
A Review on the Molecular Mechanisms of Action of Natural Products in Preventing Bone Diseases

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
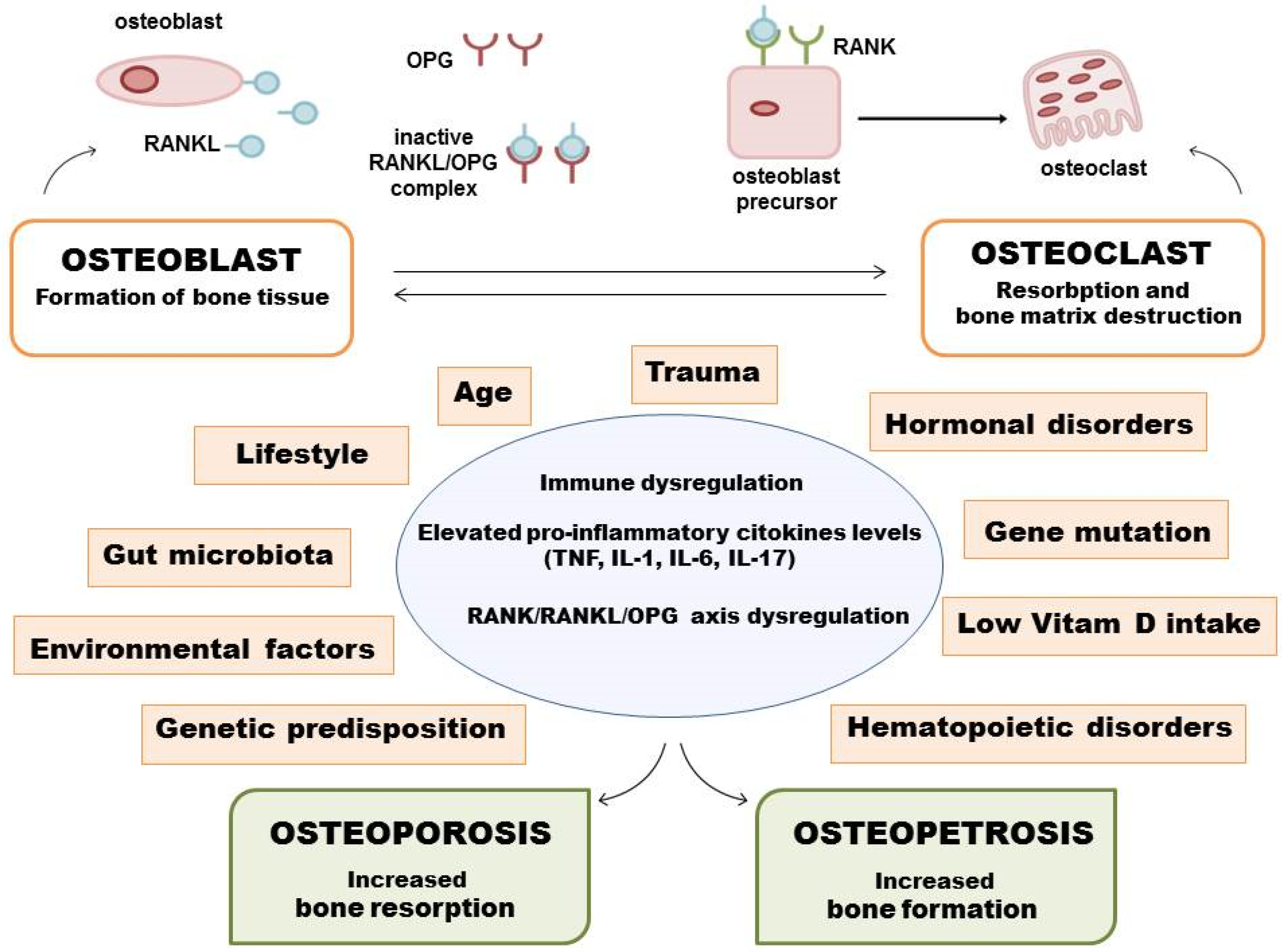
2. Pathological Mechanism of Development of Some Bone Diseases
3. Cellular and Molecular Mechanisms of Bone Health-Promoting Properties of Some Natural Products
3.1. Targeting the RANKL/M-CSF/OPG and MAPKs/JNK/NF-κB Signaling Pathways
3.2. Targeting Keap-1/Nrf2/HO-1 Signaling Pathway
3.3. Targeting BMP2-Wnt/β-Catenin Signaling Pathway
3.4. Targeting PI3K/Akt/GSK3β Signaling Pathway
3.5. Targeting Rev-Erbs Signaling Pathway
3.6. Targeting Calcium Ion Signaling Pathway
3.7. Targeting Endogenous Molecules with Bone Promoting Properties
3.8. Targeting Vitamin D Receptor (VDR)
4. Clinical Trials of Natural Products for Bone Diseases
5. Safety Concerns of Applying Some Natural Products for Clinical Management of Bone Diseases and Limitations of Some of the Current Studies
6. Conclusions and Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barbosa, J.S.; Almeida Paz, F.A.; Braga, S.S. Bisphosphonates, Old Friends of Bones and New Trends in Clinics. J. Med. Chem. 2021, 64, 1260–1282. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, L.; Goyal, R. Molecular Signaling Pathways and Essential Metabolic Elements in Bone Remodeling: An Implication of Therapeutic Targets for Bone Diseases. Curr. Drug Targets 2021, 22, 77–104. [Google Scholar] [CrossRef]
- Papapoulos, S.E. Bisphosphonate Actions: Physical Chemistry Revisited. Bone 2006, 38, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Baroncelli, G.I.; Bertelloni, S. The Use of Bisphosphonates in Pediatrics. Horm. Res. Paediatr. 2014, 82, 290–302. [Google Scholar] [CrossRef]
- Lim, A.; Simm, P.J.; James, S.; Lee, S.L.K.; Zacharin, M. Outcomes of Zoledronic Acid Use in Paediatric Conditions. Horm. Res. Paediatr. 2020, 93, 442–452. [Google Scholar] [CrossRef]
- Migliorati, C.A. Oral Complications in Cancer Patients–Medication-Related Osteonecrosis of the Jaw (MRONJ). Front. Oral Health 2022, 3, 866871. [Google Scholar] [CrossRef]
- Ruggiero, S.L.; Dodson, T.B.; Aghaloo, T.; Carlson, E.R.; Ward, B.B.; Kademani, D. American Association of Oral and Maxillofacial Surgeons’ Position Paper on Medication-Related Osteonecrosis of the Jaws—2022 Update. J. Oral Maxillofac. Surg. 2022, 80, 920–943. [Google Scholar] [CrossRef]
- Wick, A.; Bankosegger, P.; Otto, S.; Hohlweg-Majert, B.; Steiner, T.; Probst, F.; Ristow, O.; Pautke, C. Risk Factors Associated with Onset of Medication-Related Osteonecrosis of the Jaw in Patients Treated with Denosumab. Clin. Oral Investig. 2022, 26, 2839–2852. [Google Scholar] [CrossRef]
- Suresh, E.; Pazianas, M.; Abrahamsen, B. Safety Issues with Bisphosphonate Therapy for Osteoporosis. Rheumatology 2014, 53, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Van Weelden, W.J.; Massuger, L.F.A.G.; Pijnenborg, J.M.A.; Romano, A. Anti-Estrogen Treatment in Endometrial Cancer: A Systematic Review. Front. Oncol. 2019, 9, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, I.R.; Hosking, D.J. Bisphosphonates in Paget’s Disease. Bone 2011, 49, 89–94. [Google Scholar] [CrossRef]
- Reid, I.R. Management of Paget’s Disease of Bone. Osteoporos. Int. 2020, 31, 827–837. [Google Scholar] [CrossRef]
- George, A.J. The Actions and Side Effects of Anabolic Steroids in Sport and Social Abuse. Andrologie 2003, 13, 354–380. [Google Scholar] [CrossRef] [Green Version]
- Albano, G.D.; Amico, F.; Cocimano, G.; Liberto, A.; Maglietta, F.; Esposito, M.; Rosi, G.L.; Di Nunno, N.; Salerno, M.; Montana, A. Review Adverse Effects of Anabolic-Androgenic Steroids: A Literature Review. Healthcare 2021, 9, 97. [Google Scholar] [CrossRef]
- Pouresmaeili, F.; Kamalidehghan, B.; Kamarehei, M.; Goh, Y.M. A Comprehensive Overview on Osteoporosis and Its Risk Factors. Ther. Clin. Risk Manag. 2018, 14, 2029–2049. [Google Scholar] [CrossRef] [Green Version]
- Streicher, C.; Heyny, A.; Andrukhova, O.; Haigl, B.; Slavic, S.; Schüler, C.; Kollmann, K.; Kantner, I.; Sexl, V.; Kleiter, M.; et al. Estrogen Regulates Bone Turnover by Targeting RANKL Expression in Bone Lining Cells. Sci. Rep. 2017, 7, 6460. [Google Scholar] [CrossRef] [Green Version]
- Rachner, T.; Khosla, S.; Hofbauer, L. Osteoporosis: Now and the Future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef] [Green Version]
- Hofbauer, L.C.; Khosla, S.; Dunstan, C.R.; Lacey, D.L.; Spelsberg, T.C.; Riggs, B.L. Estrogen Stimulates Gene Expression and Protein Production of Osteoprotegerin in Human Osteoblastic Cells. Endocrinology 1999, 140, 4367–4370. [Google Scholar] [CrossRef]
- Demontiero, O.; Vidal, C.; Duque, G. Aging and Bone Loss: New Insights for the Clinician. Ther. Adv. Musculoskelet. Dis. 2012, 4, 61. [Google Scholar] [CrossRef] [Green Version]
- Putnam, S.E.; Scutt, A.M.; Bicknell, K.; Priestley, C.M.; Williamson, E.M. Natural Products as Alternative Treatments ForMetabolic Bone Disorders and for Maintenanceof Bone Health. Phyther. Res. 2007, 21, 99–112. [Google Scholar] [CrossRef]
- Buhrmann, C.; Brockmueller, A.; Mueller, A.L.; Shayan, P.; Shakibaei, M. Curcumin Attenuates Environment-Derived Osteoarthritis by Sox9/NF-KB Signaling Axis. Int. J. Mol. Sci. 2021, 22, 7645. [Google Scholar] [CrossRef]
- Park, Y.; Moon, H.J.; Paik, D.J.; Kim, D.Y. Effect of Dietary Legumes on Bone-Specific Gene Expression in Ovariectomized Rats. Nutr. Res. Pract. 2013, 7, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Shevkani, K.; Chourasia, S. Dietary Proteins: Functions, Health Benefits and Healthy Aging. In Nutrition, Food and Diet in Ageing and Longevity. Healthy Ageing and Longevity; Rattan, S.I., Kaur, G., Eds.; Springer: Cham, Switzerland, 2021; pp. 3–37. [Google Scholar]
- Shams-White, M.M.; Chung, M.; Fu, Z.; Insogna, K.L.; Karlsen, M.C.; LeBoff, M.S.; Shapses, S.A.; Sackey, J.; Shi, J.; Wallace, T.C.; et al. Animal versus Plant Protein and Adult Bone Health: A Systematic Review and Meta-Analysis from the National Osteoporosis Foundation. PLoS ONE 2018, 13, 1528–1543. [Google Scholar] [CrossRef] [Green Version]
- Groenendijk, I.; den Boeft, L.; van Loon, L.J.C.; de Groot, L.C.P.G.M. High Versus Low Dietary Protein Intake and Bone Health in Older Adults: A Systematic Review and Meta-Analysis. Comput. Struct. Biotechnol. J. 2019, 17, 1101–1112. [Google Scholar] [CrossRef]
- He, J.; Li, X.; Wang, Z.; Bennett, S.; Chen, K.; Xiao, Z.; Zhan, J.; Chen, S.; Hou, Y.; Chen, J.; et al. Therapeutic Anabolic and Anticatabolic Benefits of Natural Chinese Medicines for the Treatment of Osteoporosis. Front. Pharmacol. 2019, 10, 1344. [Google Scholar] [CrossRef]
- Guo, Y.; Li, Y.; Xue, L.; Severino, R.P.; Gao, S.; Niu, J.; Qin, L.P.; Zhang, D.; Brömme, D. Salvia Miltiorrhiza: An Ancient Chinese Herbal Medicine as a Source for Anti-Osteoporotic Drugs. J. Ethnopharmacol. 2014, 155, 1401–1416. [Google Scholar] [CrossRef] [PubMed]
- Rufus, P.; Mohamed, N.; Shuid, A. Beneficial Effects of Traditional Chinese Medicine on the Treatment of Osteoporosis on Ovariectomised Rat Models. Curr. Drug Targets 2013, 14, 1689–1693. [Google Scholar] [CrossRef]
- Saito, F.; Hirayasu, K.; Satoh, T.; Wang, C.W.; Lusingu, J.; Arimori, T.; Shida, K.; Palacpac, N.M.Q.; Itagaki, S.; Iwanaga, S.; et al. Immune Evasion of Plasmodium Falciparum by RIFIN via Inhibitory Receptors. Nature 2017, 552, 101–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, J.M.; Lee, A.Y.; Nam, J.Y.; Lim, K.S.; Choe, M.S.; Lee, M.Y.; Kim, C.; Kim, J.S. Effects of Dipsacus Asperoides Extract on Monosodium Iodoacetate–Induced Osteoarthritis in Rats Based on Gene Expression Profiling. Front. Pharmacol. 2021, 12, 615157. [Google Scholar] [CrossRef]
- Saar-Reismaa, P.; Bragina, O.; Kuhtinskaja, M.; Reile, I.; Laanet, P.R.; Kulp, M.; Vaher, M. Extraction and Fractionation of Bioactives from Dipsacus Fullonum L. Leaves and Evaluation of Their Anti-Borrelia Activity. Pharmaceuticals 2022, 15, 87. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Lee, A.; Kim, T.; Jang, S.A.; Ha, H. Anti-Osteoporotic Effects of Commiphora Myrrha and Its Poly-Saccharide via Osteoclastogenesis Inhibition. Plants 2021, 10, 945. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Jang, S.A.; Lee, A.; Cho, C.W.; Song, Y.R.; Hong, H.-D.; Ha, H.; Kim, T. Polysaccharides Isolated from Lotus Leaves (LLEP) Exert Anti-Osteoporotic Effects by Inhibiting Osteoclastogenesis. Int. J. Biol. Macromol. 2020, 161, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.M.; Lee, A.Y.; Moon, B.C.; Choi, G.; Kim, J.S. Effects of Dipsacus Asperoides and Phlomis Umbrosa Extracts in a Rat Model of Osteoarthritis. Plants 2021, 10, 2030. [Google Scholar] [CrossRef] [PubMed]
- Phromnoi, K.; Yodkeeree, S.; Pintha, K.; Mapoung, S.; Suttajit, M.; Saenjum, C.; Dejkriengkraikul, P. Anti-Osteoporosis Effect of Perilla Frutescens Leaf Hexane Fraction through Regulating Osteoclast and Osteoblast Differentiation. Molecules 2022, 27, 824. [Google Scholar] [CrossRef] [PubMed]
- Seo, I.; Lee, K.P.; Park, S.; Ahn, S. Therapeutic Effect of Shinkiwhan, Herbal Medicine, Regulates OPG/RANKL/RANK System on Ovariectomy-Induced Bone Loss Rat. Phys. Act. Nutr. 2020, 24, 19. [Google Scholar] [CrossRef]
- Dirckx, N.; Moorer, M.C.; Clemens, T.L.; Riddle, R.C. The Role of Osteoblasts in Energy Homeostasis. Nat. Rev. Endocrinol. 2019, 15, 651–665. [Google Scholar] [CrossRef]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.-H.; Inada, M. Targeted Disruption of Cbfa1 Results in a Complete Lack of Bone Formation Owing to Maturational Arrest of Osteoblasts. Cell 1997, 89, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Katagiri, T.; Takahashi, N. Regulatory Mechanisms of Osteoblast and Osteoclast Differentiation. Oral Dis. 2002, 8, 147–159. [Google Scholar] [CrossRef]
- Fröbel, J.; Landspersky, T.; Percin, G.; Schreck, C.; Rahmig, S.; Ori, A.; Nowak, D.; Essers, M.; Waskow, C.; Oostendorp, R.A.J. The Hematopoietic Bone Marrow Niche Ecosystem. Front. Cell Dev. Biol. 2021, 9, 1958. [Google Scholar] [CrossRef]
