
Expression and Prognostic Significance of CD47–SIRPA Macrophage Checkpoint Molecules in Colorectal Cancer

 , ,
, ,
Abstract
:1. Introduction
2. Results
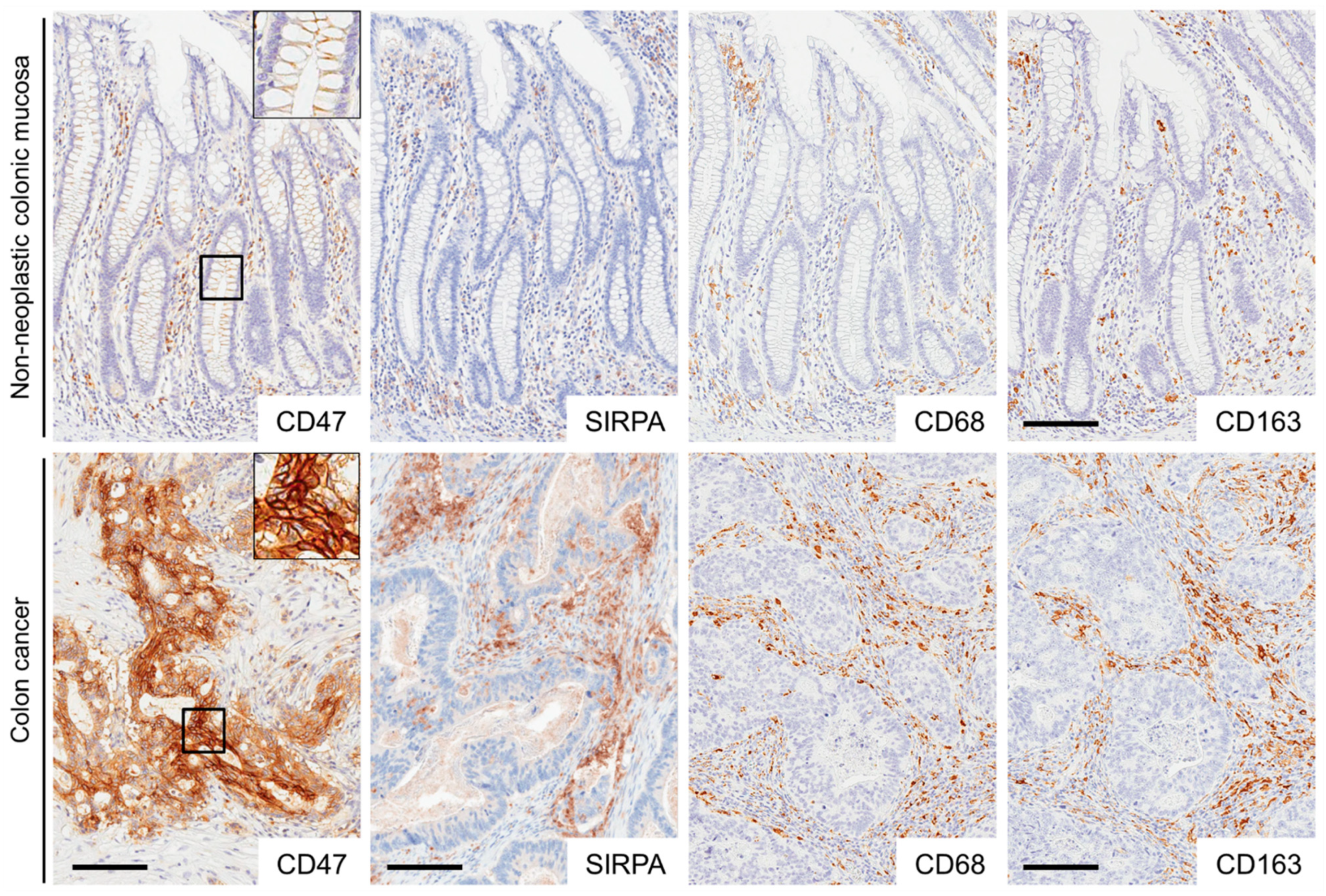
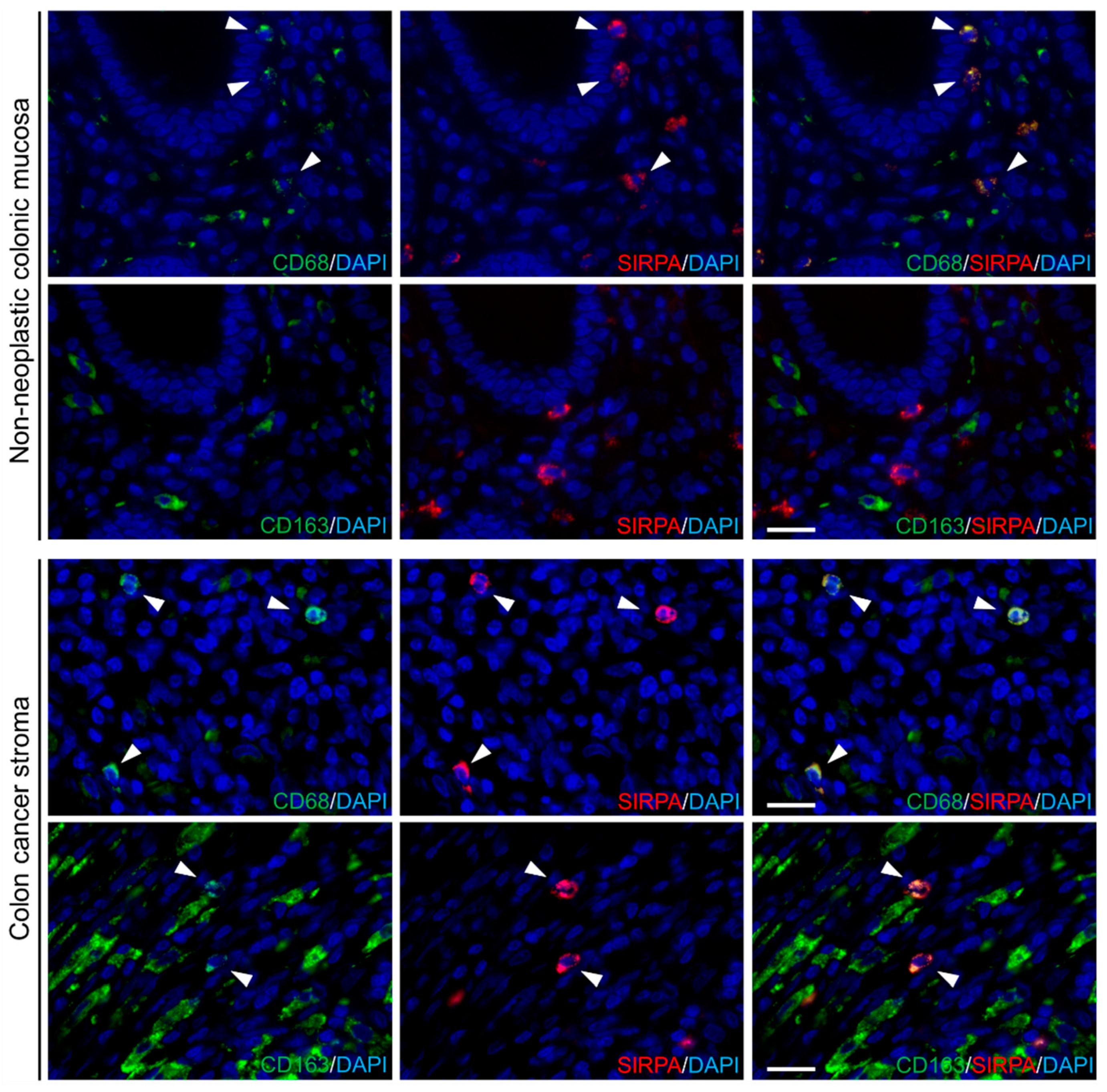
2.1. Expression of CD47, SIRPA, CD68, and CD163 in Non-Neoplastic Colonic Mucosae and CRCs
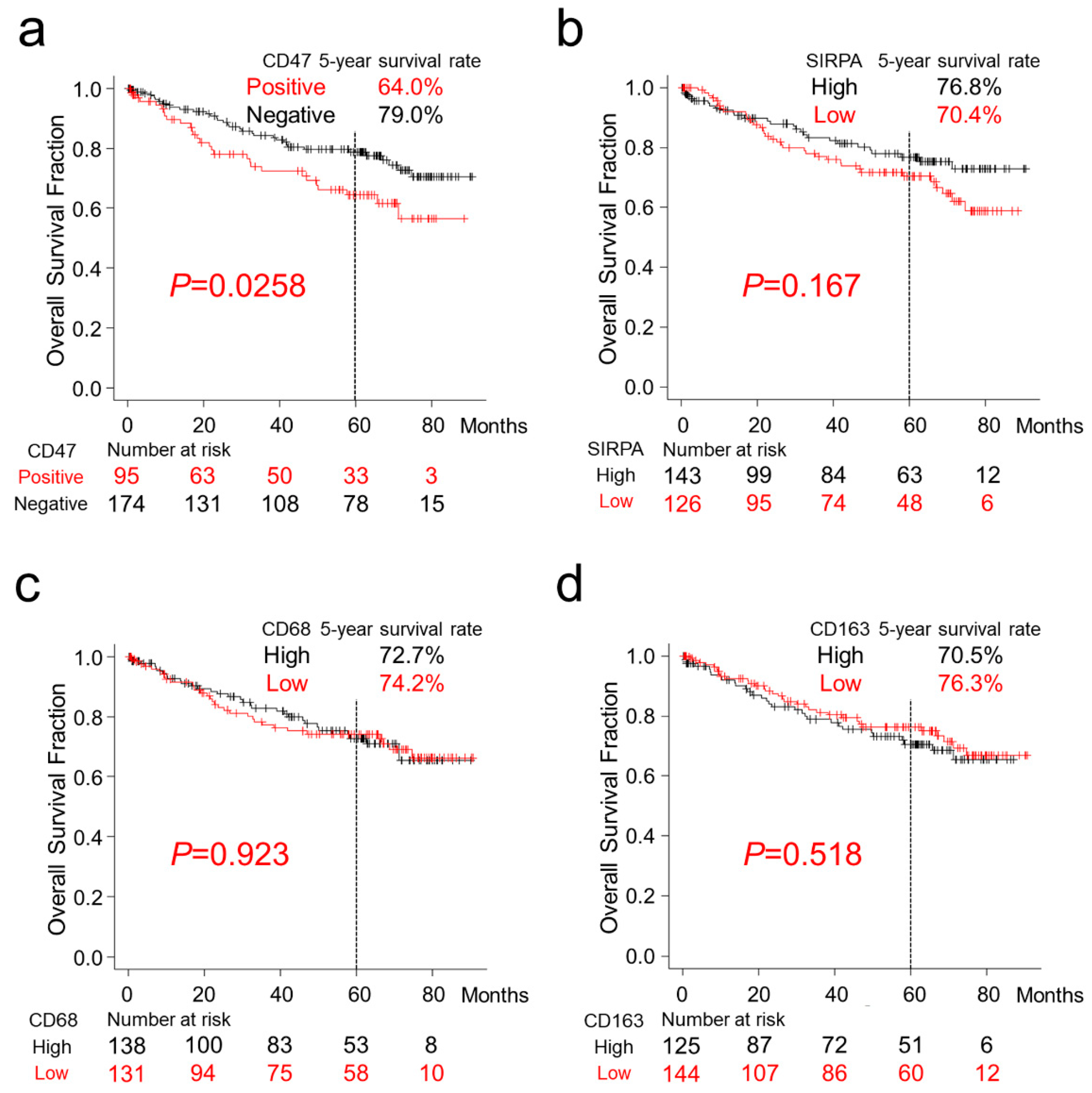
2.2. Survival Analyses of Patients with CRC
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. Immunohistochemistry
4.3. Fluorescent Immunohistochemistry
4.4. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CRC | colorectal cancer |
| SIRPA | signal regulatory protein-alpha |
| HR | hazard ratio |
| CI | confidence interval |
| EMT | epithelial–mesenchymal transition |
References
- Callahan, M.K.; Postow, M.A.; Wolchok, J.D. CTLA-4 and PD-1 Pathway Blockade: Combinations in the Clinic. Front. Oncol. 2014, 4, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luke, J.J.; Flaherty, K.T.; Ribas, A.; Long, G.V. Targeted agents and immunotherapies: Optimizing outcomes in melanoma. Nat. Rev. Clin. Oncol. 2017, 14, 463–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, D.T.; Uram, J.N.; Wang, H.; Bartlett, B.R.; Kemberling, H.; Eyring, A.D.; Skora, A.D.; Luber, B.S.; Azad, N.S.; Laheru, D.; et al. PD-1 Blockade in Tumors with Mismatch-Repair Deficiency. N. Engl. J. Med. 2015, 372, 2509–2520. [Google Scholar] [CrossRef] [Green Version]
- Isidro, R.A.; Appleyard, C.B. Colonic macrophage polarization in homeostasis, inflammation, and cancer. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G59–G73. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, D.; Iida, T.; Nakase, H. The Phagocytic Function of Macrophage-Enforcing Innate Immunity and Tissue Homeostasis. Int. J. Mol. Sci. 2017, 19, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhold, M.I.; Lindberg, F.P.; Plas, D.; Reynolds, S.; Peters, M.G.; Brown, E.J. In vivo expression of alternatively spliced forms of integrin-associated protein (CD47). J. Cell Sci. 1995, 108 (Pt 11), 3419–3425. [Google Scholar]
