
Extensive Sampling of Molecular Dynamics Simulations to Identify Reliable Protein Structures for Optimized Virtual Screening Studies: The Case of the hTRPM8 Channel

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Docking Simulations
2.2. VS Campaigns by Using the Non-Optimized Frames
2.3. VS Campaigns by Using the SCRWL4 Optimized Frames
2.4. Comparison of the Two Sets of Simulations
2.5. Multiple Frames Consensus Strategy
2.6. Structural Analysis of the Sampled Frames
3. Discussion
4. Methods and Materials
4.1. Frame Selection and Optimization
4.2. Virtual Screening by Using LiGen
4.3. Rescoring and Consensus Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| APBS | Adaptive Poisson–Boltzmann Solver |
| CS | Chemical Score |
| CSopt | Optimized Chemical Score |
| EF | Enrichment Factor |
| EFO | Enrichment Factor Optimization |
| MCC | Matthews correlation coefficient |
| MD | Molecular Dynamics |
| MLPInS | Molecular Lipophilicity Potential Interaction Score |
| PLP | Piecewise Linear Potential |
| PS | Pacman Score |
| TRPM8 | Transient receptor potential cation channel subfamily M (melastatin) 8 |
| VS | Virtual Screening |
References
- Maia, E.H.B.; Assis, L.C.; de Oliveira, T.A.; da Silva, A.M.; Taranto, A.G. Structure-Based Virtual Screening: From Classical to Artificial Intelligence. Front. Chem. 2020, 8, 343. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, A. Latest trends in structure based drug design with protein targets. Adv. Protein Chem. Struct. Biol. 2020, 121, 1–23. [Google Scholar] [PubMed]
- Sulimov, V.B.; Kutov, D.C.; Sulimov, A.V. Advances in Docking. Curr. Med. Chem. 2019, 26, 7555–7580. [Google Scholar] [CrossRef] [PubMed]
- Pozharski, E.; Deller, M.C.; Rupp, B. Validation of Protein-Ligand Crystal Structure Models: Small Molecule and Peptide Ligands. Methods Mol. Biol. 2017, 1607, 611–625. [Google Scholar]
- Deller, M.C.; Rupp, B. Models of protein-ligand crystal structures: Trust, but verify. J. Comput. Aided Mol. Des. 2015, 29, 817–836. [Google Scholar] [CrossRef]
