
Embryonic Exposure to Cigarette Smoke Extract Impedes Skeletal Development and Evokes Craniofacial Defects in Zebrafish


Abstract
:
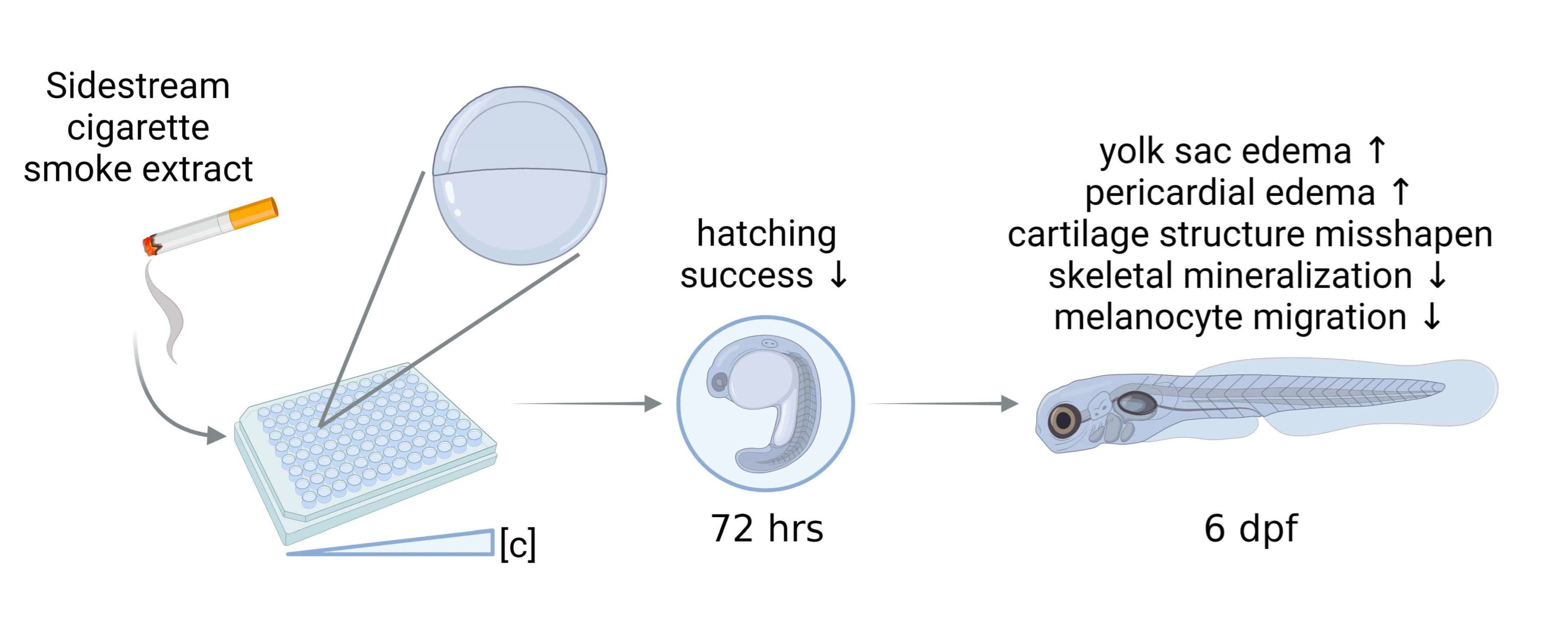{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Sidestream Tobacco Smoke Solutions Alter Zebrafish Embryo Survival and Hatching In Vivo
2.2. All Tobacco Products, Including Harm-Reduction Products, Significantly Increased General Developmental Defects In Vivo
2.3. Both Conventional and Harm-Reduction Tobacco Products Affect Cartilage Formation
2.4. Different Tobacco Products Affect Distinct Bone Formation during Zebrafish Development
2.5. Melanocyte Migration
3. Discussion
4. Materials and Methods
4.1. Production of Smoke Solutions
4.2. Zebrafish Rearing and Collection
4.3. Hatching Success and Survival
4.4. Skeletal Staining
4.5. Image Morphometry
4.5.1. Cartilage Lengths
4.5.2. Scoring of Melanocyte Travel Distance and Area
4.5.3. Mineralization Abundance
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Committee on the Public Health Implications of Raising the Minimum Age for Purchasing Tobacco Products; Board on Population Health and Public Health Practice; Institute of Medicine. The Effects of Tobacco Use on Health. In Public Health Implications of Raising the Minimum Age of Legal Access to Tobacco Products; Bonnie, R.J., Stratton, K., Kwan, L.Y., Eds.; National Academies Press: Washington, DC, USA, 2015. Available online: https://www.ncbi.nlm.nih.gov/books/NBK310413/ (accessed on 21 May 2022).
