
Clinical and Genetic Characteristics of Pediatric Patients with Hypophosphatasia in the Russian Population

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
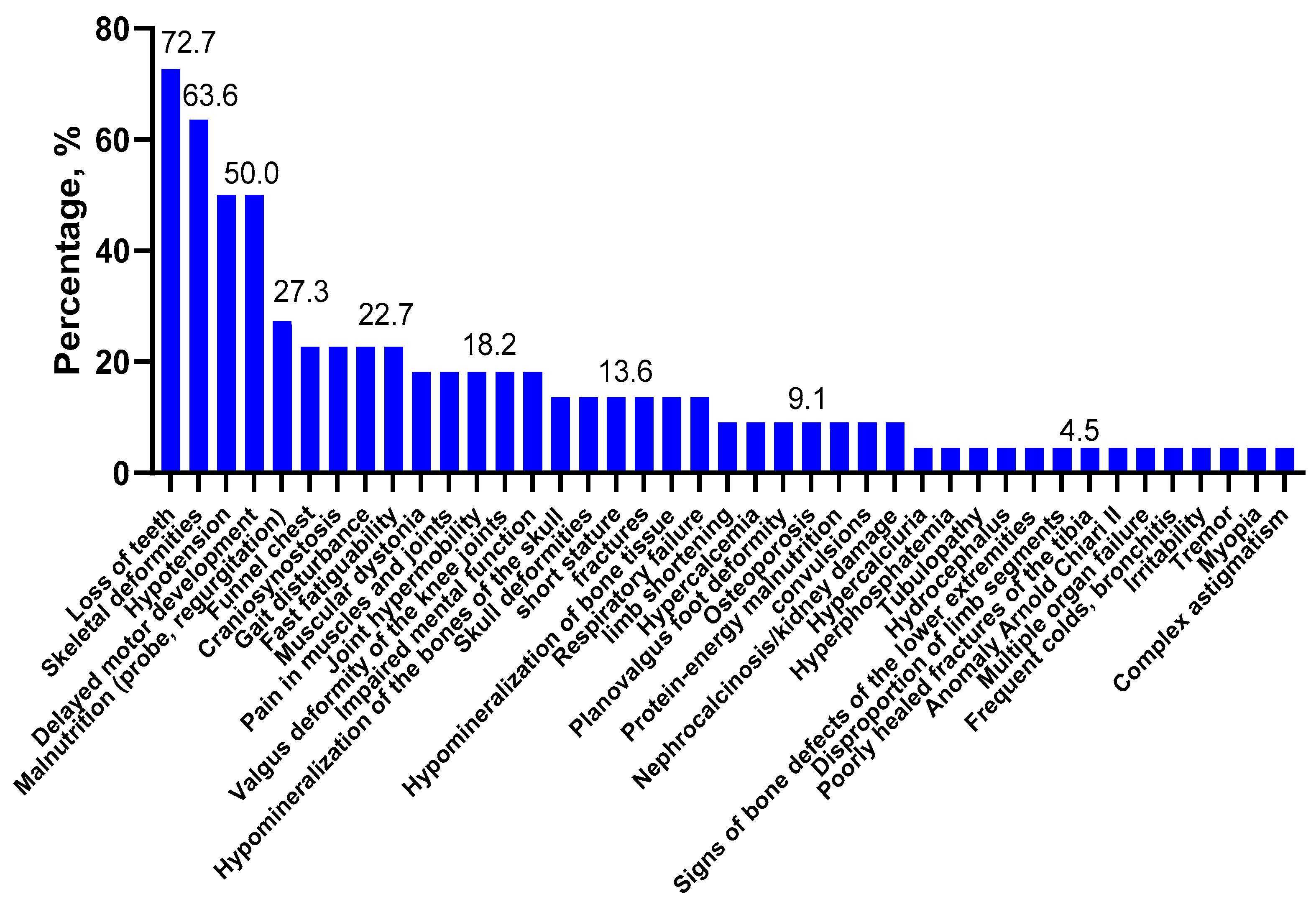
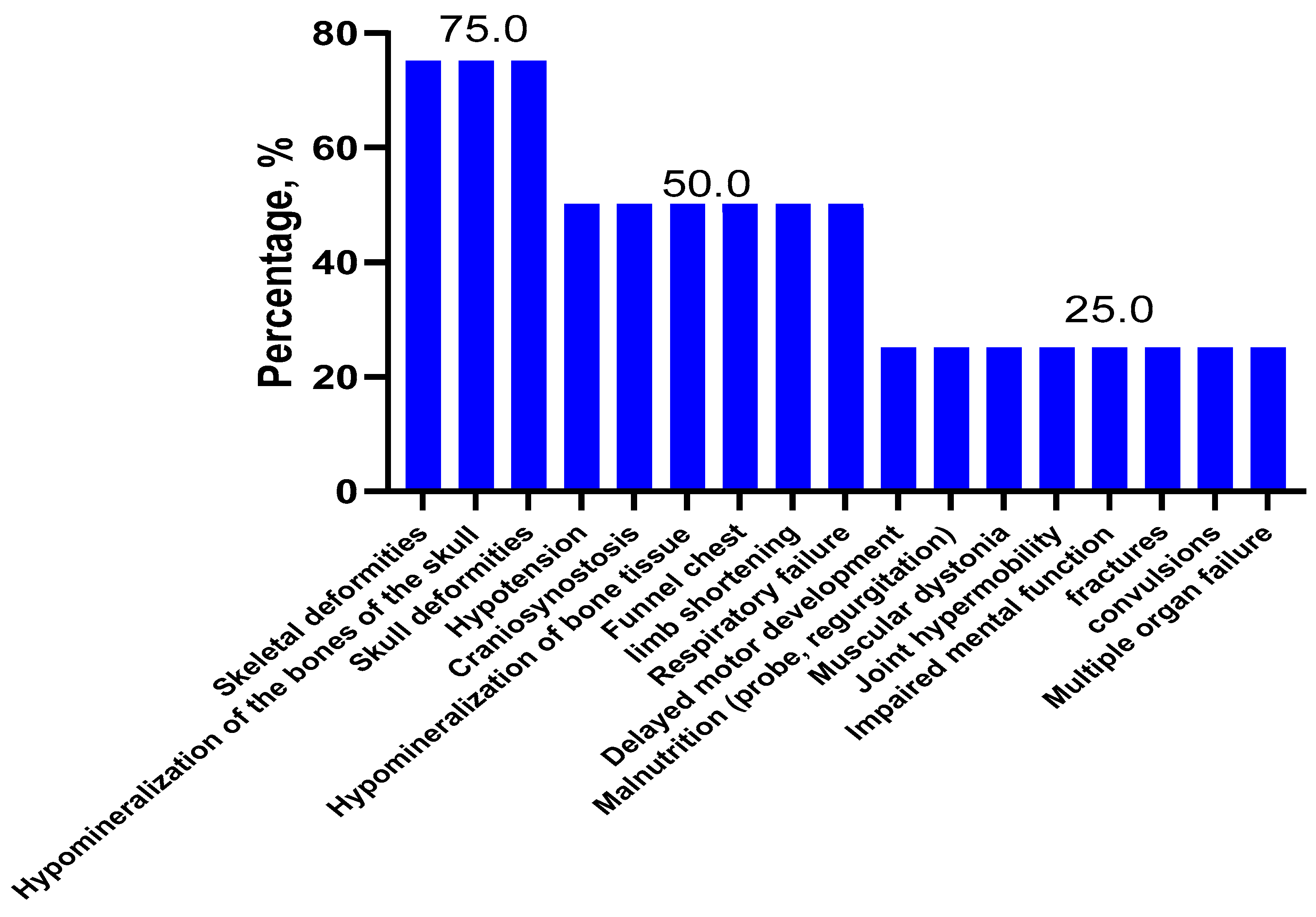
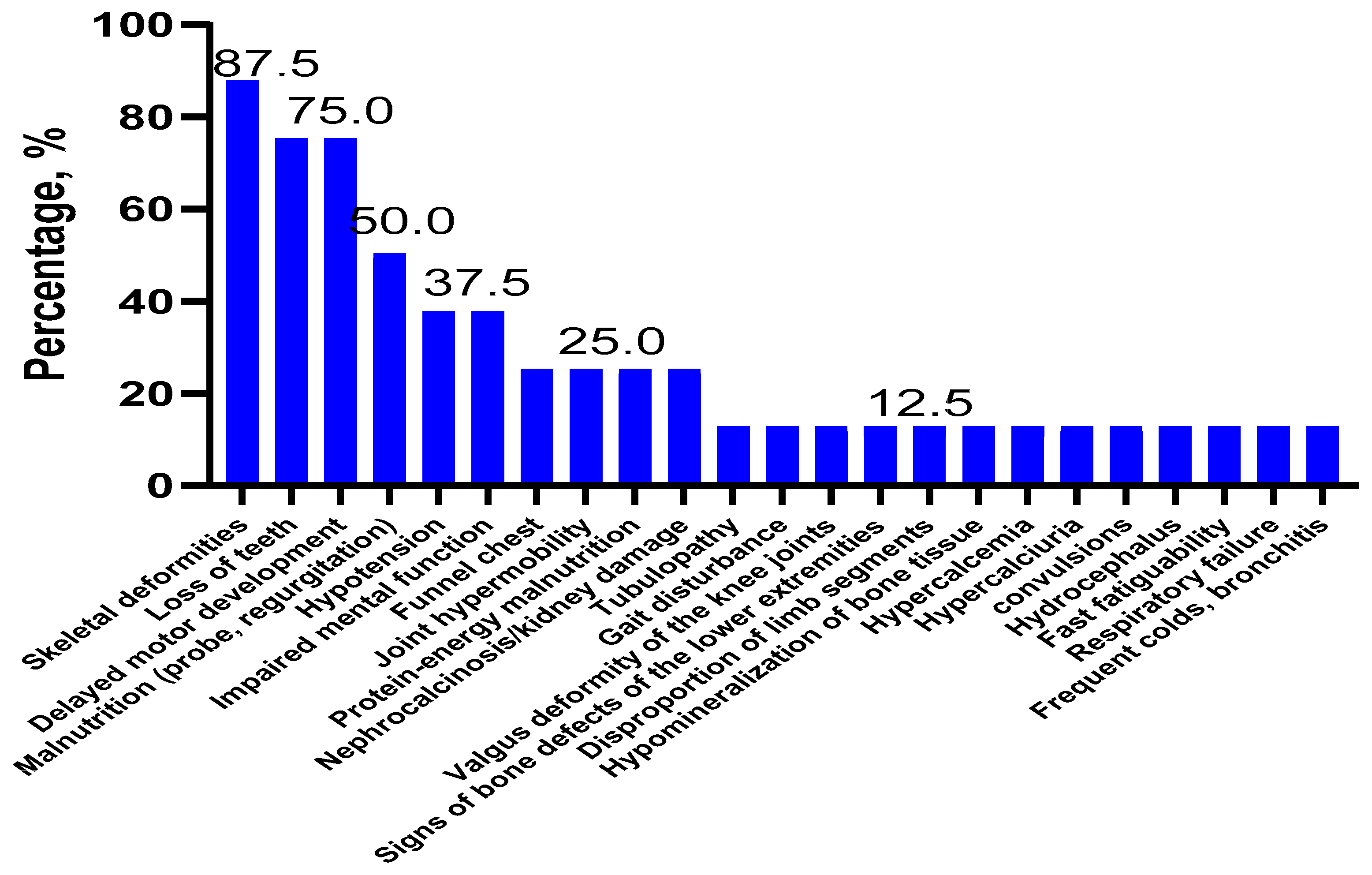
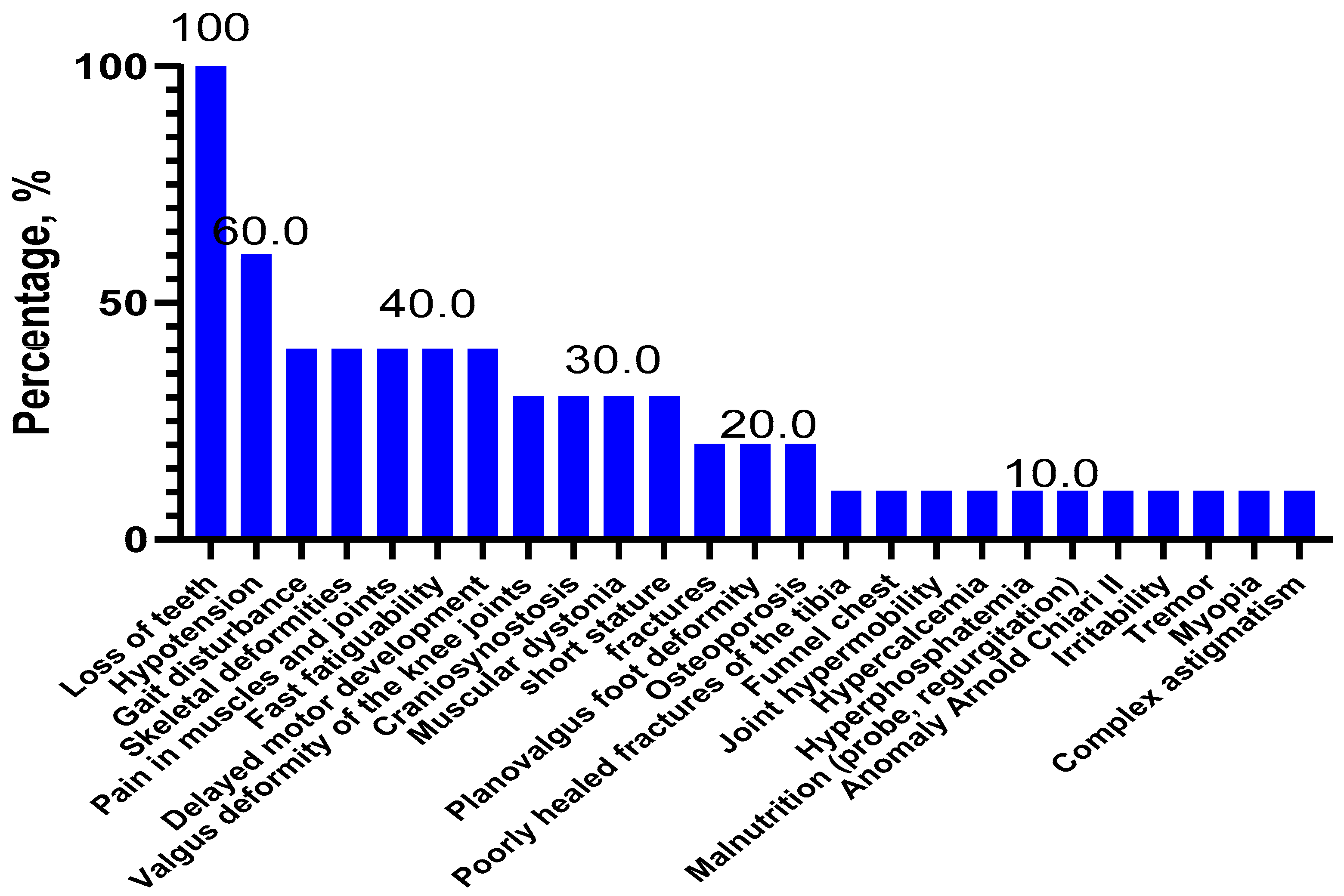
2.1. Clinical Features and Forms of Hypophosphatasia
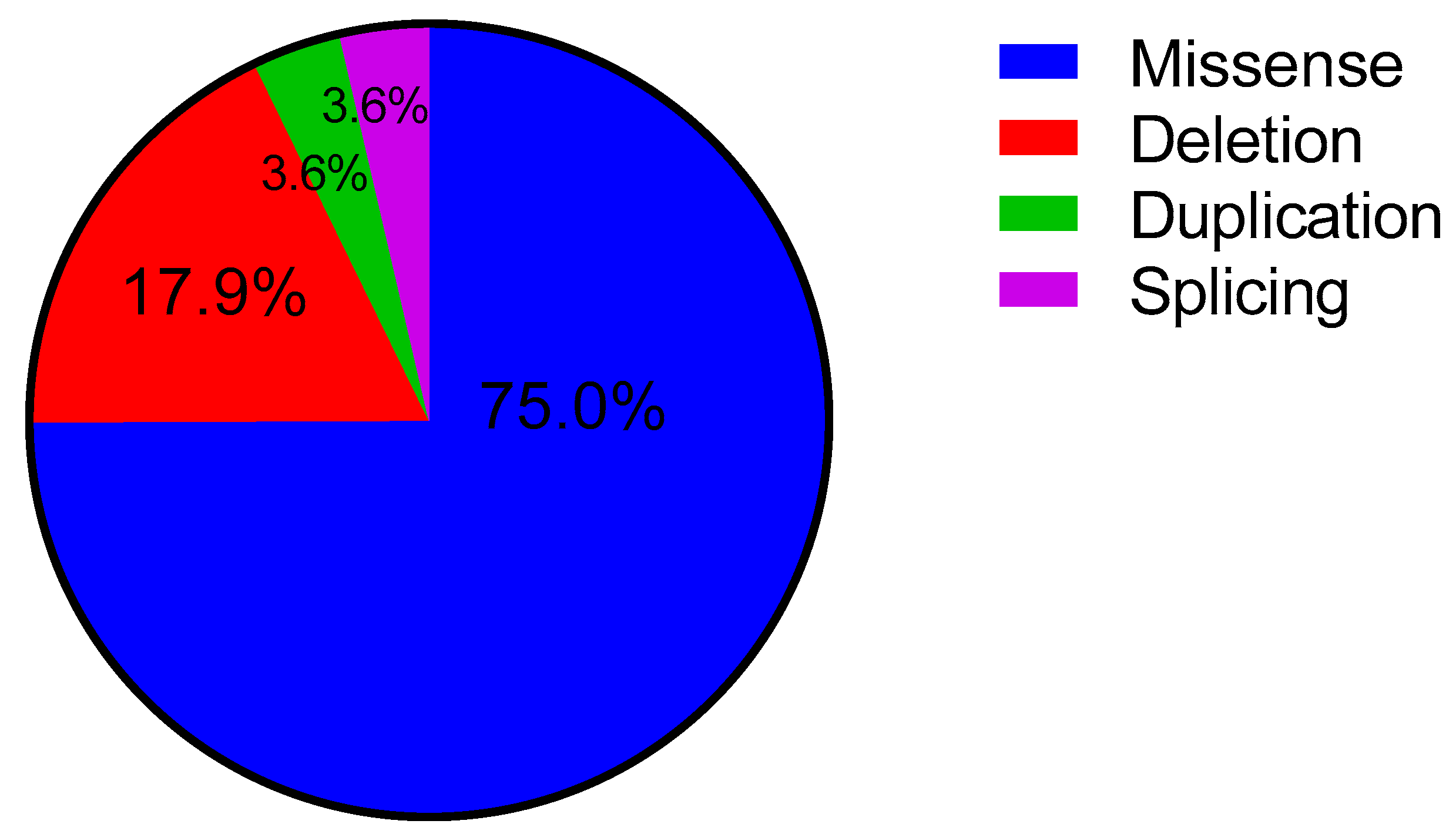
2.2. Variant Frequencies in the ALPL Gene at HPP
2.3. Analysis of the Variants Depending on Type and Localization in the ALPL Gene
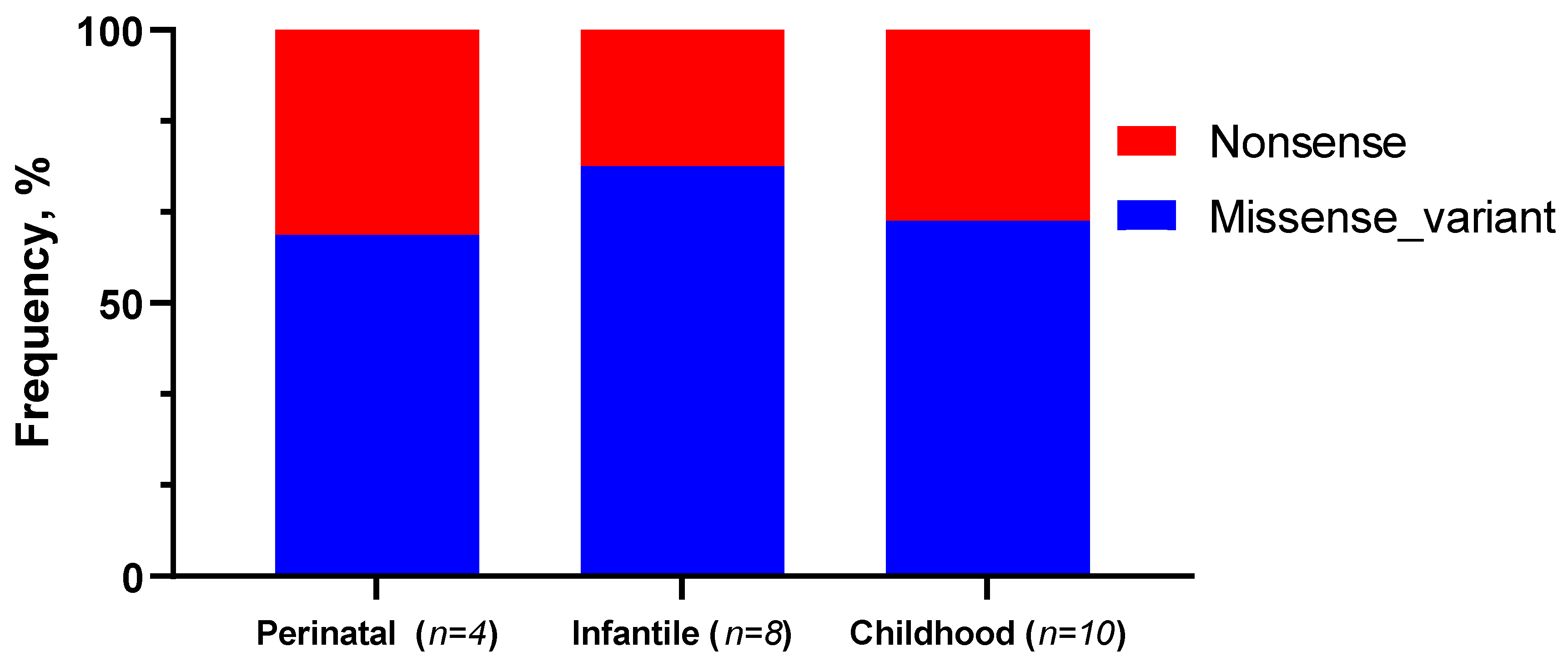
2.4. Analysis of ALPL Gene Variants Depending on the Form of HPP and Clinical Symptoms in Patients
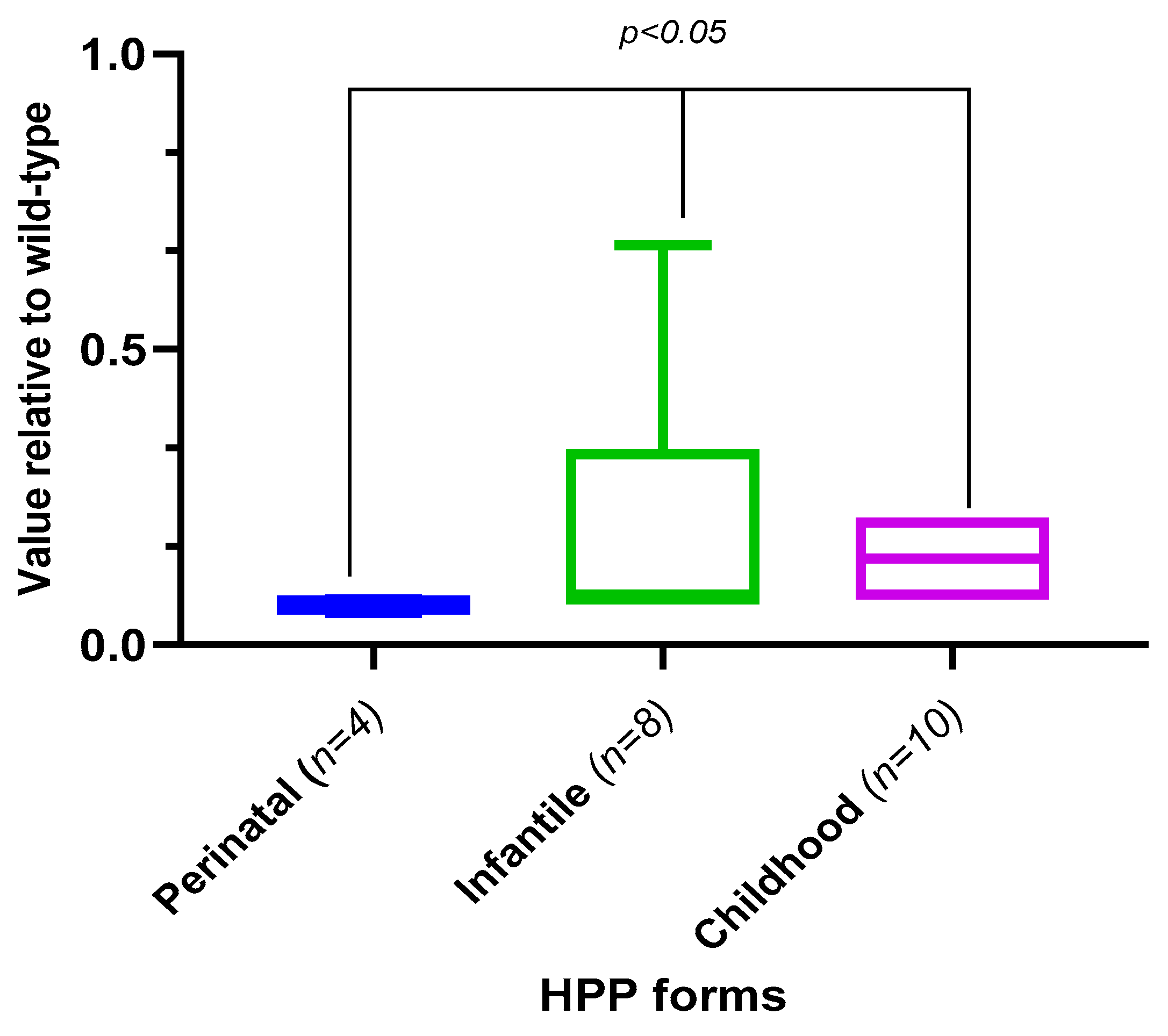
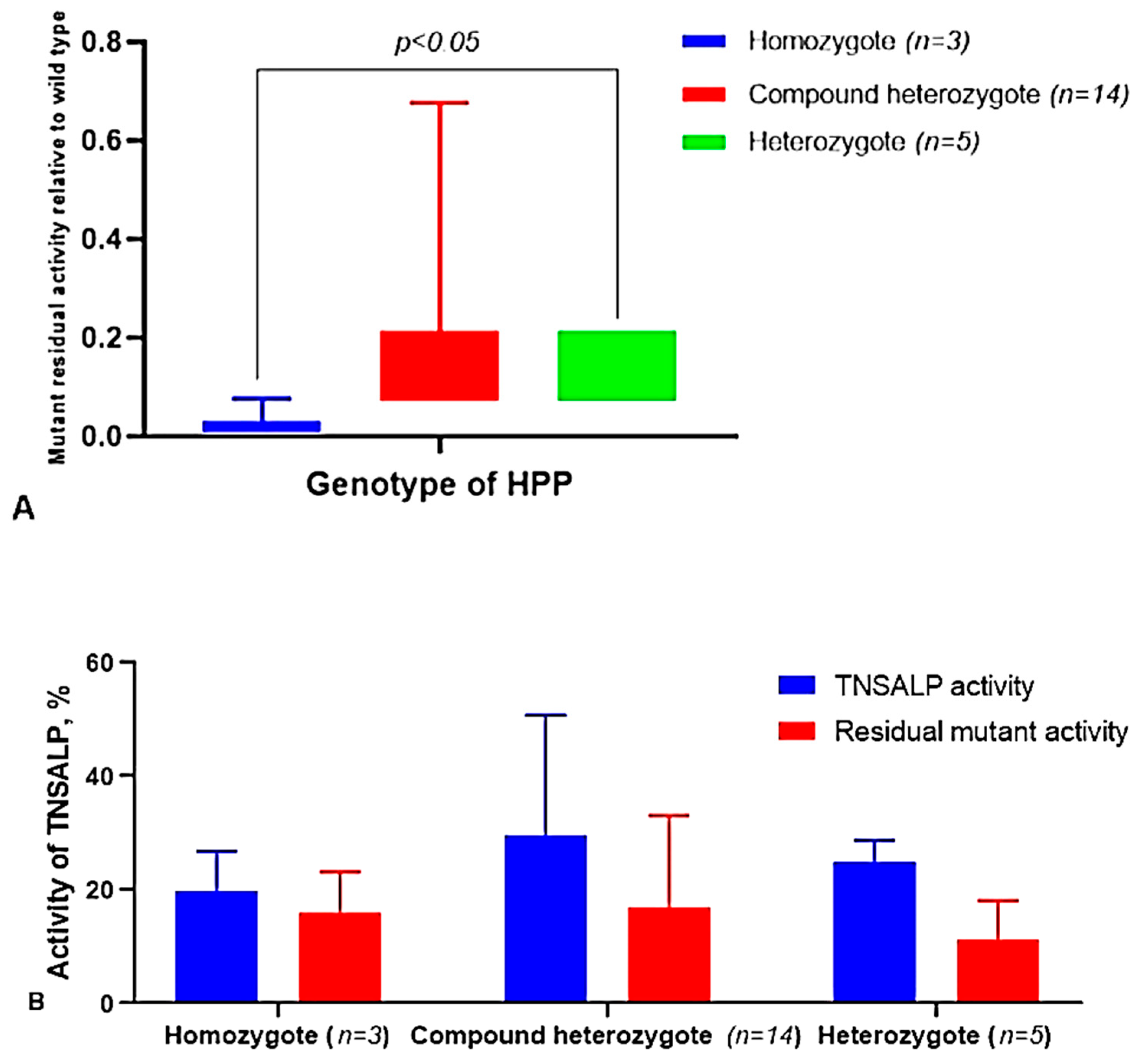
2.5. Analysis of TNSALP Activity Depending on the Form, Type of Variants, and the Genotype of the Patients
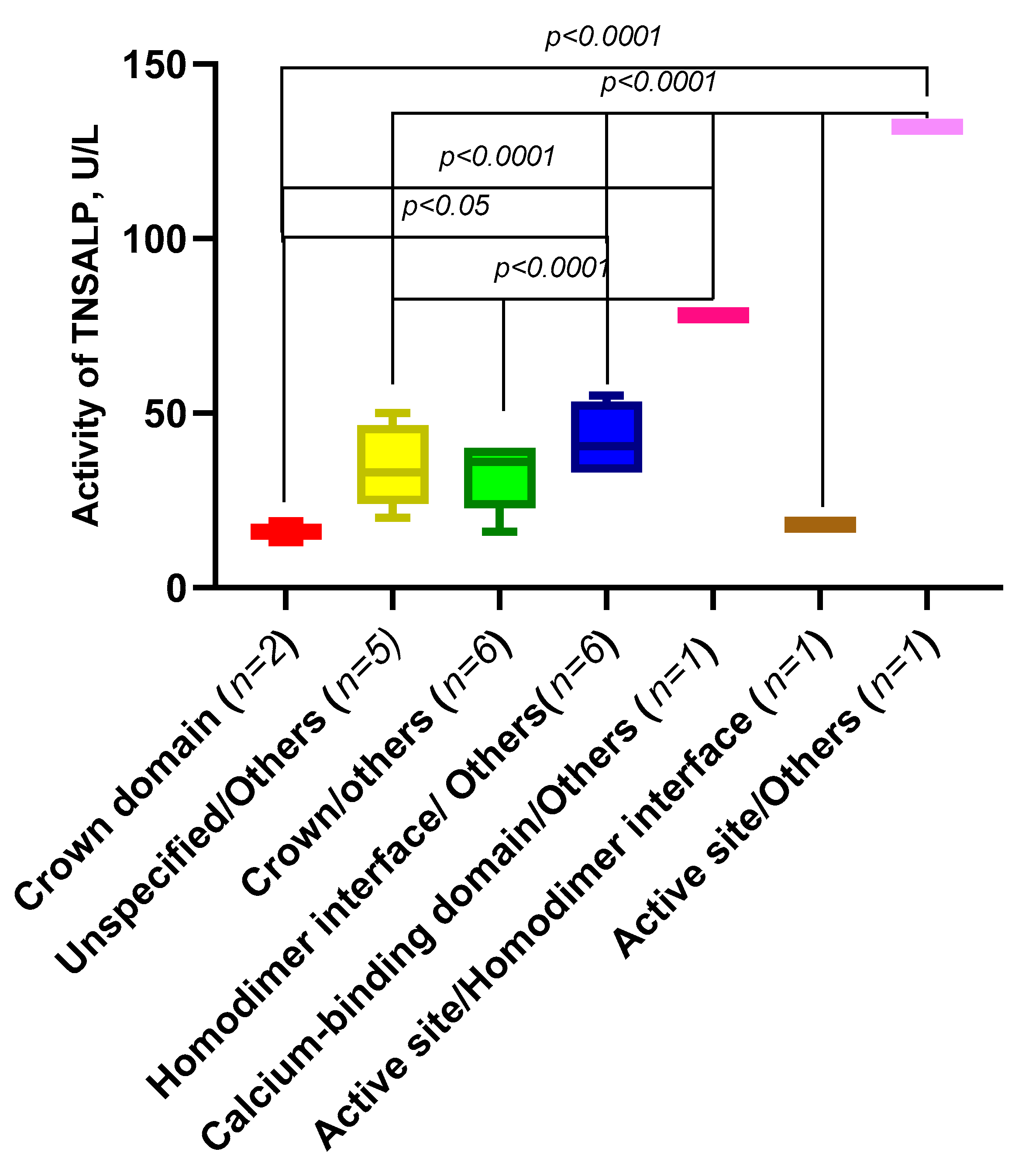
2.6. Analysis of the Activity of the ALPL Gene Variants Depending on Their Domain Localization and Origin
3. Discussion
