induced injury models. After injury, FAPs are activated and begin to proliferate and then differentiate into fibroblasts and adipocytes. Fibroblasts secrete extracellular matrix proteins and growth factors, and differentiate into myofibroblasts to increase α-SMA expression and ECM synthesis, which may provide scaffolds for the skeletal muscle regeneration while abnormal deposition of ECM finally results in fibrosis. In CTX-induced injury muscle, increases in miR-199a-5p, growth differentiation factor 11 (GDF11), platelet-derived growth factor receptor beta (PDGFRβ), lack of GDF-associated serum protein-1 (Gasp1) and/or Gasp2, and a prior burst of double homeobox 4 (DUX40) induce the deposition of collagen and contribute to fibrosis. In contrast, laminin-111 and Losartam therapy reduce fibrosis. Additionally, IL-4/IL-13 signaling, EGF and BTC, and the RA signaling pathway also regulate the fate of FAPs in skeletal muscle regeneration.
Figure 3.
Fibrosis in CTX-induced injury models. After injury, FAPs are activated and begin to proliferate and then differentiate into fibroblasts and adipocytes. Fibroblasts secrete extracellular matrix proteins and growth factors, and differentiate into myofibroblasts to increase α-SMA expression and ECM synthesis, which may provide scaffolds for the skeletal muscle regeneration while abnormal deposition of ECM finally results in fibrosis. In CTX-induced injury muscle, increases in miR-199a-5p, growth differentiation factor 11 (GDF11), platelet-derived growth factor receptor beta (PDGFRβ), lack of GDF-associated serum protein-1 (Gasp1) and/or Gasp2, and a prior burst of double homeobox 4 (DUX40) induce the deposition of collagen and contribute to fibrosis. In contrast, laminin-111 and Losartam therapy reduce fibrosis. Additionally, IL-4/IL-13 signaling, EGF and BTC, and the RA signaling pathway also regulate the fate of FAPs in skeletal muscle regeneration.

Figure 4.
Non-SC stem cells regulate regeneration in CTX-induced injury muscle. Transplantation of hematopoietic stem cells, adipose-derived mesenchymal stem cells, bone marrow-derived mesenchymal stem cells, and amniotic fluid stem cells stimulate regeneration in the damaged skeletal muscle.
Figure 4.
Non-SC stem cells regulate regeneration in CTX-induced injury muscle. Transplantation of hematopoietic stem cells, adipose-derived mesenchymal stem cells, bone marrow-derived mesenchymal stem cells, and amniotic fluid stem cells stimulate regeneration in the damaged skeletal muscle.


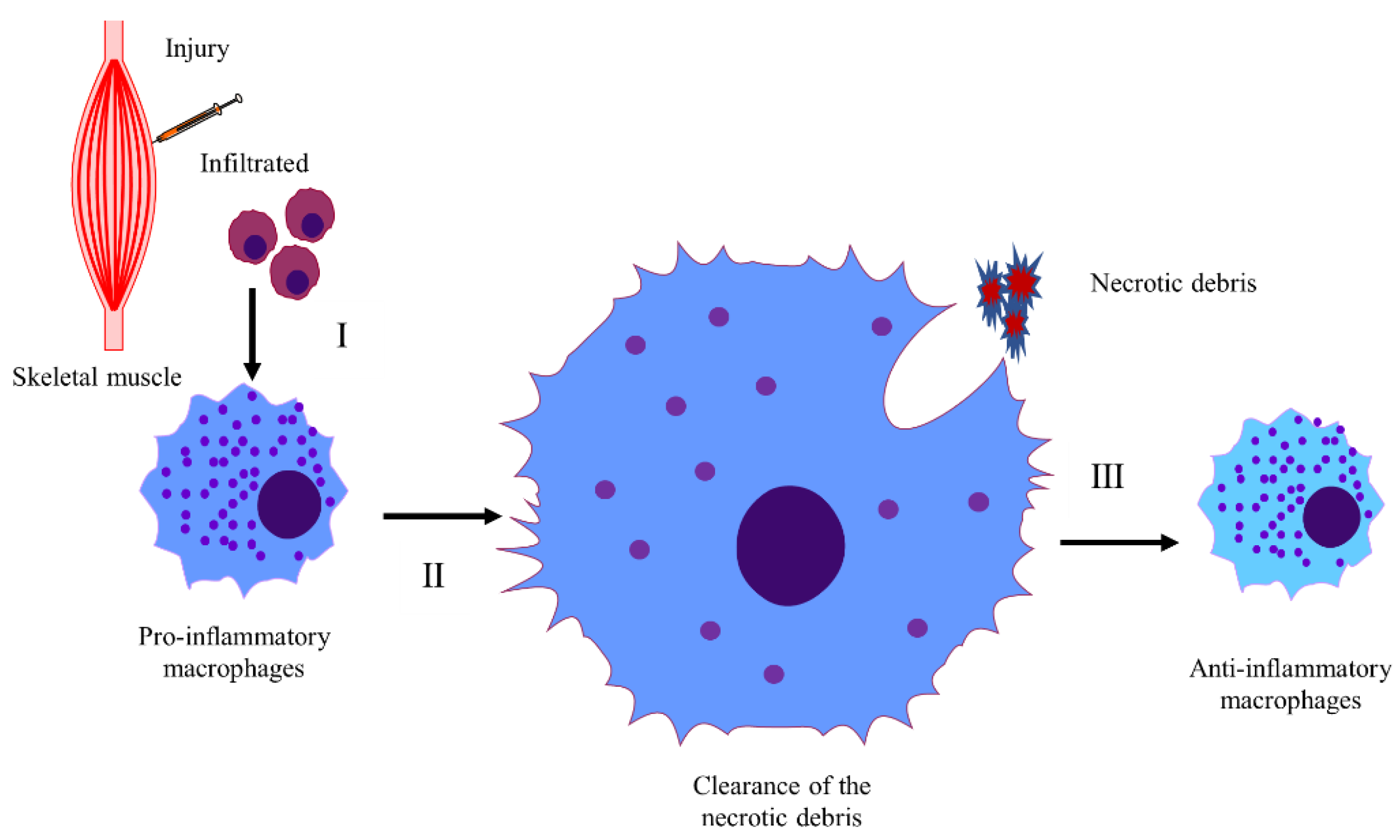{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Different mice models on CTX-induced regeneration in skeletal muscle.