- Jha, P.; Ramasundarahettige, C.; Landsman, V.; Rostron, B.; Thun, M.; Anderson, R.N.; McAfee, T.; Peto, R. 21st-Century Hazards of Smoking and Benefits of Cessation in the United States. N. Engl. J. Med. 2013, 368, 341–350. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute. Harms of Cigarette Smoking and Health Benefits of Quitting. 2017. Available online: https://www.cancer.gov/about-cancer/causes-prevention/risk/tobacco/cessation-fact-sheet (accessed on 2 June 2022).
- Daniell, H.W. Osteoporosis of the slender smoker. Vertebral compression fractures and loss of metacarpal cortex in relation to postmenopausal cigarette smoking and lack of obesity. Arch. Intern. Med. 1976, 136, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Seeman, E.; Melton, L.J., 3rd; O’Fallon, W.; Riggs, B. Risk factors for spinal osteoporosis in men. Am. J. Med. 1983, 75, 977–983. [Google Scholar] [CrossRef]
- Wickham, C.A.; Walsh, K.; Cooper, C.; Barker, D.J.; Margetts, B.M.; Morris, J.; Bruce, S.A. Dietary calcium, physical activity, and risk of hip fracture: A prospective study. BMJ 1989, 299, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Bergström, J.; Eliasson, S.; Preber, H. Cigarette Smoking and Periodontal Bone Loss. J. Periodontol. 1991, 62, 242–246. [Google Scholar] [CrossRef]
- Glynn, W.N.; Meilahn, E.N.; Charron, M.; Anderson, S.J.; Kuller, L.H.; Cauley, J.A. Determinants of bone mineral density in older men. J. Bone Miner. Res. 1995, 10, 1769–1777. [Google Scholar] [CrossRef]
- Egger, P.; Duggleby, S.; Hobbs, R.; Fall, C.; Cooper, C. Cigarette smoking and bone mineral density in the elderly. J. Epidemiology Community Health 1996, 50, 47–50. [Google Scholar] [CrossRef]
- Law, M.R.; Hackshaw, A.K. A meta-analysis of cigarette smoking, bone mineral density and risk of hip fracture: Recognition of a major effect. BMJ 1997, 315, 841–846. [Google Scholar] [CrossRef]
- Ward, K.D.; Klesges, R.C. A meta-analysis of the effects of cigarette smoking on bone mineral density. Calcif. Tissue Res. 2001, 68, 259–270. [Google Scholar] [CrossRef]
- Al-Bashaireh, A.M.; Haddad, L.G.; Weaver, M.; Chengguo, X.; Kelly, D.L.; Yoon, S. The Effect of Tobacco Smoking on Bone Mass: An Overview of Pathophysiologic Mechanisms. J. Osteoporos. 2018, 2018, 1206235. [Google Scholar] [CrossRef] [Green Version]
- Lorentzon, M.; Mellström, D.; Haug, E.; Ohlsson, C. Smoking Is Associated with Lower Bone Mineral Density and Reduced Cortical Thickness in Young Men. J. Clin. Endocrinol. Metab. 2007, 92, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Krall, E.A.; Dawson-Hughes, B. Smoking and bone loss among postmenopausal women. J. Bone Miner. Res. 1991, 6, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Naeem, Z. Second-Hand Smoke: Ignored Implications. Int. J. Health Sci. 2015, 9, 24103. [Google Scholar] [CrossRef] [PubMed]
- Acuff, L.; Fristoe, K.; Hamblen, J.; Smith, M.; Chen, J. Third-Hand Smoke: Old Smoke, New Concerns. J. Community Health 2016, 41, 680–687. [Google Scholar] [CrossRef]
- U.S. Department of Health and Human Services. The Health Consequences of Smoking—50 Years of Progress: A Report of the Surgeon General; U.S. Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Chronic Disease Prevention and Health Promotion, Office on Smoking and Health: Atlanta, GA, USA, 2014.
