
Characteristics of Ventricular Electrophysiological Substrates in Metabolic Mice Treated with Empagliflozin

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Metabolic Profiles of the Study Groups
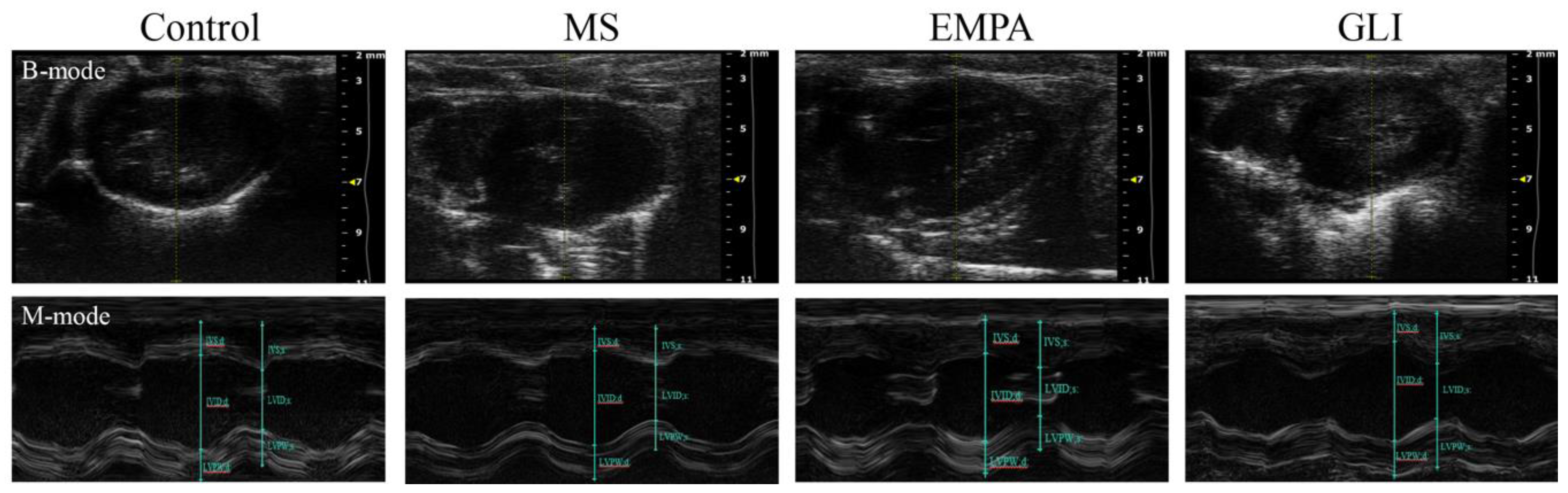
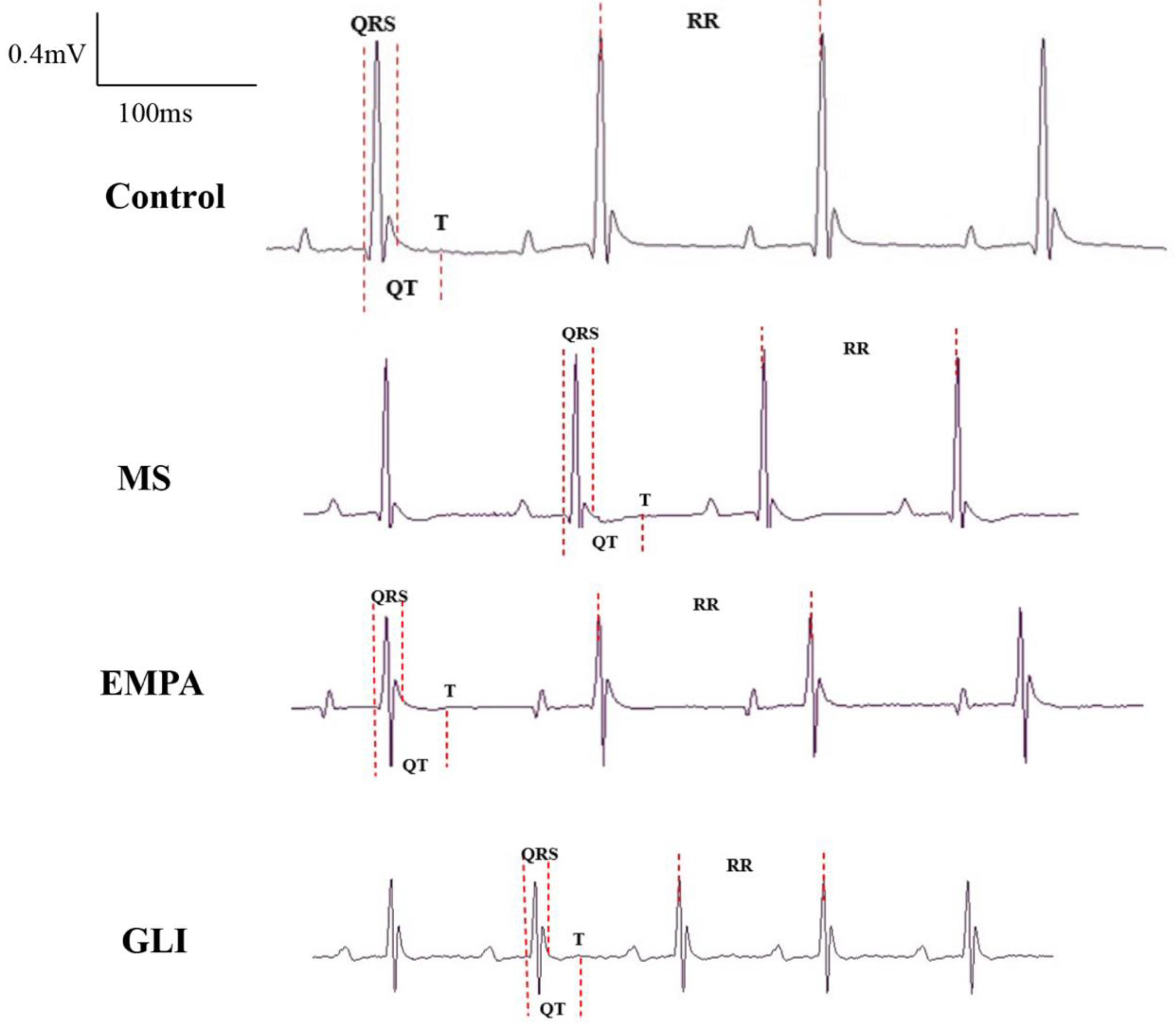
2.2. Characteristics of the Echocardiography and Electrocardiography (ECG) in the Ventricles of the Study Groups
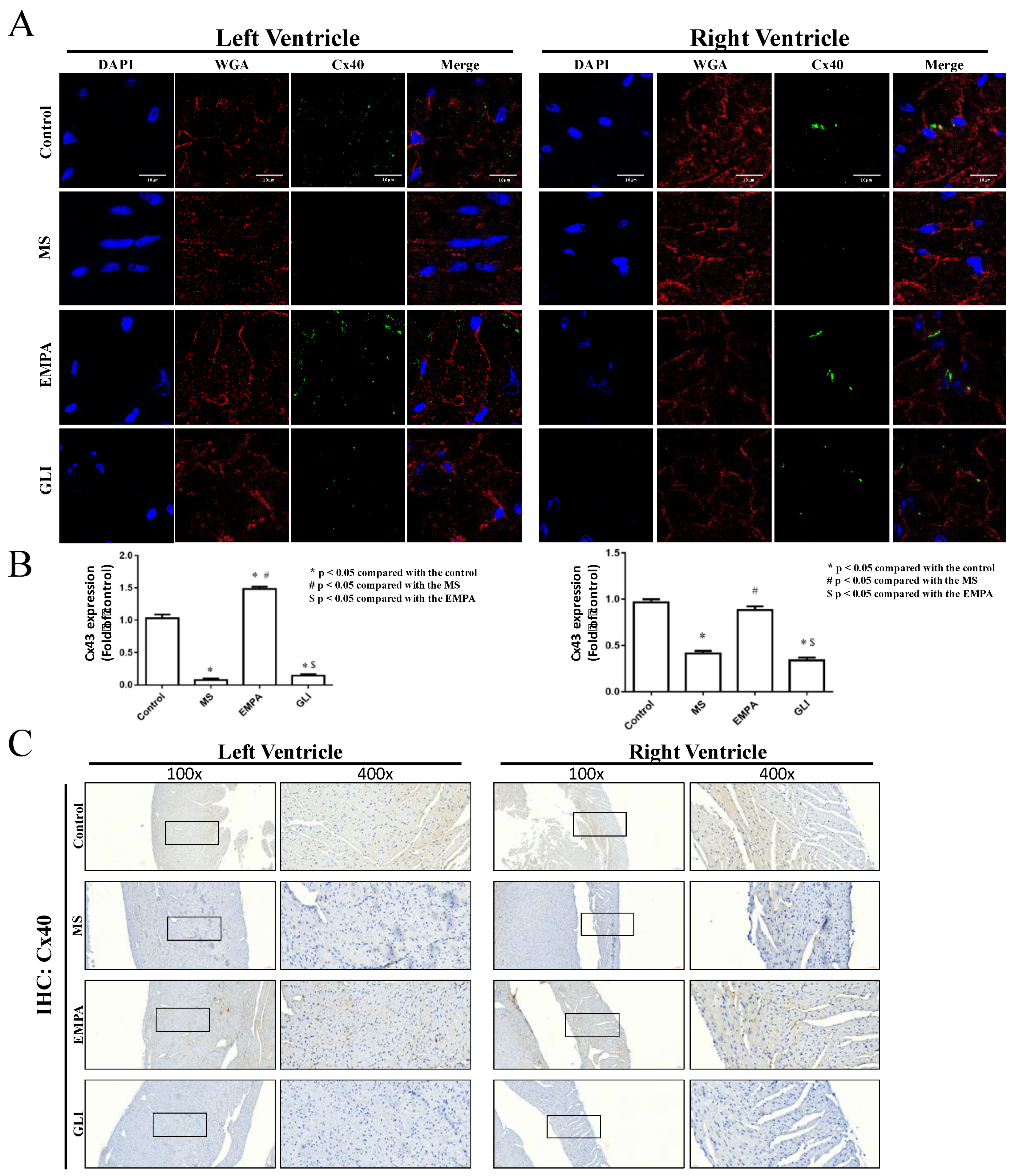
2.3. Expression of Connexin 40 (Cx40) and Connexin 43 (Cx43) in Cardiac Ventricles
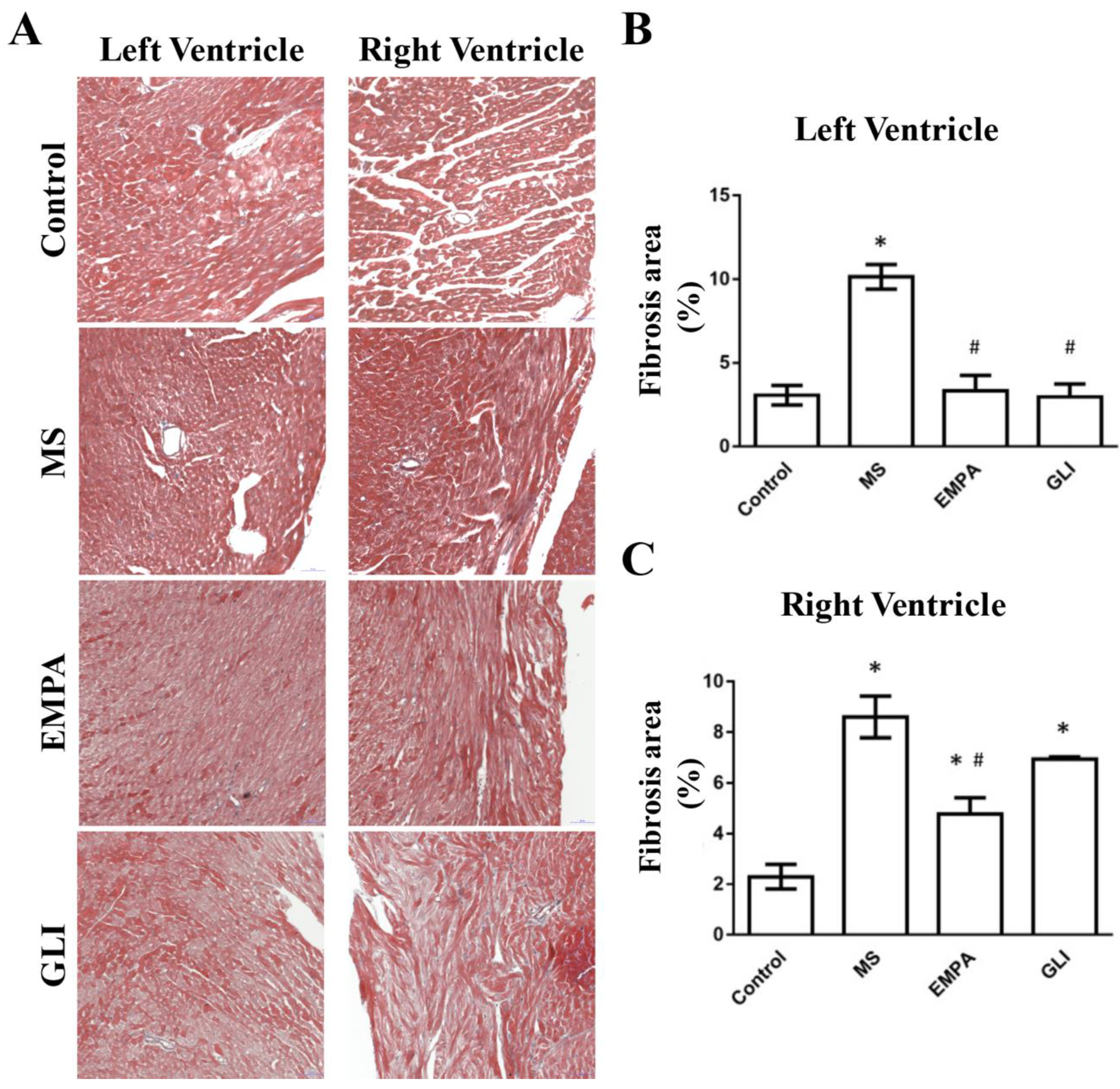
2.4. Fibrotic Areas of the Study Groups
3. Discussion
3.1. General Discussion
3.2. The Change of Electrophysiology and Electrocardiography in the Metabolic Syndrome
3.3. The Change in Electrophysiology and Electrocardiography after Sulfonylurea Treatment
3.4. The Change of Electrophysiology and Electrocardiography after EMPA Treatment
3.5. The Changes in Cx40 and Cx43 Expression in Ventricular Tissue after EMPA Treatment
3.6. The Changes in Fibrotic Areas of Ventricular Tissue after EMPA Treatment
3.7. Limitations
4. Materials and Methods
4.1. Study Animal Preparation
4.2. Characteristics of ECG and Echocardiography in the Ventricles
4.3. Determination of the ERP of the RV and LV by Langendorff Heart Perfusion
4.4. Ventricle Expression of Connexin 40 and Connexin 43
4.5. Histological and Immunohistochemistry (IHC) Staining
4.6. The Measurement of Fibrotic Area in Ventricles
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asghar, O.; Alam, U.; Hayat, S.A.; Aghamohammadzadeh, R.; Heagerty, A.; Malik, R.A. Obesity, Diabetes and Atrial Fibrillation; Epidemiology, Mechanisms and Interventions. Curr. Cardiol. Rev. 2012, 8, 253–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzella, D.; Barbieri, M.; Rizzo, M.R.; Ragno, E.; Passariello, N.; Gambardella, A.; Marfella, R.; Giugliano, D.; Paolisso, G. Role of free fatty acids on cardiac autonomic nervous system in noninsulin-dependent diabetic patients: Effects of metabolic control. J. Clin. Endocrinol. Metab. 