| Author, Year | Injury Portions | Mouse Models | Targets | Regeneration (Impair/Improve) | Ref. |
|---|---|---|---|---|---|
| Mouisel E, 2010 | Right tibialis anterior | 5 weeks,5, 12 and 18–24 months mdx mice | Different ages | Impair | [28] |
| Fearing CM, 2016 | Right hind limb anterior and post compartments | Male and female C57BL/6J mice; young:4-6 months, middle:12–19 months, old 25–30 months and very old:32–33 months | Ages and sex | Impair; —— | [40] |
| Takahashi Y, 2021 | Left tibialis anterior | 22 weeks C57BL/6-WT, C57BL/6-Akita, KK/Ta-WT, and KK/Ta-Akita male mice | Diabetes | Impair | [20] |
| Vignaud A, 2007 | Right tibialis anterior | 3–4 months STZ-treated Swiss and Akita male mice | Diabetes | Impair | [21] |
| Chaiyasing R, 2021 | Right tibialis anterior | 12 weeks ovariectomized and normal C5BL/6JJcl female mice | Estrogen | Impair | [41] |
| Rebalka IA, 2017 | Tibialis anterior | 10–12W STZ-treated and normal male C57BL/6J mice | Fluvastatin | Impair | [42] |
| Nguyen MH, 2011 | Extensor digitorum longus | 14–16 weeks leptin-deficient, leptin receptor-mutant mice and a group of C57BL/6 mice fed a high-fat diet | ob/ob and db/db | Impair | [22] |
| D’Souza DM, 2015 | Left gastrocnemius-plantaris, tibialis anterior, quadriceps | 16 weeks male C57BL/6J mice fed a high-fat diet | Diet-induced obesity | Impair | [24] |
| Jinno N, 2014 | Right gastrocnemius | 8 weeks male C57BL/6 mice: a marginally zinc-deficient diet-fed group, a zinc-adequate diet-fed group and a zinc-high diet-fed group | Zinc | Impair | [36] |
| Matsuba Y, 2009 | Soleus muscle | gravitational unloading | Gravitational unloading | Impair | [32] |
| Jeong J, 2013 | Tibialis anterior and gastrocnemius | 2–4 months C57BL/6 male mice treated with STZ, 20–24 months C57BL/6 male mice and C57BL/6J-Ins2Akita mice | Diabetes | Impair | [43] |
| Inaba S, 2018 | Tibialis anterior | cancer cachexia | Cancer cachexia | Impair | [26] |
| McHale MJ, 2012 | Right hind limb anterior and posterior compartment | 4–6 months ovariectomized, castrated and normal male and female C57BL/6J mice | Sex hormones | —— | [44] |
| Patsalos A, 2017 | Tibialis anterior | 2–6 months BoyJ, C57BL/6J male mice with or without irradiation and Pax7 Cre-Rosa26 DTA mice | Irradiation | Impair | [29] |
| Ikeda Y, 2019 | Gastrocnemius | 8 weeks male C57BL/6J mice with or without iron overload | Iron | Impair | [45] |
| Attia M, 2017 | Right tibialis anterior | 10 weeks myasthenia gravis and normal female C57BL/6J mice | Myasthenia gravis | Impair | [46] |
| Saliu TP, 2022 | Left tibialis anterior and gastrocnemius | 9 weeks normal and NAFLD male CD-1 mice | NAFLD | Impair | [47] |
| Rahman FA, 2020 | Left tibialis anterior | Young (3 months) and old (18 months) male C57BL/6J mice | Aging | Impair | [48] |
| Kohno S, 2012 | Soleus | C57BL/6J mice with or without tail suspension | Unloading | Impair | [38] |
| Kimoloi S, 2022 | Tibialis anterior | Both male and female K320Eskm and K320Emsc transgenic mice | Mitochondrial DNA alterations | Impair | [39] |
| Paiva-Oliveira EL, 2017 | Right gastrocnemius | Isogenic 6–8 weeks C3H/HeJ (TLR4 defective), C3H/HeN(TLR4 WT), C57BL/6 (WT) and TLR-4 knockout male mice | Different strains | —— | [49] |
| Yoshioka K, 2021 | Tibialis anterior and masseter | C57BL/6J and mdx mice | Position specificity | —— | [50] |
| Joanisse S, 2016 | Tibialis anterior | 24 months old male C57BL/6J mice with or without exercise and 8 weeks young male mice | Exercise and age | Improve; impair | [33] |
| Nagata K, 2013 | Tibialis anterior | 8 weeks C57BL/6J mice with ultrasound exposure | Ultrasound | Improve | [51] |
| Morioka S, 2008 | Soleus | 10 weeks male C57BL/6J mice with or without functional overloading | Functional overloading | Improve | [37] |
| Fujiya H, 2015 | Left tibialis anterior | 7 weeks male C57BL/6J mice with or without microcurrent electrical neuromuscular stimulation (MENS) | MENS | Improve | [35] |
Table 2.
The mechanism of inflammatory infiltration, conversion of macrophages, and elimination of the necrotic debris.
Table 2.
The mechanism of inflammatory infiltration, conversion of macrophages, and elimination of the necrotic debris.