4. Materials and Methods
4.1. Patients with Hypophosphatasia
4.2. Biochemical Studies
4.3. Molecular Genetic Analysis of a ALPL Gene
4.4. Predicting Theoretical Activity of Patient Genotypes
4.5. Statistical Analysis
4.6. Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Fenn, J.S.; Lorde, N.; Martin Ward, J.; Borovickova, I. Hypophosphatasia. J. Clin. Pathol. 2021, 74, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Conti, F.; Ciullini, L.; Pugliese, G. Hypophosphatasia: Clinical manifestation and burden of disease in adult patients. Clin. Cases Miner. Bone Metab. 2017, 14, 230–234. [Google Scholar] [CrossRef] [PubMed]
- RUSeq. Available online: http://ruseq.ru/#/gene/249 (accessed on 20 September 2022).
- Rathbun, J.C. Hypophosphatasia; a new developmental anomaly. Am. J. Dis. Child. 1948, 75, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.J.; Cole, D.E.; Ray, K.; Whyte, M.P.; Lafferty, M.A.; Mulivor, R.A.; Harris, H. A missense mutation in the human liver/bone/kidney alkaline phosphatase gene causing a lethal form of hypophosphatasia. Proc. Natl. Acad. Sci. USA 1988, 85, 7666–7669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whyte, M.P.; Zhang, F.; Wenkert, D.; McAlister, W.H.; Mack, K.E.; Benigno, M.C.; Coburn, S.P.; Wagy, S.; Griffin, D.M.; Ericson, K.L.; et al. Hypophosphatasia: Validation and expansion of the clinical nosology for children from 25 years experience with 173 pediatric patients. Bone 2015, 75, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Liu, S.; Lin, Y.; Chen, Z.; Shao, Y.; Yu, Q.; Liu, H.; Lu, Z.; Sheng, H.; Lu, X.; et al. Two novel mutations in the ALPL gene of unrelated Chinese children with Hypophosphatasia: Case reports and literature review. BMC Pediatr. 2019, 19, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangura, A.; Wright, L.; Shuler, T. Hypophosphatasia: Current Literature for Pathophysiology, Clinical Manifestations, Diagnosis, and Treatment. Cureus 2020, 12, e8594. [Google Scholar] [CrossRef]