- McGovern, S.L.; Shoichet, B.K. Information decay in molecular docking screens against holo, apo and modeled conformations of enzymes. J. Med. Chem. 2003, 46, 2895–2907. [Google Scholar] [CrossRef]
- Seeliger, D.; de Groot, B.L. Conformational transitions upon ligand binding: Holo-structure prediction from apo conformations. PLoS Comput. Biol. 2010, 6, e1000634. [Google Scholar] [CrossRef]
- Salmaso, V.; Moro, S. Bridging Molecular Docking to Molecular Dynamics in Exploring Ligand-Protein Recognition Process: An Overview. Front. Pharmacol. 2018, 9, 923. [Google Scholar] [CrossRef] [Green Version]
- Amaro, R.E.; Baudry, J.; Chodera, J.; Demir, Ö.; McCammon, J.A.; Miao, Y.; Smith, J.C. Ensemble Docking in Drug Discovery. Biophys. J. 2018, 114, 2271–2278. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Lill, M.A. Utilizing Experimental Data for Reducing Ensemble Size in Flexible-Protein Docking. J. Chem. Inf. Model. 2012, 52, 187–198. [Google Scholar] [CrossRef]
- Evangelista Falcon, W.; Ellingson, S.R.; Smith, J.C.; Baudry, J. Ensemble Docking in Drug Discovery: How Many Protein Configurations from Molecular Dynamics Simulations are Needed To Reproduce Known Ligand Binding? J. Phys. Chem. B 2019, 123, 5189–5195. [Google Scholar] [CrossRef] [PubMed]
- Amadei, A.; Linssen, A.B.; Berendsen, H.J. Essential dynamics of proteins. Proteins 1993, 17, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Talarico, C.; Gervasoni, S.; Manelfi, C.; Pedretti, A.; Vistoli, G.; Beccari, A.R. Combining Molecular Dynamics and Docking Simulations to Develop Targeted Protocols for Performing Optimized Virtual Screening Campaigns on The hTRPM8 Channel. Int. J. Mol. Sci. 2020, 21, 2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Muñiz, R.; Bonache, M.A.; Martín-Escura, C.; Gómez-Monterrey, I. Recent Progress in TRPM8 Modulation: An Update. Int. J. Mol. Sci. 2019, 28, 2618. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Lee, S.Y. Current View of Ligand and Lipid Recognition by the Menthol Receptor TRPM8. Trends Biochem. Sci. 2020, 45, 806–819. [Google Scholar] [CrossRef]