| Author, Year | Injury Portions | Target Molecule/ Drug | Target Process | Expression | Effects (Positive/Negative) | Regeneration (Impair/Improve) | Ref. |
|---|---|---|---|---|---|---|---|
| Shi D, 2018 | Tibialis anterior | CaMKIV | Infiltration of macrophages | Up/Down | Positive/Negative | Impair/improve | [57] |
| Neves Jde C, 2015 | Right tibialis anterior | Neuraminidase-1 | Inflammatory response; myofiber maturation | Down | Positive; Negative | Impair | [58] |
| Liao ZH, 2019 | Tibialis anterior | Estrogen signaling | Inflammation infiltration; conversion of macrophages from M1 to M2 | Down | Positive; Negative | Impair | [59] |
| Kohno S, 2011 | Tibialis anterior | Cbl-b | Cytotoxic T-cell infiltration | Down | Positive | Impair | [60] |
| Wang H, 2014 | Right tibialis anterior and hindlimb posterior compartment | Monocyte/ macrophage | Monocyte and macrophages recruitment; conversion of macrophages from M1 to M2 | —— | Positive; Negative | Impair | [61] |
| Park CY, 2010 | Gastrocnemius and soleus | skNAC | Inflammation infiltration and myonecrosis | Down | Positive | Impair | [62] |
| Shi H, 2010 | Right tibialis anterior | MKP-1 | Inflammation; myoblast proliferation; differentiation | Down | Positive; Negative; Positive | Impair | [63] |
| Hu J, 2019 | Tibialis anterior | CaM signaling | Inflammatory response | Up | Positive | Improve | [64] |
| Manoharan P, 2019 | Gastrocnemius | Klf2 | Inflammatory response | Down | Positive | Improve | [65] |
| Gao Y, 2012 | Unilateral tibialis anterior | STAT1 | Inflammatory response | Down | Positive | Improve | [66] |
| Kozakowska M, 2018 | Gastrocnemius | Hmox1 | Inflammation and SC proliferation | Down | Positive | Improve | [67] |
| Koh, 2005 | Extensor digitorum longus | PAI-1 | Macrophage and SC migration | Down | Positive | Improve | [2] |
| Zhang, 2020 | Tibialis anterior and gastrocnemius | IFN-γ/CXCL10/ CXCR3 | Macrophages and myoblast proliferation | —— | Positive | Improve | [68] |
| Yaden BC, 2014 | Right Gastrocnemius | Activin A | Macrophage infiltration | Up | Positive | Improve | [69] |
| Mothe-Satney I, 2017 | Left Tibialis Anterior | PPARβ | Macrophage recruitment | Up | Positive | Improve | [70] |
| Tanaka Y, 2019 | Tibialis Anterior | APN | Elimination of the necrotic fibers | Up | Positive | Improve | [71] |
| Dinulovic I, 2016 | Tibialis Anterior | PGC-1α | Conversion of macrophages from M1 to M2 | Up/Down | Positive; Negative | Improve/Impair | [56] |
| Sugihara H, 2018 | Tibialis Anterior | PGRN | Prolonged Persistence of M2 Macrophages | Down | Positive | Improve | [72] |
| Lo Sicco, 2017 | Tibialis Anterior | Extracellular vesicles released by human adipose derived-MSCs | Conversion of macrophages from M1 to M2 | Up | Positive | Improve | [73] |
| Yang M, 2022 | tibialis anterior | Balenine | Phagocytosis ability of macrophages | —— | Positive | improve | [74] |
| Cardoso ES, 2016 | Gastrocnemius | Thymol | Inflammatory response | —— | Negative | Improve | [75] |
| Wang ZG, 2021 | Gastrocnemius | Conversion of n-6 to n-3 PUFAs | Inflammatory response; SC activation | Up | Negative; Positive | Improve | [76] |
| Chaweewannakorn C, 2018 | Unilaterally tibialis anterior | IL-1a/β | Inflammatory response | Down | Negative | Impair | [77] |
| Senf SM, 2013 | Tibialis anterior | Hsp70 | Inflammatory response | Down | Negative | Impair | [78] |
| Mojumdar K, 2016 | Tibialis anterior | TLR2 | Macrophage accumulation; elimination of the necrotic fibers | Down | Negative; Negative | Impair | [79] |
| Varga T, 2013 | Tibialis anterior | NUR77 | Macrophage development | Down | —— | Impair | [80] |
| Ochoa O, 2007 | Right anterior and posterior compartment | CCR2 | Macrophage recruitment and angiogenesis and VEGF production | Down | Negative | Impair | [81] |
| Zhang C, 2013 | Tibialis anterior and gastrocnemius | IL-6/STAT3 | Infiltration of macrophages and myoblast proliferation | Down | Negative | Impair | [82] |
| Zhang J, 2014 | Tibialis anterior | CD8 | Macrophage recruitment | Down | Negative | Impair | [83] |
| Martinez CO, 2010 | Right tibialis anterior and hindlimb posterior compartment | CCR-2/MCP-1 | Macrophage recruitment | Down | Negative | Impair | [84] |
| Krause MP, 2013 | Left tibialis anterior and gastrocnemius-plantaris-soleus | Diabetes | Macrophages infiltration | —— | —— | Impair | [23] |
| Cheng M, 2008 | Extensor digitorum longus and tibialis anterior | IFN-γ | Macrophages infiltration; myoblast proliferation | Down | Negative | Impair | [85] |
| Zhang, 2017 | Tibialis anterior and gastrocnemius | C3a | Monocyte/macrophage infiltration | Down | Negative | Impair | [86] |
| Sun D, 2009 | Right hind limb anterior and posterior compartment | CCR2 | Recruitment of macrophages and neutrophils | Down | Negative | Impair | [87] |
| Nishimura D, 2015 | Tibialis anterior | ADAM8 | Elimination of the necrotic fibers | Down | Negative | Impair | [88] |
| AI-Zaeed N, 2021 | Tibialis anterior | TAM kinase receptor Mer | Elimination of the necrotic fibers and conversion of macrophage from M1 to m2s | Down | Negative | Impair | [89] |
| Zhang J, 2019 | Tibialis anterior | SRB1 | Elimination of the necrotic fibers and conversion of macrophage from M1 to M2 | Down | Negative | Impair | [90] |
| Jin R M, 2018 | One or more hindlimb muscles | Preexisting inflammatory environment | Conversion of macrophage from M1 to M2 | —— | Negative | Impair | [91] |
| Bronisz-Budzyńska I, 2020 | Gastrocnemius | Nrf2 | Inflammatory response | Down | Positive | No effects | [92] |
| Tarban N, 2022 | Tibialis anterior | Retinol saturase | Phagocytosis ability of macrophages | Down | Negative | No effects | [93] |
| Dalle S, 2020 | Tibialis anterior | Ibuprofen | Inflammatory response | —— | Negative | No effects | [94] |
| Shen W, 2008 | Gastrocnemius | Macrophage, TFG-β1 and COX-2 | Inflammatory response | —— | —— | —— | [95] |
| Rousseau AS, 2021 | Left tibialis anterior | PPARβ/δ | T cell dynamic | Down | —— | —— | [96] |
Table 3.
The mechanism of SC activation, myoblast proliferation, differentiation, and fusion.
