
The Human Gut Microbiota: A Key Mediator of Osteoporosis and Osteogenesis

 ,
, 
Abstract
:1. Introduction
2. Osteoporosis
3. Clinical Treatment of Osteoporosis
3.1. Lifestyle Modifications
3.2. Nutraceutical Supplementation
3.2.1. Probiotic Supplementation
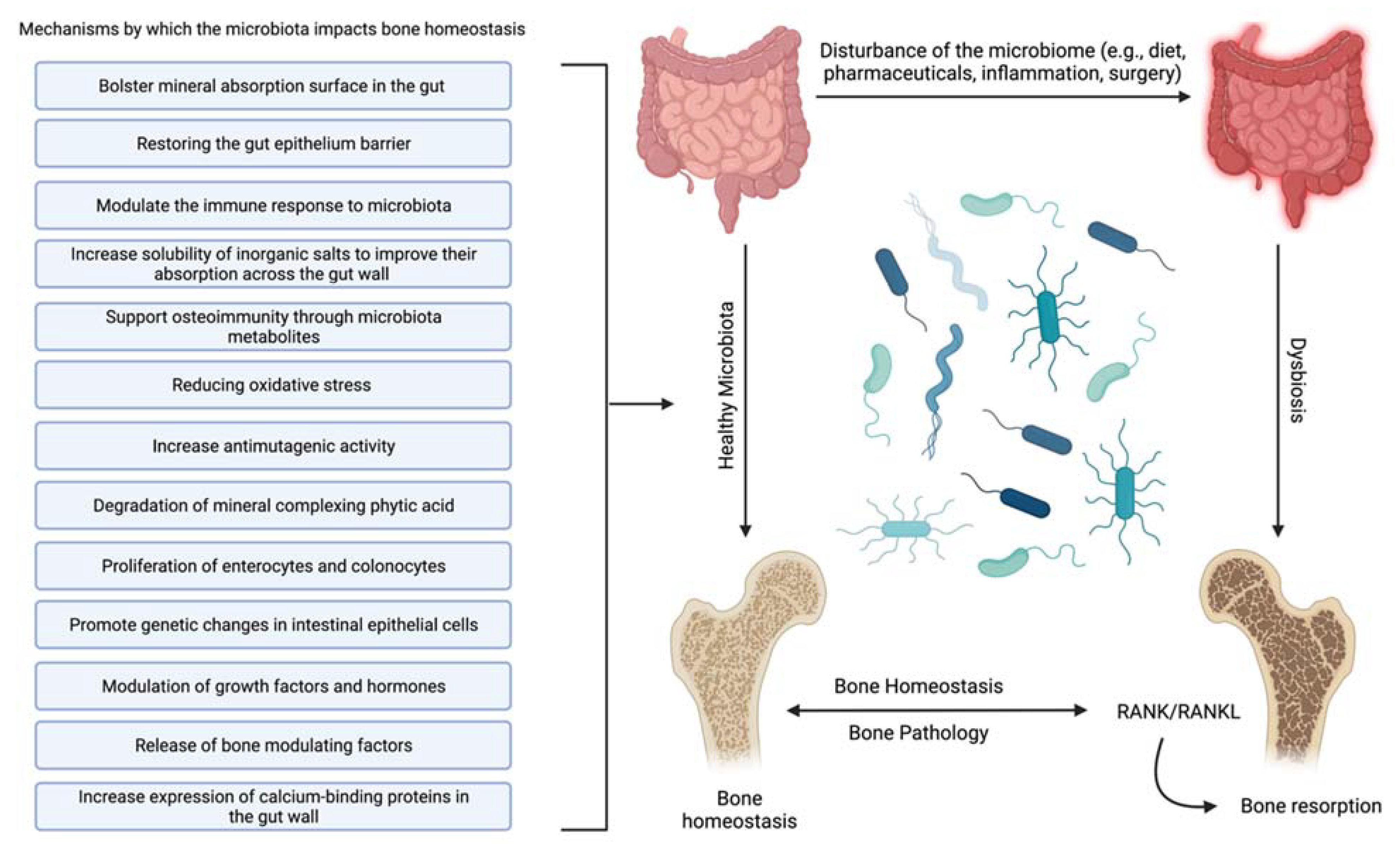
3.2.2. Mechanisms of Microbiota Impacts
- Increase solubility of inorganic salts to improve their absorption across the gut wall [56]—The availability of inorganic salts such as phosphate is critical for bone mineral deposition by osteoblasts and bone homeostasis [57]. This effect may be partially due to the metabolism of mineral complexed phytic acid [58] by microbial synthesized phytase into inorganic phosphate and a myoinositol phosphate derivative [52].
- Bolster mineral absorption surface in the gut—By promoting the proliferation of enterocytes and colonocytes, gut microbiota homeostasis is mediated, and mineral absorption in the gut is supported. Furthermore, increased colonocyte metabolism has been documented to promote obligate anaerobes, which are known to metabolize fiber, thereby increasing SCFAs, which is critical for bone homeostasis, among other physiological functions [59].
- Restore and maintain gut epithelium barrier—Enhancing the gut’s barrier function is integral to gastrointestinal immunity [60]. A healthy gut epithelial barrier prevents the hyperpermeability that comes with damaged tight junctions [61]. Hyperpermeability or a “leaky gut” leaches unusually high levels of inflammatory cytokines, producing systemic inflammation and leading to hyperactive osteoclasts and bone degradation [62].
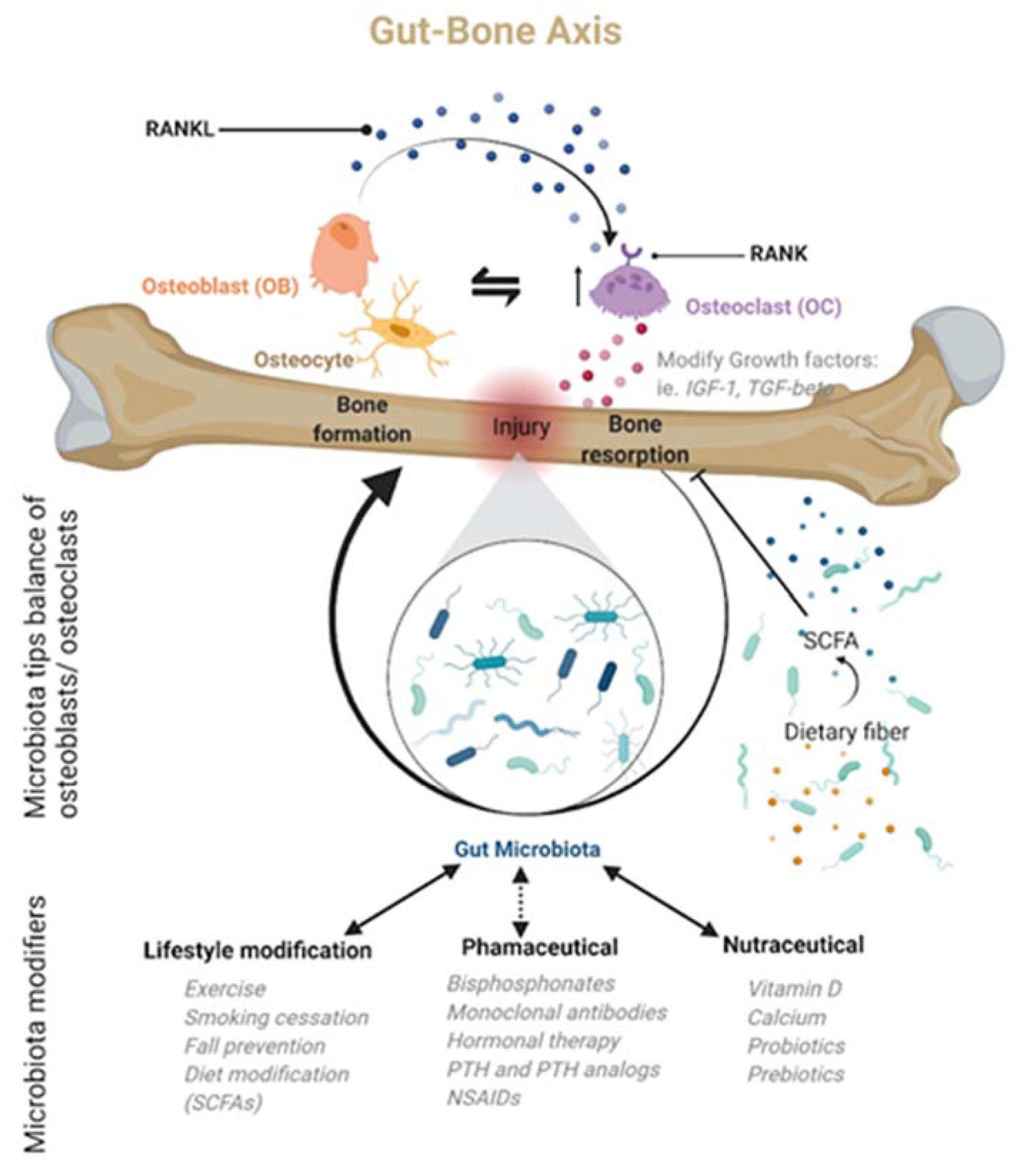
- Support osteoimmunity through microbiota metabolites—Short-chain fatty acids (SCFAs) produced by the gut microbiota offer anti-inflammatory effects by inhibiting the activation of nuclear factor kappa-light-chain-enhancer of activated B cells, reducing auto-immune inflammation [63]. Additionally, SCFAs, specifically propionate and butyrate, metabolically reprogram osteoclasts by downregulating TRAF6 and NFATc1 to inhibit osteoclastogenesis and bone resorption, effectively increasing bone density without directly altering osteoblasts [30].