- Brown, E.; Hooper, L.; Ho, T.; Gresham, H. Integrin-associated protein: A 50-kD plasma membrane antigen physically and functionally associated with integrins. J. Cell. Biol. 1990, 111 (6 Pt 1), 2785–2794. [Google Scholar] [CrossRef]
- Gao, A.G.; Lindberg, F.P.; Finn, M.B.; Blystone, S.D.; Brown, E.J.; Frazier, W.A. Integrin-associated protein is a receptor for the C-terminal domain of thrombospondin. J. Biol. Chem. 1996, 271, 21–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindberg, F.P.; Bullard, D.C.; Caver, T.E.; Gresham, H.D.; Beaudet, A.L.; Brown, E.J. Decreased resistance to bacterial infection and granulocyte defects in IAP-deficient mice. Science 1996, 274, 795–798. [Google Scholar] [CrossRef]
- Liu, Y.; Merlin, D.; Burst, S.L.; Pochet, M.; Madara, J.L.; Parkos, C.A. The role of CD47 in neutrophil transmigration. Increased rate of migration correlates with increased cell surface expression of CD47. J. Biol. Chem. 2001, 276, 40156–40166. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, M.; Ohnishi, H.; Okazawa, H.; Tomonaga, H.; Hayashi, A.; Fujimoto, T.T.; Furuya, N.; Matozaki, T. Promotion of neurite and filopodium formation by CD47: Roles of integrins, Rac, and Cdc42. Mol. Biol. Cell 2004, 15, 3950–3963. [Google Scholar] [CrossRef]
- Reinhold, M.I.; Lindberg, F.P.; Kersh, G.J.; Allen, P.M.; Brown, E.J. Costimulation of T cell activation by integrin-associated protein (CD47) is an adhesion-dependent, CD28-independent signaling pathway. J. Exp. Med. 1997, 185, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, M.P.; Alizadeh, A.A.; Tang, C.; Myklebust, J.H.; Varghese, B.; Gill, S.; Jan, M.; Cha, A.C.; Chan, C.K.; Tan, B.T.; et al. Anti-CD47 antibody synergizes with rituximab to promote phagocytosis and eradicate non-Hodgkin lymphoma. Cell 2010, 142, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Willingham, S.B.; Volkmer, J.P.; Gentles, A.J.; Sahoo, D.; Dalerba, P.; Mitra, S.S.; Wang, J.; Contreras-Trujillo, H.; Martin, R.; Cohen, J.D.; et al. The CD47-signal regulatory protein alpha (SIRPa) interaction is a therapeutic target for human solid tumors. Proc. Natl. Acad. Sci. USA 2012, 109, 6662–6667. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, Y.; Matozaki, T.; Noguchi, T.; Iwamatsu, A.; Yamao, T.; Takahashi, N.; Tsuda, M.; Takada, T.; Kasuga, M. A novel membrane glycoprotein, SHPS-1, that binds the SH2-domain-containing protein tyrosine phosphatase SHP-2 in response to mitogens and cell adhesion. Mol. Cell. Biol. 1996, 16, 6887–6899. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, S.; Jamieson, C.H.; Pang, W.W.; Park, C.Y.; Chao, M.P.; Majeti, R.; Traver, D.; van Rooijen, N.; Weissman, I.L. CD47 is upregulated on circulating hematopoietic stem cells and leukemia cells to avoid phagocytosis. Cell 2009, 138, 271–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majeti, R.; Chao, M.P.; Alizadeh, A.A.; Pang, W.W.; Jaiswal, S.; Gibbs, K.D., Jr.; van Rooijen, N.; Weissman, I.L. Weissman. CD47 is an adverse prognostic factor and therapeutic antibody target on human acute myeloid leukemia stem cells. Cell 2009, 138, 286–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Advani, R.; Flinn, I.; Popplewell, L.; Forero, A.; Bartlett, N.L.; Ghosh, N.; Kline, J.; Roschewski, M.; LaCasce, A.; Collins, G.P.; et al. CD47 Blockade by Hu5F9-G4 and Rituximab in Non-Hodgkin’s Lymphoma. N. Engl. J. Med. 2018, 379, 1711–1721. [Google Scholar] [CrossRef]
- Johnson, L.D.S.; Banerjee, S.; Kruglov, O.; Viller, N.N.; Horwitz, S.M.; Lesokhin, A.; Zain, J.; Querfeld, C.; Chen, R.; Okada, C.; et al. Targeting CD47 in Sezary syndrome with SIRPalphaFc. Blood Adv. 2019, 3, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Sikic, B.I.; Lakhani, N.; Patnaik, A.; Shah, S.A.; Chandana, S.R.; Rasco, D.; Colevas, A.D.; O’Rourke, T.; Narayanan, S.; Papadopoulos, K.; et al. First-in-Human, First-in-Class Phase I Trial of the Anti-CD47 Antibody Hu5F9-G4 in Patients with Advanced Cancers. J. Clin. Oncol. 2019, 37, 946–953. [Google Scholar] [CrossRef]