- Xu, F.; Teitelbaum, S.L. Osteoclasts: New Insights. Bone Res. 2013, 1, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Föger-Samwald, U.; Dovjak, P.; Azizi-Semrad, U.; Kerschan-Schindl, K.; Pietschmann, P. Osteoporosis: Pathophysiology and Therapeutic Options. EXCLI J. 2020, 19, 1017–1037. [Google Scholar] [CrossRef] [PubMed]
- Llorente, I.; García-Castañeda, N.; Valero, C.; González-Álvaro, I.; Castañeda, S. Osteoporosis in Rheumatoid Arthritis: Dangerous Liaisons. Front. Med. 2020, 7, 802. [Google Scholar] [CrossRef] [PubMed]
- Rotta, D.; Fassio, A.; Rossini, M.; Giollo, A.; Viapiana, O.; Orsolini, G.; Bertoldo, E.; Gatti, D.; Adami, G. Osteoporosis in Inflammatory Arthritides: New Perspective on Pathogenesis and Treatment. Front. Med. 2020, 7, 896. [Google Scholar] [CrossRef] [PubMed]
- Ralston, S.H.; Langston, A.L.; Reid, I.R. Pathogenesis and Management of Paget’s Disease of Bone. Lancet 2008, 372, 155–163. [Google Scholar] [CrossRef]
- Ralston, S.H.; Fraser, J. Diagnosis and Management of Osteoporosis. Practitioner 2015, 259, 15–19. [Google Scholar] [CrossRef]
- Broadhead, M.L.; Clark, J.C.M.; Myers, D.E.; Dass, C.R.; Choong, P.F.M. The Molecular Pathogenesis of Osteosarcoma: A Review. Sarcoma 2011, 2011, 959248. [Google Scholar] [CrossRef]
- Berenson, J.R.; Rajdev, L.; Broder, M. Pathophysiology of Bone Metastases. Cancer Biol. Ther. 2006, 5, 1078–1081. [Google Scholar] [CrossRef] [Green Version]
- Hambli, R. Connecting Mechanics and Bone Cell Activities in the Bone Remodeling Process: An Integrated Finite Element Modeling. Front. Bioeng. Biotechnol. 2014, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, P. Physiology of Bone. Endocr. Dev. 2015, 28, 33–55. [Google Scholar] [CrossRef]
- Komatsu, N.; Okamoto, K.; Sawa, S.; Nakashima, T.; Oh-Hora, M.; Kodama, T.; Tanaka, S.; Bluestone, J.A.; Takayanagi, H. Pathogenic Conversion of Foxp3+ T Cells into TH17 Cells in Autoimmune Arthritis. Nat. Med. 2014, 20, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Amarasekara, D.S.; Yu, J.; Rho, J. Bone Loss Triggered by the Cytokine Network in Inflammatory Autoimmune Diseases. J. Immunol. Res. 2015, 2015, 832127. [Google Scholar] [CrossRef] [PubMed]
- Eghbali-Fatourechi, G.; Khosla, S.; Sanyal, A.; Boyle, W.J.; Lacey, D.L.; Riggs, B.L. Role of RANK Ligand in Mediating Increased Bone Resorption in Early Postmenopausal Women. J. Clin. Investig. 2003, 111, 1221–1230. [Google Scholar] [CrossRef]
- Walsh, M.C.; Choi, Y. Biology of the RANKL-RANK-OPG System in Immunity, Bone, and Beyond. Front. Immunol. 2014, 5, 511. [Google Scholar] [CrossRef] [Green Version]
- Behera, J.; Ison, J.; Tyagi, S.C.; Tyagi, N. The Role of Gut Microbiota in Bone Homeostasis. Bone 2020, 135, 115317. [Google Scholar] [CrossRef] [PubMed]
- Sjögren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Bäckhed, F.; Ohlsson, C. The Gut Microbiota Regulates Bone Mass in Mice. J. Bone Miner. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef] [Green Version]
- Guss, J.D.; Taylor, E.; Rouse, Z.; Roubert, S.; Higgins, C.H.; Thomas, C.J.; Baker, S.P.; Vashishth, D.; Donnelly, E.; Shea, M.K.; et al. The Microbial Metagenome and Bone Tissue Composition in Mice with Microbiome-Induced Reductions in Bone Strength. Bone 2019, 127, 146–154. [Google Scholar] [CrossRef]
- Rozental, T.D.; Shah, J.; Chacko, A.T.; Zurakowski, D. Prevalence and Predictors of Osteoporosis Risk in Orthopaedic Patients. Clin. Orthop. Relat. Res. 2010, 468, 1765–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penna, S.; Capo, V.; Palagano, E.; Sobacchi, C.; Villa, A. One Disease, Many Genes: Implications for the Treatment of Osteopetroses. Front. Endocrinol. 2019, 10, 85. [Google Scholar] [CrossRef] [Green Version]
- Stark, Z.; Savarirayan, R. Osteopetrosis. Orphanet J. Rare Dis. 2009, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Sobacchi, C.; Schulz, A.; Coxon, F.P.; Villa, A.; Helfrich, M.H. Osteopetrosis: Genetics, Treatment and New Insights into Osteoclast Function. Nat. Rev. Endocrinol. 2013, 9, 522–536. [Google Scholar] [CrossRef]
- Sobacchi, C.; Frattini, A.; Guerrini, M.M.; Abinun, M.; Pangrazio, A.; Susani, L.; Bredius, R.; Mancini, G.; Cant, A.; Bishop, N.; et al. Osteoclast-Poor Human Osteopetrosis Due to Mutations in the Gene Encoding RANKL. Nat. Genet. 2007, 39, 960–962. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.; Nelson, C.A.; Ross, F.P.; Teitelbaum, S.L.; Fremont, D.H. Crystal Structure of the TRANCE/RANKL Cytokine Reveals Determinants of Receptor-Ligand Specificity. J. Clin. Investig. 2001, 108, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Econs, M.J.; DiMeglio, L.A.; Insogna, K.L.; Levine, M.A.; Orchard, P.J.; Miller, W.P.; Petryk, A.; Rush, E.T.; Shoback, D.M.; et al. Diagnosis and Management of Osteopetrosis: Consensus Guidelines From the Osteopetrosis Working Group. J. Clin. Endocrinol. Metab. 2017, 102, 3111–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Bari, A.A.; Mamun, A. Al Current Advances in Regulation of Bone Homeostasis. FASEB BioAdvances 2020, 2, 668–679. [Google Scholar] [CrossRef]
- Emmanuelle, N.E.; Marie-Cécile, V.; Florence, T.; Jean-François, A.; Françoise, L.; Coralie, F.; Alexia, V. Critical Role of Estrogens on Bone Homeostasis in Both Male and Female: From Physiology to Medical Implications. Int. J. Mol. Sci. 2021, 22, 1568. [Google Scholar] [CrossRef]
- Drake, M.T.; Clarke, B.L.; Lewiecki, E.M. The Pathophysiology and Treatment of Osteoporosis. Clin. Ther. 2015, 37, 1837–1850. [Google Scholar] [CrossRef]
- Redlich, K.; Smolen, J.S. Inflammatory Bone Loss: Pathogenesis and Therapeutic Intervention. Nat. Rev. Drug Discov. 2012, 11, 234–250. [Google Scholar] [CrossRef]
- Hienz, S.A.; Paliwal, S.; Ivanovski, S. Mechanisms of Bone Resorption in Periodontitis. J. Immunol. Res. 2015, 2015, 615486. [Google Scholar] [CrossRef] [Green Version]
- Di Benedetto, A.; Gigante, I.; Colucci, S.; Grano, M. Periodontal Disease: Linking the Primary Inflammation to Bone Loss. Clin. Dev. Immunol. 2013, 2013, 503754. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zheng, H.; Ma, R. Natural Occurring Compounds Inhibit Osteoclastogenesis via Targeting NFATc1-Related Signaling Pathways. Curr. Drug Targets 2020, 21, 358–364. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Ima-Nirwana, S. The Osteoprotective Effects Of Kaempferol: The Evidence From In Vivo And In Vitro Studies. Drug Des. Dev. Ther. 2019, 13, 3497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J. The Osteoprotective Effects of Artemisinin Compounds and the Possible Mechanisms Associated with Intracellular Iron: A Review of in Vivo and in Vitro Studies. Environ. Toxicol. Pharmacol. 2020, 76, 103358. [Google Scholar] [CrossRef]
- Curtis, E.M.; Moon, R.J.; Dennison, E.M.; Harvey, N.C.; Cooper, C. Recent Advances in the Pathogenesis and Treatment of Osteoporosis. Clin. Med. 2015, 15, s92–s96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, M.M.; Khoo, W.H.; Ng, P.Y.; Xiao, Y.; Zamerli, J.; Thatcher, P.; Kyaw, W.; Pathmanandavel, K.; Grootveld, A.K.; Moran, I.; et al. Osteoclasts Recycle via Osteomorphs during RANKL-Stimulated Bone Resorption. Cell 2021, 184, 1330–1347.e13. [Google Scholar] [CrossRef]
- Cawley, K.M.; Bustamante-Gomez, N.C.; Guha, A.G.; MacLeod, R.S.; Xiong, J.; Gubrij, I.; Liu, Y.; Mulkey, R.; Palmieri, M.; Thostenson, J.D.; et al. Local Production of Osteoprotegerin by Osteoblasts Suppresses Bone Resorption. Cell Rep. 2020, 32, 108052. [Google Scholar] [CrossRef]
- Kim, J.-M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.-H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Park, C.; Lee, H.; Noh, J.S.; Jin, C.Y.; Kim, G.Y.; Hyun, J.W.; Leem, S.H.; Choi, Y.H. Hemistepsin A Protects Human Keratinocytes against Hydrogen Peroxide-Induced Oxidative Stress through Activation of the Nrf2/HO-1 Signaling Pathway. Arch. Biochem. Biophys. 2020, 691, 108512. [Google Scholar] [CrossRef] [PubMed]
- Epsley, S.; Tadros, S.; Farid, A.; Kargilis, D.; Mehta, S.; Rajapakse, C.S. The Effect of Inflammation on Bone. Front. Physiol. 2021, 11, 511799. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Lee, I.H.; Iimura, T.; Kong, S.W. Two Macrophages, Osteoclasts and Microglia: From Development to Pleiotropy. Bone Res. 2021, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Nakamichi, Y.; Nakamura, M.; Sato, N.; Ninomiya, T.; Muto, A.; Nakamura, V.; Ozawa, H.; Iwasaki, Y.; Kobayashi, E.; et al. New 19-nor-(20S)-1alpha,25-Dihydroxyvitamin D3 Analogs Strongly Stimulate Osteoclast Formation Both in Vivo and in Vitro. Bone 2007, 40, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Corbo, F.; Brunetti, G.; Crupi, P.; Bortolotti, S.; Storlino, G.; Piacente, L.; Carocci, A.; Catalano, A.; Milani, G.; Colaianni, G.; et al. Effects of Sweet Cherry Polyphenols on Enhanced Osteoclastogenesis Associated With Childhood Obesity. Front. Immunol. 2019, 10, 1001. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kobayashi, Y.; Nakamichi, Y.; Udagawa, N.; Takahashi, N.; Im, N.; Seo, H.; Jeon, W.B.; Yonezawa, T.; Cha, B.Y.; et al. Alisol-B, a Novel Phyto-Steroid, Suppresses the RANKL-Induced Osteoclast Formation and Prevents Bone Loss in Mice. Biochem. Pharmacol. 2010, 80, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Fu, B.; Ji, N.; Pan, S.; Zhao, X.; Zhang, Z.; Qiu, Y.; Wang, R.; Jin, M.; Wen, K.; et al. Alisol B 23-Acetate Inhibits IgE/Ag-Mediated Mast Cell Activation and Allergic Reaction. Int. J. Mol. Sci. 2018, 19, 4092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, P.T.; Park, D.H.; Kim, O.; Kwon, S.H.; Min, B.S.; Lee, J.H. Desoxyrhapontigenin Inhibits RANKL-Induced Osteoclast Formation and Prevents Inflammation-Mediated Bone Loss. Int. J. Mol. Med. 2018, 42, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wilk, S.A.; Wang, A.; Zhou, L.; Wang, R.H.; Ogawa, W.; Deng, C.; Dong, L.Q.; Liu, F. Resveratrol Inhibits MTOR Signaling by Promoting the Interaction between MTOR and DEPTOR. J. Biol. Chem. 2010, 285, 36387–36394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Cao, Z.; Zhang, W.; Tickner, J.; Qiu, H.; Wang, C.; Chen, K.; Wang, Z.; Tan, R.; Dong, S.; et al. Lumichrome Inhibits Osteoclastogenesis and Bone Resorption through Suppressing RANKL-Induced NFAT Activation and Calcium Signaling. J. Cell. Physiol. 2018, 233, 8971–8983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepan, J.; Hruskova, H.; Kverka, M. Update on Menopausal Hormone Therapy for Fracture Prevention. Curr. Osteoporos. Rep. 2019, 17, 465–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara-Castillo, N. Estrogen Signaling in Bone. Appl. Sci. 2021, 11, 4439. [Google Scholar] [CrossRef]