- Ignatovich, O.N.; Namazova-Baranova, L.S.; Margieva, T.V.; Yakhyaeva, G.T.; Zhurkova, N.V.; Savostyanov, K.V.; Pushkov, A.A.; Krotov, I.A. Osteogenesis Imperfecta: Diagnostic Feature. Pediatr. Pharmacol. 2018, 15, 224–232. [Google Scholar] [CrossRef]
- García-Fontana, C.M.; Villa-Suárez, J.; Andújar-Vera, F.; González-Salvatierra, S.; Martínez-Navajas, G.; Real, P.J.; Gómez Vida, J.M.; de Haro, T.; García-Fontana, B.; Muñoz-Torres, M. Epidemiological, Clinical and Genetic Study of Hypophosphatasia in A Spanish Population: Identification of Two Novel Mutations in The Alpl Gene. Sci Rep. 2019, 9, 9569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranov, A.A.; Namazova-Baranova, L.S.; Savostianov, K.V.; Margieva, T.V.; Vishneva, E.A.; Yakhyaeva, G.T. CLinical Recomendation to the Diagnostics and Treatment of Hypophosphatasia in Children. Pediatr. Pharmacol. 2016, 13, 539–543. (In Russian) [Google Scholar] [CrossRef]
- Martos-Moreno, G.A.; Calzada, J.; Couce, M.L.; Argente, J. Hypophosphatasia: Clinical manifestations, diagnostic recommendations and therapeutic options. An. Pediatr. (Engl. Ed.) 2018, 88, e1–e356. [Google Scholar] [CrossRef]
- Collmann, H.; Mornet, E.; Gattenlöhner, S.; Beck, C.; Girschick, H. Neurosurgical aspects of childhood hypophosphatasia. Childs Nerv. Syst. 2009, 25, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Kishnani, P.S.; Rush, E.T.; Arundel, P.; Bishop, N.; Dahir, K.; Fraser, W.; Harmatz, P.; Linglart, A.; Munns, C.F.; Nunes, M.E.; et al. Monitoring guidance for patients with hypophosphatasia treated with asfotase alfa. Mol. Genet. Metab. 2017, 122, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Mornet, E. Genetics of hypophosphatasia. Arch. Pediatr. 2017, 24, 5s51–5s56. [Google Scholar] [CrossRef]
- Mornet, E.; Hofmann, C.; Bloch-Zupan, A.; Girschick, H.; Le Merrer, M. Clinical utility gene card for: Hypophosphatasia—Update 2013. Eur. J. Hum. Genet. 2014, 22, 572. [Google Scholar] [CrossRef] [PubMed]
- Global Variome Shared LOVD ALPL (Alkaline Phosphatase, Liver/Bone/Kidney). Available online: https://databases.lovd.nl/shared/variants/ALPL/unique (accessed on 20 September 2022).