- Yin, Y.; Wu, M.; Zubcevic, L.; Borschel, W.F.; Lander, G.C.; Lee, S.Y. Structure of the cold- and menthol-sensing ion channel TRPM8. Science 2018, 359, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Beccari, A.R.; Cavazzoni, C.; Beato, C.; Costantino, G. LiGen: A high performance workflow for chemistry driven de novo design. J. Chem. Inf. Model. 2013, 53, 1518–1527. [Google Scholar] [CrossRef]
- Krivov, G.G.; Shapovalov, M.V.; Dunbrack, R.L., Jr. Improved prediction of protein side-chain conformations with SCWRL4. Proteins 2009, 77, 778–795. [Google Scholar] [CrossRef] [Green Version]
- Vistoli, G.; Mazzolari, A.; Testa, B.; Pedretti, A. Binding Space Concept: A New Approach To Enhance the Reliability of Docking Scores and Its Application to Predicting Butyrylcholinesterase Hydrolytic Activity. J. Chem. Inf. Model. 2017, 57, 1691–1702. [Google Scholar] [CrossRef]
- Mazzolari, A.; Vistoli, G.; Testa, B.; Pedretti, A. Prediction of the Formation of Reactive Metabolites by A Novel Classifier Approach Based on Enrichment Factor Optimization (EFO) as Implemented in the VEGA Program. Molecules 2018, 23, 2955. [Google Scholar] [CrossRef] [Green Version]
- Pedretti, A.; Mazzolari, A.; Gervasoni, S.; Vistoli, G. Rescoring and Linearly Combining: A Highly Effective Consensus Strategy for Virtual Screening Campaigns. Int. J. Mol. Sci. 2019, 20, 2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manelfi, C.; Gossen, J.; Gervasoni, S.; Talarico, C.; Albani, S.; Philipp, B.J.; Musiani, F.; Vistoli, G.; Rossetti, G.; Beccari, A.R.; et al. Combining Different Docking Engines and Consensus Strategies to Design and Validate Optimized Virtual Screening Protocols for the SARS-CoV-2 3CL Protease. Molecules 2021, 26, 797. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.R.; Sørensen, J.; Hensley, N.; Wong, C.; Zhu, C.; Perison, T.; Amaro, R.E. POVME 3.0: Software for Mapping Binding Pocket Flexibility. J. Chem. Theory Comput. 2017, 13, 4584–4592. [Google Scholar] [CrossRef] [PubMed]