- Reduce oxidative stress [64]—Oxidative stress is documented to cause excessive osteocyte apoptosis, which generates an imbalance favoring osteoclastogenesis, leading to increased bone remodeling, turnover, and loss [65]. Strain-specific probiotics can relieve oxidative stress by producing several antioxidant molecules (e.g., glutathione, folate, and exopolysaccharide). In addition, the SCFAs produced by several gut microbiota can also help to relieve oxidative stress by promoting the production of antioxidant molecules [66,67].
- Modulate the immune response to microbiota [68]—The effects of normal gut microbiota are appreciated when discussing the abnormal rather than the normal. In an abnormal state, the immune system’s reaction to microbiota stimulation leads to an increase in circulating osteoclastogenic cytokines through the action of T-cells. This degradative process is not active in normal gut microbiota states [8].
- Promote genetic changes in intestinal epithelial cells [69]—Although it is not completely clear how they accomplish it, specific gut microbiota can prompt the genetic modification of cells. Recently, it was shown that Bifidobacterium lactis species upregulated cyclooxygenase-1 (Cox-1) and downregulated Cox-2 gene expression in a Caco-2 cell culture model. This outcome is thought to lead to a decrease in tissue damage and inflammation [70].
- Increase antimutagenic activity [52,71,72]—Although this capability has largely been explored in the context of cancer [73], certain species of lactic acid microbiota can bind potent mutagens such as pyrolyzates [74,75] and heterocyclic amines [76,77] in the gut to decrease the mutagenic activity of these compounds. Reducing DNA damage reduces inflammation, protects the gut wall, increases mineral absorption, and suppresses osteoporosis [78].
- Increase expression of calcium-binding proteins in the gut wall—Increasing calbindin-D9k gene expression in the gut wall can increase the ability to absorb calcium [79], effectively suppressing bone degradation and promoting bone deposition by suppressing the actions of parathyroid hormone [39]. In addition, enhanced calcium absorption and inhibition of parathyroid hormone activity and insulin-like growth factor 1 production can also modify the development of osteoclasts and osteoblasts [80,81].
- Modulation of growth factors and hormones [82]—The gut microbiota should be considered an endocrine organ based on the plethora of secreted molecules. Specifically, the gut microbiota promotes the production of IGF-1 through a proposed SCFA-mediated pathway [83]. IGF-1 is known to stimulate the differentiation of osteoblasts, osteoclasts, and chondrocytes. The gut microbiota may also enhance bone degradation through a cortisol-mediated interaction [82,84]; however, the evidence is indirect, and the precise mechanism is unclear. The gut microbiota also modulates gut serotonin production, a molecule that interacts with bone cells and has been suggested to act as a bone mass regulator [85].
3.3. Pharmaceuticals
3.4. Fracture in Osteoporotic Patients: Surgical Fracture Reduction

{kind=link}
{kind=link}
| Intervention | Mechanism | Treatment Recommendation | References |
|---|---|---|---|
| Lifestyle Modification | |||
| Exercise | Exercise-induced mechanical loading increases bone mass by regulating the hormones, cytokines, signaling pathways, and noncoding RNAs in bone metabolism. | Patients who have osteopenia or osteoporosis, or would like to prevent it, should exercise for at least 30 min three times a week. Resistance training, jogging, jumping, and walking are generally considered effective. | [102,103,104,105] |
| Diet | 1. Reactive oxygen species induce the apoptosis of osteoblasts and osteocytes. Excessive osteocyte apoptosis is linked to oxidative stress, which causes an imbalance in favor of osteoclastogenesis, resulting in more significant bone remodeling turnover and loss. | 1. A diet high in antioxidant-rich foods, such as foods high in polyphenols, supports antiresorptive therapies for the treatment and prevention of bone loss. | [62,65,106,107,108,109] |
| 2. In chronic inflammatory states, overproduction of cytokines such as tumor necrosis factor, interleukin-1 (IL-1), IL-6, and IL-17 is linked to inflammation. In addition, specific cytokines can impede osteoblast function; their overexpression during inflammation leads to excessive bone degradation, primarily due to hyperactivation of osteoclasts. | 2. An anti-inflammatory diet may delay the course of osteoporosis by regulating inflammatory activity, modifying the lipid profile, boosting antioxidant levels, and altering the gut’s microbiota. | ||
| Smoking Cessation | 1. Smoking lowers circulating estrogen levels, as seen by higher blood concentrations of follicle-stimulating hormone and luteinizing hormone. Smoking may also increase bone resorption, causing a rise in blood calcium levels, a drop in serum parathyroid hormone levels, and increased urine hydroxyproline and pyridinoline excretion. | Smoking cessation is strongly recommended to all patients concerned with their skeletal health. | [110,111,112] |
| 2. The chemical composition of smoking (and vaping to a lesser extent) can interfere with other therapeutic measures included in a comprehensive treatment regimen. | |||
| Nutraceuticals | |||
| Calcium with Vitamin D | Vitamin D3 (Cholecalciferol) increases calcium uptake in the small intestine. Increased calcium in the blood promotes bone deposition and suppresses bone resorption. | Total calcium intake (diet + supplements) should approximate 1200 mg/day. Women should also ingest a total of 800 international units of vitamin D3 daily. | [113,114,115] |
| Probiotics | Can improve calcium balance, prevent secondary hyperparathyroidism, and attenuate age-related increase in bone resorption and bone loss via multiple proposed mechanisms outlined in Section 3 of this review. | No probiotic strategy is currently included in the standard of care or as a primary treatment for osteoporosis. | [52,53,116,117] |
| It may benefit overall bone health if consumed daily as preparations with active live cultures containing bacteria, such as lactobacilli, lactococci, or bifidobacteria isolated from natural environments. | |||
| Pharmaceuticals: Antiresorptive Agents | |||
| Bisphosphonates | Nitrogen-containing bisphosphonates selectively inhibit farnesyl pyrophosphate synthase (FPPS) within osteoclasts, inhibiting osteoclast activity. | Oral bisphosphonates are considered as first-line treatment. However, alendronate or risedronate are commonly used and are considered safe and efficacious. | [95,118,119] |
| Denosumab | Binds with high specificity and affinity to the cytokine RANKL, thereby inhibiting its action; as a result, osteoclast recruitment, maturation, and action are suppressed, and bone resorption slows. | Used as initial therapy in certain patients at high risk for fracture, such as older patients who have difficulty with the dosing requirements of oral bisphosphonates or who have markedly impaired renal function. | [120,121,122] |
| Pharmaceuticals: Anabolic Agents | |||
| PTH/PTH-RP Analogs | Stimulates bone formation and activates bone remodeling through RANK/RANKL. Intermittent administration of recombinant human PTH or PTHrP has been shown to stimulate bone formation more than resorption. | Indicated in severe cases of osteoporosis (T-score of ≤−3.5 even in the absence of fractures, or T-score of ≤−2.5 plus a fragility fracture). May prescribe for patients with osteoporosis who cannot tolerate bisphosphonates or who have contraindications to oral bisphosphonates and patients who fail other osteoporosis therapies. | [123,124,125] |
| Romosozumab | Sclerostin inhibits WNT/catenin signaling in osteoblasts and osteocytes, decreasing OPG expression. Direct actions of sclerostin on osteocytes stimulate RANKL expression. Monoclonal anti-sclerostin antibody (Romosozumab) inhibits sclerostin and enhances osteoblast function to improve bone mass and reduce fractures. | Not considered initial therapy for most patients with osteoporosis. Candidates include patients with multiple fragility fractures, those at high risk for fracture, or those who cannot tolerate any other osteoporosis therapies. | [88,126,127] |