- Liu, M.; Liu, M.; Liang, X.; Wu, D.; Li, W.; Su, C.; Cao, B.; Chen, J.; Gong, C. Clinical and genetic characteristics of hypophosphatasia in Chinese children. Orphanet. J. Rare Dis. 2021, 16, 159. [Google Scholar] [CrossRef]
- Brun-Heath, I.; Lia-Baldini, A.S.; Maillard, S.; Taillandier, A.; Utsch, B.; Nunes, M.E.; Serre, J.L.; Mornet, E. Delayed transport of tissue-nonspecific alkaline phosphatase with missense mutations causing hypophosphatasia. Eur. J. Med. Genet. 2007, 50, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Mornet, E. Molecular genetics of Hypophosphatasia and phenotype-genotype correlations. Subcell. Biochem. 2015, 76, 25–43. [Google Scholar] [CrossRef]
- del Angel, G.; Reynders, J.; Negron, C.; Steinbrecher, T.; Mornet, E. Large-scale in vitro functional testing and novel variant scoring via protein modeling provide insights into alkaline phosphatase activity in hypophosphatasia. Hum. Mutat. 2020, 41, 1250–1262. [Google Scholar] [CrossRef] [Green Version]
- Whyte, M.P.; Greenberg, C.R.; Salman, N.J.; Bober, M.B.; McAlister, W.H.; Wenkert, D.; Van Sickle, B.J.; Simmons, J.H.; Edgar, T.S.; Bauer, M.L.; et al. Enzyme-replacement therapy in life-threatening hypophosphatasia. N. Engl. J. Med. 2012, 366, 904–913. [Google Scholar] [CrossRef]
- Adeli, K.; Higgins, V.; Trajcevski, K.; White-Al Habeeb, N. The Canadian laboratory initiative on pediatric reference intervals: A CALIPER white paper. Crit. Rev. Clin. Lab. Sci. 2017, 54, 358–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Högler, W.; Langman, C.; da Silva, H.G.; Fang, S.; Linglart, A.; Ozono, K.; Petryk, A.; Rockman-Greenberg, C.; Seefried, L.; Kishnani, P.S. Diagnostic delay is common among patients with hypophosphatasia: Initial findings from a longitudinal, prospective, global registry. BMC Musculoskelet. Disord. 2019, 20, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, S.M.; Tomazos, I.C.; Petryk, A.; Powell, L.C.; Donato, B.M.K.; Zarate, Y.A.; Tiulpakov, A.; Martos-Moreno, G.A. Frequency and age at occurrence of clinical manifestations of disease in patients with hypophosphatasia: A systematic literature review. Orphanet. J.Rare Dis. 2019, 14, 85. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, C.; Girschick, H.; Mornet, E.; Schneider, D.; Jakob, F.; Mentrup, B. Unexpected high intrafamilial phenotypic variability observed in hypophosphatasia. Eur. J. Hym. Genet. 2014, 22, 1160–1164. [Google Scholar] [CrossRef]
- Greenberg, C.R.; Evans, J.A.; McKendry-Smith, S.; Redekopp, S.; Haworth, J.C.; Mulivor, R.; Chodirker, B.N. Infantile hypophosphatasia: Localization within chromosome region 1p36.1-34 and prenatal diagnosis using linked DNA markers. Am. J. Hum. Genet. 1990, 46, 286–292. [Google Scholar]
- Genome Aggregation Database. Available online: https://gnomad.broadinstitute.org/ (accessed on 20 September 2022).
- Okawa, R.; Kokomoto, K.; Kitaoka, T.; Kubota, T.; Watanabe, A.; Taketani, T.; Michigami, T.; Ozono, K.; Nakano, K. Japanese nationwide survey of hypophosphatasia reveals prominent differences in genetic and dental findings between odonto and non-odonto types. PLoS ONE 2019, 14, e0222931. [Google Scholar] [CrossRef] [PubMed]
- Michigami, T.; Tachikawa, K.; Yamazaki, M.; Kawai, M.; Kubota, T.; Ozono, K. Hypophosphatasia in Japan: ALPL mutation analysis in 98 unrelated patients. Calcif. Tissue Int. 2020, 106, 221–231. [Google Scholar] [CrossRef]
- Taillandier, A.; Domingues, C.; Dufour, A.; Debiais, F.; Guggenbuhl, P.; Roux, C.; Cormier, C.; Cortet, B.; Porquet-Bordes, V.; Coury, F.; et al. Genetic analysis of adults heterozygous for ALPL mutations. J. Bone Miner. Metab. 2018, 36, 723–733. [Google Scholar] [CrossRef]
- Mornet, E.; Yvard, A.; Taillandier, A.; Fauvert, D.; Simon-Bouy, B. A molecular-based estimation of the prevalence of hypophosphatasia in the European population. Ann. Hum. Genet. 2011, 75, 439–445. [Google Scholar] [CrossRef]
- Landrum, M.J.; Lee, J.M.; Benson, M.; Brown, G.R.; Chao, C.; Chitipiralla, S.; Gu, B.; Hart, J.; Hoffman, D.; Jang, W.; et al. ClinVar: Improving access to variant interpretations and supporting evidence. Nucleic Acids Res. 2018, 46, D1062–D1067. [Google Scholar] [CrossRef] [Green Version]
- Brun-Heath, I.; Taillandier, A.; Serre, J.-L.; Mornet, E. Characterization of 11 novel mutations in the tissue non-specific alkaline phosphatase gene responsible for hypophosphatasia and genotype-phenotype correlations. Mol. Genet. Metab. 2005, 84, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Lia-Baldini, A.S.; Brun-Heath, I.; Carrion, C.; Simon-Bouy, B.; Serre, J.L.; Nunes, M.E.; Mornet, E. A new mechanism of dominance in hypophosphatasia: The mutated protein can disturb the cell localization of the wild-type protein. Hum. Genet. 2008, 123, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Schumann, G.; Klauke, R.; Canalias, F.; Bossert-Reuther, S.; Franck, P.F.H.; Gella, F.-J.; Jørgensen, P.J.; Kang, D.; Lessinger, J.-M.; Panteghini, M.; et al. IFCC primary reference procedures for the measurement of catalytic activity concentrations of enzymes at 37 °C. Part 9: Reference procedure for the measurement of catalytic concentration of alkaline phosphatase International Federation of Clinical Chemistry and Laboratory Medicine (IFCC) Scientific Division, Committee on Reference Systems of Enzymes (C-RSE) (1)). Clin. Chem. Lab. Med. 2011, 49, 1439–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Belle, G.; Fisher, L.D.; Heagerty, P.J.; Lumley, T. Biostatistics: A Methodology for the Health Sciences; Fisher, L.D., van Belle, G., Eds.; Jhon Wiley and Sons Inc.: Hoboken, NJ, USA, 2004. [Google Scholar]
- 1000 Genomes Project Consortium; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; Cummings, B.B.; et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbitoff, Y.A.; Skitchenko, R.K.; Poleshchuk, O.I.; Shikov, A.E.; Serebryakova, E.A.; Nasykhova, Y.A.; Polev, D.E.; Shuvalova, A.R.; Shcherbakova, I.V.; Fedyakov, M.A.; et al. Whole exome sequencing provides insights into monogenic disease prevalence in Northwest Russia. Mol. Genet. Genom. Med. 2019, 7, e964. [Google Scholar] [CrossRef] [Green Version]