- Walton, K.; Gentzke, A.S.; Murphy-Hoefer, R.; Kenemer, B.; Neff, L.J. Exposure to Secondhand Smoke in Homes and Vehicles Among US Youths, United States, 2011–2019. Prev. Chronic Dis. 2020, 17, 200107. [Google Scholar] [CrossRef]
- Martinez, I.K.C.; Sparks, N.R.L.; Madrid, J.V.; Talbot, P.; zur Nieden, N.I. Exposure to cigarette smoke impedes human in vitro osteoblast differentiation independently of nicotine. Nicotine Tob. Res. 2022, ntac144. [Google Scholar] [CrossRef]
- Institute of Medicine (US). Committee to Assess the Science Base for Tobacco Harm Reduction. In Clearing the Smoke: Assessing the Science Base for Tobacco Harm Reduction; Stratton, K., Shetty, P., Wallace, R., Bondurant, S., Eds.; National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Mackay, E.W.; Apschner, A.; Schulte-Merker, S. A bone to pick with zebrafish. BoneKEy Rep. 2013, 2, 445. [Google Scholar] [CrossRef]
- Hall, B.K. (Ed.) Bones and Cartilage; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar] [CrossRef]
- Keller, K.C.; zur Nieden, N.I. Osteogenesis from pluripotent stem cells: Neural crest or mesodermal origin? In Embryonic Stem Cells; Kallos, M.S., Ed.; IntechOpen: Rijeka, Croatia, 2011; pp. 323–348. [Google Scholar]
- Long, F.; Ornitz, D.M. Development of the Endochondral Skeleton. Cold Spring Harb. Perspect. Biol. 2013, 5, a008334. [Google Scholar] [CrossRef]
- Walker, L.; Preston, M.; Magnay, J.; Thomas, P.; El Haj, A. Nicotinic regulation of c-fos and osteopontin expression in human-derived osteoblast-like cells and human trabecular bone organ culture. Bone 2001, 28, 603–608. [Google Scholar] [CrossRef]
- Ma, L.; Zheng, L.W.; Sham, M.H.; Cheung, L.K. Uncoupled angiogenesis and osteogenesis in nicotine-compromised bone healing. J. Bone Miner. Res. 2010, 25, 1305–1313. [Google Scholar] [CrossRef]
- Lammer, E.J.; Shaw, G.M.; Iovannisci, D.M.; Finnell, R. Maternal Smoking, Genetic Variation of Glutathione S-Transferases, and Risk for Orofacial Clefts. Epidemiology 2005, 16, 698–701. [Google Scholar] [CrossRef] [PubMed]
- Sanbe, A.; Mizutani, R.; Miyauchi, N.; Yamauchi, J.; Nagase, T.; Yamamura, K.-I.; Tanoue, A. Inhibitory effects of cigarette smoke extract on neural crest migration occur through suppression of R-spondin1 expression via aryl hydrocarbon receptor. Naunyn-Schmiedebergs Arch. Fur Exp. Pathol. Und Pharmakol. 2009, 380, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.E.; Kandalam, S.; Olivares-Navarrete, R.; Dickinson, A.J.G. E-cigarette aerosol exposure can cause craniofacial defects in Xenopus laevis embryos and mammalian neural crest cells. PLoS ONE 2017, 12, e0185729. [Google Scholar] [CrossRef] [PubMed]
- Kuo, B.R.; Erickson, C.A. Regional differences in neural crest morphogenesis. Cell Adhes. Migr. 2010, 4, 567–585. [Google Scholar] [CrossRef] [PubMed]
- Dupin, E.; Sommer, L. Neural crest progenitors and stem cells: From early development to adulthood. Dev. Biol. 2012, 366, 83–95. [Google Scholar] [CrossRef]
- Pavan, W.J.; Raible, D.W. Specification of neural crest into sensory neuron and melanocyte lineages. Dev. Biol. 2012, 366, 55–63. [Google Scholar] [CrossRef]
- Mort, R.L.; Jackson, I.J.; Patton, E.E. The melanocyte lineage in development and disease. Development 2015, 142, 620–632. [Google Scholar] [CrossRef]
- Buck, K.; Nieden, N.I.Z. Risk Assessment Using Human Pluripotent Stem Cells: Recent Advances in Developmental Toxicity Screens. In Stem Cells in Birth Defects Research and Developmental Toxicology; Rasmussen, T.P., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2018. [Google Scholar]
- Sant, K.E.; Timme-Laragy, A.R. Zebrafish as a Model for Toxicological Perturbation of Yolk and Nutrition in the Early Embryo. Curr. Environ. Health Rep. 2018, 5, 125–133. [Google Scholar] [CrossRef]