| Pharmaceuticals: Hormone Modulators | |||
| Selective Estrogen Receptor Modulators | Acts upon estrogen-sensitive tissues and functions as an estrogen agonist in bone to prevent bone loss and has estrogen antagonist activity to block some estrogen effects in the breast and uterine tissues. Decreases bone resorption, increasing BMD and decreasing fracture incidence. | Usually recommended for osteoporosis when there is an independent need for breast cancer prophylaxis. | [128,129] |
| Estrogen/Progestin Therapy | Estrogens regulate the activity of bone-forming osteoblasts and bone-resorbing osteoclasts, which regulate bone mass and strength. The cellular actions of estrogen are mediated primarily through estrogen receptor alpha (ERα), which is present in estrogen-sensitive tissues such as bone. | No longer a first-line approach. Indicated for postmenopausal women with persistent postmenopausal symptoms and an indication for antiresorptive therapy who cannot tolerate other drugs. | [130,131,132] |
4. Human Gut Microbiota: Physiologic Role
Human Gut Microbiota: Role in Osteoporosis and Osteoclastogenesis
5. Bioactive Compounds and the Human Gut Microbiome
5.1. Pharmacological Gut Microbiota and Fracture Treatment: Known Interactions
5.1.1. Bisphosphonates
5.1.2. Monoclonal Antibodies
5.1.3. Hormonal Therapy
5.1.4. PTH-Related Analogs
5.1.5. Non-Steroidal Anti-Inflammatory Drugs
| Treatment | Gut Microbiota Interactions | References |
|---|---|---|
| Bisphosphonates | Long-term use is hypothesized to lead to gut microbiome-related immunosuppression via inhibition of the mevalonate (MVA) pathway. | [164,166] |
| Monoclonal Antibodies | Effects of Denosumab on bone health may either be enhanced or suppressed based on alterations in the gut microbiota. | [167,172] |
| Romosozumab may impact the gut microbiota through the WNT pathway. | ||
| Hormonal Therapy | Estrogen promotes a healthy and diverse mix of commensal flora, which improves T-regulatory cell function and suppresses inflammation, suggesting that manipulating the gut microbiota in conjunction with estrogen-based therapies may improve the overall inflammatory state and strengthen bone mineral density. | [177] |
| PTH-Related Analogs | More studies are needed to determine if PTH-related analogs can be used to enhance the gut microbiota’s role in osteoporotic bone healing. | [123,125] |
| Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) | May mimic broad-spectrum antimicrobial properties. | [194,195] |
| Inhibit some positive effects of microbiota through enzyme inhibition. |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bahney, C.S.; Zondervan, R.L.; Allison, P.; Theologis, A.; Ashley, J.W.; Ahn, J.; Miclau, T.; Marcucio, R.S.; Hankenson, K.D. Cellular Biology of Fracture Healing. J. Orthop. Res. 2019, 37, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Xie, Z.; Sun, J.; Huang, S.; Chen, Y.; Li, C.; Sun, X.; Xia, B.; Tian, L.; Guo, C.; et al. Gut Microbiome Reveals Specific Dysbiosis in Primary Osteoporosis. Front. Cell. Infect. Microbiol. 2020, 10, 160. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Xu, S.; Zhang, B.; Xiao, C.; Chen, Z.; Si, F.; Fu, J.; Lin, X.; Zheng, G.; Yu, G.; et al. Gut Microbiota and Metabolite Alterations Associated with Reduced Bone Mineral Density or Bone Metabolic Indexes in Postmenopausal Osteoporosis. Aging 2020, 12, 8583–8604. [Google Scholar] [CrossRef]
- Proal, A.D.; Lindseth, I.A.; Marshall, T.G. Microbe-Microbe and Host-Microbe Interactions Drive Microbiome Dysbiosis and Inflammatory Processes. Discov. Med. 2017, 23, 51–60. [Google Scholar] [PubMed]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the Gut Microbiota in Disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Gao, W.; Wang, B.; Zhao, H.; Zeng, Y.; Ji, Y.; Hao, D. Diversity Analysis of Gut Microbiota in Osteoporosis and Osteopenia Patients. PeerJ 2017, 5, e3450. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-W.; Li, Y.-J.; Lu, P.-P.; Dai, G.-C.; Chen, X.-X.; Rui, Y.-F. The Modulatory Effect and Implication of Gut Microbiota on Osteoporosis: From the Perspective of “Brain–Gut–Bone” Axis. Food Funct. 2021, 12, 5703–5718. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Pal, S.; Paterson, C.W.; Li, J.-Y.; Tyagi, A.M.; Adams, J.; Coopersmith, C.M.; Weitzmann, M.N.; Pacifici, R. Ovariectomy Induces Bone Loss via Microbial-Dependent Trafficking of Intestinal TNF+ T Cells and Th17 Cells. J. Clin. Investig. 2021, 131, e143137. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, M.M.; Jones, R.M.; Schett, G.; Pacifici, R. The Gut-Bone Axis: How Bacterial Metabolites Bridge the Distance. J. Clin. Investig. 2019, 129, 3018–3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheen, J.R.; Garla, V.V. Fracture Healing Overview; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Kenkre, J.; Bassett, J. The Bone Remodelling Cycle. Ann. Clin. Biochem. Int. J. Lab. Med. 2018, 55, 308–327. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast–Osteoclast Interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef]
- Li, J.-Y.; Yu, M.; Pal, S.; Tyagi, A.M.; Dar, H.; Adams, J.; Weitzmann, M.N.; Jones, R.M.; Pacifici, R. Parathyroid Hormone–Dependent Bone Formation Requires Butyrate Production by Intestinal Microbiota. J. Clin. Investig. 2020, 130, 1767–1781. [Google Scholar] [CrossRef]
- Castaneda, M.; Smith, K.M.; Nixon, J.C.; Hernandez, C.J.; Rowan, S. Alterations to the Gut Microbiome Impair Bone Tissue Strength in Aged Mice. Bone Rep. 2021, 14, 101065. [Google Scholar] [CrossRef] [PubMed]
- Sjögren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Bäckhed, F.; Ohlsson, C. The Gut Microbiota Regulates Bone Mass in Mice. J. Bone Miner. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielby, R.; Jones, E.; McGonagle, D. The Role of Mesenchymal Stem Cells in Maintenance and Repair of Bone. Injury 2007, 38, S26–S32. [Google Scholar] [CrossRef] [PubMed]
- Park-Min, K.-H. Mechanisms Involved in Normal and Pathological Osteoclastogenesis. Cell. Mol. Life Sci. 2018, 75, 2519–2528. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, H. Discovery of the RANKL/RANK/OPG System. J. Bone Miner. Metab. 2021, 39, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C.; et al. Osteoclast Differentiation by RANKL and OPG Signaling Pathways. J. Bone Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.-Y.; Yang, Y.; Jung, H. Molecular Mechanisms and Emerging Therapeutics for Osteoporosis. Int. J. Mol. Sci. 2020, 21, 7623. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Endo, I. RANKL as a Target for the Treatment of Osteoporosis. J. Bone Miner. Metab. 2021, 39, 91–105. [Google Scholar] [CrossRef]
- Li, W.-F.; Hou, S.-X.; Yu, B.; Jin, D.; Férec, C.; Chen, J.-M. Genetics of Osteoporosis: Perspectives for Personalized Medicine. Pers. Med. 2010, 7, 655–668. [Google Scholar] [CrossRef]
- Rachner, T.D.; Khosla, S.; Hofbauer, L.C. Osteoporosis: Now and the Future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef] [Green Version]
- Aspray, T.J.; Hill, T.R. Osteoporosis and the Ageing Skeleton. In Biochemistry and Cell Biology of Ageing: Part II Clinical Science; Harris, J.R., Korolchuk, V.I., Eds.; Subcellular Biochemistry; Springer: Singapore, 2019; Volume 91, pp. 453–476. ISBN 9789811336805. [Google Scholar]
- Langdahl, B.; Ferrari, S.; Dempster, D.W. Bone Modeling and Remodeling: Potential as Therapeutic Targets for the Treatment of Osteoporosis. Ther. Adv. Musculoskelet. Dis. 2016, 8, 225–235. [Google Scholar] [CrossRef] [Green Version]
- McCabe, L.R.; Irwin, R.; Tekalur, A.; Evans, C.; Schepper, J.D.; Parameswaran, N.; Ciancio, M. Exercise Prevents High Fat Diet-Induced Bone Loss, Marrow Adiposity and Dysbiosis in Male Mice. Bone 2019, 118, 20–31. [Google Scholar] [CrossRef]
- Park, S.G.; Jeong, S.U.; Lee, J.H.; Ryu, S.H.; Jeong, H.J.; Sim, Y.J.; Kim, D.K.; Kim, G.C. The Changes of CTX, DPD, Osteocalcin, and Bone Mineral Density During the Postmenopausal Period. Ann. Rehabil. Med. 2018, 42, 441–448. [Google Scholar] [CrossRef]