- Barbitoff, Y.A.; Khmelkova, D.N.; Pomerantseva, E.A.; Slepchenkov, A.V.; Zubashenko, N.A.; Mironova, I.V.; Kaimonov, V.S.; Polev, D.E.; Tsay, V.V.; Glotov, A.S.; et al. Expanding the Russian allele frequency reference via cross-laboratory data integration: Insights from 6096 exome samples. MedRXiv 2021. [Google Scholar] [CrossRef]
- Shikov, A.E.; Barbitoff, Y.A.; Glotov, A.S.; Danilova, M.M.; Tonyan, Z.N.; Nasykhova, Y.A.; Mikhailova, A.A.; Bespalova, O.N.; Kalinin, R.S.; Mirzorustamova, A.M.; et al. Analysis of the Spectrum of ACE2 Variation Suggests a Possible Influence of Rare and Common Variants on Susceptibility to COVID-19 and Severity of Outcome. Front. Genet. 2020, 11, 551220. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HPP Onset | Number of Patients | Age at the Time of Diagnosis, Years | TNSALP Level in Serum, U/L |
|---|---|---|---|
| Prenatal | 4 | up to 1 year | 18.7 ± 6.0 |
| Infantile | 8 | 2.0 ± 1.2 (1–4) | 31.6 ± 10.8 |
| Childhood | 10 | 5.3 ± 5.0 (1–18) | 40.1 ± 16.5 |
| Allele 1 | Allele 2 | Amino Acid Substitution | Variant Type | Number of Patients | Clinical Varinat Classification |
|---|---|---|---|---|---|
| c.182G > A | c.571G > A | Gly61Glu/Glu191Lys | Missense/Missense | 1 | Pathogenic/Pathogenic |
| c.211C > T | c.1101_1103del | Arg71Cys/Ser368del | Missense/In frame | 1 | Pathogenic/Pathogenic |
| c.228G > T | c.571G > A | Gln76His/Glu191Lys | Missense/Missense | 1 | Pathogenic/Pathogenic |
| c.253A > C | c.571G > A | Thr85Pro/Glu191Lys | Missense/Missense | 1 | VUS/Pathogenic |
| c.302A > G | c.571G > A | Tyr101Cys/Glu191Lys | Missense/Missense | 1 | VUS/Pathogenic |
| c.303C > A | c.314C > T | Tyr101Ter/Ala105Val | Nonsense/Missense | 1 | Pathogenic/VUS |
| c.303C > A | c.571G > A | Tyr101Ter/Glu191Lys | Nonsense/Missense | 1 | Pathogenic/Pathogenic |
| c.340G > A | c.571G > A | Ala114Thr/Glu191Lys | Missense/Missense | 1 | Pathogenic/Pathogenic |
| c.382G > A | c.871G > C | Val128Met/Glu291Gln | Missense/Missense | 1 | Pathogenic/VUS |
| c.508A > G | c.508A > G | Asn170Asp | Missense | 1 | Pathogenic |
| c.526G > A | c.1375G > A | Ala176Thr/Val459Met | Missense/Missense | 1 | Pathogenic/Pathogenic |
| c.571G > A | c.144_148dup | Glu191Lys/ | Missense/Frameshift | 1 | Pathogenic/VUS |
| Val50AlafsTer20 | |||||
| c.571G > A | c.662delG | Glu191Lys/ | Missense//Frameshift | 2 | Pathogenic/Pathogenic |
| Gly221Valfs*56 | |||||
| c.571G > A | c.1259G > T | Glu191Lys/Gly420Val | Missense/Missence | 1 | Pathogenic/Pathogenic |
| c.571G > A | c.1157_1175inv | Glu191Lys/GGYTPRG386_392ATGGVST | Missense/Frameshift | 1 | Pathogenic//Pathogenic |
| c.571G > A | c.1309 + 2T > C | Glu191Lys/ | Missense/Splicing | 2 | Pathogenic/Pathogenic |
| c.984_986delCT | c.371A > G | Phe328del/Asn124Ser | In frame/Missense | 1 | Pathogenic/VUS |
| c.1112C > A | c.1447G > A | Thr371Asn/Val483Met | Missense/Missense | 2 | Pathogenic/VUS |
| c.1163A > G | c.1163A > G | Tyr388Cys | Missense | 1 | VUS |
| c.1171del | c.1171del | Arg391ValfsTer12 | Frameshift | 1 | Pathogenic |
| c.1171delC | c.302A > G | Arg391Valfs*12/ | Fameshift/Missense | 1 | Pathogenic/VUS |
| Tyr101Cys | |||||
| c.1171delC | c.571G > A | Arg391Valfs*12/Glu191Lys | Fameshift/Missense | 1 | Pathogenic/Pathogenic |
| c.1349G > A | c.1349G > A | Arg450His | Missense | 1 | Pathogenic |
| c.1364G > A | c.571G > A | Glu455Asp/Glu191Lys | Missense/Missense | 1 | Pathogenic/Pathogenic |
| Variant | Amino Acid Substitution | Variant Type | Allelic | Clinical Variants |
|---|---|---|---|---|
| Frequency, % | Interpretation | |||
| c.61G > A | Glu21Lys | Missense | 0.22 (1/450) | Pathogenic |
| c.98C > T | Ala33Val | Missense | 0.22 (1/450) | Pathogenic |
| c.119C > T | Ala40Val | Missense | 0.22 (1/450) | Pathogenic |
| c.144_148dup | Val50Alafs*20 | Frameshift | 0.22 (1/450) | Pathogenic |
| c.182G > A | Gly61Glu | Missense | 0.22 (1/450) | VUS |
| c.188G > T | Gly63Val | Missense | 0.44 (2/450) | Pathogenic |
| c.202_204delACG | Thr68del | In frame | 0.22 (1/450) | VUS |
| c.203C > T | Thr68Met | Missense | 0.44 (2/450) | Pathogenic |
| c.205G > A | Ala69Thr | Missense | 1.11 (5/450) | VUS |
| c. 211C > T | Arg71Cys | Missense | 0.67 (3/450) | Pathogenic |
| c.211C > A | p.Arg71Ser | Missense | 0.22 (1/450) | Pathogenic |
| c.212G > A | p.Arg71His | Missense | 0.22 (1/450) | Pathogenic |
| c.214A > G | Ile72Val | Missense | 0.22 (1/450) | Likely Pathogenic |
| c.216C > A | p.(=) | Synonymous | 0.22 (1/450) | VUS |
| c.250G > A | Glu84Lys | Missense | 0.22 (1/450) | Pathogenic |
| c.253A > C | Thr85Pro | Missense | 0.22 (1/450) | VUS |
| c.297 + 1G > A | - | Splicing | 0.44 (2/450) | Pathogenic |
| c.298-2A > G | - | Splicing | 0.22 (1/450) | Pathogenic |
| c.302A > G | Tyr101Cys | Missense | 0.22 (1/450) | VUS |
| c.303C > A | Tyr101Ter | Nonsense | 1.11 (5/450) | Pathogenic |
| c.305A > C | Asn102Thr | Missense | 0.44 (2/450) | VUS |
| c.314C > T | Ala105Val | Missense | 0.22 (1/450) | VUS |
| c.331_332insCCGGCA | p.T113_A114insGT | In frame | 0.22 (1/450) | VUS |
| c.368C > A | Ala123Asp | Missense | 0.44 (2/450) | Pathogenic |
| c.371A > G | Asn124Ser | Missense | 0.22 (1/450) | VUS |
| c.389del | Val130GlufsTer35 | Frameshift | 0.22 (1/450) | Pathogenic |
| c.407G > A | Arg136His | Missense | 0.22 (1/450) | Pathogenic |
| c.436G > A | Glu146Lys | Missense | 0.67 (3/450) | VUS |
| c.455G > A | Arg152His | Missense | 3.33 (15/450) | VUS |
| c.500C > T | Thr167Met | Missense | 0.22 (1/450) | Pathogenic |
| c.508A > G | Asn170Asp | Missense | 0.44 (2/450) | Pathogenic |
| c.571G > A | Glu191Lys | Missense | 12.7 (57/450) | Pathogenic |
| c.595C > T | Gly199* | Nonsense | 0.44 (2/450) | Pathogenic |
| c.648 + 1G > A | Splicing | 0.22 (1/450) | Pathogenic | |