- Wang, C.L.; Lin, M.J.; Hsu, C.Y.; Lin, H.Y.; Tsai, H.P.; Long, C.Y.; Tsai, E.M.; Hsieh, T.H.; Wu, C.H. CD47 promotes cell growth and motility in epithelial ovarian cancer. Biomed. Pharmacother. 2019, 119, 109105. [Google Scholar] [CrossRef] [PubMed]
- Pai, S.; Bamodu, O.A.; Lin, Y.K.; Lin, C.S.; Chu, P.Y.; Chien, M.H.; Wang, L.S.; Hsiao, M.; Yeh, C.T.; Tsai, J.T. CD47-SIRPalpha Signaling Induces Epithelial-Mesenchymal Transition and Cancer Stemness and Links to a Poor Prognosis in Patients with Oral Squamous Cell Carcinoma. Cells 2019, 8, 1658. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.P.; Kim, H.J.; Wu, H.; Price-Troska, T.; Villasboas, J.C.; Jalali, S.; Feldman, A.L.; Novak, A.J.; Yang, Z.Z.; Ansell, S.M. SIRPalpha expression delineates subsets of intratumoral monocyte/macrophages with different functional and prognostic impact in follicular lymphoma. Blood Cancer J. 2019, 9, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, J.; Chen, F.; Liu, D.; Gu, F.; Chen, Z.; Wang, Y. Prognostic value of immune checkpoint molecules in breast cancer. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Kazama, R.; Miyoshi, H.; Takeuchi, M.; Miyawaki, K.; Nakashima, K.; Yoshida, N.; Kawamoto, K.; Yanagida, E.; Yamada, K.; Umeno, T.; et al. Combination of CD47 and signal-regulatory protein-alpha constituting the “don’t eat me signal” is a prognostic factor in diffuse large B-cell lymphoma. Cancer Sci. 2020, 111, 2608–2619. [Google Scholar] [CrossRef] [PubMed]
- Hamada, I.; Kato, M.; Yamasaki, T.; Iwabuchi, K.; Watanabe, T.; Yamada, T.; Itoyama, S.; Ito, H.; Okada, K. Clinical effects of tumor-associated macrophages and dendritic cells on renal cell carcinoma. Anticancer Res. 2002, 22, 4281–4284. [Google Scholar] [PubMed]
- Hanada, T.; Nakagawa, M.; Emoto, A.; Nomura, T.; Nasu, N.; Nomura, Y. Prognostic value of tumor-associated macrophage count in human bladder cancer. Int. J. Urol. 2000, 7, 263–269. [Google Scholar] [CrossRef]
- Jensen, T.O.; Schmidt, H.; Moller, H.J.; Hoyer, M.; Maniecki, M.B.; Sjoegren, P.; Christensen, I.J.; Steiniche, T. Macrophage markers in serum and tumor have prognostic impact in American Joint Committee on Cancer stage I/II melanoma. J. Clin. Oncol. 2009, 27, 3330–3337. [Google Scholar] [CrossRef]