- Pinheiro-Da-Silva, J.; Luchiari, A.C. Embryonic ethanol exposure on zebrafish early development. Brain Behav. 2021, 11, e02062. [Google Scholar] [CrossRef]
- Kiziltan, T.; Baran, A.; Kankaynar, M.; Şenol, O.; Sulukan, E.; Yildirim, S.; Ceyhun, S.B. Effects of the food colorant carmoisine on zebrafish embryos at a wide range of concentrations. Arch. Toxicol. 2022, 96, 1089–1099. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhong, Z.; Wang, M.; Zhang, X. 5-Hydroxymethyl-2-furaldehyde induces developmental toxicology and decreases bone mineralization in zebrafish larvae. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 254, 109254. [Google Scholar] [CrossRef] [PubMed]
- Knecht, A.L.; Goodale, B.C.; Truong, L.; Simonich, M.T.; Swanson, A.J.; Matzke, M.M.; Anderson, K.A.; Waters, K.M.; Tanguay, R.L. Comparative developmental toxicity of environmentally relevant oxygenated PAHs. Toxicol. Appl. Pharmacol. 2013, 271, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Chlebowski, A.C.; Garcia, G.R.; La Du, J.K.; Bisson, W.H.; Truong, L.; Simonich, S.L.M.; Tanguay, R.L. Mechanistic Investigations Into the Developmental Toxicity of Nitrated and Heterocyclic PAHs. Toxicol. Sci. 2017, 157, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Van Duuren, B.L. Identification of Some Polynuclear Aromatic Hydrocarbons in Cigarette-Smoke Condensate2,3. JNCI J. Natl. Cancer Inst. 1958, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lyons, M.J. Presence of 1,2,3,4-Dibenzopyrene in Cigarette Smoke. Nature 1958, 182, 178. [Google Scholar] [CrossRef]
- Nogales, F.F.; Beltran, E.; Fernandez, P.L. The pathology of secondary human yolk sac in spontaneous abortion: Findings in 103 cases. In Progress in Surgical Pathology; Fenoglio-Preiser, C.M., Wolff, M., Rilke, F., Eds.; Volume XII; Springer: Berlin/Heidelberg, Germany, 1992; pp. 291–303. [Google Scholar]
- Riveles, K.; Tran, V.; Roza, R.; Kwan, D.; Talbot, P. Smoke from traditional commercial, harm reduction and research brand cigarettes impairs oviductal functioning in hamsters (Mesocricetus auratus) in vitro. Hum. Reprod. 2007, 22, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Tran, V.; Talbot, P. Comparison of toxicity of smoke from traditional and harm-reduction cigarettes using mouse embryonic stem cells as a novel model for preimplantation development. Hum. Reprod. 2009, 24, 386–397. [Google Scholar] [CrossRef]
- Valenca, S.S.; Castro, P.; Pimenta, W.A.; Lanzetti, M.; Silva, S.V.; Barja-Fidalgo, C.; Koatz, V.L.G.; Porto, L.C. Light cigarette smoke-induced emphysema and NFκB activation in mouse lung. Int. J. Exp. Pathol. 2006, 87, 373–381. [Google Scholar] [CrossRef]
- Denlinger-Apte, R.L.; Joel, D.L.; Strasser, A.A.; Donny, E.C. Low Nicotine Content Descriptors Reduce Perceived Health Risks and Positive Cigarette Ratings in Participants Using Very Low Nicotine Content Cigarettes. Nicotine Tob. Res. 2017, 19, 1149–1154. [Google Scholar] [CrossRef]
- Kozlowski, L.T.; Sweeney, C.T. Low Yield, Light, and Ultra Light Cigarettes: Let’s Understand the Product Before We Promote. In Social Marketing; Goldberg, M.E., Fishbein, M., Middlestadt, S.E., Eds.; Psychology Press: New York, NY, USA, 2018; pp. 231–244. [Google Scholar] [CrossRef]
- Epperson, A.E.; Lambin, E.F.; Henriksen, L.; Baiocchi, M.; Flora, J.A.; Prochaska, J.J. Natural American Spirit’s pro-environment packaging and perceptions of reduced-harm cigarettes. Prev. Med. 2019, 126, 105782. [Google Scholar] [CrossRef]
- National Cancer Institute. ‘Light’ Cigarettes and Cancer Risk. Available online: https://www.cancer.gov/about-cancer/causes-prevention/risk/tobacco/light-cigarettes-fact-sheet (accessed on 2 June 2022).