- Hardy, R.S.; Zhou, H.; Seibel, M.J.; Cooper, M.S. Glucocorticoids and Bone: Consequences of Endogenous and Exogenous Excess and Replacement Therapy. Endocr. Rev. 2018, 39, 519–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, S.; Omata, Y.; Hofmann, J.; Böttcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Krönke, G.; et al. Short-Chain Fatty Acids Regulate Systemic Bone Mass and Protect from Pathological Bone Loss. Nat. Commun. 2018, 9, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montalvany-Antonucci, C.C.; Duffles, L.F.; de Arruda, J.A.A.; Zicker, M.C.; de Oliveira, S.; Macari, S.; Garlet, G.P.; Madeira, M.F.M.; Fukada, S.Y.; Andrade, I.; et al. Short-Chain Fatty Acids and FFAR2 as Suppressors of Bone Resorption. Bone 2019, 125, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Mei, F.; Meng, K.; Gu, Z.; Yun, Y.; Zhang, W.; Zhang, C.; Zhong, Q.; Pan, F.; Shen, X.; Xia, G.; et al. Arecanut (Areca catechu L.) Seed Polyphenol-Ameliorated Osteoporosis by Altering Gut Microbiome via LYZ and the Immune System in Estrogen-Deficient Rats. J. Agric. Food Chem. 2021, 69, 246–258. [Google Scholar] [CrossRef]
- Lee, T.; Suh, H.S. Associations between Dietary Fiber Intake and Bone Mineral Density in Adult Korean Population: Analysis of National Health and Nutrition Examination Survey in 2011. J. Bone Metab. 2019, 26, 151. [Google Scholar] [CrossRef]
- Dai, Z.; Zhang, Y.; Lu, N.; Felson, D.T.; Kiel, D.P.; Sahni, S. Association between dietary fiber intake and bone loss in the Framingham Offspring Study: Association between dietary fiber intake and bone loss. J. Bone Miner. Res. 2018, 33, 241–249. [Google Scholar] [CrossRef]
- Adhikary, S.; Choudhary, D.; Ahmad, N.; Karvande, A.; Kumar, A.; Banala, V.T.; Mishra, P.R.; Trivedi, R. Dietary Flavonoid Kaempferol Inhibits Glucocorticoid-Induced Bone Loss by Promoting Osteoblast Survival. Nutrition 2018, 53, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Alayat, M.S.M.; Abdel-Kafy, E.M.; Thabet, A.A.M.; Abdel-Malek, A.S.; Ali, T.H.; Header, E.A. Long-Term Effect of Pulsed Nd-YAG Laser Combined with Exercise on Bone Mineral Density in Men with Osteopenia or Osteoporosis: 1 Year of Follow-Up. Photomed. Laser Surg. 2018, 36, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Daly, R.M.; Dalla Via, J.; Duckham, R.L.; Fraser, S.F.; Helge, E.W. Exercise for the Prevention of Osteoporosis in Postmenopausal Women: An Evidence-Based Guide to the Optimal Prescription. Braz. J. Phys. Ther. 2019, 23, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Pagnotti, G.M.; Styner, M.; Uzer, G.; Patel, V.S.; Wright, L.E.; Ness, K.K.; Guise, T.A.; Rubin, J.; Rubin, C.T. Combating Osteoporosis and Obesity with Exercise: Leveraging Cell Mechanosensitivity. Nat. Rev. Endocrinol. 2019, 15, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shen, S.; Mao, H.; Chen, C.; Zhao, J.; Han, C.; Shi, L.; Li, Y. Effects of Calcium on the Structure of Intestinal Microbiota in Ovariectomized Rats. Wei Sheng Yan Jiu 2021, 50, 267–273. [Google Scholar]
- Weaver, C.M.; Alexander, D.D.; Boushey, C.J.; Dawson-Hughes, B.; Lappe, J.M.; LeBoff, M.S.; Liu, S.; Looker, A.C.; Wallace, T.C.; Wang, D.D. Calcium plus Vitamin D Supplementation and Risk of Fractures: An Updated Meta-Analysis from the National Osteoporosis Foundation. Osteoporos. Int. 2016, 27, 367–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, I.R.; Bristow, S.M.; Bolland, M.J. Calcium Supplements: Benefits and Risks. J. Intern. Med. 2015, 278, 354–368. [Google Scholar] [CrossRef]
- Cerani, A.; Zhou, S.; Forgetta, V.; Morris, J.A.; Trajanoska, K.; Rivadeneira, F.; Larsson, S.C.; Michaëlsson, K.; Richards, J.B. Genetic Predisposition to Increased Serum Calcium, Bone Mineral Density, and Fracture Risk in Individuals with Normal Calcium Levels: Mendelian Randomisation Study. BMJ 2019, 366, l4410. [Google Scholar] [CrossRef] [Green Version]
- Uenishi, K.; Tokiwa, M.; Kato, S.; Shiraki, M. Stimulation of Intestinal Calcium Absorption by Orally Administrated Vitamin D3 Compounds: A Prospective Open-Label Randomized Trial in Osteoporosis. Osteoporos. Int. 2018, 29, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Pilz, S.; Zittermann, A.; Trummer, C.; Theiler-Schwetz, V.; Lerchbaum, E.; Keppel, M.H.; Grübler, M.R.; März, W.; Pandis, M. Vitamin D Testing and Treatment: A Narrative Review of Current Evidence. Endocr. Connect. 2019, 8, R27–R43. [Google Scholar] [CrossRef] [Green Version]
- Whisner, C.M.; Castillo, L.F. Prebiotics, Bone and Mineral Metabolism. Calcif. Tissue Int. 2018, 102, 443–479. [Google Scholar] [CrossRef] [Green Version]
- Schepper, J.D.; Collins, F.; Rios-Arce, N.D.; Kang, H.J.; Schaefer, L.; Gardinier, J.D.; Raghuvanshi, R.; Quinn, R.A.; Britton, R.; Parameswaran, N.; et al. Involvement of the Gut Microbiota and Barrier Function in Glucocorticoid-Induced Osteoporosis. J. Bone Miner. Res. 2020, 35, 801–820. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Jia, X.; Mo, L.; Liu, C.; Zheng, L.; Yuan, Q.; Zhou, X. Intestinal Microbiota: A Potential Target for the Treatment of Postmenopausal Osteoporosis. Bone Res. 2017, 5, 17046. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, T.; Hatanaka, M.; Hoshino, T.; Takara, T.; Tanaka, K.; Shimizu, A.; Morita, H.; Nakamura, T. Effect of Bacillus subtilis C-3102 on Bone Mineral Density in Healthy Postmenopausal Japanese Women: A Randomized, Placebo-Controlled, Double-Blind Clinical Trial. Biosci. Microbiota Food Health 2018, 37, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, A.G.; Sundh, D.; Bäckhed, F.; Lorentzon, M. Lactobacillus reuteri Reduces Bone Loss in Older Women with Low Bone Mineral Density: A Randomized, Placebo-Controlled, Double-Blind, Clinical Trial. J. Intern. Med. 2018, 284, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafarnejad, S.; Djafarian, K.; Fazeli, M.R.; Yekaninejad, M.S.; Rostamian, A.; Keshavarz, S.A. Effects of a Multispecies Probiotic Supplement on Bone Health in Osteopenic Postmenopausal Women: A Randomized, Double-Blind, Controlled Trial. J. Am. Coll. Nutr. 2017, 36, 497–506. [Google Scholar] [CrossRef]
- Jansson, P.-A.; Curiac, D.; Ahrén, I.L.; Hansson, F.; Niskanen, T.M.; Sjögren, K.; Ohlsson, C. Probiotic Treatment Using a Mix of Three Lactobacillus Strains for Lumbar Spine Bone Loss in Postmenopausal Women: A Randomised, Double-Blind, Placebo-Controlled, Multicentre Trial. Lancet Rheumatol. 2019, 1, e154–e162. [Google Scholar] [CrossRef]
- Nath, A.; Molnár, M.A.; Csighy, A.; Kőszegi, K.; Galambos, I.; Huszár, K.P.; Koris, A.; Vatai, G. Biological Activities of Lactose-Based Prebiotics and Symbiosis with Probiotics on Controlling Osteoporosis, Blood-Lipid and Glucose Levels. Medicina 2018, 54, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, R.; Biver, E. Are Probiotics the New Calcium and Vitamin D for Bone Health? Curr. Osteoporos. Rep. 2020, 18, 273–284. [Google Scholar] [CrossRef]
- Abboud, M.; Papandreou, D. Gut Microbiome, Probiotics and Bone: An Updated Mini Review. Open Access Maced. J. Med. Sci. 2019, 7, 478–481. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.; Park, C.; Lee, J.; Park, D.H.; Jeong, S.; Yun, C.-H.; Park, O.-J.; Han, S.H. Regulation of Bone Cell Differentiation and Activation by Microbe-Associated Molecular Patterns. Int. J. Mol. Sci. 2021, 22, 5805. [Google Scholar] [CrossRef] [PubMed]
- Bongers, A.; van den Heuvel, E.G. Prebiotics and the Bioavailability of Minerals and Trace Elements. Food Rev. Int. 2003, 19, 397–422. [Google Scholar] [CrossRef]
- Qin, L.; Liu, W.; Cao, H.; Xiao, G. Molecular Mechanosensors in Osteocytes. Bone Res. 2020, 8, 23. [Google Scholar] [CrossRef]
- Pallauf, J.; Rimbach, G. Nutritional Significance of Phytic Acid and Phytase. Arch. Tierernaehrung 1997, 50, 301–319. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Bäumler, A.J. Colonocyte Metabolism Shapes the Gut Microbiota. Science 2018, 362, eaat9076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaboriau-Routhiau, V.; Cerf-Bensussan, N. Gut microbiota and development of the immune system. Med. Sci. 2016, 32, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T. Regulation of the Intestinal Barrier by Nutrients: The Role of Tight Junctions. Anim. Sci. J. Nihon Chikusan Gakkaiho 2020, 91, e13357. [Google Scholar] [CrossRef] [Green Version]
- Redlich, K.; Smolen, J.S. Inflammatory Bone Loss: Pathogenesis and Therapeutic Intervention. Nat. Rev. Drug Discov. 2012, 11, 234–250. [Google Scholar] [CrossRef]