- Leek, R.D.; Lewis, C.E.; Whitehouse, R.; Greenall, M.; Clarke, J.; Harris, A.L. Association of macrophage infiltration with angiogenesis and prognosis in invasive breast carcinoma. Cancer Res. 1996, 56, 4625–4629. [Google Scholar]
- Tsutsui, S.; Yasuda, K.; Suzuki, K.; Tahara, K.; Higashi, H.; Era, S. Macrophage infiltration and its prognostic implications in breast cancer: The relationship with VEGF expression and microvessel density. Oncol. Rep. 2005, 14, 425–431. [Google Scholar] [CrossRef]
- Edin, S.; Wikberg, M.L.; Dahlin, A.M.; Rutegard, J.; Oberg, A.; Oldenborg, P.A.; Palmqvist, R. The distribution of macrophages with a M1 or M2 phenotype in relation to prognosis and the molecular characteristics of colorectal cancer. PLoS ONE 2012, 7, e47045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Ge, X.; Xu, X.; Yu, S.; Wang, J.; Sun, L. Prognostic value and clinicopathological roles of phenotypes of tumour-associated macrophages in colorectal cancer. J. Cancer Res. Clin. Oncol. 2019, 145, 3005–3019. [Google Scholar] [CrossRef]
- Chao, M.P.; Jaiswal, S.; Weissman-Tsukamoto, R.; Alizadeh, A.A.; Gentles, A.J.; Volkmer, J.; Weiskopf, K.; Willingham, S.B.; Raveh, T.; Park, C.Y.; et al. Calreticulin is the dominant pro-phagocytic signal on multiple human cancers and is counterbalanced by CD47. Sci. Transl. Med. 2010, 2, 63ra94. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Fang, X.; Jiang, C.; Yao, N.; Fang, X. Thrombospondin promoted anti-tumor of adenovirus-mediated calreticulin in breast cancer: Relationship with anti-CD47. Biomed. Pharmacother. 2015, 73, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, L.; Zhao, F.; Tseng, S.; Narayanan, C.; Shura, L.; Willingham, S.; Howard, M.; Prohaska, S.; Volkmer, J.; et al. Pre-Clinical Development of a Humanized Anti-CD47 Antibody with Anti-Cancer Therapeutic Potential. PLoS ONE 2015, 10, e0137345. [Google Scholar] [CrossRef] [Green Version]