2001, 86, 2769–2774. [Google Scholar] [CrossRef]
- Gallo, L.A.; Wright, E.M.; Vallon, V. Probing SGLT2 as a therapeutic target for diabetes: Basic physiology and consequences. Diabetes Vasc. Dis. Res. 2015, 12, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Zinman, B.; Wanner, C.; Lachin, J.M.; Fitchett, D.H.; Bluhmki, E.; Hantel, S.; Mattheus, M.; Devins, T.; Johansen, O.E.; Woerle, H.J.; et al. Empagliflozin, Cardiovascular Outcomes, and Mortality in Type 2 Diabetes. N. Engl. J. Med. 2015, 373, 2117–2128. [Google Scholar] [CrossRef]
- McMurray, J.J.; Solomon, S.D.; Inzucchi, S.E.; Køber, L.; Kosiborod, M.N.; Martinez, F.A.; Ponikowski, P.; Sabatine, M.S.; Anand, I.S.; Bělohlávek, J. Dapagliflozin in patients with heart failure and reduced ejection fraction. N. Engl. J. Med. 2019, 381, 1995–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Aizawa, Y.; Yuasa, S.; Kishi, S.; Fuse, K.; Fujita, S.; Ikeda, Y.; Kitazawa, H.; Takahashi, M.; Sato, M.; et al. The effect of dapagliflozin treatment on epicardial adipose tissue volume. Cardiovasc. Diabetol. 2018, 17, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inzucchi, S.E.; Zinman, B.; Wanner, C.; Ferrari, R.; Fitchett, D.; Hantel, S.; Espadero, R.-M.; Woerle, H.-J.; Broedl, U.C.; Johansen, O.E. SGLT-2 inhibitors and cardiovascular risk: Proposed pathways and review of ongoing outcome trials. Diabetes Vasc. Dis. Res. 2015, 12, 90–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Q.; Meng, L.; Lee, S.; Tse, G.; Gong, M.; Zhang, Z.; Zhao, J.; Zhao, Y.; Li, G.; Liu, T. Empagliflozin, a sodium glucose co-transporter-2 inhibitor, alleviates atrial remodeling and improves mitochondrial function in high-fat diet/streptozotocin-induced diabetic rats. Cardiovasc. Diabetol. 