| Author, Year | Injury Portions | Target Molecule/ Drug | Target Process | Expression | Effects (Positive/Negative) | Regeneration (Impair/Improve) | Ref. |
|---|---|---|---|---|---|---|---|
| Schaaf G J, 2018 | Quadriceps femoris and gastrocnemius | Acid alpha glucosidase | SC activation | Down | Negative | Impair | [115] |
| Serra C, 2013 | Left tibialis anterior | Testosterone | SC activation | Up | Positive | Improve | [116] |
| Liu Q, 2021 | Left tibialis anterior | Salvador | SC activation and angiogenesis | Down | Positive | Improve | [117] |
| Zeng L, 2010 | Tibialis anterior or gastrocnemius | Insl6 | SC activation and proliferation | Up/Down | Positive/Negative | Improve/impair | [108] |
| Shelar SB, 2016 | Tibialis anterior | Nrf2 | SC activation and proliferation | Down | Negative | Impair | [113] |
| Rebalka IA, 2018 | Left tibialis anterior or left gastrocnemius | Lcn2 | SC activation and fibrosis | Down | Negative | Impair | [114] |
| Zeng P, 2016 | Right tibialis anterior | Mir-378/IGF1R | SC activation and differentiation | Up | Negative | Impair | [118] |
| Nissar AA, 2011 | Tibialis anterior | Xin | SC activation | Down | Negative | Impair | [111] |
| Lagalice L, 2018 | Tibialis anterior | Acid alpha glucosidase | SC activation | Down | Negative | Impair | [119] |
| Mizbani A, 2016 | Tibialis anterior | Mirna-501 | SC activation | Down | Negative | Impair | [120] |
| Fiore PF, 2020 | Tibialis anterior | Pkcθ | SC self-renewal | Down | Positive | Improve | [121] |
| Fortier M, 2013 | Tibialis anterior | S1pr3 | SC proliferation | Down | Positive | Improve | [122] |
| Cai S, 2020 | Tibialis anterior | Mll1/myf5 | SC proliferation | Up | Positive | Improve | [123] |
| Sincennes MC, 2021 | Tibialis anterior | Pax7 acetylation | SC pool | Down | Positive | Improve | [124] |
| Naito T, 2009 | Tibialis anterior | G-csf | SC number | Up | Positive | Improve | [125] |
| Ohno Y, 2016 | Left soleus | Mstn | SC number | Down | Positive | Improve | [31] |
| Price FD, 2014 | Tibialis anterior | Jak/stat | SC number | Down | Positive | Improve | [126] |
| Hillege MMG, 2022 | Tibialis anterior | TGF-β signaling | SC number | Down | Positive | Improve | [112] |
| Angione AR, 2011 | Tibialis anterior and gastrocnemius | Pparδ | SC number and proliferation | Down | Negative | Impair | [127] |
| Nishizawa S, 2013 | Left soleus | Hsf1 | SC number and proinflammatory response | Down | Negative | Impair | [128] |
| Ahrens HE, 2018 | Right tibialis anterior | Klotho | SC number and function | Down | Negative | Impair | [129] |
| Sakamoto K, 2019 | Forearm muscle | R3hdml | SC number | down | Negative | Impair | [130] |
| Bye-A-Jee H, 2018 | Tibialis anterior | ZFP36L1 and ZFP36L2 | SC number | Down | Negative | Impair | [131] |
| Tonami K, 2013 | Left tibialis anterior | Capn6 | Myoblast differentiation | Down | Positive | Improve | [132] |
| Accornero F, 2014 | Tibialis anterior | TGF-β | SC number and activity; decreased degeneration | Down | Positive; Positive | Improve | [133] |
| Van Ry PM, 2014 | Tibialis anterior | Laminin-111 | SC pool; fibrosis | Up | Positive; Negative | Improve | [134] |
| Hosoyama T, 2011 | Tibialis anterior | Rb1 | SC pool; differentiation | Down | Positive; Negative | Impair | [135] |
| Rion N, 2019 | Unilaterally tibialis anterior | mTORC2 | SC pool replenishment | Down | Negative | Impair | [136] |
| Yoshida T, 2013 | Unilateral gastrocnemius | Angiotensin II | SC pool and proliferation | Up | Negative | Impair | [137] |
| Armand AS, 2011 | Soleus or EDL | AIF | SC pool | Down | Negative | Impair | [138] |
| Milanesi A, 2017 | Right tibialis anterior or quadriceps femoris | Thyroid hormone receptor alpha | SC pool | Down | Negative | Impair | [139] |
| Castets P, 2011 | Unilateral tibialis anterior and soleus muscles | SelN | SC pool | Down | Negative | Impair | [140] |
| Fujimaki S, 2018 | Right tibialis anterior | Notch1/Notch2 | SC pool and proliferation; myoblast differentiation; fibrosis | Down | Negative; Positive; Positive | Impair | [141] |
| Johnston AP, 2011 | Tibialis anterior | Ang II | SC number; myoblast differentiation | Down | Positive; Negative | Impair | [142] |
| Jaafar M N, 2016 | Right tibialis anterior | Primary cilium | SC self-renewal | —— | —— | —— | [143] |
| Leung C, 2020 | Tibialis anterior/Extensor digitorum longus | Lgr5 | SC replenish and myofiber formation | —— | Positive | Improve | [144] |
| Buono R, 2012 | Tibialis anterior and quadriceps | NO signaling | SC self-renewal and proliferation | Down | Negative | Impair | [145] |
| Urciuolo A, 2014 | Tibialis anterior | Collagen VI | SC self-renewal | Down | Negative | Impair | [146] |
| Alexeev V, 2014 | Left gastrocnemius muscle | Adipose-derived stem cells | Migration of SCs | —— | Positive | Improve | [147] |
| Brien P, 2013 | Tibialis anterior/extensor digitorum longus muscle group | P38α | Myoblast proliferation; differentiation | Down | Positive; Negative | Impair | [148] |
| Hawke TJ, 2003 | Tibialis anterior | p21 | Myoblast proliferation; differentiation | Down | Positive; Negative | Impair | [149] |
| Cortez-Toledo O, 2017 | Tibialis anterior | Nur77 | Myoblast proliferation | Down | Negative | No effects | [150] |
| Alves, 2019 | Rectus femoral muscle | Kinin-B2 receptor | Myoblast proliferation; differentiation | Down | Positive; Negative | Impair | [4] |
| Wu, 2014 | Right tibialis anterior | Duxbl | Myoblast proliferation and differentiation | Up | Positive; Negative | Impair | [151] |
| Chen Y, 2012 | Tibialis anterior | miR-351 | Myoblast proliferation and differentiation | Up/Down | Positive/Negative | Improve/impair | [152] |
| Tseng C, 2019 | Gastrocnemius | Sod1/Cat | Myoblast proliferation and differentiation | Up | Positive | Improve | [153] |
| Jia Y, 2012 | Gastrocnemius | EPO | Proliferation and survival of the SCs | —— | Positive | Improve | [154] |