- Tseng, D.; Volkmer, J.P.; Willingham, S.B.; Contreras-Trujillo, H.; Fathman, J.W.; Fernhoff, N.B.; Seita, J.; Inlay, M.A.; Weiskopf, K.; Miyanishi, M.; et al. Anti-CD47 antibody-mediated phagocytosis of cancer by macrophages primes an effective antitumor T-cell response. Proc. Natl. Acad. Sci. USA 2013, 110, 11103–11108. [Google Scholar] [CrossRef] [Green Version]
- Brierley, J.; Gospodarowicz, M.K.; Wittekind, C. TNM Classification of Malignant Tumours, 8th ed.; John Wiley & Sons, Inc.: Chichester, West Sussex, UK; Hoboken, NJ, USA, 2017. [Google Scholar]
- Kanda, Y. Investigation of the freely available easy-to-use software ’EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total No. | CD47 Positive | CD47 Negative | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 269 | (100%) | 95 | (35%) | 174 | (65%) | ||||
| Sex | a | ||||||||
| Male | 143 | (53%) | 56 | (59%) | 87 | (50%) | 0.20 | ||
| Female | 126 | (47%) | 39 | (41%) | 87 | (50%) | |||
| Age, years (mean ± S.D.) | 68.6 ± 12.6 | 67.1 ± 14.6 | 69.4 ± 11.3 | 0.15 | b | ||||
| Size, cm (mean ± S.D.) | 5.0 ± 2.6 | 4.8 ± 2.4 | 5.1 ± 2.6 | 0.32 | b | ||||
| Tumor location | a | ||||||||
| Right-sided colon | 124 | (46%) | 42 | (44%) | 82 | (47%) | 0.89 | ||
| Left-sided colon | 86 | (32%) | 31 | (33%) | 55 | (32%) | |||
| Rectum | 59 | (22%) | 22 | (23%) | 37 | (21%) | |||
| pT stage | c | ||||||||
| pT2 | 36 | (13%) | 17 | (18%) | 19 | (11%) | 0.50 | ||
| pT3 | 189 | (70%) | 61 | (64%) | 128 | (74%) | |||
| pT4 | 44 | (16%) | 17 | (18%) | 27 | (15%) | |||
| Histological differentiation | a | ||||||||
| Well to moderately | 242 | (90%) | 80 | (84%) | 162 | (93%) | 0.032 | ||
| Poorly | 27 | (10%) | 15 | (16%) | 12 | (7%) | |||
| Mucus production | a | ||||||||
| Positive | 14 | (5%) | 9 | (9%) | 5 | (3%) | 0.040 | ||
| Negative | 255 | (95%) | 86 | (91%) | 163 | (97%) | |||
| Lymph node metastasis | a | ||||||||
| Positive | 124 | (49%) | 52 | (58%) | 72 | (44%) | 0.049 | ||
| Negative | 129 | (51%) | 38 | (42%) | 91 | (56%) | |||
| Peritoneal metastasis | a | ||||||||