- Nasibeh, Y.; Khosrow, K.; Sajad, J.; Asghar, G. Ovariectomized Rat Model of Osteoporosis: A Practical Guide. EXCLI J. 2020, 19, 89–117. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, Q.; Liu, T.; Zhao, H.; Wang, R.; Li, H.; Zhang, Y.; Shan, L.; He, B.; Wang, X.; et al. Effect of Vicenin-2 on Ovariectomy-Induced Osteoporosis in Rats. Biomed. Pharmacother. 2020, 129, 110474. [Google Scholar] [CrossRef]
- Li, C.; Yang, Z.; Li, Z.; Ma, Y.; Zhang, L.; Zheng, C.; Qiu, W.; Wu, X.; Wang, X.; Li, H.; et al. Maslinic Acid Suppresses Osteoclastogenesis and Prevents Ovariectomy-Induced Bone Loss by Regulating RANKL-Mediated NF-ΚB and MAPK Signaling Pathways. J. Bone Miner. Res. 2011, 26, 644–656. [Google Scholar] [CrossRef]
- Chen, S.; Jin, G.; Huang, K.M.; Ma, J.J.; Wang, Q.; Ma, Y.; Tang, X.Z.; Zhou, Z.J.; Hu, Z.J.; Wang, J.Y.; et al. Lycorine Suppresses RANKL-Induced Osteoclastogenesis in Vitro and Prevents Ovariectomy-Induced Osteoporosis and Titanium Particle-Induced Osteolysis in Vivo. Sci. Rep. 2015, 5, 12853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Wei, H.; Wu, Y.; Yang, S.; Xiao, L.; Zhang, J.; Peng, B. Licorice Isoliquiritigenin Suppresses RANKL-Induced Osteoclastogenesis in Vitro and Prevents Inflammatory Bone Loss in Vivo. Int. J. Biochem. Cell Biol. 2012, 44, 1139–1152. [Google Scholar] [CrossRef]
- Jin, H.; Wang, Q.; Chen, K.; Xu, K.; Pan, H.; Chu, F.; Ye, Z.; Wang, Z.; Tickner, J.; Qiu, H.; et al. Astilbin Prevents Bone Loss in Ovariectomized Mice through the Inhibition of RANKL-Induced Osteoclastogenesis. J. Cell. Mol. Med. 2019, 23, 8355–8368. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Shi, H.; Yang, W. Osteoprotective Effect of Cimiracemate in Glucocorticoid-Induced Osteoporosis by Osteoprotegerin/Receptor Activator of Nuclear Factor κ B/Receptor Activator of Nuclear Factor Kappa-β Ligand Signaling. Pharmacology 2019, 103, 163–172. [Google Scholar] [CrossRef]
- Chen, D.; Wang, Q.; Li, Y.; Sun, P.; Kuek, V.; Yuan, J.; Yang, J.; Wen, L.; Wang, H.; Xu, J.; et al. Notopterol Attenuates Estrogen Deficiency-Induced Osteoporosis via Repressing RANKL Signaling and Reactive Oxygen Species. Front. Pharmacol. 2021, 12, 664836. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.A.; Song, H.S.; Kwon, J.E.; Baek, H.J.; Koo, H.J.; Sohn, E.H.; Lee, S.R.; Kang, S.C. Protocatechuic Acid Attenuates Trabecular Bone Loss in Ovariectomized Mice. Oxid. Med. Cell. Longev. 2018, 2018, 7280342. [Google Scholar] [CrossRef]
- Peng, J.; Zhao, K.; Zhu, J.; Wang, Y.; Sun, P.; Yang, Q.; Zhang, T.; Han, W.; Hu, W.; Yang, W.; et al. Sarsasapogenin Suppresses Rankl-Induced Osteoclastogenesis in Vitro and Prevents Lipopolysaccharide-Induced Bone Loss in Vivo. Drug Des. Dev. Ther. 2020, 14, 3435–3447. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Cao, Z.; Wu, B.; Chen, F.; Tickner, J.; Wang, Z.; Qiu, H.; Wang, C.; Chen, K.; Tan, R.; et al. Modulating Calcium-Mediated NFATc1 and Mitogen-Activated Protein Kinase Deactivation Underlies the Inhibitory Effects of Kavain on Osteoclastogenesis and Bone Resorption. J. Cell. Physiol. 2018, 234, 789–801. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Yuan, Y.; Wang, Z.; Song, D.; Zhao, J.; Cao, Z.; Chen, J.; Guo, Q.; Chen, L.; Tickner, J.; et al. Helvolic Acid Attenuates Osteoclast Formation and Function via Suppressing RANKL-Induced NFATc1 Activation. J. Cell. Physiol. 2019, 234, 6477–6488. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Kim, Y.K. Berberine Suppresses RANKL-Induced Osteoclast Differentiation by Inhibiting c-Fos and NFATc1 Expression. Am. J. Chin. Med. 2019, 47, 439–455. [Google Scholar] [CrossRef]
- Chen, X.; Wang, C.; Qiu, H.; Yuan, Y.; Chen, K.; Cao, Z.; Tan, R.X.; Tickner, J.; Xu, J.; Zou, J. Asperpyrone A Attenuates RANKL-Induced Osteoclast Formation through Inhibiting NFATc1, Ca2+ Signalling and Oxidative Stress. J. Cell. Mol. Med. 2019, 23, 8269–8279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Hong, G.; Zhou, L.; Zhang, J.; Fang, J.; He, W.; Tickner, J.; Han, X.; Zhao, L.; Xu, J. Asiaticoside, a Component of Centella Asiatica Attenuates RANKL-Induced Osteoclastogenesis via NFATc1 and NF-ΚB Signaling Pathways. J. Cell. Physiol. 2019, 234, 4267–4276. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Ye, Z.; Wang, C.; Wang, Q.; Wang, H.; Kuek, V.; Wang, Z.; Qiu, H.; Yuan, J.; Kenny, J.; et al. Arctiin Abrogates Osteoclastogenesis and Bone Resorption via Suppressing RANKL-Induced ROS and NFATc1 Activation. Pharmacol. Res. 2020, 159, 104944. [Google Scholar] [CrossRef]
- Wang, Q.; Yao, L.; Xu, K.; Jin, H.; Chen, K.; Wang, Z.; Liu, Q.; Cao, Z.; Kenny, J.; Liu, Y.; et al. Madecassoside Inhibits Estrogen Deficiency-Induced Osteoporosis by Suppressing RANKL-Induced Osteoclastogenesis. J. Cell. Mol. Med. 2019, 23, 380–394. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Lee, K.S.; Yi, S.H.; Kook, S.H.; Lee, J.C. Acteoside Suppresses RANKL-Mediated Osteoclastogenesis by Inhibiting c-Fos Induction and NF-ΚB Pathway and Attenuating ROS Production. PLoS ONE 2013, 8, e80873. [Google Scholar] [CrossRef]
- Jin, G.; Wang, F.F.; Li, T.; Jia, D.D.; Shen, Y.; Xu, H.C. Neogambogic Acid Suppresses Receptor Activator of Nuclear Factor KB Ligand (RANKL)-Induced Osteoclastogenesis by Inhibiting the Jnk and NF-KB Pathways in Mouse Bone Marrow-Derived Monocyte/Macrophages. Med. Sci. Monit. 2018, 24, 2569–2577. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.T.; Kang, W.B.; Zhao, J.N.; Liu, G.; Zhao, M.G. Osteoprotective Effect of Echinocystic Acid, a Triterpone Component from Eclipta Prostrata, in Ovariectomy-Induced Osteoporotic Rats. PLoS ONE 2015, 10, e0136572. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Tang, X.; Liu, Z.; Song, X.; Peng, D.; Zhu, W.; Ouyang, Z.; Wang, W. Hesperetin Prevents Bone Resorption by Inhibiting RANKL-Induced Osteoclastogenesis and Jnk Mediated Irf-3/c-Jun Activation. Front. Pharmacol. 2018, 9, 1028. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Jiang, J.; Yang, Z.; Jin, S.; Lu, X.; Qian, Y. Galangin Suppresses RANKL-Induced Osteoclastogenesis via Inhibiting MAPK and NF-ΚB Signalling Pathways. J. Cell. Mol. Med. 2021, 25, 4988–5000. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.K.; Hyun, C.G. 4-Hydroxy-7-Methoxycoumarin Inhibits Inflammation in LPS-Activated RAW264.7 Macrophages by Suppressing NF-ΚB and MAPK Activation. Molecules 2020, 25, 4424. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Z.; Zhai, Z.; Li, H.; Liu, X.; Qu, X.; Li, X.; Fan, Q.; Tang, T.; Qin, A.; Dai, K. Hypericin Suppresses Osteoclast Formation and Wear Particle-Induced Osteolysis via Modulating ERK Signalling Pathway. Biochem. Pharmacol. 2014, 90, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Kim, H.S.; Lee, J.M.; Choi, J.H.; Kim, S.N.; Kim, D.K.; Kim, J.H.; Mun, S.H.; Kim, J.W.; Jeon, H.S.; et al. Arctigenin Suppresses Receptor Activator of Nuclear Factor Κb Ligand (RANKL)-Mediated Osteoclast Differentiation in Bone Marrow-Derived Macrophages. Eur. J. Pharmacol. 2012, 682, 29–36. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Chen, K.; Deng, T.; Xie, J.; Zhong, K.; Yuan, J.; Wang, Z.; Xiao, Z.; Gu, R.; Chen, D.; et al. Inhibitory Effects of Rhaponticin on Osteoclast Formation and Resorption by Targeting RANKL-Induced NFATc1 and ROS Activity. Front. Pharmacol. 2021, 12, 645140. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, H.; Shinohara, F.; Kajiya, M.; Kodama, T. The Keap1/Nrf2 Protein Axis Plays a Role in Osteoclast Differentiation by Regulating Intracellular Reactive Oxygen Species Signaling. J. Biol. Chem. 2013, 288, 23009–23020. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Corry, K.A.; Loughran, J.P.; Li, J. Moderate Nrf2 Activation by Genetic Disruption of Keap1 Has Sex-Specific Effects on Bone Mass in Mice. Sci. Rep. 2020, 10, 348. [Google Scholar] [CrossRef] [Green Version]
- Pajares, M.; Jiménez-Moreno, N.; García-Yagüe, A.; Escoll, M.; de Ceballos, M.; Van Leuven, F.; Rábano, A.; Yamamoto, M.; Rojo, A.; Cuadrado, A. Transcription Factor NFE2L2/NRF2 Is a Regulator of Macroautophagy Genes. Autophagy 2016, 12, 1902–1916. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Saha, S.; Buttari, B.; Profumo, E.; Saso, L. Activation of Nrf2 Signaling Pathway by Natural and Synthetic Chalcones: A Therapeutic Road Map for Oxidative Stress. Expert Rev. Clin. Pharmacol. 2021, 14, 465–480. [Google Scholar] [CrossRef]
- Sun, Y.-X.; Xu, A.-H.; Yang, Y.; Li, J. Role of Nrf2 in Bone Metabolism. J. Biomed. Sci. 2015, 22, 101. [Google Scholar] [CrossRef] [Green Version]
- Süntar, I.; Çetinkaya, S.; Panieri, E.; Saha, S.; Buttari, B.; Profumo, E.; Saso, L. Regulatory Role of Nrf2 Signaling Pathway in Wound Healing Process. Molecules 2021, 26, 2424. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.-H.; Yang, J.-J.; Tang, P.-J.; Zhu, Y.; Chen, Z.; She, C.; Chen, G.; Cao, P.; Xu, X.-Y. A Novel Keap1 Inhibitor IKeap1 Activates Nrf2 Signaling and Ameliorates Hydrogen Peroxide-Induced Oxidative Injury and Apoptosis in Osteoblasts. Cell Death Dis. 2021, 12, 679. [Google Scholar] [CrossRef]
- Schündeln, M.M.; Höppner, J.; Meyer, F.L.; Schmuck, W.; Kauther, M.D.; Hilken, G.; Levkau, B.; Rauner, M.; Grasemann, C. Prednisone Prevents Particle Induced Bone Loss in the Calvaria Mouse Model. Heliyon 2021, 7, e07828. [Google Scholar] [CrossRef]
- Cho, S.; Sung, Y. Update on Glucocorticoid Induced Osteoporosis. Endocrinol. Metab. 2021, 36, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.; Chen, S.; Peng, H. Glucocorticoid Induces Osteonecrosis of the Femoral Head in Rats through GSK3β-Mediated Osteoblast Apoptosis. Biochem. Biophys. Res. Commun. 2019, 511, 693–699. [Google Scholar] [CrossRef]