| Shibasaki H, 2019 | Tibialis anterior | miR-188 | Myoblast fusion | Up/Down | Positive/Negative | Improve/impair | [155] |
| Hawke TJ, 2007 | Tibialis anterior | Xin | Myoblast proliferation and migration | Down | Positive | Improve | [109] |
| Meng, 2014 | Tibialis anterior | RNF13 | Myoblast proliferation and differentiation | Up | Positive | Improve | [104] |
| Lee EJ, 2021 | Left gastrocnemius | Glycyrrhiza uralensis-extracted compounds | Myoblast proliferation and differentiation | —— | Positive | Improve | [156] |
| Galimov A, 2016 | Tibialis anterior | FGF2 | Myoblast proliferation | Up | Positive | Improve | [107] |
| Armand, 2003 | Soleus | FGF6 | Myoblast proliferation | Down/Up | Negative/Positive | Impair/improve | [157] |
| Shi, 2010 | Tibialis anterior and gastrocnemius | Tceal7 | Myoblast proliferation; differentiation | Up | Negative; Positive | —— | [1] |
| Pessemesse L, 2019 | Right tibialis anterior | p43 | Myoblast proliferation | Down/Up | Negative/Positive | Impair/improve | [158] |
| Ye, 2016 | Gastrocnemius | CaMKK2 | Myoblast proliferation and differentiation | Up/Down | Negative/Positive | Impair/ Improve | [159] |
| Minetti GC, 2014 | Tibialis anterior | Gαi2 | Myoblast proliferation, differentiation and fusion | Down | Negative | Impair | [160] |
| Zhang CC, 2021 | Tibialis anterior | Cyp4a14 | Myoblast proliferation and differentiation and inflammatory response | Down | Negative | Impair | [161] |
| Ding, 2021 | Tibialis anterior | Tfr1 | Myoblast proliferation and differentiation | Down | Negative | Impair | [162] |
| Naito M, 2016 | Tibialis anterior | Dnmt3a | Myoblast proliferation | Down | Negative | Impair | [163] |
| Bae, 2020 | Tibialis anterior | Cdon | Myoblast proliferation and senescence | Down | Negative | Impair | [164] |
| Rooney JE, 2009 | Left tibialis anterior | α7 integrin | Myoblast proliferation and differentiation | Down | Negative | Impair | [165] |
| Katsushi, 2020 | Tibialis anterior | Bach 1 | Myoblast proliferation and differentiation | Down | Negative | Impair | [166] |
| Yamashita, 2016 | Gastrocnemius | FOXO1 | Myoblast proliferation | Up | Negative | Impair | [167] |
| Al-Sajee D, 2015 | Left tibialis anterior, gastrocnemius/ Plantaris/soleus, quadriceps muscles | Xin | Myoblast proliferation | Down | Negative | Impair | [110] |
| Girgenrath, 2006 | Tibialis anterior | Fn14 | Myoblast proliferation | Down | Negative | Impair | [168] |
| Martinet C, 2016 | Tibialis anterior | H19 | Myoblast proliferation | Down | Negative | Impair | [169] |
| Yahiaoui, 2008 | Tibialis anterior | MCP-1 | Myoblast proliferation | Up | Negative | Impair | [170] |
| Kursaka, 2017 | One leg of tibialis anterior | Egr3 | Myoblast proliferation | Down | Negative | Impair | [171] |
| Ochiai N, 2016 | Tibialis anterior | fad24 | Myoblast proliferation | Down | Negative | Impair | [172] |
| Zanou, 2012 | Tibialis anterior and extensor digitorium longus | Trpc1 | Myoblast migration and differentiation | Down | Negative | Impair | [173] |
| Yablonka-Reuveni Z, 2015 | Unilateral tibialis anterior | FGFR1 | Myoblast proliferation | Down | Negative | No effects | [174] |
| Ohtsubo, 2017 | Gastrocnemius | APOBEC2 | Myoblast differentiation and fusion | Down | Positive | Improve | [8] |
| He, 2019 | Extensor digitorum longus | Nicotine | Myoblast differentiation | —— | Positive | Improve | [175] |
| Wu, 2007 | Tibialis anterior | Sema4C | Myoblast differentiation | Up/Down | Positive/Negative | Improve/impair | [176] |
| Liu, 2012 | Tibialis anterior | miR-206 | Myoblast differentiation | Up | Positive | Improve | [177] |
| Song, 2018 | Tibialis anterior | Linc-smad7 | Myoblast differentiation | Up | Positive | Improve | [178] |
| Gatta L, 2017 | Right tibialis anterior | Trimetazidine | Myoblast differentiation | —— | Positive | Improve | [179] |
| Gagan, 2011 | Tibialis anterior | miR-378 | Myoblast differentiation | Up | positive | Improve | [180] |
| Mikami T, 2012 | Tibialis anterior | Chondroitin sulfate | Myoblast differentiation | Down | Positive | Improve | [181] |
| Lee KP, 2015 | Hindlimb muscle | miR-431 | Myoblast differentiation | Up | positive | Improve | [27] |
| Storbeck CJ, 2013 | Tibialis anterior | SLK | Myoblast differentiation | Down | Positive | Improve | [182] |
| Lei, 2013 | One leg of tibialis anterior | MMP-13 | Myoblast migration | Down/Up | Negative/Positive | Impair/improve | [183] |
| Das, 2017 | Tibialis anterior | ACL | Myoblast differentiation | Down/Up | Negative/Positive | Impair/improve | [184] |
| Yoshida T, 2014 | Gastrocnemius | AT2R | Myoblast differentiation | Down/Up | Negative/Positive | Impair/improve | [185] |
| Huang Y, 2016 | Right tibialis anterior | Ccndbp1 | Myoblast differentiation | Down/Up | Negative/Positive | Impair/ Improve | [186] |
| Chen SE, 2007 | Soleus | TNF-α | Myoblast differentiation | Down/Up | Negative/Positive | Impair/ Improve | [187] |
| He, 2021 | Tibialis anterior | IRE1a | Myoblast differentiation and hypertrophy | Down/Up | Negative/ Positive | Impair/improve | [188] |
| Luca E, 2020 | Tibialis anterior | miRNA network | Myoblast differentiation | Down | Negative | Improve | [189] |
| Esteca MV, 2020 | Left tibialis anterior | Parkin | Myoblast differentiation | Down | Negative | Impair | [190] |
| Ishii A, 2001 | Tibialis anterior | Tensin | Myoblast differentiation and fusion | Down | Negative | Impair | [191] |
| Zhang M, 2020 | Tibialis anterior of one limb | Rbm24 | Myoblast differentiation | Down | Negative | Impair | [106] |
| Lee, 2015 | Tibialis anterior | miR-431 | Myoblast differentiation | Down | Negative | Impair | [192] |
| Fan, 2018 | Tibialis anterior | Hsp70 | Myoblast differentiation | Down | Negative | Impair | [16] |
| Cerquone, 2018 | Tibialis anterior | PAK1 | Myoblast differentiation | Down | Negative | Impair | [193] |
| Lee, 2020 | Tibialis anterior | PHF20 | Myoblast differentiation | Up | Negative | Impair | [194] |