- Park, J.-S.; Lee, E.-J.; Lee, J.-C.; Kim, W.-K.; Kim, H.-S. Anti-Inflammatory Effects of Short Chain Fatty Acids in IFN-Gamma-Stimulated RAW 264.7 Murine Macrophage Cells: Involvement of NF-KappaB and ERK Signaling Pathways. Int. Immunopharmacol. 2007, 7, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Tomasello, G.; Mazzola, M.; Leone, A.; Sinagra, E.; Zummo, G.; Farina, F.; Damiani, P.; Cappello, F.; Gerges Geagea, A.; Jurjus, A.; et al. Nutrition, Oxidative Stress and Intestinal Dysbiosis: Influence of Diet on Gut Microbiota in Inflammatory Bowel Diseases. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czechoslov. 2016, 160, 461–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative Stress in Bone Remodeling: Role of Antioxidants. Clin. Cases Miner. Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, E521. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.M.A.E.; Bast, A.; Vanhoutvin, S.A.L.W.; Fischer, M.A.J.G.; Kodde, A.; Troost, F.J.; Venema, K.; Brummer, R.-J.M. Butyrate Modulates Oxidative Stress in the Colonic Mucosa of Healthy Humans. Clin. Nutr. Edinb. Scotl. 2009, 28, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Rooks, M.G.; Garrett, W.S. Gut Microbiota, Metabolites and Host Immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Mahkonen, A.; Putaala, H.; Mustonen, H.; Rautonen, N.; Puolakkainen, P. Lactobacillus Acidophilus 74-2 and Butyrate Induce Cyclooxygenase (COX)-1 Expression in Gastric Cancer Cells. Immunopharmacol. Immunotoxicol. 2008, 30, 503–518. [Google Scholar] [CrossRef] [PubMed]
- Nurmi, J.T.; Puolakkainen, P.A.; Rautonen, N.E. Bifidobacterium Lactis Sp. 420 up-Regulates Cyclooxygenase (Cox)-1 and down-Regulates Cox-2 Gene Expression in a Caco-2 Cell Culture Model. Nutr. Cancer 2005, 51, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Motta, G.E.; Molognoni, L.; Daguer, H.; Angonese, M.; da Silva Correa Lemos, A.L.; Dafre, A.L.; De Dea Lindner, J. The Potential of Bacterial Cultures to Degrade the Mutagen 2-Methyl-1,4-Dinitro-Pyrrole in a Processed Meat Model. Food Res. Int. 2020, 136, 109441. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-González, P.F.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Aguilar-Toalá, J.E.; Hall, F.G.; Urbizo-Reyes, U.C.; Liceaga, A.M.; Hernandez-Mendoza, A.; García, H.S. Protective Role of Lactic Acid Bacteria and Yeasts as Dietary Carcinogen-Binding Agents—A Review. Crit. Rev. Food Sci. Nutr. 2020, 1–21, in press. [Google Scholar] [CrossRef]
- Śliżewska, K.; Markowiak-Kopeć, P.; Śliżewska, W. The Role of Probiotics in Cancer Prevention. Cancers 2020, 13, 20. [Google Scholar] [CrossRef]
- Morotomi, M.; Mutai, M. In Vitro Binding of Potent Mutagenic Pyrolysates to Intestinal Bacteria. J. Natl. Cancer Inst. 1986, 77, 195–201. [Google Scholar]
- Zhang, X.B.; Ohta, Y. In Vitro Binding of Mutagenic Pyrolyzates to Lactic Acid Bacterial Cells in Human Gastric Juice. J. Dairy Sci. 1991, 74, 752–757. [Google Scholar] [CrossRef]
- Orrhage, K.; Sillerström, E.; Gustafsson, J.-Å.; Nord, C.E.; Rafter, J. Binding of Mutagenic Heterocyclic Amines by Intestinal and Lactic Acid Bacteria. Mutat. Res. Mol. Mech. Mutagen. 1994, 311, 239–248. [Google Scholar] [CrossRef]
- Kim, H.S.; Hur, S.J. Changes in Carcinogenic Heterocyclic Amines during in Vitro Digestion. J. Heterocycl. Chem. 2019, 56, 759–764. [Google Scholar] [CrossRef]
- Weaver, C.M. Diet, Gut Microbiome, and Bone Health. Curr. Osteoporos. Rep. 2015, 13, 125–130. [Google Scholar] [CrossRef]
- Fukushima, A.; Aizaki, Y.; Sakuma, K. Short-Chain Fatty Acids Increase the Level of Calbindin-D9k Messenger RNA in Caco-2 Cells. J. Nutr. Sci. Vitaminol. 2012, 58, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Charles, J.F. Gut Microbiota and IGF-1. Calcif. Tissue Int. 2018, 102, 406–414. [Google Scholar] [CrossRef]
- Wang, Y.; Nishida, S.; Elalieh, H.Z.; Long, R.K.; Halloran, B.P.; Bikle, D.D. Role of IGF-I Signaling in Regulating Osteoclastogenesis. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2006, 21, 1350–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Charles, J.F. Gut Microbiome and Bone: To Build, Destroy or Both? Curr. Osteoporos. Rep. 2017, 15, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Herzog, J.W.; Tsang, K.; Brennan, C.A.; Bower, M.A.; Garrett, W.S.; Sartor, B.R.; Aliprantis, A.O.; Charles, J.F. Gut Microbiota Induce IGF-1 and Promote Bone Formation and Growth. Proc. Natl. Acad. Sci. USA 2016, 113, E7554–E7563. [Google Scholar] [CrossRef] [Green Version]
- Soldi, S.; Tagliacarne, S.C.; Valsecchi, C.; Perna, S.; Rondanelli, M.; Ziviani, L.; Milleri, S.; Annoni, A.; Castellazzi, A. Effect of a Multistrain Probiotic (Lactoflorene® Plus) on Inflammatory Parameters and Microbiota Composition in Subjects with Stress-Related Symptoms. Neurobiol. Stress 2018, 10, 100138. [Google Scholar] [CrossRef]
- D’Amelio, P.; Sassi, F. Gut Microbiota, Immune System, and Bone. Calcif. Tissue Int. 2018, 102, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Cosman, F.; de Beur, S.J.; LeBoff, M.S.; Lewiecki, E.M.; Tanner, B.; Randall, S.; Lindsay, R. Clinician’s Guide to Prevention and Treatment of Osteoporosis. Osteoporos. Int. 2014, 25, 2359–2381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, P.D.; Pannacciulli, N.; Malouf-Sierra, J.; Singer, A.; Czerwiński, E.; Bone, H.G.; Wang, C.; Huang, S.; Chines, A.; Lems, W.; et al. Efficacy and Safety of Denosumab vs. Bisphosphonates in Postmenopausal Women Previously Treated with Oral Bisphosphonates. Osteoporos. Int. 2020, 31, 181–191. [Google Scholar] [CrossRef]
- Li, X.; Ominsky, M.S.; Villasenor, K.S.; Niu, Q.-T.; Asuncion, F.J.; Xia, X.; Grisanti, M.; Wronski, T.J.; Simonet, W.S.; Ke, H.Z. Sclerostin Antibody Reverses Bone Loss by Increasing Bone Formation and Decreasing Bone Resorption in a Rat Model of Male Osteoporosis. Endocrinology 2018, 159, 260–271. [Google Scholar] [CrossRef]
- Lewiecki, E.M. Bisphosphonates for the Treatment of Osteoporosis: Insights for Clinicians. Ther. Adv. Chronic Dis. 2010, 1, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Conley, R.B.; Adib, G.; Adler, R.A.; Åkesson, K.E.; Alexander, I.M.; Amenta, K.C.; Blank, R.D.; Brox, W.T.; Carmody, E.E.; Chapman-Novakofski, K.; et al. Secondary Fracture Prevention: Consensus Clinical Recommendations from a Multistakeholder Coalition. J. Bone Miner. Res. 2020, 35, 36–52. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Shoback, D. Mechanisms Underlying Normal Fracture Healing and Risk Factors for Delayed Healing. Curr. Osteoporos. Rep. 2019, 17, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-P.; Wu, P.-K.; Lee, C.-H.; Shih, C.-M.; Chiu, Y.-C.; Hsu, C.-E. Association of Osteoporosis and Varus Inclination of the Tibial Plateau in Postmenopausal Women with Advanced Osteoarthritis of the Knee. BMC Musculoskelet. Disord. 2021, 22, 223. [Google Scholar] [CrossRef]
- Féron, J.-M.; Mauprivez, R. Fracture Repair: General Aspects and Influence of Osteoporosis and Anti-Osteoporosis Treatment. Injury 2016, 47, S10–S14. [Google Scholar] [CrossRef]
- Hak, D.J. The Biology of Fracture Healing in Osteoporosis and in the Presence of Anti-Osteoporotic Drugs. Injury 2018, 49, 1461–1465. [Google Scholar] [CrossRef]
- Kates, S.L.; Ackert-Bicknell, C.L. How Do Bisphosphonates Affect Fracture Healing? Injury 2016, 47, S65–S68. [Google Scholar] [CrossRef] [Green Version]
- Vannucci, L.; Brandi, M.L. Healing of the Bone with Anti-Fracture Drugs. Expert Opin. Pharmacother. 2016, 17, 2267–2272. [Google Scholar] [CrossRef]
- Casp, A.J.; Montgomery, S.R.; Cancienne, J.M.; Brockmeier, S.F.; Werner, B.C. Osteoporosis and Implant-Related Complications After Anatomic and Reverse Total Shoulder Arthroplasty. J. Am. Acad. Orthop. Surg. 2020, 28, 121–127. [Google Scholar] [CrossRef]
- Bernatz, J.T.; Krueger, D.C.; Squire, M.W.; Illgen, R.L.; Binkley, N.C.; Anderson, P.A. Unrecognized Osteoporosis Is Common in Patients with a Well-Functioning Total Knee Arthroplasty. J. Arthroplast. 2019, 34, 2347–2350. [Google Scholar] [CrossRef]