2019, 18, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ring, A.; Brand, T.; Macha, S.; Breithaupt-Groegler, K.; Simons, G.; Walter, B.; Woerle, H.J.; Broedl, U.C. The sodium glucose cotransporter 2 inhibitor empagliflozin does not prolong QT interval in a thorough QT (TQT) study. Cardiovasc. Diabetol. 2013, 12, 70. [Google Scholar] [CrossRef] [Green Version]
- Jhuo, S.-J.; Liu, I.-H.; Tsai, W.-C.; Chou, T.-W.; Lin, Y.-H.; Wu, B.-N.; Lee, K.-T.; Lai, W.-T. Effects of Secretome from Fat Tissues on Ion Currents of Cardiomyocyte Modulated by Sodium-Glucose Transporter 2 Inhibitor. Molecules 2020, 25, 3606. [Google Scholar] [CrossRef]
- Tam, W.-C.; Lin, Y.-K.; Chan, W.-P.; Huang, J.-H.; Hsieh, M.-H.; Chen, S.-A.; Chen, Y.-J. Pericardial Fat Is Associated with the Risk of Ventricular Arrhythmia in Asian Patients. Circ. J. 2016, 80, 1726–1733. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.; Zhang, S.; Yang, D.; Gao, L.; Lin, Y.; Chu, Z.; Jiang, X.; Yin, X.; Zheng, Z.; Wei, X.; et al. Effect of Epicardial Fat Pad Ablation on Acute Atrial Electrical Remodeling and Inducibility of Atrial Fibrillation. Circ. J. 2010, 74, 885–894. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-T.; Tang, P.W.-H.; Tsai, W.-C.; Liu, I.-H.; Yen, H.-W.; Voon, W.-C.; Wu, B.-N.; Sheu, S.-H.; Lai, W.-T. Differential Effects of Central and Peripheral Fat Tissues on the Delayed Rectifier K + Outward Currents in Cardiac Myocytes. Cardiology 2013, 125, 118–124. [Google Scholar] [CrossRef]