- Beccari, A.R.; Gemei, M.; Lo Monte, M.; Menegatti, N.; Fanton, M.; Pedretti, A.; Bovolenta, S.; Nucci, C.; Molteni, A.; Rossignoli, A.; et al. Novel selective, potent naphthyl TRPM8 antagonists identified through a combined ligand- and structure-based virtual screening approach. Sci. Rep. 2017, 7, 10999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Le, S.C.; Hsu, A.L.; Borgnia, M.J.; Yang, H.; Lee, S.Y. Structural basis of cooling agent and lipid sensing by the cold-activated TRPM8 channel. Science 2019, 363, 6430. [Google Scholar] [CrossRef]
- Pedretti, A.; Mazzolari, A.; Gervasoni, S.; Fumagalli, L.; Vistoli, G. The VEGA suite of programs: An versatile platform for cheminformatics and drug design projects. Bioinformatics 2021, 37, 1174–1175. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Time (ns) | EF1% | Consensus Equation |
|---|---|---|
| 25 | nd | The frame was fully discarded |
| 50 | nd | The frame was fully discarded |
| 75 | nd | The frame was fully discarded |
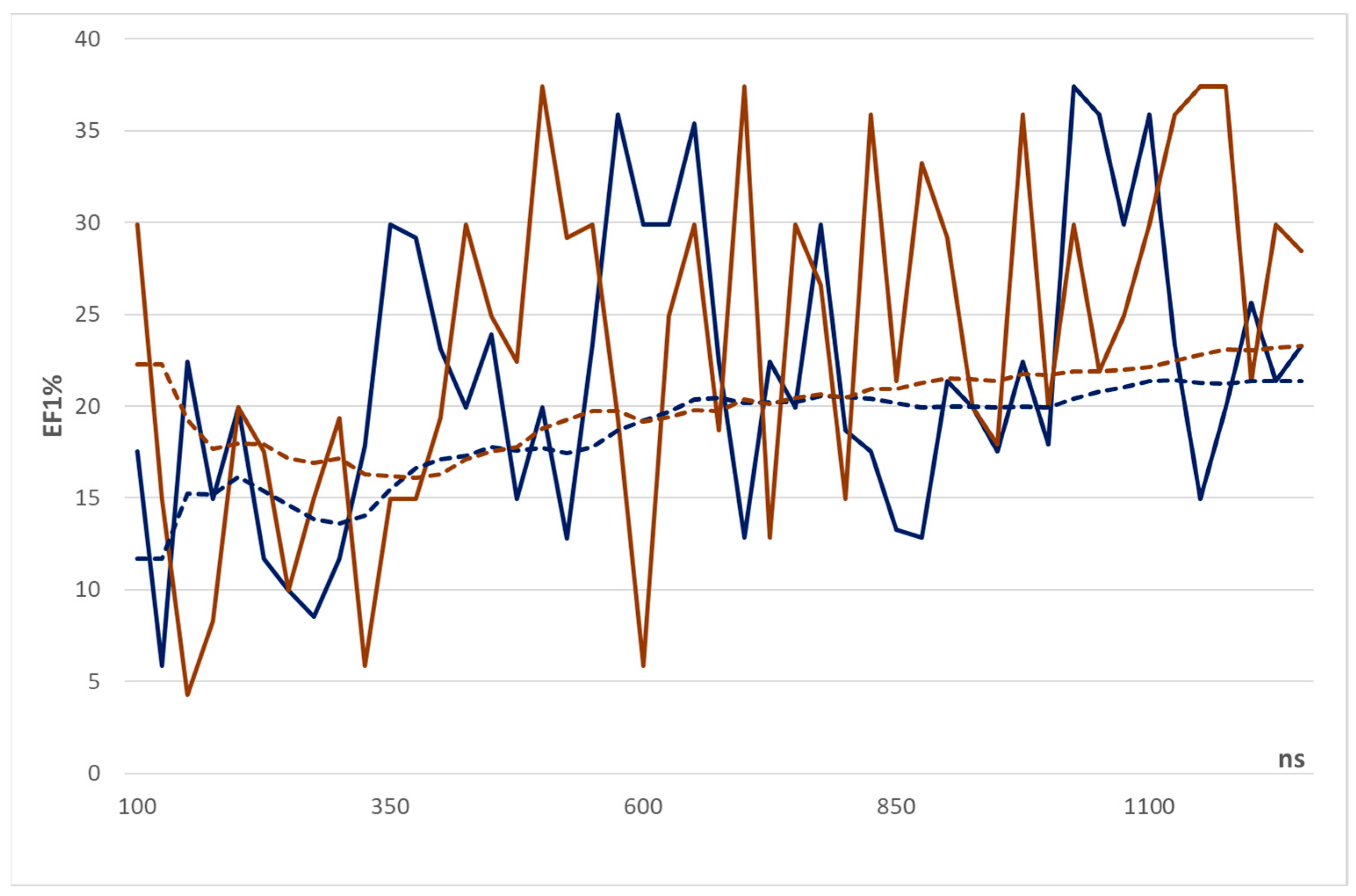
| 100 | 17.51 | 1.00 PSmeanB |
| 125 | 5.84 | 1.00 CSbestC |
| 150 | 22.42 | −1.00 CSbestC − 0.36 CsoptbestB |
| 175 | 14.95 | 1.00 CsoptmeanC + 2.96 CSbestB |
| 200 | 19.93 | 1.00 CSmeanB − 1.10 CSbestB − 1.38 CsoptmeanA |
| 225 | 11.67 | 1.00 PSmeanA |
| 250 | 9.96 | −1.00 CSmeanC − 2.44 CSmeanB |
| 275 | 8.54 | −1.00 CSmeanC + 0.46 CSbestB |
| 300 | 11.67 | 1.00 CSbestC |
| 325 | 17.81 | 1.00 CSbestD − 1.11 CSbestC |
| 350 | 29.90 | −1.00 CsoptbestB + 2.15 CsoptbestA |
| 375 | 29.18 | 1.00 CSbestD |
| 400 | 23.12 | −1.00 CSmeanD − 8.80 CsoptbestC |
| 425 | 19.93 | 1.00 CsoptmeanB − 3.53 CSmeanA |
| 450 | 23.92 | −1.00 CSmeanD − 0.41 CsoptbestB − 0.11 CSmeanA |
| 475 | 14.95 | −1.00 CSmeanD − 9.86 CsoptbestC + 3.87 CsoptbestA |
| 500 | 19.93 | 1.00 CSmeanD − 12.42 CsoptbestA |
| 525 | 12.81 | 1.00 PSmeanD − 71.23 CSmeanD − 58.51 CsoptbestC |
| 550 | 23.35 | 1.00 CSbestA |
| 575 | 35.88 | −1.00 CsoptmeanC + 1.30 CSbestA |
| 600 | 29.90 | −1.00 CsoptbestC − 0.39 CSmeanA |
| 625 | 29.90 | −1.00 CsoptmeanD − 0.082 CSmeanB + 1.26 CsoptmeanA |
| 650 | 35.88 | −1.00 CSbestD − 1.71 CSmeanC + 1.16 CsoptbestB |
| 675 | 22.42 | −1.00 CsoptmeanC + 0.24 CSbestA |
| 700 | 12.82 | −1.00 CSbestB + 0.82 CSmeanA |
| 725 | 22.42 | −1.00 CSmeanB + 0.36 CSbestB |
| 750 | 19.93 | −1.00 CSmeanC − 6.04 CsoptmeanB − 1.51 CsoptmeanA |
| 775 | 29.90 | −1.00 CSmeanC + 0.67 CsoptbestB |