| Hayashi, 2016 | Tibialis anterior | Klf5 | Myoblast differentiation | Down | Negative | Impair | [3] |
| Lee, 2015 | Tibialis anterior | AKAP6 | Myoblast differentiation | Down | Negative | Impair | [195] |
| Li, 2020 | Tibialis anterior | LRTM1 | Myoblast differentiation | Down | Negative | Impair | [196] |
| Lin, 2019 | Left gastrocnemius | MPM | Myoblast differentiation | Down | Negative | Impair | [197] |
| Harada, 2018 | Tibialis anterior | H3mm7 | Myoblast differentiation | Down | Negative | Impair | [198] |
| Tetsuaki, 2009 | Tibialis anterior | CT-1 | Myoblast differentiation | Up | Negative | Impair | [199] |
| Paul, 2012 | Tibialis anterior | COPR5 | Myoblast differentiation | Down | Negative | Impair | [200] |
| Faralli, 2011 | Tibialis anterior and gastrocnemius of one hind limb | Tshz3 | Myoblast differentiation | Up | Negative | Impair | [201] |
| Liu N, 2014 | Tibialis anterior | MEF2A, C and D | Myoblast differentiation | Down | Negative | Impair | [202] |
| Kielbasa OM, 2011 | Unilateral tibialis anterior | Myospryn | Myoblast differentiation | Up | Negative | Impair | [203] |
| Verpoorten S, 2020 | Tibialis anterior | CD36 | Myoblast differentiation | Down | Negative | Impair | [204] |
| Andrée B, 2002 | Right gastrocnemius and soleus | Pop | Myoblast differentiation | Down | Negative | Impair | [205] |
| Paolini A, 2018 | Tibialis anterior | Autophagy | Myoblast differentiation | Down | Negative | Impair | [206] |
| Clow C, 2010 | Tibialis anterior | BDNF | Myoblast differentiation | Down | Negative | Impair | [207] |
| Marshall JL, 2012 | Left quadriceps | SSPN | Myoblast differentiation | Down | Negative | Impair | [208] |
| Chen SE, 2005 | Soleus | TNF-α | Myoblast differentiation | Down | Negative | Impair | [209] |
| Ravel, 2014 | Tibialis anterior | Staufen1 | Myoblast differentiation | Up | Negative | Impair | [210] |
| Langsdorf A, 2007 | Tibialis anterior | Sulfs | Myoblast differentiation | Down | Negative | Impair | [211] |
| Liu H, 2011 | Tibialis anterior and soleus | β3-Integrin | Myoblast differentiation | Down | Negative | Impair | [212] |
| Watanabe S, 2007 | Right tibialis anterior | Lbx1 | Myoblast differentiation | Down | Negative | Impair | [213] |
| Fu D, 2015 | Tibialis anterior | Mdm2 | Myoblast differentiation | Down | Negative | Impair | [214] |
| Schroer, 2019 | Tibialis anterior | RGS12 | A switch from myoblast proliferation to differentiation | Down | Negative | Impair | [215] |
| Lim S, 2013 | Left tibialis anterior | CBR1 | Myoblast differentiation | Down | Negative | —— | [216] |
| Mammen AL, 2009 | Right tibialis anterior | Mi-2 | Myoblast differentiation | Up | Negative | —— | [217] |
| Wu Z, 2020 | Tibialis anterior | Andrographolide | Myoblast differentiation and fusion | —— | —— | Improve | [218] |
| Kurosaka, 2021 | Left tibialis anterior | STAT6 | Myoblast differentiation and fusion | Down | —— | Improve | [219] |
| Budai Z, 2021 | Tibialis anterior | TG2 | Myoblast fusion and conversion of macrophages from M1 to M2 | Down | Negative | Impair | [102] |
| Vijayakumar A, 2013 | Unilateral tibialis anterior | IGF-1R | Myoblast fusion | Down | Negative | Impair | [220] |
| Pryce BR, 2017 | Unilateral tibialis anterior | SLK | Myoblast fusion | Down | Negative | Impair | [221] |
| Redelsperger, 2016 | One tibialis anterior | Syncytin | Myoblast fusion | Down | Negative | Impair | [222] |
| Kaspar, 2013 | Tibialis anterior | 3′ untranslated region of c-Myb | Myoblast fusion | Down | Negative | Impair | [223] |
| Griffin, 2016 | Left tibialis anterior and gastrocnemius | ANO5 | Myoblast fusion | Down | Negative | Impair | [224] |
| Trapani L, 2012 | Right tibialis anterior | HMGR | Myoblast fusion | Down | Negative | Impair | [225] |
| Hamoud, 2018 | Tibialis anterior | BAI3 | Myoblast fusion | Down | Negative | Impair | [226] |
| Tamilarasan K P, 2012 | Gastrocnemius | Lipid accumulation | Myoblast fusion | Up | Negative | Impair | [227] |
| Yoshida N, 2019 | Tibialis anterior | (P)RR | Myoblast fusion | Up | Negative | Impair | [228] |
| Shibasaki H, 2019 | Tibialis anterior | miR-188 | Myoblast fusion | Up/Down | Positive/Negative | Improve/impair | [155] |
| Youm TH, 2019 | tibialis anterior | Nox4/ROS | Myoblast fusion | —— | Positive | Improve | [229] |
| Teng, 2015 | One tibialis anterior | PLD1 | Myoblast fusion | Up | Positive | Improve | [230] |
| Krause MP, 2013 | Left tibialis anterior | Mustn1 | Myoblast fusion | Up | Positive | Improve | [231] |
| Kurosaka, 2016 | Tibialis anterior | TRPV I | Myoblast fusion | Up | Positive | Improve | [232] |
| Singhal N, 2015 | Gastrocnemius, quadriceps, tibialis anterior | Galgt1 | Myoblast fusion; SC apoptosis | Down | Negative; Positive | Impair | [233] |
| Yalvac ME, 2017 | Left tibialis anterior and left gastrocnemius | Calpain-3 | Myoblast fusion; fibrosis | Down | Negative; Positive | Impair | [234] |
| Ogawa, 2015 | Tibialis anterior | Dcx | Myofiber maturation | Down | Negative | Impair | [235] |
| Ohno Y, 2019 | Right tibialis anterior | Lactate | Myotube formation | Up | Positive | Improve | [236] |
| Hoshino S, 2013 | Right tibialis anterior | CHC22 | Myofiber maturation | Up | Negative | Impair | [237] |
| Hu Z, 2010 | Unilateral tibialis anterior | PTEN | Myofiber maturation; fibrosis | Down | Positive | Improve | [25] |
| Agbulut O, 2001 | Gastrocnemius and soleus or tibialis anterior | Desmin | Myofiber maturation; neuromuscular junctions | Down | Negative | Impair | [238] |
| Cicchillitti, 2012 | Tibialis anterior | miR-210 | Myoblast differentiation | Down | —— | No effects | [239] |
| Piccioni A, 2014 | Tibialis anterior | Shh | Activated SCs | Up | —— | Improve | [240] |
| Ceco E, 2021 | Left tibialis anterior | Elevated CO2 exposure | Myoblast differentiation and fusion | —— | —— | Impair | [30] |
| Liu, 2017 | Tibialis anterior | Twist2 | Maintain SC state | —— | —— | —— | [241] |
Table 4.