- Wong, C.X.; Ganesan, A.; Selvanayagam, J.B. Epicardial fat and atrial fibrillation: Current evidence, potential mechanisms, clinical implications, and future directions. Eur. Heart J. 2016, 38, 1294–1302. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, J.; Gomez, J.F.; Martinez-Mateu, L.; Romero, L.; Saiz, J.; Trenor, B. Heterogeneous Effects of Fibroblast-Myocyte Coupling in Different Regions of the Human Atria Under Conditions of Atrial Fibrillation. Front. Physiol. 2019, 10, 847. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Bai, Y.; Sun, K.; Xue, H.; Wang, Y.; Song, X.; Fan, X.; Song, H.; Han, Y.; Hui, R. Patients with Metabolic Syndrome Have Prolonged Corrected QT Interval (QTc). Clin. Cardiol. 2009, 32, E93–E99. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, Y.; Tu, D.; Liu, X.; Niu, S.; Suo, Y.; Liu, T.; Li, G.; Liu, C. Role of NLRP3-Inflammasome/Caspase-1/Galectin-3 Pathway on Atrial Remodeling in Diabetic Rabbits. J. Cardiovasc. Transl. Res. 2020, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Brady, P.A.; Terzic, A. The sulfonylurea controversy: More questions from the heart. J. Am. Coll. Cardiol. 1998, 31, 950–956. [Google Scholar] [CrossRef] [Green Version]
- Hayami, N.; Sekiguchi, A.; Iwasaki, Y.-K.; Murakawa, Y.; Yamashita, T. No Additional Effect of DPP-4 Inhibitor on Preventing Atrial Fibrosis in Streptozotocin-Induced Diabetic Rat as Compared with Sulfonylurea. Int. Heart J. 2016, 57, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Leonard, C.E.; Brensinger, C.M.; Aquilante, C.L.; Bilker, W.B.; Boudreau, D.M.; Deo, R.; Flory, J.H.; Gagne, J.J.; Mangaali, M.J.; Hennessy, S. Comparative Safety of Sulfonylureas and the Risk of Sudden Cardiac Arrest and Ventricular Arrhythmia. Diabetes Care 2018, 41, 713–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosati, B.; Rocchetti, M.; Zaza, A.; Wanke, E. Sulfonylureas