- Ashraf, M.W. Levels of Heavy Metals in Popular Cigarette Brands and Exposure to These Metals via Smoking. Sci. World, J. 2012, 2012, 729430. [Google Scholar] [CrossRef] [PubMed]
- Pappas, R.S.; Fresquez, M.R.; Martone, N.; Watson, C.H. Toxic Metal Concentrations in Mainstream Smoke from Cigarettes Available in the USA. J. Anal. Toxicol. 2014, 38, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, T.; Yamada, H.; Takeuchi, M.; Kozuka, H.; Kato, T.; Sudo, H. Inhibitory effects of cadmium on in vitro calcification of a clonal osteogenic cell, MC3T3-E1. Toxicol. Appl. Pharmacol. 1988, 96, 52–59. [Google Scholar] [CrossRef]
- Kaji, T.; Takata, M.; Miyahara, T.; Kozuka, H.; Koizumi, F. Interaction between cadmium and copper on ossification of embryonic chick bone in tissue culture. Toxicol. Lett. 1991, 55, 255–262. [Google Scholar] [CrossRef]
- Puzas, J.E.; Sickel, M.J.; Felter, M.E. Osteoblasts and chondrocytes are important target cells for the toxic effects of lead. Neurotoxicology 1992, 13, 783–788. [Google Scholar]
- Al-Ghafari, A.; Elmorsy, E.; Fikry, E.; Alrowaili, M.; Carter, W.G. The heavy metals lead and cadmium are cytotoxic to human bone osteoblasts via induction of redox stress. PLoS ONE 2019, 14, e0225341. [Google Scholar] [CrossRef]
- Ciosek, Z.; Kot, K.; Kosik-Bogacka, D.; Łanocha-Arendarczyk, N.; Rotter, I. The Effects of Calcium, Magnesium, Phosphorus, Fluoride, and Lead on Bone Tissue. Biomolecules 2021, 11, 506. [Google Scholar] [CrossRef]
- Xu, C.; Xu, J.; Zhang, X.; Xu, S.; Liu, Q.; Weng, Z.; Gu, A. Serum nickel is associated with craniosynostosis risk: Evidence from humans and mice. Environ. Int. 2020, 146, 106289. [Google Scholar] [CrossRef]
- Patrone, C.; Pollio, G.; Vegeto, E.; Enmark, E.; de Curtis, I.; Gustafsson, J.; Maggi, A. Estradiol Induces Differential Neuronal Phenotypes by Activating Estrogen Receptor α or β. Endocrinology 2000, 141, 1839–1845. [Google Scholar] [CrossRef]
- Kim, H.; Kim, Y.Y.; Ku, S.-Y.; Kim, S.H.; Choi, Y.M.; Moon, S.Y. The Effect of Estrogen Compounds on Human Embryoid Bodies. Reprod. Sci. 2013, 20, 661–669. [Google Scholar] [CrossRef]
- Varshney, M.K.; Inzunza, J.; Lupu, D.; Ganapathy, V.; Antonson, P.; Rüegg, J.; Nalvarte, I.; Gustafsson, J.Å. Role of estrogen receptor beta in neural differentiation of mouse embryonic stem cells. Proc. Natl. Acad. Sci. USA 2017, 114, E10428–E10437. [Google Scholar] [CrossRef]
- Pölkki, M.; Rantala, M.J. Smoking affects womens’ sex hormone–Regulated body form. Am. J. Public Health 2009, 99, 1350. [Google Scholar] [CrossRef] [PubMed]
- Varea, O.; Garrido, J.J.; Dopazo, A.; Mendez, P.; Garcia-Segura, L.M.; Wandosell, F. Estradiol Activates β-Catenin Dependent Transcription in Neurons. PLoS ONE 2009, 4, e5153. [Google Scholar] [CrossRef]
- Xiong, W.; Zhang, L.; Yu, L.; Xie, W.; Man, Y.; Xiong, Y.; Liu, H.; Liu, Y. Estradiol promotes cells invasion by activating β-catenin signaling pathway in endometriosis. Reproduction 2015, 150, 507–516. [Google Scholar] [CrossRef]
- Haegel, H.; Larue, L.; Ohsugi, M.; Fedorov, L.; Herrenknecht, K.; Kemler, R. Lack of beta-catenin affects mouse development at gastrulation. Development 1995, 121, 3529–3537. [Google Scholar] [CrossRef]