| Positive | 50 | (19%) | 18 | (19%) | 32 | (18%) | 1 | ||
| Negative | 219 | (81%) | 77 | (81%) | 142 | (82%) | |||
| Distant organ metastasis | a | ||||||||
| Positive | 44 | (16%) | 17 | (18%) | 27 | (16%) | 0.61 | ||
| Negative | 225 | (84%) | 78 | (82%) | 147 | (84%) | |||
| Operation status | a | ||||||||
| Complete resection | 237 | (88%) | 85 | (89%) | 152 | (87%) | 0.70 | ||
| Incomplete resection | 32 | (12%) | 10 | (11%) | 22 | (13%) | |||
| MMR system status | a | ||||||||
| Deficient | 31 | (12%) | 14 | (15%) | 17 | (10%) | 0.24 | ||
| Preserved | 238 | (88%) | 81 | (85%) | 157 | (90%) | |||
| Total No. | SIRPA High | SIRPA Low | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 269 | (100%) | 143 | (35%) | 126 | (65%) | ||||
| Sex | a | ||||||||
| Male | 143 | (53%) | 73 | (51%) | 70 | (56%) | 0.50 | ||
| Female | 126 | (47%) | 70 | (49%) | 56 | (44%) | |||
| Age, years (mean ± S.D.) | 68.6 ± 12.6 | 67.9 ± 12.6 | 69.4 ± 12.6 | 0.37 | b | ||||
| Size, cm (mean ± S.D.) | 5.0 ± 2.6 | 5.3 ± 2.8 | 4.7 ± 2.4 | 0.037 | b | ||||
| Tumor location | a | ||||||||
| Right-sided colon | 124 | (46%) | 67 | (47%) | 57 | (45%) | 0.78 | ||
| Left-sided colon | 86 | (32%) | 47 | (33%) | 39 | (31%) | |||
| Rectum | 59 | (22%) | 29 | (20%) | 30 | (24%) | |||
| pT stage | c | ||||||||
| pT2 | 36 | (13%) | 22 | (15%) | 14 | (11%) | 0.37 | ||
| pT3 | 189 | (70%) | 100 | (70%) | 89 | (71%) | |||
| pT4 | 44 | (16%) | 21 | (15%) | 23 | (18%) | |||
| Histological differentiation | a | ||||||||
| Well to moderately | 242 | (90%) | 128 | (90%) | 114 | (90%) | 0.95 | ||
| Poorly | 27 | (10%) | 15 | (10%) | 12 | (10%) | |||
| Mucus production | a | ||||||||
| Positive | 14 | (5%) | 4 | (3%) | 10 | (8%) | 0.11 | ||
| Negative | 255 | (95%) | 139 | (97%) | 116 | (92%) | |||
| Lymph node metastasis | a | ||||||||
| Positive | 124 | (49%) | 64 | (47%) | 60 | (51%) | 0.59 | ||
| Negative | 129 | (51%) | 72 | (53%) | 57 | (49%) | |||
| Peritoneal metastasis | a | ||||||||
| Positive | 50 | (19%) | 25 | (17%) | 25 | (20%) | 0.71 | ||