blockade of neural and cardiac HERG channels. FEBS Lett. 1998, 440, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Leonard, C.E.; Hennessy, S.; Han, X.; Siscovick, D.S.; Flory, J.H.; Deo, R. Pro- and Antiarrhythmic Actions of Sulfonylureas: Mechanistic and Clinical Evidence. Trends Endocrinol. Metab. 2017, 28, 561–586. [Google Scholar] [CrossRef]
- Halkin, A.; Roth, A.; Jonas, M.; Behar, S. Sulfonylureas Are Not Associated with Increased Mortality in Diabetics Treated with Thrombolysis for Acute Myocardial Infarction. J. Thromb. Thrombolysis 2001, 12, 177–184. [Google Scholar] [CrossRef]
- Holman, R.R.; Paul, S.; Bethel, M.A.; Matthews, D.R.; Neil, H.A.W. 10-Year Follow-up of Intensive Glucose Control in Type 2 Diabetes. N. Engl. J. Med. 2008, 359, 1577–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garvey, W.T.; Van Gaal, L.; Leiter, L.A.; Vijapurkar, U.; List, J.; Cuddihy, R.; Ren, J.; Davies, M.J. Effects of canagliflozin versus glimepiride on adipokines and inflammatory biomarkers in type 2 diabetes. Metablism 2018, 85, 32–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najeed, S.A.; Khan, I.A.; Molnar, J.; Somberg, J.C. Differential effect of glyburide (glibenclamide) and metformin on QT dispersion: A potential adenosine triphosphate sensitive K+ channel effect. Am. J. Cardiol. 2002, 90, 1103–1106. [Google Scholar] [CrossRef]
- Epifantseva, I.; Shaw, R.M. Intracellular trafficking pathways of Cx43 gap junction channels. Biochim. Biophys. Acta (BBA)-Biomembr. 2018, 1860, 40–47. [Google Scholar] [CrossRef]