- Liu, S.; Fang, T.; Yang, L.; Chen, Z.; Mu, S.; Fu, Q. Gastrodin Protects MC3T3-E1 Osteoblasts from Dexamethasone-Induced Cellular Dysfunction and Promotes Bone Formation via Induction of the NRF2 Signaling Pathway. Int. J. Mol. Med. 2018, 41, 2059–2069. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Mao, Z.; He, S.; Zhan, Y.; Ning, R.; Liu, W.; Yan, B.; Yang, J. Icariin Protects against Glucocorticoid Induced Osteoporosis, Increases the Expression of the Bone Enhancer DEC1 and Modulates the PI3K/Akt/GSK3β/β-Catenin Integrated Signaling Pathway. Biochem. Pharmacol. 2017, 136, 109–121. [Google Scholar] [CrossRef]
- Han, D.; Gu, X.; Gao, J.; Wang, Z.; Liu, G.; Barkema, H.W.; Han, B. Chlorogenic Acid Promotes the Nrf2/HO-1 Anti-Oxidative Pathway by Activating P21 Waf1/Cip1 to Resist Dexamethasone-Induced Apoptosis in Osteoblastic Cells. Free Radic. Biol. Med. 2019, 137, 1–12. [Google Scholar] [CrossRef]
- Yang, J.; Wu, Q.; Lv, J.; Nie, H. 4-Phenyl Butyric Acid Prevents Glucocorticoid-Induced Osteoblast Apoptosis by Attenuating Endoplasmic Reticulum Stress. J. Bone Miner. Metab. 2017, 35, 366–374. [Google Scholar] [CrossRef]
- Jang, W.-G.; Kim, E.-J.; Kim, D.-K.; Ryoo, H.-M.; Lee, K.-B.; Kim, S.-H.; Choi, H.-S.; Koh, J.-T. BMP2 Protein Regulates Osteocalcin Expression via Runx2-Mediated Atf6 Gene Transcription. J. Biol. Chem. 2012, 287, 905–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, V.S.; Ohte, V.; Capelo, L.P.; Gamer, L.; Rosen, V. Specification of Osteoblast Cell Fate by Canonical Wnt Signaling Requires Bmp2. Development 2016, 143, 4352–4367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haxaire, C.; Haÿ, E.; Geoffroy, V. Runx2 Controls Bone Resorption through the Down-Regulation of the Wnt Pathway in Osteoblasts. Am. J. Pathol. 2016, 186, 1598–1609. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.J.; Shen, Y.S.; He, M.C.; Yang, F.; Yang, P.; Pang, F.X.; He, W.; Cao, Y.M.; Wei, Q.S. Polydatin Promotes the Osteogenic Differentiation of Human Bone Mesenchymal Stem Cells by Activating the BMP2-Wnt/β-Catenin Signaling Pathway. Biomed. Pharmacother. 2019, 112, 108746. [Google Scholar] [CrossRef] [PubMed]
- Gough, N.R. Focus Issue: Wnt and β-Catenin Signaling in Development and Disease. Sci. Signal. 2012, 5, eg2. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.A. The Role of the PI3K-AKT Pathway in Melanoma. Cancer J. 2012, 18, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Vega, O.A.; Lucero, C.M.J.; Araya, H.F.; Jerez, S.; Tapia, J.C.; Antonelli, M.; Salazar-Onfray, F.; Las Heras, F.; Thaler, R.; Riester, S.M.; et al. Wnt/β-Catenin Signaling Activates Expression of the Bone-Related Transcription Factor RUNX2 in Select Human Osteosarcoma Cell Types. J. Cell. Biochem. 2017, 118, 3662–3674. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, E.F.; Díez-Pérez, A.; Boonen, S. Update on Long-Term Treatment with Bisphosphonates for Postmenopausal Osteoporosis: A Systematic Review. Bone 2014, 58, 126–135. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.; Suhaimi, F.; Ahmad, F.; Ima-Nirwana, S. The Relationship between Metabolic Syndrome and Osteoporosis: A Review. Nutrients 2016, 8, 347. [Google Scholar] [CrossRef] [Green Version]
- Karadeniz, F.; Oh, J.H.; Lee, J.I.; Seo, Y.; Kong, C.S. 3,5-Dicaffeoyl-epi-Quinic Acid from Atriplex Gmelinii Enhances the Osteoblast Differentiation of Bone Marrow-Derived Human Mesenchymal Stromal Cells via WnT/BMP Signaling and Suppresses Adipocyte Differentiation via AMPK Activation. Phytomedicine 2020, 71, 153225. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, K.; Venkatesan, J.; Kim, S.K. Marine Derived Natural Products for Osteoporosis. Biomed. Prev. Nutr. 2014, 4, 1–7. [Google Scholar] [CrossRef]
- Suvarna, V.; Sarkar, M.; Chaubey, P.; Khan, T.; Sherje, A.; Patel, K.; Dravyakar, B. Bone Health and Natural Products- An Insight. Front. Pharmacol. 2018, 9, 981. [Google Scholar] [CrossRef]
- Zhuang, S.; Yu, R.; Zhong, J.; Liu, P.; Liu, Z. Rhein from Rheum Rhabarbarum Inhibits Hydrogen-Peroxide-Induced Oxidative Stress in Intestinal Epithelial Cells Partly through PI3K/Akt-Mediated Nrf2/HO-1 Pathways. J. Agric. Food Chem. 2019, 67, 2519–2529. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, M.; Hong, S.Y.; Kim, E.Y.; Lee, H.; Jung, H.S.; Sohn, Y. Albiflorin Promotes Osteoblast Differentiation and Healing of Rat Femoral Fractures Through Enhancing BMP-2/Smad and Wnt/β-Catenin Signaling. Front. Pharmacol. 2021, 12, 690113. [Google Scholar] [CrossRef]
- Guo, C.; Yang, R.J.; Jang, K.; Zhou, X.L.; Liu, Y.Z. Protective Effects of Pretreatment with Quercetin Against Lipopolysaccharide-Induced Apoptosis and the Inhibition of Osteoblast Differentiation via the MAPK and Wnt/β-Catenin Pathways in MC3T3-E1 Cells. Cell. Physiol. Biochem. 2017, 43, 1547–1561. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, A.; Posa, F.; De Maria, S.; Ravagnan, G.; Ballini, A.; Porro, C.; Trotta, T.; Grano, M.; Lo Muzio, L.; Mori, G. Polydatin, Natural Precursor of Resveratrol, Promotes Osteogenic Differentiation of Mesenchymal Stem Cells. Int. J. Med. Sci. 2018, 15, 944–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakibaei, M.; Shayan, P.; Busch, F.; Aldinger, C.; Buhrmann, C.; Lueders, C.; Mobasheri, A. Resveratrol Mediated Modulation of Sirt-1/Runx2 Promotes Osteogenic Differentiation of Mesenchymal Stem Cells: Potential Role of Runx2 Deacetylation. PLoS ONE 2012, 7, e35712. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Hu, Y.; Yang, L.; Zhou, J.; Tang, Y.; Zheng, L.; Qin, P. Runx2 Alleviates High Glucose-Suppressed Osteogenic Differentiation via PI3K/AKT/GSK3β/β-Catenin Pathway. Cell Biol. Int. 2017, 41, 822–832. [Google Scholar] [CrossRef]
- Li, N.; Gong, Y. The Mechanism of the Yigutang-Mediated P13K/AKT/GSK-3β Signal Pathway to Regulate Osteogenic Differentiation of Bone Marrow Stromal Stem Cells to Treat Osteoporosis. Evid. Based Complement. Alternat. Med. 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Siroy, A.E.; Davies, M.A.; Lazar, A.J. The PI3K-AKT Pathway in Melanoma. In Genetics of Melanoma; Springer: Berlin, Germany, 2016; pp. 165–180. [Google Scholar]
- Yan, D.Y.; Tang, J.; Chen, L.; Wang, B.; Weng, S.; Xie, Z.; Wu, Z.Y.; Shen, Z.; Bai, B.; Yang, L. Imperatorin Promotes Osteogenesis and Suppresses Osteoclast by Activating AKT/GSK3 β/β-Catenin Pathways. J. Cell. Mol. Med. 2020, 24, 2330–2341. [Google Scholar] [CrossRef]
- Deng, S.; Zhou, J.L.; Fang, H.S.; Nie, Z.G.; Chen, S.; Peng, H. Sesamin Protects the Femoral Head From Osteonecrosis by Inhibiting ROS-Induced Osteoblast Apoptosis in Rat Model. Front. Physiol. 2018, 9, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cakir, E.; Yilmaz, A.; Demirag, F.; Oguztuzun, S.; Sahin, S.; Yazici, U.E.; Aydin, M. Prognostic Significance of Micropapillary Pattern in Lung Adenocarcinoma and Expression of Apoptosis-Related Markers: Caspase-3, Bcl-2, and P53. APMIS 2011, 119, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.H.; Hu, X.Q.; Hu, M.J.; Pan, X.K.; Lu, H.G.; Chen, B.; Wu, B.; Chen, G. Activation of AKT Signaling via Small Molecule Natural Compound Prevents against Osteoblast Apoptosis and Osteonecrosis of the Femoral Head. Am. J. Transl. Res. 2020, 12, 7211–7222. [Google Scholar]
- Wang, J.; Xie, Y.; Feng, Y.; Zhang, L.; Huang, X.; Shen, X.; Luo, X. (-)-Epigallocatechingallate Induces Apoptosis in B Lymphoma Cells via Caspase-Dependent Pathway and Bcl-2 Family Protein Modulation. Int. J. Oncol. 2015, 46, 1507–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, J.; Kim, C.; Cho, K.; Chai, J.; Kim, C.; Choi, S. (-)-Epigallocatechin Gallate Induces Apoptosis, via Caspase Activation, in Osteoclasts Differentiated from RAW 264.7 Cells. J. Periodontal Res. 2007, 42, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Fu, S.; Chen, X.; Chen, C.; Wang, H.; Zhong, L. Epigallocatechin Gallate Affects the Proliferation of Human Alveolar Osteoblasts and Periodontal Ligament Cells, as Well as Promoting Cell Differentiation by Regulating PI3K/Akt Signaling Pathway. Odontology 2021, 109, 729–740. [Google Scholar] [CrossRef]
- Vali, B.; Rao, L.G.; El-Sohemy, A. Epigallocatechin-3-Gallate Increases the Formation of Mineralized Bone Nodules by Human Osteoblast-like Cells. J. Nutr. Biochem. 2007, 18, 341–347. [Google Scholar] [CrossRef]
- Jia, Y.; Jiang, J.; Lu, X.; Zhang, T.; Zhao, K.; Han, W.; Yang, W.; Qian, Y. Garcinol Suppresses RANKL-Induced Osteoclastogenesis and Its Underlying Mechanism. J. Cell. Physiol. 2019, 234, 7498–7509. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Chen, W.; Gu, X.; Shan, R.; Zou, J.; Liu, G.; Shahid, M.; Gao, J.; Han, B. Cytoprotective Effect of Chlorogenic Acid against Hydrogen Peroxide-Induced Oxidative Stress in MC3T3-E1 Cells through PI3K/Akt-Mediated Nrf2/HO-1 Signaling Pathway. Oncotarget 2017, 8, 14680–14692. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Sapkota, M.; Gao, M.; Choi, H.; Soh, Y. Macrolactin F Inhibits RANKL-Mediated Osteoclastogenesis by Suppressing Akt, MAPK and NFATc1 Pathways and Promotes Osteoblastogenesis through a BMP-2/Smad/Akt/Runx2 Signaling Pathway. Eur. J. Pharmacol. 2017, 815, 202–209. [Google Scholar] [CrossRef]
- Wang, S.; Li, F.; Lin, Y.; Wu, B. Targeting REV-ERBα for Therapeutic Purposes: Promises and Challenges. Theranostics 2020, 10, 4168–4182. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Kim, J.; Kim, I.; Seong, S.; Kim, N. Rev-Erbα Negatively Regulates Osteoclast and Osteoblast Differentiation through P38 MAPK Signaling Pathway. Mol. Cells 2020, 43, 34–47. [Google Scholar] [CrossRef]