Fibrosis, calcification, and apoptosis.
| Author, Year | Injury Portions | Target Molecule/ Drug | Target Process | Expression | Effects (Positive/Negative) | Regeneration (Impair/Improve) | Ref. |
|---|---|---|---|---|---|---|---|
| Heredia JE, 2013 | Unilateral tibialis anterior | IL-4 | FAP proliferation | Down | Negative | Impair | [55] |
| Vumbaca S, 2021 | Tibialis anterior, quadriceps, and gastrocnemius | IL1a/IL1β and extracellular vesicles | FAP proliferation and differentiation | Up | Positive | —— | [248] |
| Zhao L, 2020 | Tibialis anterior | RA signaling | FAP proliferation | Up | positive | Improve | [246] |
| Zanotti S, 2018 | Tibialis anterior | Exosome miR-199a-5p/CAV1 | Fibrosis | Up | Positive | Impair | [247] |
| Horii N, 2018 | Tibialis anterior | C1q/Wnt and resistance training | Fibrosis | Up | positive | Impair | [34] |
| Murray, 2017 | Tibialis anterior | αV intergin | Fibrosis | Down | Negative | Improve | [245] |
| Burks, 2011 | Tibialis anterior | Losartam | Fibrosis | —— | Negative | Improve | [250] |
| Stepien DM, 2020 | Left tibialis anterior | TGF-β1 | Fibrosis | Down | Negative | Improve | [249] |
| Bosnakovski D, 2022 | Tibialis anterior | a prior DUX4 burst | Fibrosis | —— | Positive | —— | [251] |
| Ding, 2016 | Tibialis anterior | TAR RNA-binding protein (Trbp) | Fibrosis; myofiber formation | Down | Positive; Negative | Impair | [244] |
| Ogasawara S, 2018 | Left gastrocnemius | CatK | Fibrosis; inflammation and cell apoptosis | Down | Negative | Improve | [252] |
| Lee SJ, 2010 | Gastrocnemius | Follistatin | Fibrosis; myofiber maturation | Down | Negative | Impair | [253] |
| Rinaldi F, 2016 | Tibialis anterior | GDF11 | Collagen deposition | Up | Positive | No effects | [254] |
| Mignemi NA, 2017 | Posterior compartments of the lower extremities | Plasmin | Calcification | Down | Negative | Impair | [255] |
| Lee YS, 2013 | Right gastrocnemius | Gasp1 and/or Gasp2 | Calcified fibers and fibrosis | Down | Positive | Impair | [256] |
| Zhao Y, 2009 | Tibialis anterior | —— | Dystrophic calcification | —— | —— | —— | [257] |
| Lounev V Y, 2009 | Quadriceps | Tie2-expressing endothelial precursors | Heterotopic ossification | —— | —— | —— | [258] |
| Drouin G, 2019 | —— | Hypoxic state | Heterotopic ossification | —— | —— | —— | [259] |
| Arsic N, 2004 | Tibialis anterior | VEGF | Apoptosis | Up | Negative | improve | [260] |
| Sinha-Hikim I, 2007 | Gastrocnemius | JNK and iNOS signaling | Cell apoptosis | Down | Negative | improve | [261] |
| Min K, 2017 | Gastrocnemius/ Soleus | MKP-5 | myofiber apoptosis | Down | Negative | improve | [262] |
| Tjondrokoesoemo A, 2016 | Tibialis anterior | serpina3n | Stabilization of myofiber plasm membrane | Up | Positive | Improve | [263] |
Table 5.
Angiogenesis and neurogenesis in CTX-induced injury models.
| Author, Year | Injury Portions | Target Molecule/ Drug | Target Process | Expression | Effects (Positive/Negative) | Regeneration (Impair/Improve) | Ref. |
|---|---|---|---|---|---|---|---|
| Bellamy LM, 2010 | Unilateral tibialis anterior | Angiotensin II | Angiogenesis | Down | Negative | Impair | [266] |
| Ieronimakis N, 2012 | Tibialis anterior, quadriceps, gastrocnemius | Bone marrow-derived cells | Angiogenesis | —— | Positive | Improve | [265] |
| Mellows B, 2017 | Tibialis anterior | Extracellular vesicles-derived from amniotic fluid stem cell | Angiogenesis | —— | Positive | Improve | [267] |
| Hosaka Y, 2002 | Right tibialis anterior | α1-Syntrophin | Hypertrophy and neuromuscular junctions | Down | Positive; Negative | Improve | [268] |
| Daneshvar N, 2020 | Left tibialis anterior | Premature satellite cell activation | Maturation of neuromuscular junctions | Up | Negative | Improve | [269] |
| Kurosaka M, 2021 | Tibialis anterior | M2 macrophage | Motor innervation regeneration | —— | —— | Improve | [219] |
| Sawano S, 2014 | Tibialis anterior | M2 macrophage | Motor innervation regeneration | —— | —— | Improve | [270] |
| Randazzo D, 2019 | Unilaterally tibialis anterior | Tubb6 | Microtubule organization | Up | Negative | —— | [15] |
Table 6.
Single gene in the CTX-induced skeletal muscle injury model.