- Dorsky, R.I.; Moon, R.T.; Raible, D.W. Control of neural crest cell fate by the Wnt signalling pathway. Nature 1998, 396, 370–373. [Google Scholar] [CrossRef]
- Leung, A.W.; Murdoch, B.; Salem, A.F.; Prasad, M.S.; Gomez, G.A.; García-Castro, M.I. WNT/β-catenin signaling mediates human neural crest induction via a pre-neural border intermediate. Development 2016, 143, 398–410. [Google Scholar] [CrossRef]
- Wurzel, H.; Yeh, C.C.; Gairola, C.; Chow, C.K. Oxidative damage and antioxidant status in the lungs and bronchoalveolar lavage fluid of rats exposed chronically to cigarette smoke. J. Biochem. Toxicol. 1995, 10, 11–17. [Google Scholar]
- Chen, C. Cigarette smoke extract inhibits oxidative modification of low density lipoprotein. Atherosclerosis 1995, 112, 177–185. [Google Scholar] [CrossRef]
- Zákány, R.; Bakondi, E.; Juhász, T.; Matta, C.; Szíjgyártó, Z.; Erdélyi, K.; Szabó, E.; Módis, L.; Virág, L.; Gergely, P. Oxidative stress-induced poly(ADP-ribosyl)ation in chick limb bud-derived chondrocytes. Int. J. Mol. Med. 2007, 19, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Fitriasari, S.; Trainor, P.A. Diabetes, Oxidative Stress, and DNA Damage Modulate Cranial Neural Crest Cell Development and the Phenotype Variability of Craniofacial Disorders. Front. Cell Dev. Biol. 2021, 9, 1226. [Google Scholar] [CrossRef] [PubMed]
- Knoll, M.; Shaoulian, R.; Magers, T.; Talbot, P. Ciliary Beat Frequency of Hamster Oviducts is Decreased in Vitro by Exposure to Solutions of Mainstream and Sidestream Cigarette Smoke1. Biol. Reprod. 1995, 53, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Knoll, M.; Talbot, P. Cigarette smoke inhibits oocyte cumulus complex pick-up by the oviduct in vitro independent of ciliary beat frequency. Reprod. Toxicol. 1998, 12, 57–68. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Vliet, S.M.; Dasgupta, S.; Sparks, N.R.; Kirkwood, J.S.; Vollaro, A.; Hur, M.; Nieden, N.I.Z.; Volz, D.C. Maternal-to-zygotic transition as a potential target for niclosamide during early embryogenesis. Toxicol. Appl. Pharmacol. 2019, 380, 114699. [Google Scholar] [CrossRef]
- Aceto, J.; Nourizadeh-Lillabadi, R.; Marée, R.; Dardenne, N.; Jeanray, N.; Wehenkel, L.; Aleström, P.; van Loon, J.J.W.A.; Muller, M. Zebrafish Bone and General Physiology Are Differently Affected by Hormones or Changes in Gravity. PLoS ONE 2015, 10, e0126928. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karmach, O.; Madrid, J.V.; Dasgupta, S.; Volz, D.C.; zur Nieden, N.I. Embryonic Exposure to Cigarette Smoke Extract Impedes Skeletal Development and Evokes Craniofacial Defects in Zebrafish. Int. J. Mol. Sci. 2022, 23, 9904. https://doi.org/10.3390/ijms23179904
Karmach O, Madrid JV, Dasgupta S, Volz DC, zur Nieden NI. Embryonic Exposure to Cigarette Smoke Extract Impedes Skeletal Development and Evokes Craniofacial Defects in Zebrafish. International Journal of Molecular Sciences. 2022; 23(17):9904. https://doi.org/10.3390/ijms23179904
Chicago/Turabian StyleKarmach, Omran, Joseph V. Madrid, Subham Dasgupta, David C. Volz, and Nicole I. zur Nieden. 2022. "Embryonic Exposure to Cigarette Smoke Extract Impedes Skeletal Development and Evokes Craniofacial Defects in Zebrafish" International Journal of Molecular Sciences 23, no. 17: 9904. https://doi.org/10.3390/ijms23179904