- Xu, C.; Ochi, H.; Fukuda, T.; Sato, S.; Sunamura, S.; Takarada, T.; Hinoi, E.; Okawa, A.; Takeda, S. Circadian Clock Regulates Bone Resorption in Mice. J. Bone Miner. Res. 2016, 31, 1344–1355. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Tan, P.; Zhang, Z.; Wu, W.; Dong, Y.; Zhao, L.; Liu, H.; Guan, H.; Li, F. REV-ERB Agonism Suppresses Osteoclastogenesis and Prevents Ovariectomy-Induced Bone Loss Partially via FABP4 Upregulation. FASEB J. 2018, 32, 3215–3228. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Wang, J.; Kim, B.; Lu, C.; Zhang, Z.; Liu, H.; Kang, H.; Sun, Y.; Guan, H.; Fang, Z.; et al. Insights into the Role of Circadian Rhythms in Bone Metabolism: A Promising Intervention Target? BioMed Res. Int. 2018, 2018, 9156478. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Zhu, Y.; Sun, H.; Xu, X.; Zhang, S.; Hao, Z.; Wang, G.; Mu, C.; Ren, H. Pharmacological Activation of REV-ERBα Represses LPS-Induced Microglial Activation through the NF-ΚB Pathway. Acta Pharmacol. Sin. 2019, 40, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Lin, Y.; Gao, L.; Yang, Z.; Wang, S.; Wu, B. Circadian Pharmacological Effects of Berberine on Chronic Colitis in Mice: Role of the Clock Component Rev-Erbα. Biochem. Pharmacol. 2020, 172, 113773. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, H.; Zhang, L.; Wang, S.; Lu, D.; Chen, M.; Wu, B. Chronoeffects of the Herbal Medicines Puerariae Radix and Coptidis Rhizoma in Mice: A Potential Role of REV-ERBα. Front. Pharmacol. 2021, 12, 707844. [Google Scholar] [CrossRef]
- Cheon, Y.-H.; Kim, J.-Y.; Baek, J.M.; Ahn, S.-J.; Jun, H.Y.; Erkhembaatar, M.; Kim, M.S.; Lee, M.S.; Oh, J. WHI-131 Promotes Osteoblast Differentiation and Prevents Osteoclast Formation and Resorption in Mice. J. Bone Miner. Res. 2016, 31, 403–415. [Google Scholar] [CrossRef]
- Karadeniz, F.; Ahn, B.-N.; Kim, J.-A.; Seo, Y.; Jang, M.-S.; Nam, K.-H.; Kim, M.; Lee, S.-H.; Kong, C.-S. Phlorotannins Suppress Adipogenesis in Pre-Adipocytes While Enhancing Osteoblastogenesis in Pre-Osteoblasts. Arch. Pharmacal Res. 2015, 38, 2172–2182. [Google Scholar] [CrossRef] [PubMed]
- Kifor, O.; Diaz, R.; Butters, R.; Brown, E.M. The Ca2+-Sensing Receptor (CaR) Activates Phospholipases C, A2, and D in Bovine Parathyroid and CaR-Transfected, Human Embryonic Kidney (HEK293) Cells. J. Bone Miner. Res. 1997, 12, 715–725. [Google Scholar] [CrossRef]
- Guha, S.; Paul, C.; Alvarez, S.; Mine, Y.; Majumder, K. Dietary γ-Glutamyl Valine Ameliorates TNF-α-Induced Vascular Inflammation via Endothelial Calcium-Sensing Receptors. J. Agric. Food Chem. 2020, 68, 9139–9149. [Google Scholar] [CrossRef]
- Feng, R.; Ding, F.; Mi, X.-H.; Liu, S.-F.; Jiang, A.-L.; Liu, B.-H.; Lian, Y.; Shi, Q.; Wang, Y.-J.; Zhang, Y. Protective Effects of Ligustroflavone, an Active Compound from Ligustrum Lucidum, on Diabetes-Induced Osteoporosis in Mice: A Potential Candidate as Calcium-Sensing Receptor Antagonist. Am. J. Chin. Med. 2019, 47, 457–476. [Google Scholar] [CrossRef]
- Nam, G.S.; Lee, K.S.; Nam, K.S. Morin Hydrate Inhibits Platelet Activation and Clot Retraction by Regulating Integrin AIIbβ3, TXA2, and CAMP Levels. Eur. J. Pharmacol. 2019, 865, 172734. [Google Scholar] [CrossRef] [PubMed]
- Kumrungsee, T.; Saiki, T.; Akiyama, S.; Nakashima, K.; Tanaka, M.; Kobayashi, Y.; Matsui, T. Inhibition of Calcium-Calmodulin Complex Formation by Vasorelaxant Basic Dipeptides Demonstrated by in Vitro and in Silico Analyses. Biochim. Biophys. Acta - Gen. Subj. 2014, 1840, 3073–3078. [Google Scholar] [CrossRef] [PubMed]
- Cianferotti, L.; Gomes, A.; Fabbri, S.; Tanini, A.; Brandi, M. The Calcium-Sensing Receptor in Bone Metabolism: From Bench to Bedside and Back. Osteoporos. Int. 2015, 26, 2055–2071. [Google Scholar] [CrossRef] [PubMed]
- Arora, H. Pea Protein Derived Bioactive Peptides Stimulate Bone Health Promoting Effects. Master’s Thesis, University of Alberta, Edomton, AB, Canada, 2020. [Google Scholar]
- Bayle, M.; Neasta, J.; Dall’Asta, M.; Gautheron, G.; Virsolvy, A.; Quignard, J.-F.; Youl, E.; Magous, R.; Guichou, J.-F.; Crozier, A.; et al. The Ellagitannin Metabolite Urolithin C Is a Glucose-Dependent Regulator of Insulin Secretion through Activation of L-Type Calcium Channels. Br. J. Pharmacol. 2019, 176, 4065–4078. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Chu, X.; Wang, H.; Zhang, X.; Zhang, Y.; Guo, H.; Liu, Z.; Dong, Y.; Liu, H.; Liu, Y.; et al. Crocin, a Carotenoid Component of Crocus Cativus, Exerts Inhibitory Effects on L-Type Ca2+ Current, Ca2+ Transient, and Contractility in Rat Ventricular Myocytes. Can. J. Physiol. Pharmacol. 2015, 94, 302–308. [Google Scholar] [CrossRef]
- Wong, J.; Chiang, Y.-F.; Shih, Y.-H.; Chiu, C.-H.; Chen, H.-Y.; Shieh, T.-M.; Wang, K.-L.; Huang, T.-C.; Hong, Y.-H.; Hsia, S.-M. Salvia Sclarea L. Essential Oil Extract and Its Antioxidative Phytochemical Sclareol Inhibit Oxytocin-Induced Uterine Hypercontraction Dysmenorrhea Model by Inhibiting the Ca2+–MLCK–MLC20 Signaling Cascade: An Ex Vivo and In Vivo Study. Antioxidants 2020, 9, 991. [Google Scholar] [CrossRef]
- Hardy, N.; Viola, H.M.; Johnstone, V.P.A.; Clemons, T.D.; Szappanos, H.C.; Singh, R.; Smith, N.M.; Iyer, K.S.; Hool, L.C. Nanoparticle-Mediated Dual Delivery of an Antioxidant and a Peptide against the L-Type Ca2+ Channel Enables Simultaneous Reduction of Cardiac Ischemia-Reperfusion Injury. ACS Nano 2014, 9, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaweewannakorn, W.; Ariyoshi, W.; Okinaga, T.; Fujita, Y.; Maki, K.; Nishihara, T. Ameloblastin Attenuates RANKL-Mediated Osteoclastogenesis by Suppressing Activation of Nuclear Factor of Activated T-Cell Cytoplasmic 1 (NFATc1). J. Cell. Physiol. 2019, 234, 1745–1757. [Google Scholar] [CrossRef]
- Posa, F.; Colaianni, G.; Di Cosola, M.; Dicarlo, M.; Gaccione, F.; Colucci, S.; Grano, M.; Mori, G. The Myokine Irisin Promotes Osteogenic Differentiation of Dental Bud-Derived MSCs. Biology 2021, 10, 295. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Gong, Z.; Zhao, R.; Zhu, Y. Melatonin Inhibits RANKL-Induced Osteoclastogenesis through the MiR-882/Rev-Erbα Axis in Raw264.7 Cells. Int. J. Mol. Med. 2021, 47, 633–642. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, I.J.; Tsai, H.-C.; Chang, A.-C.; Huang, C.-C.; Yang, S.-F.; Tang, C.-H. Melatonin Inhibits Osteoclastogenesis and Osteolytic Bone Metastasis: Implications for Osteoporosis. Int. J. Mol. Sci. 2021, 22, 9435. [Google Scholar] [CrossRef]
- Cheon, Y.H.; Lee, C.H.; Kim, S.; Park, G.D.; Kwak, S.C.; Cho, H.J.; Kim, J.Y.; Lee, M.S. Pitavastatin Prevents Ovariectomy-Induced Osteoporosis by Regulating Osteoclastic Resorption and Osteoblastic Formation. Biomed. Pharmacother. 2021, 139. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, D.; Jin, H.; Ye, Z.; Wang, C.; Chen, K.; Kuek, V.; Xu, K.; Qiu, H.; Chen, P.; et al. Hymenialdisine: A Marine Natural Product That Acts on Both Osteoblasts and Osteoclasts and Prevents Estrogen-Dependent Bone Loss in Mice. J. Bone Miner. Res. 2020, 35, 1582–1596. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Ma, C.; Chen, K.; Wang, Z.; Qiu, H.; Chen, D.; He, J.; Zhang, C.; Guo, D.; Lai, B.; et al. Cycloastragenol Attenuates Osteoclastogenesis and Bone Loss by Targeting RANKL-Induced Nrf2/Keap1/ARE, NF-ΚB, Calcium, and NFATc1 Pathways. Front. Pharmacol. 2022, 12, 810322. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zeng, W.; Ma, C.; Wang, Z.; Wang, C.; Li, S.; He, W.; Zhang, Q.; Xu, J.; Zhou, C. Maackiain Dampens Osteoclastogenesis via Attenuating RANKL-stimulated NF-κB Signalling Pathway and NFATc1 Activity. J. Cell. Mol. Med. 2020, 24, 12308. [Google Scholar] [CrossRef]
- Sun, K.; Zhu, J.; Deng, Y.; Xu, X.; Kong, F.; Sun, X.; Huan, L.; Ren, C.; Sun, J.; Shi, J. Gamabufotalin Inhibits Osteoclastgenesis and Counteracts Estrogen-Deficient Bone Loss in Mice by Suppressing RANKL-Induced NF-ΚB and ERK/MAPK Pathways. Front. Pharmacol. 2021, 12, 629968. [Google Scholar] [CrossRef] [PubMed]
- Carnovali, M.; Ciavatta, M.L.; Mollo, E.; Roussis, V.; Banfi, G.; Carbone, M.; Mariotti, M. Aerophobin-1 from the Marine Sponge Aplysina Aerophoba Modulates Osteogenesis in Zebrafish Larvae. Mar. Drugs 2022, 20, 135. [Google Scholar] [CrossRef]
- Hairi, H.A.; Jamal, J.A.; Aladdin, N.A.; Husain, K.; Mohd Sofi, N.S.; Mohamed, N.; Mohamed, I.N.; Shuid, A.N. Demethylbelamcandaquinone B (Dmcq B) Is the Active Compound of Marantodes Pumilum Var. Alata (Blume) Kuntze with Osteoanabolic Activities. Molecules 2018, 23, 1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Chu, F.; Zhang, G.; Wang, Q.; Li, Y.; Zhang, M.; He, Q.; Yang, J.; Wang, H.; Sun, P.; et al. 12-Deoxyphorbol 13-Acetate Inhibits RANKL-Induced Osteoclastogenesis via the Attenuation of MAPK Signaling and NFATc1 Activation. Int. Immunopharmacol. 2021, 101, 108177. [Google Scholar] [CrossRef]
- Meng, Y.; Yin, D.; Qiu, S.; Zhang, X. Abrine Promotes Cell Proliferation and Inhibits Apoptosis of Interleukin-1β-Stimulated Chondrocytes via PIM2/VEGF Signalling in Osteoarthritis. Phytomedicine 2022, 96, 153906. [Google Scholar] [CrossRef] [PubMed]
- Xian, Y.; Su, Y.; Liang, J.; Long, F.; Feng, X.; Xiao, Y.; Lian, H.; Xu, J.; Zhao, J.; Liu, Q.; et al. Oroxylin A Reduces Osteoclast Formation and Bone Resorption via Suppressing RANKL-Induced ROS and NFATc1 Activation. Biochem. Pharmacol. 2021, 193, 114761. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jin, H.; Shim, H.; Kim, H.; Ha, H.; Lee, Z. Epigallocatechin-3-Gallate Inhibits Osteoclastogenesis by down-Regulating c-Fos Expression and Suppressing the Nuclear Factor-KappaB Signal. Mol. Pharmacol. 2010, 77, 17–25. [Google Scholar] [CrossRef]
- Shen, C.L.; Chyu, M.C.; Yeh, J.K.; Zhang, Y.; Pence, B.C.; Felton, C.K.; Brismée, J.M.; Arjmandi, B.H.; Doctolero, S.; Wang, J.S. Effect of Green Tea and Tai Chi on Bone Health in Postmenopausal Osteopenic Women: A 6-Month Randomized Placebo-Controlled Trial. Osteoporos. Int. 2012, 23, 1541–1552. [Google Scholar] [CrossRef] [Green Version]
- Morato-Martínez, M.; López-Plaza, B.; Santurino, C.; Palma-Milla, S.; Gómez-Candela, C. A Dairy Product to Reconstitute Enriched with Bioactive Nutrients Stops Bone Loss in High-Risk Menopausal Women without Pharmacological Treatment. Nutrients 2020, 12, 2203. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F.; Lamb, J.J.; Lerman, R.H.; Konda, V.R.; Darland, G.; Minich, D.M.; Desai, A.; Chen, T.C.; Austin, M.; Kornberg, J.; et al. Hop Rho Iso-Alpha Acids, Berberine, Vitamin D3 and Vitamin K1 Favorably Impact Biomarkers of Bone Turnover in Postmenopausal Women in a 14-Week Trial. J. Bone Miner. Metab. 2010, 28, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Lamb, J.J.; Holick, M.F.; Lerman, R.H.; Konda, V.R.; Minich, D.M.; Desai, A.; Chen, T.C.; Austin, M.; Kornberg, J.; Chang, J.L.; et al. Nutritional Supplementation of Hop Rho Iso-Alpha Acids, Berberine, Vitamin D3, and Vitamin K1 Produces a Favorable Bone Biomarker Profile Supporting Healthy Bone Metabolism in Postmenopausal Women with Metabolic Syndrome. Nutr. Res. 2011, 31, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.N.T.; Thorup, A.C.; Hansen, E.S.S.; Jeppesen, P.B. Combined Red Clover Isoflavones and Probiotics Potently Reduce Menopausal Vasomotor Symptoms. PLoS ONE 2017, 12, e0176590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Sugihara, F.; Suzuki, K.; Inoue, N.; Venkateswarathirukumara, S. A Double-Blind, Placebo-Controlled, Randomised, Clinical Study on the Effectiveness of Collagen Peptide on Osteoarthritis. J. Sci. Food Agric. 2015, 95, 702–707. [Google Scholar] [CrossRef]