- Lee, C.K.; Choi, S.K.; An, S.B.; Ha, Y.; Yoon, S.-H.; Kim, I.; Kim, K.N. Influence of Osteoporosis Following Spine Surgery on Reoperation, Readmission, and Economic Costs: An 8-Year Nationwide Population-Based Study in Korea. World Neurosurg. 2021, 149, e360–e368. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Cha, T.; Schwab, J.; Fogel, H.; Tobert, D.; Razi, A.E.; Hecht, A.; Bono, C.M.; Hershman, S. Osteoporosis Increases the Likelihood of Revision Surgery Following a Long Spinal Fusion for Adult Spinal Deformity. Spine J. 2021, 21, 134–140. [Google Scholar] [CrossRef]
- Rosso, F.; Cottino, U.; Dettoni, F.; Bruzzone, M.; Bonasia, D.E.; Rossi, R. Revision Total Knee Arthroplasty (TKA): Mid-Term Outcomes and Bone Loss/Quality Evaluation and Treatment. J. Orthop. Surg. 2019, 14, 280. [Google Scholar] [CrossRef]
- Yoshiya, S. The Effects of Exercise and Sports Activities on Bone and Joint Morbidities. Clin. Calcium 2017, 27, 39–43. [Google Scholar]
- Gregg, E.W.; Cauley, J.A.; Seeley, D.G.; Ensrud, K.E.; Bauer, D.C. Physical Activity and Osteoporotic Fracture Risk in Older Women. Study of Osteoporotic Fractures Research Group. Ann. Intern. Med. 1998, 129, 81–88. [Google Scholar] [CrossRef]
- Feskanich, D.; Willett, W.; Colditz, G. Walking and Leisure-Time Activity and Risk of Hip Fracture in Postmenopausal Women. JAMA 2002, 288, 2300–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, T.E.; Shea, B.; Dawson, L.J.; Downie, F.; Murray, A.; Ross, C.; Harbour, R.T.; Caldwell, L.M.; Creed, G. Exercise for Preventing and Treating Osteoporosis in Postmenopausal Women. Cochrane Database Syst. Rev. 2011, 7, CD000333. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.; Walker, M.; Hancock, S.; Holliday, E.; Brogan, G.; Jones, M.; McEvoy, M.; Boyle, M.; Talley, N.; Attia, J. A Serological Diagnosis of Coeliac Disease Is Associated with Osteoporosis in Older Australian Adults. Nutrients 2018, 10, 849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Kurray, L.; Agnihotri, A.; Das, P.; Verma, A.K.; Sreenivas, V.; Dattagupta, S.; Makharia, G.K. Titers of Anti-Tissue Transglutaminase Antibody Correlate Well with Severity of Villous Abnormalities in Celiac Disease. J. Clin. Gastroenterol. 2015, 49, 212–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratajczak, A.E.; Rychter, A.M.; Zawada, A.; Dobrowolska, A.; Krela-Kaźmierczak, I. Nutrients in the Prevention of Osteoporosis in Patients with Inflammatory Bowel Diseases. Nutrients 2020, 12, 1702. [Google Scholar] [CrossRef] [PubMed]
- Chisari, E.; Shivappa, N.; Vyas, S. Polyphenol-Rich Foods and Osteoporosis. Curr. Pharm. Des. 2019, 25, 2459–2466. [Google Scholar] [CrossRef]
- Ampelas, D.G. Current and Former Smokers and Hip Fractures. J. Frailty Sarcopenia Falls 2018, 03, 148–154. [Google Scholar] [CrossRef]
- Hopper, J.L.; Seeman, E. The Bone Density of Female Twins Discordant for Tobacco Use. N. Engl. J. Med. 1994, 330, 387–392. [Google Scholar] [CrossRef]
- Kiel, D.P.; Baron, J.A.; Anderson, J.J.; Hannan, M.T.; Felson, D.T. Smoking Eliminates the Protective Effect of Oral Estrogens on the Risk for Hip Fracture among Women. Ann. Intern. Med. 1992, 116, 716–721. [Google Scholar] [CrossRef]
- Paschalis, E.P.; Gamsjaeger, S.; Hassler, N.; Fahrleitner-Pammer, A.; Dobnig, H.; Stepan, J.J.; Pavo, I.; Eriksen, E.F.; Klaushofer, K. Vitamin D and Calcium Supplementation for Three Years in Postmenopausal Osteoporosis Significantly Alters Bone Mineral and Organic Matrix Quality. Bone 2017, 95, 41–46. [Google Scholar] [CrossRef]
- Trang, H.M.; Cole, D.E.; Rubin, L.A.; Pierratos, A.; Siu, S.; Vieth, R. Evidence That Vitamin D3 Increases Serum 25-Hydroxyvitamin D More Efficiently than Does Vitamin D2. Am. J. Clin. Nutr. 1998, 68, 854–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.-C.; Wu, C.-C.; Liao, M.-T.; Shyu, J.-F.; Hung, C.-F.; Yen, T.-H.; Lu, C.-L.; Lu, K.-C. Role of Nutritional Vitamin D in Osteoporosis Treatment. Clin. Chim. Acta 2018, 484, 179–191. [Google Scholar] [CrossRef] [PubMed]
- George Kerry, R.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.-S.; Das, G. Benefaction of Probiotics for Human Health: A Review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bongaerts, G.P.; Severijnen, R.S. A Reassessment of the PROPATRIA Study and Its Implications for Probiotic Therapy. Nat. Biotechnol. 2016, 34, 55–63. [Google Scholar] [CrossRef]
- Drake, M.T.; Clarke, B.L.; Khosla, S. Bisphosphonates: Mechanism of Action and Role in Clinical Practice. Mayo Clin. Proc. Mayo Clin. 2008, 83, 1032–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, D.M.; Schwartz, A.V.; Ensrud, K.E.; Cauley, J.A.; Levis, S.; Quandt, S.A.; Satterfield, S.; Wallace, R.B.; Bauer, D.C.; Palermo, L.; et al. Effects of Continuing or Stopping Alendronate after 5 Years of Treatment: The Fracture Intervention Trial Long-Term Extension (FLEX): A Randomized Trial. JAMA 2006, 296, 2927–2938. [Google Scholar] [CrossRef]
- Anastasilakis, A.D.; Polyzos, S.A.; Makras, P. THERAPY OF ENDOCRINE DISEASE: Denosumab vs Bisphosphonates for the Treatment of Postmenopausal Osteoporosis. Eur. J. Endocrinol. 2018, 179, R31–R45. [Google Scholar] [CrossRef]
- Deeks, E.D. Denosumab: A Review in Postmenopausal Osteoporosis. Drugs Aging 2018, 35, 163–173. [Google Scholar] [CrossRef]
- Hanley, D.A.; Adachi, J.D.; Bell, A.; Brown, V. Denosumab: Mechanism of Action and Clinical Outcomes. Int. J. Clin. Pract. 2012, 66, 1139–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricarte, F.R.; Le Henaff, C.; Kolupaeva, V.G.; Gardella, T.J.; Partridge, N.C. Parathyroid Hormone(1–34) and Its Analogs Differentially Modulate Osteoblastic Rankl Expression via PKA/SIK2/SIK3 and PP1/PP2A–CRTC3 Signaling. J. Biol. Chem. 2018, 293, 20200–20213. [Google Scholar] [CrossRef] [Green Version]
- Kendler, D.L.; Marin, F.; Zerbini, C.A.F.; Russo, L.A.; Greenspan, S.L.; Zikan, V.; Bagur, A.; Malouf-Sierra, J.; Lakatos, P.; Fahrleitner-Pammer, A.; et al. Effects of Teriparatide and Risedronate on New Fractures in Post-Menopausal Women with Severe Osteoporosis (VERO): A Multicentre, Double-Blind, Double-Dummy, Randomised Controlled Trial. Lancet 2018, 391, 230–240. [Google Scholar] [CrossRef]
- Hodsman, A.B.; Bauer, D.C.; Dempster, D.W.; Dian, L.; Hanley, D.A.; Harris, S.T.; Kendler, D.L.; McClung, M.R.; Miller, P.D.; Olszynski, W.P.; et al. Parathyroid Hormone and Teriparatide for the Treatment of Osteoporosis: A Review of the Evidence and Suggested Guidelines for Its Use. Endocr. Rev. 2005, 26, 688–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Calle, J.; Sato, A.Y.; Bellido, T. Role and Mechanism of Action of Sclerostin in Bone. Bone 2017, 96, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaveh, S.; Hosseinifard, H.; Ghadimi, N.; Vojdanian, M.; Aryankhesal, A. Efficacy and Safety of Romosozumab in Treatment for Low Bone Mineral Density: A Systematic Review and Meta-Analysis. Clin. Rheumatol. 2020, 39, 3261–3276. [Google Scholar] [CrossRef] [PubMed]
- Pinkerton, J.V.; Conner, E.A. Beyond Estrogen: Advances in Tissue Selective Estrogen Complexes and Selective Estrogen Receptor Modulators. Climacteric 2019, 22, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Peres-Ueno, M.J.; Fernandes, F.; Brito, V.G.B.; Nicola, Â.C.; Stringhetta-Garcia, C.T.; Castoldi, R.C.; Menezes, A.P.; Ciarlini, P.C.; Louzada, M.J.Q.; Oliveira, S.H.P.; et al. Effect of Pre-Treatment of Strength Training and Raloxifene in Periestropause on Bone Healing. Bone 2020, 134, 115285. [Google Scholar] [CrossRef] [PubMed]
- Prior, J.C.; Seifert-Klauss, V.R.; Giustini, D.; Adachi, J.D.; Kalyan, S.; Goshtasebi, A. Estrogen-Progestin Therapy Causes a Greater Increase in Spinal Bone Mineral Density than Estrogen Therapy-a Systematic Review and Meta-Analysis of Controlled Trials with Direct Randomization. J. Musculoskelet. Neuronal. Interact. 2017, 17, 146. [Google Scholar]
- Levin, V.A.; Jiang, X.; Kagan, R. Estrogen Therapy for Osteoporosis in the Modern Era. Osteoporos. Int. J. Establ. Result Coop. Eur. Found. Osteoporos. Natl. Osteoporos. Found. USA 2018, 29, 1049–1055. [Google Scholar] [CrossRef]
- Imai, Y.; Youn, M.-Y.; Kondoh, S.; Nakamura, T.; Kouzmenko, A.; Matsumoto, T.; Takada, I.; Takaoka, K.; Kato, S. Estrogens Maintain Bone Mass by Regulating Expression of Genes Controlling Function and Life Span in Mature Osteoclasts. Ann. N. Y. Acad. Sci. 2009, 1173 (Suppl. S1), E31–E39. [Google Scholar] [CrossRef]