- Pellman, J.; Zhang, J.; Sheikh, F. Myocyte-fibroblast communication in cardiac fibrosis and arrhythmias: Mechanisms and model systems. J. Mol. Cell. Cardiol. 2016, 94, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Sawaya, S.E.; Rajawat, Y.S.; Rami, T.G.; Szalai, G.; Price, R.L.; Sivasubramanian, N.; Mann, D.; Khoury, D.S. Downregulation of connexin40 and increased prevalence of atrial arrhythmias in transgenic mice with cardiac-restricted overexpression of tumor necrosis factor. Am. J. Physiol. Circ. Physiol. 2007, 292, H1561–H1567. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Novak, S.M.; Cover, C.; Wang, A.A.; Chinyere, I.R.; Juneman, E.B.; Zarnescu, D.C.; Wong, P.K.; Gregorio, C.C. Increased Cardiac Arrhythmogenesis Associated with Gap Junction Remodeling with Upregulation of RNA-Binding Protein FXR1. Circulation 2018, 137, 605–618. [Google Scholar] [CrossRef]
- Lee, H.-C.; Chen, C.-C.; Tsai, W.-C.; Lin, H.-T.; Shiao, Y.-L.; Sheu, S.-H.; Wu, B.-N.; Chen, C.-H.; Lai, W.-T. Very-Low-Density Lipoprotein of Metabolic Syndrome Modulates Gap Junctions and Slows Cardiac Conduction. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kirubakaran, S.; Chowdhury, R.A.; Hall, M.C.; Patel, P.M.; Garratt, C.J.; Peters, N.S. Fractionation of electrograms is caused by colocalized conduction block and connexin disorganization in the absence of fibrosis as AF becomes persistent in the goat model. Heart Rhythm. 2015, 12, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalliah, C.J.; Bell, J.R.; Raaijmakers, A.J.; Waddell, H.M.; Wells, S.P.; Bernasochi, G.B.; Montgomery, M.K.; Binny, S.; Watts, T.; Joshi, S.B.; et al. Epicardial Adipose Tissue Accumulation Confers Atrial Conduction Abnormality. J. Am. Coll. Cardiol. 2020, 76, 1197–1211. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, G.; Li, M.; Li, Y.; Yang, L.; Fu, Q.; Tian, Y. High Glucose Downregulates Connexin 43 Expression and Its Gap Junction and Hemichannel Function in Osteocyte-like MLO-Y4 Cells Through Activation of the p38MAPK/ERK Signal Pathway. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 545–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzerini, P.E.; Laghi-Pasini, F.; Acampa, M.; Srivastava, U.; Bertolozzi, I.; Giabbani, B.; Finizola, F.; Vanni, F.; Dokollari, A.; Natale, M.; et al. Systemic Inflammation Rapidly Induces Reversible Atrial Electrical Remodeling: The Role of Interleukin-6–Mediated Changes in Connexin Expression. J. Am. Heart Assoc. 2019, 8, e011006. [Google Scholar] [CrossRef] [PubMed]
- Ladeiras-Lopes, R.; Moreira, H.T.; Bettencourt, N.; Carvalho, R.F.; Sampaio, F.; Ambale-Venkatesh, B.; Wu, C.; Liu, K.; Bertoni, A.G.; Ouyang, P.; et al. Metabolic Syndrome Is Associated with Impaired Diastolic Function Independently of MRI-Derived Myocardial Extracellular Volume: The MESA Study. Diabetes 2018, 67, 1007–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdy, O.; Porramatikul, S.; Al-Ozairi, E. Metabolic obesity: The paradox between visceral and subcutaneous fat. Curr. Diabetes Rev. 2006, 2, 367–373. [Google Scholar]
- Eryd, S.A.; Smith, J.G.; Melander, O.; Hedblad, B.; Engström, G. Inflammation-sensitive proteins and risk of atrial fibrillation: A population-based cohort study. Eur. J. Epidemiol. 2011, 26, 449–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.-F.; Yeh, H.-I.; Tsao, H.-M.; Tai, C.-T.; Lin, Y.-J.; Chang, S.-L.; Lo, L.-W.; Tuan, T.-C.; Tzeng, C.-H.; Huang, S.-H.; et al. Impact of circulating monocyte CD36 level on atrial fibrillation and subsequent catheter ablation. Heart Rhythm. 2011, 8, 650–656. [Google Scholar] [CrossRef]