- Yong, E.L.; Cheong, W.F.; Huang, Z.; Thu, W.P.P.; Cazenave-Gassiot, A.; Seng, K.Y.; Logan, S. Randomized, Double-Blind, Placebo-Controlled Trial to Examine the Safety, Pharmacokinetics and Effects of Epimedium Prenylflavonoids, on Bone Specific Alkaline Phosphatase and the Osteoclast Adaptor Protein TRAF6 in Post-Menopausal Women. Phytomedicine 2021, 91, 153680. [Google Scholar] [CrossRef]
- Argyrou, C.; Karlafti, E.; Lampropoulou-Adamidou, K.; Tournis, S.; Makris, K.; Trovas, G.; Dontas, I.; Triantafyllopoulos, I.K. Effect of Calcium and Vitamin D Supplementation with and without Collagen Peptides on Bone Turnover in Postmenopausal Women with Osteopenia. J. Musculoskelet. Neuronal Interact. 2020, 20, 12. [Google Scholar] [PubMed]
- Tu, M.Y.; Chen, H.L.; Tung, Y.T.; Kao, C.C.; Hu, F.C.; Chen, C.M. Short-Term Effects of Kefir-Fermented Milk Consumption on Bone Mineral Density and Bone Metabolism in a Randomized Clinical Trial of Osteoporotic Patients. PLoS ONE 2015, 10, e0144231. [Google Scholar] [CrossRef]
- Al-Dashti, Y.A.; Holt, R.R.; Carson, J.G.; Keen, C.L.; Hackman, R.M. Effects of Short-Term Dried Plum (Prune) Intake on Markers of Bone Resorption and Vascular Function in Healthy Postmenopausal Women: A Randomized Crossover Trial. J. Med. Food 2019, 22, 982–992. [Google Scholar] [CrossRef]
- Zhang, T.; Han, W.; Zhao, K.; Yang, W.; Lu, X.; Jia, Y.; Qin, A.; Qiang, Y. Psoralen Accelerates Bone Fracture Healing by Activating Both Osteoclasts and Osteoblasts. FASEB J. 2019, 33, 5399–5410. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Christensen, L.A.; Legerski, R.J.; Vasquez, K.M. Mismatch Repair Participates in Error-Free Processing of DNA Interstrand Crosslinks in Human Cells; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2005; Volume 6. [Google Scholar]
- Fitzpatrick, T.K.; Momtaz, T.B. The benefits and risks of long-term puva photochemotherapy. Dermatol. Clin. 1998, 16, 227–234. [Google Scholar] [CrossRef]
- Chen, M.; Zhou, C.; Xu, H.; Zhang, T.; Wu, B. Chronopharmacological Targeting of Rev-Erbα by Puerarin Alleviates Hyperhomocysteinemia in Mice. Biomed. Pharmacother. 2020, 125, 109936. [Google Scholar] [CrossRef]
- Ebguna, C.; Alan, T.; Nwosu, O.; Olumayowa, V.; Kryeziu, T.L.; Saravanan, K.; Ifemeie, J. Toxic Plants and Phytochemicals. In Phytochemistry: Pharmacognosy, Nanomedicine, and Contemporary Issues; Apple Academic Press, Tailor & Francis: Waretown, NJ, USA, 2019; Volume 2. [Google Scholar]
- Peumans, W.J.; Van Damme, E.J.M. Lectins as Plant Defense Proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef]
- Tang, X.T.; Ibanez, F.; Tamborindeguy, C. Concanavalin A Toxicity Towards Potato Psyllid and Apoptosis Induction in Midgut Cells. Insects 2020, 11, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetri, V.; Carrotta, R.; Picone, P.; Di Carlo, M.; Militello, V. Concanavalin A Aggregation and Toxicity on Cell Cultures. Biochim. Biophys. Acta 2010, 1804, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; He, S.; Zhou, F.; Sun, H.; Cao, X.; Ye, Y.; Li, J. Detection of Lectin Protein Allergen of Kidney Beans (Phaseolus Vulgaris L.) and Desensitization Food Processing Technology. J. Agric. Food Chem. 2021, 69, 14723–14741. [Google Scholar] [CrossRef]
- Salem, M.A.; Serag, A.; El-Seedi, H.R.; Hamdan, D.I.; Ezzat, S.M.; Zayed, A. Identification and Analysis of Toxic Phytochemicals: Phytotoxins and Natural Defenses. In Phytochemistry, the Military and Health; Mtewa, A.G., Egbuna, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 443–479. ISBN 978-0-12-821556-2. [Google Scholar]
- Okagu, I.U.; Ndefo, J.C.; Aham, E.C.; Udenigwe, C.C. Zanthoxylum Species: A Review of Traditional Uses, Phytochemistry and Pharmacology in Relation to Cancer, Infectious Diseases and Sickle Cell Anemia. Front. Pharmacol. 2021, 12, 713090. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.M.; Dong, Z. Toxic Phytochemicals and Their Potential Risks for Human. Cancer Prev. Res. 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Xiao, H.; Zhang, X.; Zhou, P. Insight into the Effects of Deglycosylation and Glycation of Shrimp Tropomyosin on in Vivo Allergenicity and Mast Cell Function. Food Funct. 2019, 10, 3934–3941. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Compound | Source | Study Model | Specific Therapeutic Activity | Ref. |
|---|---|---|---|---|
| Alisol-B (1) | Alisma orientale Juzepczuk | 1α,25(OH)2D3-induced OCG in BMMs and primary osteoblasts | Prevented bone loss (50–100%) by downregulating the expression of pJNK, NFATc1, and CFos. Reduced serum Ca2+ level at 0.5–5 μM. | [84] |
| Desoxyrhapontigenin (2) | Rheum undulatum | RANKL-exposed BMMs | At 10 μM, it inhibited OCG by 90% compared to resveratrol (70%) and reduced bone resorption (60–95%) at 3–30 μM by downregulating the expression of MMP-9, TRAP, CTSK, and NFATc1. | [85] |
| Lumichrome (3) | Milk | OVX mice | Inhibited bone loss by 2-fold at 10 μM. | [87] |
| Maslinic acid (4) | Platostoma africanum P. beauv. | OVX mice | Prevented loss of bone mineral density (BMD). | [92] |
| RANKL-induced OCG in BMMs and MMCs | At 10 μM, inhibited RANKL-induced OCG (3- and 5-fold), MAPK, actin ring formation (5-fold) by blocking activation of AP-1, TRAF6, TRAP, MM9, s-Src, CTR, and CTSK, and activation of IκBα. | |||
| Lycorine (5) | Amaryllidaceae SPP. | RANKL-induced OCG in BMMs | Inhibited RANKL-induced OCs expression and bone resorption by 70–99% at 0.1–0.4 μM, thereby suppressing the expression of TRAP (70%), CTR (67%), NFATc1 (33%), CTR (60%), DC-STAMP (75%), and CTSK (90%) at 0.4 μM at day 5. | [93] |
| OVX mice | Prevented OVX-induced bone loss and titanium particle-induced osteolysis reducing expression level of TRAP, V-ATPase, NFATc1, CTR, and CTSK between 25% and 90% at 0.4 μM compared to control, but increased TN, TT, and BV/TV levels by at least 20% at 0.5–2.5 mg/kg. | |||
| Isoliquiritigenin (6) | Glycyrrhiza glabra L. | RANKL-induced OCG in BMMs and RAW 264.7 cells | Inhibited OC differentiation in both cells. At 20 μM, reduced bone resorption (95%), gene expression of NFATc1, C-Fos, TRAP, CatK, and MMP-9 between 50% and 98%. | [94] |
| C57/BL6 mice | Prevented LPS-induced bone erosion in mice by 65% and reduced OCs formation by 55%. | |||
| Astilbin (7) | Astilbe thunbergii (Siebold and Zucc.) Miq. | OVX mouse | Reduced OCs and TS levels by 10-fold while increasing the expression of BVD and TN by over 50% at 10 mg/kg for 6 weeks. | [95] |
| RANKL-induced OCG in BMMs | Decreased expression of NFATc1, CFos, ACP5, CTSK, NF-κB, JNK, ERK, and p38, and Ca2+ oscillation by 50%, 50%, 50%, 70%, 70%, 50%, and 30%, respectively, while increasing IκBα by 1% at 10 μmol/L in cell culture after five days treatment. | |||
| Notopterol (9) | Notopterygium incisum K.C.Ting ex H.T.Chang | OVX mice | Inhibited bone loss by reducing F-actin ring formation and resorption indices by 50% at 10 mg/kg for 6 weeks. | [97] |
| RANKL-induced OCG in BMMs and MMCs | The gene expression profiles of CTSK, ACP5, V-ATPase-d2, JNK, ERK, NFATc1, CFos, TRAP, CTR, MMP-9, NF-kB, and TRAF6 and ROS production were decreased by between 30% and 75% while the expressions of IκBα, GSR, CAT, NQO1, Nrf2/Keap1, and ARE were increased by 25%, 55%, 50%, 30%, 33%, and 300%, respectively, at 10 μM after 5 days treatment. | |||
| Protocatechuic acid (10) | Rubus coreanus Miquel | OVX mouse | At 20 mg/kg/d, protocatechuic acid attenuated OVX-induced bone loss by reducing gene expression profiles of RANKL, ALP, OCN, CTR, CTSK, NFATc1, and TRAF6 by 45%, 500%, 25%, 40%, and 40%, respectively, and BALP level by 5-fold, while increasing OPG level by 3-fold after 12 weeks. | [98] |
| Sarsasapogenin (11) | Anemarrhena asphodeloides Bunge | LPS-induced osteolysis in mouse | Prevented LPS-induced bone loss in a dose-dependent manner at 5–10 mg/kg/d for one week. | [99] |
| RANKL-induced OCG in BMMs | Inhibited RANKL-induced OCG by suppressing the gene expression of CFos, NFATc1, MAPK, F-actin, Nfatc1, ACP5, and CTSK by over 60% each at 4 μM. | |||
| Kavain (12) | Piper methysticum | OVX mouse | Inhibited bone loss and OCG by 50% and elevated protein levels of BVD and TN at 10 mg/kg i.p., 3x/week for 6 weeks. | [100] |
| MMCs, MCPs and BMMs | Reduced the expression of NFATc1, MMP-9, ACP5, Atp6v0d2, integrin β3, JNK, p38, ERK, and CTSK, and Ca2+ oscillation generally by more than 30% each at 20 μM on day 6. | |||
| Helvolic acid (13) | Aspergillus fumigatus | RANKL-induced OCG in BMMs | Reduced OCs by 60% through reduced expression of NFATc1, MAPK, MMP-9, ACP5, and CTSK, and Ca2+ oscillation by over 80% at 10 μM for five days | [101] |
| Berberine (14) | Coptis chinensis | LPS-generated bone loss in mice | Dose-dependently abrogated LPS-engineered bone loss in mice between 50 and 150 mg/kg/d for 10 days. | [102] |
| RANKL-induced OCG in BMMs | At 5 μM, inhibited OCG via reduction in NFATc1, CFos, p38, ERK, TRAP, OSCAR, and ATP6v0d2 expression and F-actin ring formation by more than 80% and increased IκBα expression by over 60% after 7 days of treatment. | |||
| Asperpyrone A (15) | Aspergillus niger | RANKL-induced OCG in BMMs | Prevented OCG via decrease in gene expression of NFATc1, CFos, CTSK, MAPK, and NF-κB by 66%, 66%, 40%, and 33%, respectively, and ROS production by 75% at 10 μmol/L for 6 days. | [103] |