| 800 | 18.69 | −1.00 CsoptmeanB + 0.24 CSbestA |
| 825 | 17.51 | 1.00 CSBestA |
| 850 | 13.29 | 1.00 CSbestD + 44.18 CsoptbestC |
| 875 | 12.82 | −1.00 CSoptmeanB + 7.20 CSmeanB − 29.71 CSbestA |
| 900 | 21.36 | −1.00 CSbestC + 0.95 CSbestA |
| 925 | 19.93 | 1.00 CsoptmeanD + 4.31 CSmeanC + 0.90 CSbestC |
| 950 | 17.51 | 1.00 CSbestC |
| 975 | 22.42 | −1.00 CSmeanD + 0.94 CSbestC |
| 1000 | 17.94 | −1.00 CsoptmeanC + 0.42 CSmeanB − 0.15 CSmeanA |
| 1025 | 37.38 | 1.00 CSoptmeanD − 1.27 CsoptbestC |
| 1050 | 35.88 | 1.00 CsoptmeanD + 1.69 CSmeanC − 9.10 CsoptmeanB |
| 1075 | 29.90 | 1.00 CSbestC − 1.86 CsoptmeanB |
| 1100 | 35.88 | −1.00 CSmeanB − 4.56 CsoptbestB − 3.70 CSmeanA |
| 1125 | 23.35 | 1.00 CSbestA |
| 1150 | 14.95 | −1.00 CsoptmeanD − 0.67 CsoptmeanC + 0.34 CSbestB |
| 1175 | 19.93 | −1.00 CSbestB + 0.32 CsoptmeanA |
| 1200 | 25.63 | −1.00 CsoptmeanC − 1.85 CsoptbestB + 0.95 CsoptmeanA |
| 1225 | 21.35 | −1.00 CsoptmeanD + 0.27 CSbestC |
| 1250 | 23.26 | 1.00 CsoptmeanC − 10.07 CsoptmeanB + 1.90 CsoptmeanA |
| Score | Monomer A | Monomer B | Monomer C | Monomer D | Total |
|---|---|---|---|---|---|
| Best | 11 | 14 | 16 | 4 | 45 |
| Mean | 13 | 15 | 14 | 13 | 55 |
| Total | 24 | 29 | 30 | 17 | 100 |
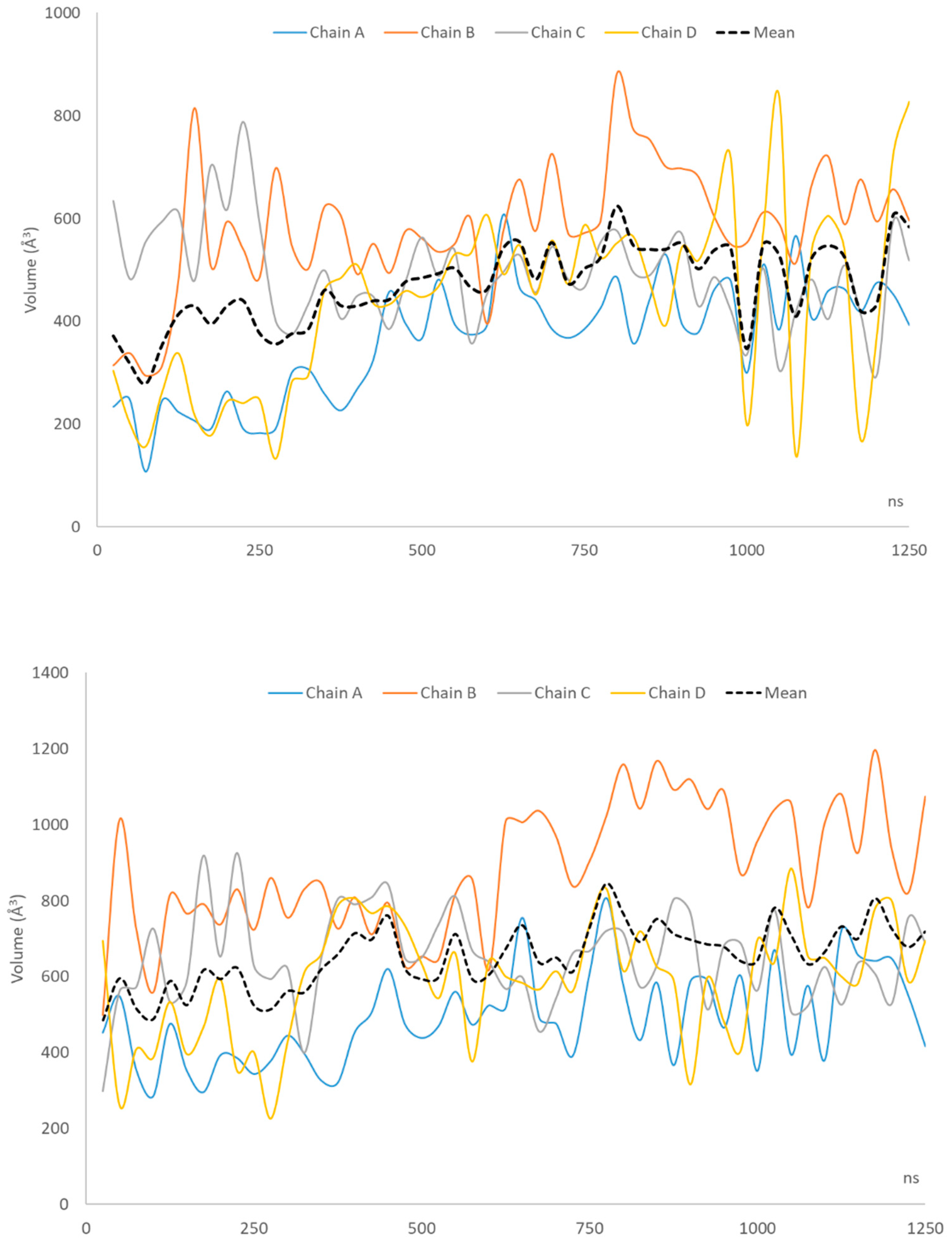
| Mean volume | 365.4 | 582.2 | 492.7 | 439.8 | 470.0 |
| Frame | EF1% | Consensus Equation |
|---|---|---|
| 25 | 29.18 | 1.00 PSmeanC |
| 50 | 11.67 | 1.00 CSbestB |
| 75 | 25.63 | −1.00 CsoptbestB − 1.27 CsoptmeanA |
| 100 | 29.90 | −1.00 CSbestC + 1.22 CSmeanB − 0.21 CSbestA |
| 125 | 14.95 | −1.00 CsoptmeanD + 0.89 CSmeanD |
| 150 | 4.27 | 1.00 CsoptbestC − 1.42 CsoptmeanB |
| 175 | 8.31 | −1.00 CSmeanD + 0.55 CSmeanC + 0.27 CSmeanB |
| 200 | 19.93 | −1.00 CsoptmeanD − 5.44 CSbestA |
| 225 | 17.51 | 1.00 CSbestA |
| 250 | 9.96 | −1.00 CSmeanC − 2.44 CSmeanB |
| 275 | nd | The frame was fully discarded |
| 300 | 19.33 | 1.00 CSmeanD + 2.91 CSbestC − 7.81 CsoptbestB |
| 325 | 5.84 | 1.00 CSmeanD |
| 350 | 14.95 | −1.00 CSbestC − 0.70 CsoptbestB |
| 375 | 14.95 | −1.00 CsoptbestC − 1.14 PSmeanA |
| 400 | 19.33 | −1.00 CSmeanC − 4.23 PSmeanB + 18.23 PSbestB |