| Negative | 219 | (81%) | 118 | (83%) | 101 | (80%) | |||
| Distant organ metastasis | a | ||||||||
| Positive | 44 | (16%) | 22 | (15%) | 22 | (17%) | 0.77 | ||
| Negative | 225 | (84%) | 121 | (85%) | 104 | (83%) | |||
| Operation status | a | ||||||||
| Complete resection | 237 | (88%) | 121 | (85%) | 116 | (92%) | 0.090 | ||
| Incomplete resection | 32 | (12%) | 22 | (15%) | 10 | (8%) | |||
| MMR system status | a | ||||||||
| Deficient | 31 | (12%) | 24 | (17%) | 7 | (6%) | 0.0072 | ||
| Preserved | 238 | (88%) | 119 | (83%) | 119 | (94%) | |||
| 95% CI | ||||
|---|---|---|---|---|
| HR | Min | Max | p-Value | |
| Well to moderately differentiated histology | 0.23 | 0.12 | 0.42 | <0.0001 |
| Age (<70) | 0.48 | 0.27 | 0.82 | 0.0087 |
| TAIs SIRPA High | 0.55 | 0.32 | 0.93 | 0.027 |
| Tumor CD47 High | 1.75 | 1.03 | 2.98 | 0.038 |
| Lymph node metastasis | 2.26 | 1.31 | 3.91 | 0.0036 |
| Peritoneal metastasis | 5.80 | 3.23 | 10.43 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugimura-Nagata, A.; Koshino, A.; Inoue, S.; Matsuo-Nagano, A.; Komura, M.; Riku, M.; Ito, H.; Inoko, A.; Murakami, H.; Ebi, M.; et al. Expression and Prognostic Significance of CD47–SIRPA Macrophage Checkpoint Molecules in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 2690. https://doi.org/10.3390/ijms22052690
Sugimura-Nagata A, Koshino A, Inoue S, Matsuo-Nagano A, Komura M, Riku M, Ito H, Inoko A, Murakami H, Ebi M, et al. Expression and Prognostic Significance of CD47–SIRPA Macrophage Checkpoint Molecules in Colorectal Cancer. International Journal of Molecular Sciences. 2021; 22(5):2690. https://doi.org/10.3390/ijms22052690
Chicago/Turabian StyleSugimura-Nagata, Akane, Akira Koshino, Satoshi Inoue, Aya Matsuo-Nagano, Masayuki Komura, Miho Riku, Hideaki Ito, Akihito Inoko, Hideki Murakami, Masahide Ebi, and et al. 2021. "Expression and Prognostic Significance of CD47–SIRPA Macrophage Checkpoint Molecules in Colorectal Cancer" International Journal of Molecular Sciences 22, no. 5: 2690. https://doi.org/10.3390/ijms22052690