| Asiaticoside (16) | Centella asiatica (L.) Urb. | RANKL-induced OCG in BMMs | Reduced OCG by 50% at 20 μM via suppression of NFATc1, CFos, MAPKs, MMP-9, ACP5, CTSK, NF-κB, TRAP, and ATP6v0d2 expression, and Ca2+ oscillation by over 30% each at 20 μM after 3 days. | [104] |
| Arctiin (17) | Arctium lappa L. | OVX mice | At 10 mg/kg/48 h, i.p., arctiin prevented bone loss and RANKL-induced OCG through the reduction in expression of NFATc1, ACP5, CFos, TRAP, ERK, JNK, MMP-9, and CTSK, and Ca2+ oscillation and F-actin ring formation by over 25% each at 20 μM for six days. | [105] |
| RANKL-induced OCG in BMMs | ||||
| Madecassoside (18) | Centella asiatica (L.) Urb | OVX mice | Prevented bone loss and increased BMD, trabecular number, Oc.S/BS, and N.Oc/BS by over 25% each 10 mg/kg/48 h, i.p. for 6 days. | [106] |
| RANKL-induced OCG in BMMs | At 10 μmol/L, madecassoside reduced OCs by over 15% via suppression of NFATc1, CFos, ACP5 TRAP, ERK, NF-κB, JNK, MMP-9, CTSK, and integrin β3 expression, and Ca2+ oscillation and F-actin ring formation by more than 30% while upregulating IκBα expression by 50% for 3–6 days. | |||
| Acteoside (19) | Rehmannia glutinosa | OVX mice | Prevented bone loss in mice and decreased serum levels of TNF-α, IL-1β, TRAP, and OCN, and Ca2+ oscillation by more than 30% at 1 mM/72 h for eight weeks. | [107] |
| RANKL-induced OCG in BMMs and MMCs | It also attenuated OC by over 80% at 20 µM through reduction in expression of NF-kB, TNF-α, IL-1β, TRAP, Ca2+ oscillation, CFos, NFATc1, ERK, and JNK by over 70% each after 24 h. | |||
| Isoliquiritigenin (20) | Glycyrrhiza glabra L. | LPS-induced mice | Inhibited LPS-induced bone erosion by 90% via reduction in F-actin ring formation, lacunar resorption pits formation, and OcS/BS by over 30% after 8 weeks of treatment. | [94] |
| RANKL-induced OCG in BMMs and MMCs | Inhibited OC via suppression of gene expression profiles of NF-kB, TRAF6, MAPKs, AP-1, CTSK, ERK, NFATc1, CFos, TRAP, CatK, MMP-9, and OSCAR by over 30% each and upregulated IκBα, OPG, and osteoblastic marker genes by over 30% at 50 μM for 72 h. | |||
| Neogambogic acid (21) | Garcinia hanburyi | RANKL-induced OCG in BMMs | Neogambogic acid inhibited RANKL-induced OCG by 87.5% via the suppression of gene expression of NF-kB, JNK, TRAPs, CTR, CTSK, and NFATc1 by 60%, 40%, 50%, and 40%, respectively, at 0.4 μg/mL for 6 days. | [108] |
| Echinocystic acid (22) | Eclipta prostrata | OVX mouse model | Inhibited loss in bone mass, strength, and density, and reduced serum OCN, ALP, and deoxypyridinoline levels by 60%, 25%, and 35%, respectively, and improved bone histology with concurrent downregulation of expression profiles of IL-1β and TNF-α by 40% and 30%, respectively, in bone tissues at 15 mg/kg/d for 12 weeks. | [109] |
| Cimiracemate A (23) | Cimicifuga racemose L. | Glucocorticoid-induced OP in rats | Inhibited bone loss as characterized by increase in levels and gene expression profiles of TN, TT, BV/TV, HDL, OCN, and OPG by 50%, 40%, 150%, 136%, 130%, and 150%, respectively, with concurrent reduction in serum level and gene expression of TS, SIM, LDL, TC, ApoA1, ApoB, TAG, CTX, IL-1β, TNF-α, TRAP, RANK, and RANKL between 26% and 50%, respectively, at 10 mg/kg for 6 weeks. | [96] |
| Hesperetin (24) | Citrus fruit | OVX mice | Inhibited bone resorption by increasing the levels of TN, TT, BV/TV, and ALP by 35%, 17%, 125%, and 40%, respectively, with concurrent reductions in levels of TS, ALP, OC, TRAP, and CTX by over 17% each at 11 mg/kg i.p., 3x/week for 8 weeks. Inhibited OC by suppressing gene expression of CTSK, JNK, ERK, NFATc1, CFos, TRAP, ACP5, CTX, MMP-9, NF-kB, TRAF6, and Irf-3, by over 22% in each at 60 μM for 10 days. | [110] |
| RANKL-induced OCG in BMCs, BMMs, and splenocytes | ||||
| Galangin (25) | Alpinia officinarum | LPS-induced osteolysis in mice | Inhibited bone loss by reducing F-actin ring formation and resorption indices by 90% and 75%, respectively, and increasing TN, TT, and BV/TV by 5% each at 10 mg/kg for 7 days. | [111] |
| RANKL-induced OCG in BMCs | Inhibited OC differentiation by reducing the gene expression of TRAP, CTSK, p38, ERK, NFATc1, and CFos by 40%, 55%, 57%, 50%, and 75%, respectively, while increasing IκBα and p65 gene expression profiles at 12 μmol/L for 6 days. | |||
| 4-hydroxy-7-methoxycoumarin (26) | Cynara scolymus L. | RANKL-induced OCG in MMCs | Inhibited OCG via promotion of gene expression of IκBα by 10-fold while downregulating the expression of TNF-α, IL-1β, IL-6, NO, PGE2, iNOS, COX-2; NF-kB, ERK, and JNK by over 45% each at 1 mM. | [112] |
| Hypericin (27) | Hypericum perforatum L. | Titanium particle-induced osteolysis in mice | Inhibited bone loss reducing bone level of NP and PP, with concomitant increases in TN, TT, and BV/TV levels by at least 85% at 15 μg/kg for 10 days. | [113] |
| RANKL-induced OCG in BMCs and BMMs | Inhibited OC formation by 88% by reducing the gene expression of resorption indices, NFATc1, CFos, ERK, MMP-9, ACP5, CTSK, NF-κB, CTR, TRAP, and ATP6v0d2, and F-actin ring formation and Ca2+ oscillation by over 65% each at 1.2 μM for 2 days. | |||
| Arctigenin (28) | Arctium lappa L. | LPS-induced bone erosion in mice | Inhibited bone loss by increasing bone levels of TN, TT, and BV/TV by 85% each, while decreasing the levels of F-actin ring formation, resorption indices, and OC over 50% at 10 mg/kg for one week. | [114] |
| RANKL-induced OCG in BMMs | Inhibited OCG by suppressing the gene expression of Syk, PLCγ2, Gab2, ERK, NFATc1, CFos, TRAF6, c-Src, and CTSK by 30% and above at 10 μM. | |||
| Rhaponticin (29) | Rheum undulatum L. | RANKL-induced OCG in BMMs | Inhibited OC formation by increasing the expression of IκBα b 50% while downregulating the expression of F-actin ring formation, OC, resorption indices, NFATc1, CFos, CTSK, ATP6v0d2, and integrin β3, and ROS production, and Ca2+ oscillation by over 30% at 50 μM for 6 days. | [115] |
| Natural Compound | Source | Study Model | Specific Therapeutic Activity | Ref. |
|---|---|---|---|---|
| Icariin (31) | Herba Epimedii | Hydrogen peroxide-induced MC3T3-E1 cell oxidative damage | Alleviated H2O2-induced cytotoxicity and inhibitory osteogenic effect and promoted osteogenesis by increasing the levels of GSH and SOD by 140% and 120%, respectively, and decreasing MDA level by 47%, while the gene expression profiles of ALP, RUNX2-2, OSX, β-catenin, and cyclin D1 increased by 167–233%, at 0.1 μM for 48 h. Promoted cell proliferation and improved osteogenic differentiation and calcium deposit by enhancing the expression and levels of RUNX2, DEC1, BV/TV, p-GSK3β p-Akt, and Tb.Th by 100%, 90%, 11%, and 10%, respectively, at 1 nM. Protected against glucocorticoid-induced osteoporosis by enhancing gene expression of DEC1, ALP, and RUNX2 by 50%, 40%, and 50%, respectively, in bone tissues and mineralization by ~50% at 250 mg/kg for 60 days. | [128] |
| SaoS-2 cells and mice | ||||
| Chlorogenic acid (32) | Flos Lonicerae Japonicae, Eucommia Ulmoides | MC3T3-E1 cells | Reversed inhibition of cell viability and reduced H2O2-induced oxidative damage and suppressed apoptosis (caspase-3 level) by ~44%, while increasing gene expression of HO-1, Nrf-2, and p-Akt by over 2-fold each at 400 μM for 48 h. | [160] |
| Albiflorin (34) | Paeonia lactiflora | MC3T3-E1 cells | Induced and promoted OBG by 40%, and improved bone union by upregulating the gene expression of RUNX2, ALP, OCN, OSN, OSX, BSP, OPN, BMP-2, p-Smad1/5, Wnt10b, β-catenin, LRP5, LRP6, Dvl2, and cyclin D1 by 50%, 20%, 40%, 50%, 25%, 50%, 33%, 22.2%, and 50%, respectively, in bone tissues at 10 mg/kg for 3 weeks. | [144] |
| Quercetin (36) | Allium cepa | MC3T3-E1 cells | Reversed LPS-induced suppression of bone mineralization and cell viability and reduced LPS-induced osteoblast apoptosis by 53% and totally restored inhibited osteoblast differentiation by increasing OSX, ALP, RUNX2, OCN, Wnt3, β-catenin, p-ERK1/2, Bcl-2, and Bcl-XL gene expression by 300%, 600%, 500%, 320%, 57%, 200%, and 150%, respectively, while suppressing gene expression of caspase-3, BAX, Cyt-C, MAPK, ERK, p-p38, and GSK3β by over 32% each at 10–50 μM for 7 days. | [145] |
| Polydatin (37) | Polygonum Cuspidatum | hBMSCs | Improved the proliferation and osteogenic differentiation by 183% at 10 μM and induced the formation of calcium nodules through the elevated expression of ALP, RUNX2, OPN, DLX5, OCN, COL1A1, BMP2, β-catenin, LEF1, TCF7, c-jun, c-myc, and cyclin D between 30% and 475% at 30 μM for 14 days. | [134] |
| 3,5-dicaffeoyl-epi-quinic acid (38) | Atriplex gmelinii | hBM-MSCs | Enhanced osteoblast differentiation of osteo-induced hBM-MSCs by upregulating ALP, OCN, RUNX2, BMP2, Wnt 10a, p-p38, p-ERK, p-JNK, β-catenin, and Smad1/5 gene expression between 20% and 180%, while suppressing the expression of PPARγs and C/EBPα, and SREBP1c by 27% and 10%, respectively, at 10 mg/kg for 7 days. | [140] |
| Epigallocatechin (40) | Green tea | hPDLCs and hAOBs | Increased proliferation, differentiation, and osteoblast formation in hAOBs and hPDLCs by 100% and 150% via enhancement of ALP, RUNX2, BMP2, OSX, OCN, p-P13K, and p-Akt gene expression by 40%, 200%, 400%, 600%, 400%, 150%, and 150%, respectively, in hAOBs and 133%, 600%, 500%, 400%, 300%, 180%, and 160%, respectively, in hPDLCs at 10 μM for 5 days. | [157] |
| Sesamin (42) | Sesamum indicum | Rat primary osteoblasts | Inhibited ROS-induced osteoblast apoptosis by 75% by upregulating p-AKT and Bcl-2 gene expression by 300% and 400%, respectively, while downregulating the expression of Bax, caspase-3, and ROS production by 63%, 67%, and 70%, respectively, at 5–20 µM for 24 h. | [152] |
| Garcinol (43) | Garcinia indica | RANKL-induced OCG in BMMs | Attenuated bone resorption and Suppressed osteoclast differentiation through the downregulation of TRAP, CTSK, NFATc1, V-ATPase d2, p-ERK, p-p38, p-JNK, p-p65, p-Akt, C-jun, and Cfos gene expression and F-actin ring formation by 81%, 75%, 75%, 75%, 90%, 55%, 40%, 50%, 100%, 50%, 50%, and 69%, respectively, at 32 μM for 7 days. Prevented bone loss and RANKL-induced OCG by improving BV/TV by 2-fold and suppressing porosity and TRAP-positive OCs/BS by 54% and 71%, respectively, at 1 and 5 mg/kg for 7 days. | [159] |