- Sun, S.; Jones, R.B.; Fodor, A.A. Inference-Based Accuracy of Metagenome Prediction Tools Varies across Sample Types and Functional Categories. Microbiome 2020, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A New Genomic Blueprint of the Human Gut Microbiota. Nature 2019, 568, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal Barrier and Gut Microbiota: Shaping Our Immune Responses throughout Life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Milani, C.; De Giori, G.S.; Sesma, F.; Van Sinderen, D.; Ventura, M. Bacteria as Vitamin Suppliers to Their Host: A Gut Microbiota Perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- Weingarden, A.R.; Vaughn, B.P. Intestinal Microbiota, Fecal Microbiota Transplantation, and Inflammatory Bowel Disease. Gut Microbes 2017, 8, 238–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.J.; Sears, C.L.; Maruthur, N. Gut Microbiome and Its Role in Obesity and Insulin Resistance. Ann. N. Y. Acad. Sci. 2020, 1461, 37–52. [Google Scholar] [CrossRef]
- Zinöcker, M.; Lindseth, I. The Western Diet–Microbiome-Host Interaction and Its Role in Metabolic Disease. Nutrients 2018, 10, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, V.J.; Miller, K.R.; Martindale, R.G. Food Insecurity, Malnutrition, and the Microbiome. Curr. Nutr. Rep. 2020, 9, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Osadchiy, V.; Martin, C.R.; Mayer, E.A. The Gut–Brain Axis and the Microbiome: Mechanisms and Clinical Implications. Clin. Gastroenterol. Hepatol. 2019, 17, 322–332. [Google Scholar] [CrossRef]
- Ge, Y.; Wang, X.; Guo, Y.; Yan, J.; Abuduwaili, A.; Aximujiang, K.; Yan, J.; Wu, M. Gut Microbiota Influence Tumor Development and Alter Interactions with the Human Immune System. J. Exp. Clin. Cancer Res. 2021, 40, 42. [Google Scholar] [CrossRef] [PubMed]
- Witkowski, M.; Weeks, T.L.; Hazen, S.L. Gut Microbiota and Cardiovascular Disease. Circ. Res. 2020, 127, 553–570. [Google Scholar] [CrossRef]
- Huttenhower, C.; Knight, R.; Brown, C.T.; Caporaso, J.G.; Clemente, J.C.; Gevers, D.; Franzosa, E.A.; Kelley, S.T.; Knights, D.; Ley, R.E.; et al. Advancing the Microbiome Research Community. Cell 2014, 159, 227–230. [Google Scholar] [CrossRef] [Green Version]
- Hills, R.; Pontefract, B.; Mishcon, H.; Black, C.; Sutton, S.; Theberge, C. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef] [Green Version]
- Overby, H.B.; Ferguson, J.F. Gut Microbiota-Derived Short-Chain Fatty Acids Facilitate Microbiota:Host Cross Talk and Modulate Obesity and Hypertension. Curr. Hypertens. Rep. 2021, 23, 8. [Google Scholar] [CrossRef] [PubMed]
- Hsu, E.; Pacifici, R. From Osteoimmunology to Osteomicrobiology: How the Microbiota and the Immune System Regulate Bone. Calcif. Tissue Int. 2018, 102, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Chen, J.; Li, X.; Sun, Q.; Qin, P.; Wang, Q. Compositional and Functional Features of the Female Premenopausal and Postmenopausal Gut Microbiota. FEBS Lett. 2019, 593, 2655–2664. [Google Scholar] [CrossRef]
- Wang, H.; Sun, Z.; Wang, Y.; Hu, Z.; Zhou, H.; Zhang, L.; Hong, B.; Zhang, S.; Cao, X. MiR-33-5p, a Novel Mechano-Sensitive MicroRNA Promotes Osteoblast Differentiation by Targeting Hmga2. Sci. Rep. 2016, 6, 23170. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; He, X.; Wei, W.; Zhou, X. MicroRNA-194 Promotes Osteoblast Differentiation via Downregulating STAT1. Biochem. Biophys. Res. Commun. 2015, 460, 482–488. [Google Scholar] [CrossRef]
- Tang, X.; Lin, J.; Wang, G.; Lu, J. MicroRNA-433-3p Promotes Osteoblast Differentiation through Targeting DKK1 Expression. PLoS ONE 2017, 12, e0179860. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Chen, W.-D.; Wang, Y.-D. The Roles of the Gut Microbiota–MiRNA Interaction in the Host Pathophysiology. Mol. Med. 2020, 26, 101. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zeng, Y.; Zeng, D.; Wang, H.; Zhou, M.; Sun, N.; Xin, J.; Khalique, A.; Rajput, D.S.; Pan, K.; et al. Probiotics and MicroRNA: Their Roles in the Host–Microbe Interactions. Front. Microbiol. 2021, 11, 604462. [Google Scholar] [CrossRef]
- Uchida, Y.; Irie, K.; Fukuhara, D.; Kataoka, K.; Hattori, T.; Ono, M.; Ekuni, D.; Kubota, S.; Morita, M. Commensal Microbiota Enhance Both Osteoclast and Osteoblast Activities. Molecules 2018, 23, 1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzer, M.; Makki, K.; Storelli, G.; Machuca-Gayet, I.; Srutkova, D.; Hermanova, P.; Martino, M.E.; Balmand, S.; Hudcovic, T.; Heddi, A.; et al. Lactobacillus Plantarum Strain Maintains Growth of Infant Mice during Chronic Undernutrition. Science 2016, 351, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Pacifici, R. Bone Remodeling and the Microbiome. Cold Spring Harb. Perspect. Med. 2018, 8, a031203. [Google Scholar] [CrossRef]
- Brüssow, H. Probiotics and Prebiotics in Clinical Tests: An Update. F1000Research 2019, 8, 1157. [Google Scholar] [CrossRef] [Green Version]
- McCabe, L.; Britton, R.A.; Parameswaran, N. Prebiotic and Probiotic Regulation of Bone Health: Role of the Intestine and Its Microbiome. Curr. Osteoporos. Rep. 2015, 13, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.-Z. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell 2018, 174, 1388–1405.e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vich Vila, A.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.R.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of Commonly Used Drugs on the Composition and Metabolic Function of the Gut Microbiota. Nat. Commun. 2020, 11, 362. [Google Scholar] [CrossRef]
- Das, M.; Cronin, O.; Keohane, D.M.; Cormac, E.M.; Nugent, H.; Nugent, M.; Molloy, C.; O’Toole, P.W.; Shanahan, F.; Molloy, M.G.; et al. Gut Microbiota Alterations Associated with Reduced Bone Mineral Density in Older Adults. Rheumatology 2019, 58, 2295–2304. [Google Scholar] [CrossRef] [Green Version]
- Branco Santos, J.C.; de Melo, J.A.; Maheshwari, S.; de Medeiros, W.M.T.Q.; de Freitas Oliveira, J.W.; Moreno, C.J.; Mario Amzel, L.; Gabelli, S.B.; Sousa Silva, M. Bisphosphonate-Based Molecules as Potential New Antiparasitic Drugs. Molecules 2020, 25, 2602. [Google Scholar] [CrossRef]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and Disease Markers Correlate with Gut Microbiome Composition across Thousands of People. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef]
- Lombard, J.; Moreira, D. Origins and Early Evolution of the Mevalonate Pathway of Isoprenoid Biosynthesis in the Three Domains of Life. Mol. Biol. Evol. 2011, 28, 87–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayakawa, H.; Sobue, F.; Motoyama, K.; Yoshimura, T.; Hemmi, H. Identification of Enzymes Involved in the Mevalonate Pathway of Flavobacterium Johnsoniae. Biochem. Biophys. Res. Commun. 2017, 487, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Kalyan, S.; Wang, J.; Quabius, E.S.; Huck, J.; Wiltfang, J.; Baines, J.F.; Kabelitz, D. Systemic Immunity Shapes the Oral Microbiome and Susceptibility to Bisphosphonate-Associated Osteonecrosis of the Jaw. J. Transl. Med. 2015, 13, 212. [Google Scholar] [CrossRef] [Green Version]
- Khafipour, A.; Eissa, N.; Munyaka, P.M.; Rabbi, M.F.; Kapoor, K.; Kermarrec, L.; Khafipour, E.; Bernstein, C.N.; Ghia, J.-E. Denosumab Regulates Gut Microbiota Composition and Cytokines in Dinitrobenzene Sulfonic Acid (DNBS)-Experimental Colitis. Front. Microbiol. 2020, 11, 1405. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.S.; Mazur, C.M.; Wein, M.N. Sclerostin and Osteocalcin: Candidate Bone-Produced Hormones. Front. Endocrinol. 2021, 12, 584147. [Google Scholar] [CrossRef] [PubMed]
- Kobza, A.O.; Papaioannou, A.; Lau, A.N.; Adachi, J.D. Romosozumab in the Treatment of Osteoporosis. Immunotherapy 2020, 12, 965–981. [Google Scholar] [CrossRef]
- Florio, M.; Gunasekaran, K.; Stolina, M.; Li, X.; Liu, L.; Tipton, B.; Salimi-Moosavi, H.; Asuncion, F.J.; Li, C.; Sun, B.; et al. A Bispecific Antibody Targeting Sclerostin and DKK-1 Promotes Bone Mass Accrual and Fracture Repair. Nat. Commun. 2016, 7, 11505. [Google Scholar] [CrossRef]
- Peng, J.; Dong, Z.; Hui, Z.; Aifei, W.; Lianfu, D.; Youjia, X. Bone Sclerostin and Dickkopf-Related Protein-1 Are Positively Correlated with Bone Mineral Density, Bone Microarchitecture, and Bone Strength in Postmenopausal Osteoporosis. BMC Musculoskelet. Disord. 2021, 22, 480. [Google Scholar] [CrossRef]