| 425 | 29.90 | −1.00 CsoptmeanC − 10.07 CSmeanB + 1.95 CsoptbestA |
| 450 | 24.92 | 1.00 CsoptmeanD − 3.48 CSmeanA |
| 475 | 22.42 | −1.00 CSmeanD − 21.50 CSmeanC |
| 500 | 37.38 | −1.00 CsoptmeanC + 1.73 CSmeanA |
| 525 | 29.18 | 1.00 PSmeanD |
| 550 | 29.90 | −1.00 CsoptmeanC − 8.69 CsoptbestC + 0.27 CSmeanB |
| 575 | nd | The frame was fully discarded |
| 600 | 5.84 | 1.00 CbestA |
| 625 | 24.92 | 1.00 CSbestD − 14.04 CsoptmeanA |
| 650 | 29.90 | 1.00 CsoptbestB − 1.66 CSmeanA − 0.94 CsoptbestA |
| 675 | 18.69 | 1.00 CsoptbestC − 1.09 CsoptmeanB |
| 700 | 37.38 | 1.00 CsoptmeanC − 4.44 CsoptmeanB |
| 725 | 12.82 | −1.00 CSmeanD − 1.38 CsoptbestB |
| 750 | 29.90 | 1.00 CSmeanD − 0.12 CsoptbestD − 1.14 CsoptmeanA |
| 775 | 26.58 | 1.00 CsoptmeanD − 1.25 CSoptbestC |
| 800 | 14.95 | 1.00 CsoptmeanD − 2.89 CsoptmeanA − 9.20 CSbestA |
| 825 | 35.88 | 1.00 CsoptmeanD − 1.05 CsoptmeanB |
| 850 | 21.35 | 1.00 CsoptmeanC − 3.50 CsoptmeanB |
| 875 | 33.22 | −1.00 CsoptbestD − 0.49 CsoptmeanA |
| 900 | 29.18 | 1.00 CSbestA |
| 925 | 19.93 | −1.00 CSbestD − 1.34 CSmeanC − 2.32 CSmeanB |
| 950 | 17.94 | 1.00 CsoptbestC − 12.25 CSbestA |
| 975 | 35.88 | −1.00 CsoptmeanD − 0.68 CsoptmeanB |
| 1000 | 19.93 | −1.00 CsoptmeanC − 0.48 CSmeanB − 4.85 CsoptbestB |
| 1025 | 29.90 | −1.00 CsoptmeanB + 0.060 CsoptbestA |
| 1050 | Nd | The frame was fully discarded |
| 1075 | 24.92 | −1.00 CsoptbestD − 3.30 CSmeanB |
| 1100 | 29.90 | −1.00 CSmeanC − 0.59 CsoptmeanA |
| 1125 | 35.88 | −1.00 CSmeanC − 4.84 CSbestB |
| 1150 | 19.93 | 1.00 CSmeanD − 15.28 CSmeanB |
| 1175 | 37.38 | 1.00 CsoptbestB − 1.66 CsoptmeanA |
| 1200 | 21.35 | 1.00 CsoptmeanD − 1.73 CsoptmeanB |
| 1225 | 29.90 | −1.00 CsoptmeanC + 0.50 CsoptbestB |
| 1250 | 28.47 | 1.00 CsoptmeanD − 0.21 CsoptbestC − 0.98 CSmeanB |
| Score | Monomer A | Monomer B | Monomer C | Monomer D | Total |
|---|---|---|---|---|---|
| Best | 10 | 11 | 10 | 5 | 36 |
| Mean | 11 | 19 | 15 | 18 | 63 |
| Total | 21 | 30 | 25 | 23 | 99 |
| Mean volume | 491.2 | 884.8 | 648.2 | 591.0 | 653.8 |
| Time (ns) | Non-Optimized Frames | Optimized Frames | ||