| c.659G > T | Gly220Ala | Missense | 0.67 (3/450) | Pathogenic |
| c.662_663insG | Gly222Trpfs | Frameshift | 0.44 (2/450) | Pathogenic |
| c.815G > T | Arg272Leu | Missense | 0.22 (1/450) | Pathogenic |
| c.818C > T | Thr273Met | Missense | 0.88 (4/450) | VUS |
| c.889T > A | Tyr297Asn | Missense | 0.44 (2/450) | VUS |
| c.902delG | Asn302Thrfs | Frameshift | 0.67 (3/450) | Pathogenic |
| c.928_929delTC | Ser310Argfs | Frameshift | 0.44 (2/450) | Pathogenic |
| c.968A > T | Asn323Ile | Missense | 0.22 (1/450) | VUS |
| c.979T > C | Phe327Leu | Missense | 0.22 (1/450) | Pathogenic |
| c.984_986delCTT | Phe328del | In frame | 0.22 (1/450) | Pathogenic |
| c.997 + 2T > A | - | Splicing | 0.44 (2/450) | Pathogenic |
| c.998-1G > A | - | Splicing | 0.67 (3/450) | Pathogenic |
| c.1001G > A | Gly334Asp | Missense | 0.44 (1/225) | Pathogenic |
| c.1066G > A | Asp356Asn | Missense | 0.22 (1/450) | VUS |
| c.1072G > A | Ala358Thr | Missense | 0.22 (1/450) | VUS |
| c.1101_1103del | Ser368del | In frame | 0.22 (1/450) | Pathogenic |
| c.1130C > T | Ala377Val | Missense | 0.22 (1/450) | Pathogenic |
| c.1163A > G | Tyr388Cys | Missense | 0.22 (1/450) | VUS |
| c. 1171del (16) | Arg391Valfs*12 | Frameshift | 3.11 (14/450) | Pathogenic |
| c. 1190-2_1190-1delinsCT | - | Splicing | 0.44 (2/450) | Pathogenic |
| c.1216G > A | Asp406Asn | Missense | 0.22 (1/450) | VUS |
| c.1247G > T | Gly416Val | Missense | 0.22 (1/450) | VUS |
| c.1259G > T | Gly420Val | Missense | 0.44 (2/450) | VUS |
| c.1276G > A | p.Gly426Ser | Missense | 0.44 (2/450) | Pathogenic |
| c.1309 + 2T > C | - | Splicing | 0.44 (2/450) | Pathogenic |
| c.1310C > T | Ala437Val | Missense | 0.22 (1/450) | VUS |
| c.1328C > T | Ala443Val | Missense | 0.22 (1/450) | Pathogenic |
| c.1349G > A | Arg450His | Missense | 0.67 (3/450) | Pathogenic |
| c.1364G > A | Glu455Asp | Missense | 0.44 (2/450) | Pathogenic |
| c.1447G > A | Val483Met | Missense | 1.11 (5/450) | VUS |
| c.1483G > A | Gly495Ser | Missense | 0.22 (1/450) | VUS |
| c.1489T > C | Cys497Arg | Missense | 0.22 (1/450) | VUS |
| Clinical Symptoms | Patients, Forms of HPP | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Perinatal | Infantile | Childhood | ||||||||||||||||||||
| Loss of teeth | ||||||||||||||||||||||
| Skeletal deformities | ||||||||||||||||||||||
| Hypotension | ||||||||||||||||||||||
| Delayed motor development | ||||||||||||||||||||||
| Malnutrition (probe, regurgitation) | ||||||||||||||||||||||
| Funnel chest | ||||||||||||||||||||||
| Craniosynostosis | ||||||||||||||||||||||
| Gait disturbance | ||||||||||||||||||||||
| Fast fatiguability | ||||||||||||||||||||||
| Muscular dystonia | ||||||||||||||||||||||
| Pain in muscles and joints | ||||||||||||||||||||||
| Joint hypermobility | ||||||||||||||||||||||
| Valgus deformity of the knee joints | ||||||||||||||||||||||
| Impaired mental function | ||||||||||||||||||||||
| Hypomineralization of the bones | ||||||||||||||||||||||
| Skull deformities | ||||||||||||||||||||||
| short stature | ||||||||||||||||||||||
| fractures | ||||||||||||||||||||||
| Hypomineralization of bone tissue | ||||||||||||||||||||||
| Respiratory failure | ||||||||||||||||||||||
| limb shortening | ||||||||||||||||||||||
| Hypercalcemia | ||||||||||||||||||||||
| Planovalgus foot deformity | ||||||||||||||||||||||
| Osteoporosis | ||||||||||||||||||||||
| Protein-energy malnutrition | ||||||||||||||||||||||
| convulsions | ||||||||||||||||||||||
| Nephrocalcinosis/kidney damage | ||||||||||||||||||||||
| Hypercalciuria | ||||||||||||||||||||||
| Hyperphosphatemia | ||||||||||||||||||||||
| Tubulopathy | ||||||||||||||||||||||
| Hydrocephalus | ||||||||||||||||||||||
| Signs of bone defects | ||||||||||||||||||||||
| Poorly healed fractures of the tibia | ||||||||||||||||||||||
| Anomaly Arnold Chiari II, | ||||||||||||||||||||||
| Multiple organ failure | ||||||||||||||||||||||
| Frequent colds, bronchitis | ||||||||||||||||||||||
| Irritability | ||||||||||||||||||||||
| Tremor | ||||||||||||||||||||||
| Myopia | ||||||||||||||||||||||
| Complex astigmatism | ||||||||||||||||||||||
| Genotype | c.1163A>G/c.1163A>G | c.1171delC/c.1171delC | c.1171delC/c.302A>G | c.508A>G/c.508A>G | c.571G>A/c.1259G>T | c.303C>A/c.314C>T | c.253A>C/c.571G>A | c.571G>A/c.1157_1175inv | c.984_986delCT/c.371A>G | c.526G>A/c.1375G>A | c.211C>T/c.1101_1103del | c.1364G>A/c.571G>A | c.340G>A/c.571G>A | c.303C>A/c.571G>A | c.571G>A/c.144_148dup | c.571G>A/c.1309+2T>C | c.571G>A/c.1309+2T>C | c.382G>A/c.871G>C | c.571G>A/c.662delG | c.571G>A/c.662delG | c.302A>G/c.571G>A | c.1171delC/c.571G>A |
 indicates homozygotes,
indicates homozygotes,  —compound heterozygotes.
—compound heterozygotes.Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glotov, O.S.; Savostyanov, K.V.; Nagornova, T.S.; Chernov, A.N.; Fedyakov, M.A.; Raspopova, A.N.; Krasnoukhov, K.N.; Danilov, L.G.; Moiseeva, N.V.; Kalinin, R.S.; et al. Clinical and Genetic Characteristics of Pediatric Patients with Hypophosphatasia in the Russian Population. Int. J. Mol. Sci. 2022, 23, 12976. https://doi.org/10.3390/ijms232112976
Glotov OS, Savostyanov KV, Nagornova TS, Chernov AN, Fedyakov MA, Raspopova AN, Krasnoukhov KN, Danilov LG, Moiseeva NV, Kalinin RS, et al. Clinical and Genetic Characteristics of Pediatric Patients with Hypophosphatasia in the Russian Population. International Journal of Molecular Sciences. 2022; 23(21):12976. https://doi.org/10.3390/ijms232112976
Chicago/Turabian StyleGlotov, Oleg S., Kirill V. Savostyanov, Tatyana S. Nagornova, Alexandr N. Chernov, Mikhail A. Fedyakov, Aleksandra N. Raspopova, Konstantin N. Krasnoukhov, Lavrentii G. Danilov, Nadegda V. Moiseeva, Roman S. Kalinin, and et al. 2022. "Clinical and Genetic Characteristics of Pediatric Patients with Hypophosphatasia in the Russian Population" International Journal of Molecular Sciences 23, no. 21: 12976. https://doi.org/10.3390/ijms232112976