| LPS-induced calvarial osteolysis mice | ||||
| Macrolactins F (44) | Bacillus amyloliquefaciens Deep sea bacterium | RANKL-induced OCG in primary BMMs | Inhibited RANKL-induced osteoclast formation, and inhibited bone resorption activity in BMMs via increased mineralization by 80% at 20 μM for 5 days. Decreased the expression profiles of c-myc, RANK, TRAP, NFATc1, CTSK, MMP-9, CFos, p-JNK, p-p38, and p-Akt, and F-actin ring formation by 90% and promoted pre-osteoblast cell differentiation, and upregulated ALP, NF-kB, RUNX2, osterix, Smad4, BMP-2 COL1A1, OPN, and OCN levels by over 50% at 0–40 μM for 21 days. | [161] |
| MC3T3-E1 cells | ||||
| Vinpocetine | Phyllostachys pubescens | DEX-induced rat osteoblasts apoptosis and ONFH | Suppressed DEX-induced over-regulation of ROS level and inhibited DEX-induced apoptosis in osteoblast by 50% each and protected against ONFH in rats. Increased gene expression of BV/TV, TN, p-Akt, Bcl-xl, and Bcl2 by 40%, 33.3%, 120%, 50%, and 50%, respectively, while suppressing ROS production and JNK, caspase3, and Bax gene expression by over 30% at 10 μM for 24–48 h. | [154] |
| Natural Product | Source | Cell Line/Animal Model | Specific Therapeutic Activity | Ref. |
|---|---|---|---|---|
| Ligustroflavone (46) | Ligustrum lucidum | HEK-293 cells and diabetic mice | Reduced CaSR and serum PTH level by 42.95% while BV/TV, serum Ca, and bone Ca levels were increased by 88.9%, 7.1%, and 41.67%, respectively, at 10−10–10−4 M for 2–24 h. | [174] |
| Leu-Arg-Trp | Pea protein | Pre-osteoblast MC3T3-E1 cells | Increased COL1, ALP, RUNX2-2, and OPG levels, and cell proliferation while inhibiting bone resorption by over 20% each at 50 µM in cell culture for 24 h. | [178] |
| Trp-His | Pea protein | Ca2+-encapsulation fluorescence assay and in silico molecular dynamic simulation | Reduced CaMKII level by 31.67%, and Hills binding coefficient of Ca2+ to CaM from 2.81 to 1.92 (in silico approach). | [176] |
| Crocin (47) | Crocus sativus | Rat cardiomyocytes | Inhibited L-type Ca2+-gated channel while increasing Ca2+-signaling axis by more than 47% each at 1–300 µM for 12 min. | [180] |
| Urolithin C (48) | Crocus sativus | INS-1 beta-cells | Increased intracellular calcium uptake by over 50%. | [179] |
| Sclareol (49) | Salvia sclarea | Oxytocin-induced uterine hypercontraction dysmenorrhea model | Promoted Ca2+–MLCK–MLC20 signaling cascade by 50% at 10–100 µM every 10 min. | [181] |
| γ-Glutamyl valine (γ-EV) | Edible beans | HAoECs | Increased CaSR and β-arrestin by ~40 and 46%, respectively, while inhibiting GIT inflammation by over 30% at 0.01–1 mM for 2 h. | [173] |
| Curcumin (50) and Alpha interacting domain of L-type Ca2+ channel packaged in a poly(glycidyl methacrylate) (PGMA) nanoparticles | Crocus sativus | Rat cardiomyocytes | Decreased L-type Ca2+-gated channel and release of superoxides from 53% to 7.32% for 0.2–20 μM for 20 min. | [182] |
| Natural Products/Sources | Clinical Trial Design | Number of Individuals That Started/Completed the Trial | Parameter Investigated and Outcomes (Bone Formation Marker (BFM) and Bone Resorption Marker (BRM)) | Outcome | Ref. |
|---|---|---|---|---|---|
| Polyphenols/Green tea | 24-week (daily treatment) randomized and placebo-controlled interventional trial | Postmenopausal women with osteopenia (171 started and 150 completed) | (a) Change from baseline (100%) in a ratio of BFM (BSAP) to BRM (TRAP) | (a) 103.6 ± 2.9% | [198] |
| Reconstituted dairy products enriched with Ca, Vit D, Vit K, Vit C, Zn, Mg, Leu, and probiotic (Lactobacillus plantarum) | 24-week (daily treatment) randomized, parallel, double-blind clinical trial with two intervention groups | Both healthy menopausal women with risk of osteoporosis and those with untreated osteopenia (78 started and completed) | (a) Bone mass of the EG vs. CG significantly (p < 0.05) increased (b) BMD was maintained in the EG and depleted in the CG (c) BFM (P1NP) significantly (p < 0.05) increased in the EG compared to the CG (d) BRM (CTx) significantly (p < 0.05) decreased in the EG compared to the CG | (a) 0.01 ± 0.03 (EG) vs. −0.01 ± 0.03 kg (CG) (b) Data not available (c) 13.19 ± 25.17 (EG) vs. −4.21 ± 15.62 (CG) ng/mL (d) −0.05 ± 0.19 (EG) vs. 0.04 ± 0.14 (CG) ng/mL | [199] |
| Hop rho iso-alpha acids (200 mg) and berberine sulphate trihydrate (100 mg) | 14-week, single-blinded, 2-arm placebo-controlled pilot study | Postmenopausal women on Mediterranean low-glycemic diet and limited aerobic exercise (33 started and 32 completed) | (a) BRM (osteocalcin) significantly (p < 0.01) decreased in EG compared to CG (b) Serum 25(OH)D-significantly increased in the ED compared to the control (c) Serum IGF-1 was significantly higher in the EG compared to the control | (a) −31% (EG) vs. +19% (CG) (b) 13% (EG) vs. −25% (CG) (c) Data not shown | [200] |
| Hop rho iso-alpha acids (200 mg) and berberine sulphate trihydrate (100 mg) | 14-week, randomized trial, 2-arm placebo-controlled interventional study | Postmenopausal women on Mediterranean low-glycemic diet and limited aerobic exercise (51 started and 45 completed) | (a) BRM (osteocalcin) significantly (p < 0.01) decreased in EG compared to CG (b) Serum 25(OH)D significantly increased in the ED compared to the control (c) Serum IGF-1 was non-significantly higher in the EG compared to the control | (a) −25% (EG) vs. + 21% (CG) (b) 23% (EG) vs. −12% decrease (c) Data not available | [201] |
| Red clover extract (RCE) rich in Isoflavone aglycones and probiotics | 12-month, double-blind, parallel design, placebo-controlled, randomized controlled trial | Postmenopausal osteopenic women supplemented with calcium (1200 mg/d), magnesium (550 mg/d), and calcitriol (25 μg/d); 87 started and 78 completed | (a) BMD loss was significantly reduced in the EG compared to the CG in the following regions
| (a)
| [202] |
| Collagen peptide from pork skin and bovine bone | 13 weeks, double-blind, placebo-controlled, randomized trial (5 g investigational product or placebo dissolved in 250 mL water or milk administered orally twice daily) | 30 patients (male and female) with knee osteoarthritis (VAS score ≥ 4 and KLG—2 to 4) | (a) Questionnaire assessment of pain, stiffness, and physical function using the WOMAC, VAS, and QOL score. The outcome was reported as the percentage number of patients with
| (a) Significant reduction in WOMAC score at visit 7 (p < 0.05 at visit 4, p < 0.01 at visit 5–7) (b) Significant reduction in VAS score at visit 7 (p < 0.01 at visit 4–7) (c) Significant reduction in QOL score at visit (p < 0.01 at visit 4–7) | [203] |
| Prenylflavonoids from Epimedium spp. | 6 weeks randomized, double-blinded clinical trial prenylflavonoid extract (740 mg daily) or placebo daily for 6 weeks | Healthy postmenopausal women administered prenylflavonoid extract (740 mg daily) or placebo daily (58 started and completed the trial) | (a) Adverse effects on administering prenylflavonoid capsule (b) Metabolites in the sera (c) Cmax and AUC0→∞ for metabolites (d) Increase in BFM (BSAP) in EG compared to the CG (e) Decrease in BRM (TRAF6) in the EG compared to the CG | (a) No adverse symptoms or changes in hepatic, hematological, and renal parameters (b) Desmethylicaritin, icaritin, and icariside II were detected in the sera (c) Cmax and AUC0→∞ for desmethylicaritin of 60.9 nM, and 157.9 nM ×day (d) Significant higher BSAP (p < 0.05) (e) Non-significantly (p = 0.068) lower level of TRAF6 | [204] |
| Collagen peptide hydrolysate (Fortibone®) | 3-month randomized, parallel assignment, interventional clinical trial (administered with 5 mg of collagen peptides) | 51 postmenopausal women within the osteopenic T-score range of (−1.0 > T-score > −2.5) at either the lumbar spine (LS) or femur | (a) Non-significant increase in BMI, and decrease in OC of the EG compared to CG (b) Significant percentage decrease from baseline to 3 months in P1NP and CTX of the EG when compared to the control group | (a) BMI (p = 0.842) and OC (p = 0.578) (b) −13.1 ± 12.3 (EG) vs. −2.1 ± 12.6 (CG) (p = 0.011) for P1NPand −11.4 ± 24 (EG) vs. 3.5 ± 29.9 (CG) for CTx | [205] |
| Kefir-fermented milk peptides | 6 months randomized, parallel, double-blind intervention study | Osteoporosis patients (male and female) of age between 55 and 70 years; 69 patients started and 40 completed the trial | (a) BMD after 6 months forspine, total hip, and hip femoral neck significantly increased in EG compared to CG (b) Serum level of β-CTX and OC | (a) Non-significant difference in spine (p = 0.909); total hip (p = 0.520); and femoral neck (p = 0.501) (b) Non-significant difference in OC and CTX (p = 0.325) | [206] |
| Dried plum (DP) (prune) | Randomized crossover trial (a group consuming ∼42 g DPs per day vs. a second group consuming ∼14 g DPs per day for 2 weeks) | Healthy, postmenopausal women (27 started and completed the trial) | (a) BRM (CTX) decrease in the group that consumed more DPs (∼42 g) after the crossover phase than the group that consumed fewer DPs (∼14 g) | (a) A significant decrease (p = 0.006) | [207] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okagu, I.U.; Ezeorba, T.P.C.; Aguchem, R.N.; Ohanenye, I.C.; Aham, E.C.; Okafor, S.N.; Bollati, C.; Lammi, C. A Review on the Molecular Mechanisms of Action of Natural Products in Preventing Bone Diseases. Int. J. Mol. Sci. 2022, 23, 8468. https://doi.org/10.3390/ijms23158468
Okagu IU, Ezeorba TPC, Aguchem RN, Ohanenye IC, Aham EC, Okafor SN, Bollati C, Lammi C. A Review on the Molecular Mechanisms of Action of Natural Products in Preventing Bone Diseases. International Journal of Molecular Sciences. 2022; 23(15):8468. https://doi.org/10.3390/ijms23158468
Chicago/Turabian StyleOkagu, Innocent U., Timothy P. C. Ezeorba, Rita N. Aguchem, Ikenna C. Ohanenye, Emmanuel C. Aham, Sunday N. Okafor, Carlotta Bollati, and Carmen Lammi. 2022. "A Review on the Molecular Mechanisms of Action of Natural Products in Preventing Bone Diseases" International Journal of Molecular Sciences 23, no. 15: 8468. https://doi.org/10.3390/ijms23158468