- Tyagi, A.M.; Yu, M.; Darby, T.M.; Vaccaro, C.; Li, J.-Y.; Owens, J.A.; Hsu, E.; Adams, J.; Weitzmann, M.N.; Jones, R.M.; et al. The Microbial Metabolite Butyrate Stimulates Bone Formation via T Regulatory Cell-Mediated Regulation of WNT10B Expression. Immunity 2018, 49, 1116–1131.e7. [Google Scholar] [CrossRef] [Green Version]
- D’Amelio, P.; Grimaldi, A.; Di Bella, S.; Brianza, S.Z.M.; Cristofaro, M.A.; Tamone, C.; Giribaldi, G.; Ulliers, D.; Pescarmona, G.P.; Isaia, G. Estrogen Deficiency Increases Osteoclastogenesis Up-Regulating T Cells Activity: A Key Mechanism in Osteoporosis. Bone 2008, 43, 92–100. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Piemontese, M.; Liu, Y.; Thostenson, J.D.; Xiong, J.; O’Brien, C.A. RANKL (Receptor Activator of NFκB Ligand) Produced by Osteocytes Is Required for the Increase in B Cells and Bone Loss Caused by Estrogen Deficiency in Mice. J. Biol. Chem. 2016, 291, 24838–24850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R.; Probiotic, L. Reuteri Treatment Prevents Bone Loss in a Menopausal Ovariectomized Mouse Model. J. Cell. Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.L.A.; Liu, X.; Zhao, Y.C.; Hieronymi, K.; Rossi, G.; Auvil, L.S.; Welge, M.; Bushell, C.; Smith, R.L.; Carlson, K.E.; et al. Long-Term Administration of Conjugated Estrogen and Bazedoxifene Decreased Murine Fecal β-Glucuronidase Activity Without Impacting Overall Microbiome Community. Sci. Rep. 2018, 8, 8166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papageorgiou, M.; Biver, E. Interactions of the Microbiome with Pharmacological and Non-Pharmacological Approaches for the Management of Ageing-Related Musculoskeletal Diseases. Ther. Adv. Musculoskelet. Dis. 2021, 13, 1759720X2110090. [Google Scholar] [CrossRef]
- Li, J.-Y.; Chassaing, B.; Tyagi, A.M.; Vaccaro, C.; Luo, T.; Adams, J.; Darby, T.M.; Weitzmann, M.N.; Mulle, J.G.; Gewirtz, A.T.; et al. Sex Steroid Deficiency–Associated Bone Loss Is Microbiota Dependent and Prevented by Probiotics. J. Clin. Investig. 2016, 126, 2049–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacifici, R. Role of Gut Microbiota in the Skeletal Response to PTH. J. Clin. Endocrinol. Metab. 2021, 106, 636–645. [Google Scholar] [CrossRef]
- Gaboriau-Routhiau, V.; Rakotobe, S.; Lécuyer, E.; Mulder, I.; Lan, A.; Bridonneau, C.; Rochet, V.; Pisi, A.; De Paepe, M.; Brandi, G.; et al. The Key Role of Segmented Filamentous Bacteria in the Coordinated Maturation of Gut Helper T Cell Responses. Immunity 2009, 31, 677–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Malik Tyagi, A.; Li, J.-Y.; Adams, J.; Denning, T.L.; Weitzmann, M.N.; Jones, R.M.; Pacifici, R. PTH Induces Bone Loss via Microbial-Dependent Expansion of Intestinal TNF+ T Cells and Th17 Cells. Nat. Commun. 2020, 11, 468. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.D.; Hattersley, G.; Riis, B.J.; Williams, G.C.; Lau, E.; Russo, L.A.; Alexandersen, P.; Zerbini, C.A.F.; Hu, M.; Harris, A.G.; et al. Effect of Abaloparatide vs Placebo on New Vertebral Fractures in Postmenopausal Women with Osteoporosis: A Randomized Clinical Trial. JAMA 2016, 316, 722. [Google Scholar] [CrossRef]
- Saffouri, G.B.; Shields-Cutler, R.R.; Chen, J.; Yang, Y.; Lekatz, H.R.; Hale, V.L.; Cho, J.M.; Battaglioli, E.J.; Bhattarai, Y.; Thompson, K.J.; et al. Small Intestinal Microbial Dysbiosis Underlies Symptoms Associated with Functional Gastrointestinal Disorders. Nat. Commun. 2019, 10, 2012. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Boudreau, D.M.; Freedman, A.N. Trends in the Use of Aspirin and Nonsteroidal Anti-Inflammatory Drugs in the General U.S. Population. Pharmacoepidemiol. Drug Saf. 2014, 23, 43–50. [Google Scholar] [CrossRef]
- Lisowska, B.; Kosson, D.; Domaracka, K. Positives and Negatives of Nonsteroidal Anti-Inflammatory Drugs in Bone Healing: The Effects of These Drugs on Bone Repair. Drug Des. Devel. Ther. 2018, 12, 1809–1814. [Google Scholar] [CrossRef] [PubMed]
- Herbenick, M.A.; Sprott, D.; Stills, H.; Lawless, M. Effects of a Cyclooxygenase 2 Inhibitor on Fracture Healing in a Rat Model. Am. J. Orthop. 2008, 37, E133–E137. [Google Scholar] [PubMed]
- Simon, A.M.; Manigrasso, M.B.; O’Connor, J.P. Cyclo-Oxygenase 2 Function Is Essential for Bone Fracture Healing. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2002, 17, 963–976. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.-K.; Li, C.-J.; Liao, H.-J.; Wang, C.-K.; Wang, G.-J.; Ho, M.-L. Anti-Inflammatory Drugs Suppress Proliferation and Induce Apoptosis through Altering Expressions of Cell Cycle Regulators and pro-Apoptotic Factors in Cultured Human Osteoblasts. Toxicology 2009, 258, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Nagano, A.; Arioka, M.; Takahashi-Yanaga, F.; Matsuzaki, E.; Sasaguri, T. Celecoxib Inhibits Osteoblast Maturation by Suppressing the Expression of Wnt Target Genes. J. Pharmacol. Sci. 2017, 133, 18–24. [Google Scholar] [CrossRef]
- Marlicz, W.; Loniewski, I.; Grimes, D.S.; Quigley, E.M. Nonsteroidal Anti-Inflammatory Drugs, Proton Pump Inhibitors, and Gastrointestinal Injury: Contrasting Interactions in the Stomach and Small Intestine. Mayo Clin. Proc. 2014, 89, 1699–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maseda, D.; Ricciotti, E. NSAID–Gut Microbiota Interactions. Front. Pharmacol. 2020, 11, 1153. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.M.; Aronoff, D.M. The Influence of Nonsteroidal Anti-Inflammatory Drugs on the Gut Microbiome. Clin. Microbiol. Infect. 2016, 22, 178.e1–178.e9. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Battaglia, T.; Aleman, J.O.; Walker, J.M.; Blaser, M.J.; Holt, P.R. Celecoxib Does Not Alter Intestinal Microbiome in a Longitudinal Diet-Controlled Study. Clin. Microbiol. Infect. 2016, 22, 464–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thangamani, S.; Younis, W.; Seleem, M.N. Repurposing Celecoxib as a Topical Antimicrobial Agent. Front. Microbiol. 2015, 6, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Sanabria, E.; Heiremans, E.; Calatayud Arroyo, M.; Props, R.; Leclercq, L.; Snoeys, J.; Van de Wiele, T. Short-Term Supplementation of Celecoxib-Shifted Butyrate Production on a Simulated Model of the Gut Microbial Ecosystem and Ameliorated in Vitro Inflammation. NPJ Biofilms Microbiomes 2020, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edogawa, S.; Peters, S.A.; Jenkins, G.D.; Gurunathan, S.V.; Sundt, W.J.; Johnson, S.; Lennon, R.J.; Dyer, R.B.; Camilleri, M.; Kashyap, P.C.; et al. Sex Differences in NSAID-Induced Perturbation of Human Intestinal Barrier Function and Microbiota. FASEB J. 2018, 32, 6615–6625. [Google Scholar] [CrossRef]
- Persson, P.; Sisask, G.; Nilsson, O. Indomethacin Inhibits Bone Formation in Inductive Allografts but Not in Autografts. Acta Orthop. 2005, 76, 465–469. [Google Scholar] [CrossRef]
- Wang, J.-W.; Kuo, C.-H.; Kuo, F.-C.; Wang, Y.-K.; Hsu, W.-H.; Yu, F.-J.; Hu, H.-M.; Hsu, P.-I.; Wang, J.-Y.; Wu, D.-C. Fecal Microbiota Transplantation: Review and Update. J. Formos. Med. Assoc. Taiwan Yi Zhi 2019, 118 (Suppl. S1), S23–S31. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seely, K.D.; Kotelko, C.A.; Douglas, H.; Bealer, B.; Brooks, A.E. The Human Gut Microbiota: A Key Mediator of Osteoporosis and Osteogenesis. Int. J. Mol. Sci. 2021, 22, 9452. https://doi.org/10.3390/ijms22179452
Seely KD, Kotelko CA, Douglas H, Bealer B, Brooks AE. The Human Gut Microbiota: A Key Mediator of Osteoporosis and Osteogenesis. International Journal of Molecular Sciences. 2021; 22(17):9452. https://doi.org/10.3390/ijms22179452
Chicago/Turabian StyleSeely, Kevin D., Cody A. Kotelko, Hannah Douglas, Brandon Bealer, and Amanda E. Brooks. 2021. "The Human Gut Microbiota: A Key Mediator of Osteoporosis and Osteogenesis" International Journal of Molecular Sciences 22, no. 17: 9452. https://doi.org/10.3390/ijms22179452
APA StyleSeely, K. D., Kotelko, C. A., Douglas, H., Bealer, B., & Brooks, A. E. (2021). The Human Gut Microbiota: A Key Mediator of Osteoporosis and Osteogenesis. International Journal of Molecular Sciences, 22(17), 9452. https://doi.org/10.3390/ijms22179452