| Author, Year | Injury Portions | Target Molecule | Expression | Regeneration (Impair/Improve) | Ref. |
|---|---|---|---|---|---|
| Kim DS, 2015 | Left tibialis anterior | TLR2 | Down | Improve | [272] |
| Oikawa S, 2019 | Tibialis anterior | Dicer | Down | Impair | [273] |
| Hiramuki Y, 2015 | Tibialis anterior | Mest | Down | Impair | [274] |
| Norton CR, 2013 | Tibialis anterior | Snai1/Snai3 | Down | No effects | [275] |
| Call JA, 2017 | Left tibialis anterior and left flexor digitorum longus | Ulk1 | Down | —— | [276] |
| Chaturvedi N, 2020 | Left gastrocnemius | S100A1 | Down | —— | [277] |
| Parks CA, 2019 | Left tibialis anterior | Trim33 | Down | No effects | [278] |
| Goetsch SC, 2005 | Gastrocnemius | Filamin C | Up | —— | [279] |
| Wardrop KE, 2011 | Left tibialis anterior | LYVE-1 | Down | —— | [280] |
| Merkulova T, 2000 | Extensor digitorum longus and tibialis anterior | β enolase | Up | —— | [281] |
| Yuasa K, 2008 | Tibialis anterior | miR-206 | Up | —— | [282] |
| Casciola-Rosen L, 2012 | Right anterior tibilias | Aldolase A | Up | —— | [283] |
| Mammen AL, 2011 | Right tibialis anterior | UFD2a | Up | —— | [284] |
| Nakamura K, 2010 | Right gastrocnemius | GNE | Up | —— | [285] |
| Sato Y, 2013 | Left gastrocnemius | Sema3A | Up | —— | [286] |
| Garry, 2000 | Hind limbs | MNF | Down | Impair | [287] |
| Kemp MW, 2009 | Tibialis anterior | Syncoilin | Up | —— | [288] |
| Miura P, 2005 | Right tibialis anterior | Utrophin A | Up | —— | [289] |
| Wang Q, 2022 | Tibialis anterior | Tsukushi | Down | Impair | [290] |
| McCullagh KJ, 2008 | Unilateral tibialis anterior | Syncoilin | Down | No effects | [291] |
| Demonbreun AR, 2010 | Quadriceps or gastrocnemius/soleus | Ferlin | Up | —— | [292] |
| Maeda Y, 2017 | Right tibialis anterior | CXCL12 | Up | Improve | [293] |
| Darabi R, 2008 | Tibialis anterior | Pax3 | Up | Improve | [294] |
| Di Rocco A, 2015 | Tibialis anterior | RARγ | Down | Impair | [295] |
| Bryer SC, 2007 | Extensor digitorum longus and tibialis anterior | uPAR | —— | No effects | [296] |
| Bryan BA, 2005 | Tibialis anterior | GEFT | Up | Improve | [297] |
| Mathes AL, 2011 | Tibialis anterior | TLR-3 | Down | Impair | [298] |
| Wu G, 2010 | Forelimb leg muscle | Chkb | Down | No effects | [299] |
| Yan Z, 2003 | Tibialis anterior | E2f1 | Down | Impair | [5] |
| Fujita R, 2014 | Left tibialis anterior | IL-6R | Down | Improve | [300] |
| Wu G, 2009 | Gastrocnemius | Chkb | Down | Impair | [301] |
| Wada E, 2019 | Tibialis anterior | Emerin and lamin A/C | Down | Impair | [302] |
| Mofarrahi M, 2015 | Unilateral tibialis anterior | Ang-1 | Up | Improve | [303] |
| Gattazzo F, 2014 | Tibialis anterior | Cyclosporin A | Up | Improve | [304] |
| Kim MH, 2011 | Unilaterally tibialis anterior | Akt | Up | Improve | [243] |
| Laziz I, 2007 | Soleus | Spry | Down | —— | [305] |
| Armand AS, 2003 | Unilateral soleus | Follistatin and myostatin | Up/Down | Improve | [306] |
| Li C, 2013 | Right gastrocnemius | Prosaposin | Up | —— | [307] |
Table 7.
Non-SCs in the CTX-induced skeletal muscle injury models.
| Author, Year | Injury Portions | Target Molecule/Drug | Regeneration | Ref |
|---|---|---|---|---|
| Kano K, 2020 | Gastrocnemius | Capillary stem cells | Improve | [315] |
| Liu Y, 2007 | Unilateral tibialis anterior | Flk-1+ AD-MSCs | Improve | [316] |
| Kim JA, 2013 | Left tibialis anterior | hAFS cells transfected with MyoD | Improve | [318] |
| Xuan W, 2021 | Tibialis anterior | Pluripotent stem cells-induced skeletal muscle progenitor cells with givinostat | Improve | [320] |
| Naldaiz-Gastesi N, 2019 | Tibialis anterior | Human cremaster muscle-derived precursor cells | Improve | [321] |
| Mori J, 2008 | Tibialis anterior | CD45+: Sca-1+ hematopoietic stem cells | Improve | [312] |
| Abedi M, 2007 | Tibialis anterior | Hematopoietic stem cells | Improve | [322] |
| Hwang Y, 2014 | Tibialis anterior | Human embryonic stem cells | Improve | [323] |
| Piccoli M, 2012 | Tibialis anterior mice transplanted with bone marrow or amniotic fluid stem cells | Amniotic fluid stem cells | Improve | [324] |
| Yang R, 2010 | Right tibialis anterior | Clones of ectopic stem cells | Improve | [325] |
| Rousseau J, 2010 | EDL | Muscle precursor cells | Improve | [313] |
| Gang EJ, 2009 | Tibialis anterior | Mesenchymal stem cells | Improve | [314] |
| Jung JE, 2017 | Gastrocnemius and masseter | Pulp-derived cell | Improve | [310] |
| de la Garza-Rodea AS, 2011 | Tibialis anterior | BM-hMSCs | improve | [311] |
| Bossolasco, 2004 | Tibialis anterior | Human adult BM | Improve | [326] |
| Ma, 2012 | Tibialis anterior | Human AF-amniotic fluid stem cells | Improve | [317] |
| Fukada S, 2002 | Tibialis anterior | Bone marrow and fetal liver cells | Improve | [327] |
| Luth ES, 2008 | Tibialis anterior, quadriceps, and gastrocnemius | Bone marrow side population cells | Improve | [328] |
| Zheng JK, 2006 | Tibialis anterior | Human embryonic stem cells | Improve | [329] |
| Cížková D, 2011 | Right tibialis anterior | BMCs | Improve | [308] |
| Meeson AP, 2004 | Hindlimbs | Skeletal muscle side population | Improve | [330] |
| Drapeau C, 2010 | Right tibialis anterior | Mobilization of bone marrow stem cells | Improve | [319] |
| Kowalski K, 2016 | Gastrocnemius | Sdf-1 and granulocyte-colony stimulating factor | Improve | [331] |
| Mitchell R, 2019 | Right tibialis anterior | ADSC secretome | Improve | [332] |
| Tobin S, 2021 | Tibialis anterior; quadriceps; gastrocnemius | Young/aging macrophages | Improve/impair | [333] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.; Lu, J.; Liu, Y. Skeletal Muscle Regeneration in Cardiotoxin-Induced Muscle Injury Models. Int. J. Mol. Sci. 2022, 23, 13380. https://doi.org/10.3390/ijms232113380
AMA Style
Wang Y, Lu J, Liu Y. Skeletal Muscle Regeneration in Cardiotoxin-Induced Muscle Injury Models. International Journal of Molecular Sciences. 2022; 23(21):13380. https://doi.org/10.3390/ijms232113380
Chicago/Turabian StyleWang, Yanjie, Jianqiang Lu, and Yujian Liu. 2022. "Skeletal Muscle Regeneration in Cardiotoxin-Induced Muscle Injury Models" International Journal of Molecular Sciences 23, no. 21: 13380. https://doi.org/10.3390/ijms232113380
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.