|---|---|---|---|---|
| EF1% | Volume Average | EF1% | Volume Average | |
| 25–250 | 14.61 | 380.6 | 17.13 | 556.1 |
| 275–500 | 19.90 | 428.1 | 20.40 | 630.4 |
| 525–750 | 24.48 | 503.1 | 23.79 | 655.2 |
| 775–1000 | 19.14 | 525.8 | 25.48 | 711.8 |
| 1025–1250 | 26.75 | 512.52 | 29.70 | 715.5 |
| Averages | 21.38 | 470.0 | 23.29 | 653.8 |
| EF1% | All Monomers | Monomer A | Monomer B | Monomer C | Monomer D | Best Scores | Mean Scores |
|---|---|---|---|---|---|---|---|
| Non-optimized | 47.84 (37.07) | 29.90 (24.17) | 41.86 (29.00) | 44.85 (36.36) | 44.85 (30.65) | 35.88 (13.85) | 44.84 (26.54) |
| Optimized | 49.83 (38.62) | 44.85 (36.13) | 29.90 (26.48) | 41.86 (26.61) | 44.85 (36.63) | 35.88 (25.12) | 38.94 (29.90) |
| Metric | Best Model from Previous Study [13] | Multiple Non-Optimized Frames | Multiple Optimized Frames |
|---|---|---|---|
| EF1% | 67.11 | 47.84 | 49.83 |
| % active in top 1% | 67.11% | 80% | 90% |
| MCC | 0.66 | 0.49 | 0.64 |
| Sensitivity | 0.66 | 0.50 | 0.65 |
| Accuracy | 0.99 | 0.99 | 0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gervasoni, S.; Talarico, C.; Manelfi, C.; Pedretti, A.; Vistoli, G.; Beccari, A.R. Extensive Sampling of Molecular Dynamics Simulations to Identify Reliable Protein Structures for Optimized Virtual Screening Studies: The Case of the hTRPM8 Channel. Int. J. Mol. Sci. 2022, 23, 7558. https://doi.org/10.3390/ijms23147558
Gervasoni S, Talarico C, Manelfi C, Pedretti A, Vistoli G, Beccari AR. Extensive Sampling of Molecular Dynamics Simulations to Identify Reliable Protein Structures for Optimized Virtual Screening Studies: The Case of the hTRPM8 Channel. International Journal of Molecular Sciences. 2022; 23(14):7558. https://doi.org/10.3390/ijms23147558
Chicago/Turabian StyleGervasoni, Silvia, Carmine Talarico, Candida Manelfi, Alessandro Pedretti, Giulio Vistoli, and Andrea R. Beccari. 2022. "Extensive Sampling of Molecular Dynamics Simulations to Identify Reliable Protein Structures for Optimized Virtual Screening Studies: The Case of the hTRPM8 Channel" International Journal of Molecular Sciences 23, no. 14: 7558. https://doi.org/10.3390/ijms23147558