- Li, C.; Zhang, J.; Xue, M.; Li, X.; Han, F.; Liu, X.; Xu, L.; Lu, Y.; Cheng, Y.; Li, T.; et al. SGLT2 inhibition with empagliflozin attenuates myocardial oxidative stress and fibrosis in diabetic mice heart. Cardiovasc. Diabetol. 2019, 18, 15. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | MS | EMPA | GLI | |
|---|---|---|---|---|
| Weight (g) | 29.58 ± 0.46 | 44.62 ± 3.13 * | 37.08 ± 3.30 *,# | 44.46 ± 3.05 *,† |
| Fasting glucose (mg/dL) | 164.10 ± 34.35 | 232.72 ± 48.4 * | 200.23 ± 13.74 | 211.62 ± 28.66 |
| Cholesterol (mg/dL) | 47.5 ± 2.65 | 115.60 ± 13.09 * | 99.75 ± 15.69 * | 133.00 ± 4.24 *,#,† |
| LDL (mg/dL) | 2.55 ± 0.17 | 5.35 ± 0.29 * | 5.12 ± 0.95 * | 7.15 ± 1.84 * |
| HDL (mg/dL) | 20.45 ± 2.37 | 45.12 ± 3.59 * | 33.63 ± 9.52 *,# | 51.53 ± 2.01 *,#,† |
| Triglyceride (mg/dL) | 11.50 ± 1.73 | 20.80 ± 2.95 * | 17.25 ± 8.02 | 19.00 ± 2.31 * |
| Control | MS | EMPA | GLI | |
|---|---|---|---|---|
| RVDd (mm) | 1.08 ± 0.26 | 1.03 ± 0.08 | 0.94 ± 0.14 | 0.96 ± 0.16 |
| IVSd (mm) | 0.67 ± 0.06 | 0.72 ± 0.06 | 0.74 ± 0.08 | 0.66 ± 0.03 |
| LVIDd (mm) | 2.81 ± 0.28 | 2.85 ± 0.09 | 2.73 ± 0.21 | 2.76 ± 0.12 |
| LVPWd (mm) | 0.79 ± 0.02 | 0.85 ± 0.16 | 0.81 ± 0.06 | 0.79 ± 0.09 |
| IVSs (mm) | 0.84 ± 0.09 | 0.96 ± 0.09 | 0.90 ± 0.06 | 0.98 ± 0.11 |
| LVIDs (mm) | 1.81 ± 0.22 | 1.67 ± 0.30 | 1.63 ± 0.17 | 1.75 ± 0.34 |
| LVPWs (mm) | 0.84 ± 0.13 | 1.00 ± 0.17 | 0.98 ± 0.07 | 0.90 ± 0.14 |
| Ejection fraction (%) | 58.39 ± 6.93 | 64.71 ± 12.94 | 64.31 ± 2.97 | 59.23 ± 13.06 |
| Control | MS | EMPA | GLI | |
|---|---|---|---|---|
| QRS wave duration (ms) | 24.27 ± 2.04 | 23.73 ± 2.55 | 22.13 ± 2.24 | 23.33 ± 3.83 |
| RR interval (ms) | 144.06 ± 14.87 | 146.67 ± 21.05 | 140.40 ± 23.18 | 142.73 ± 17.91 |
| QT interval (ms) | 53.66 ± 2.32 | 57.06 ± 3.43 | 50.00 ± 2.62 # | 56.87 ± 4.04 |
| QTc interval (ms) | 142.29 ± 9.58 | 149.47 ± 8.25 | 134.15 ± 6.72 | 150.86 ± 9.47 |
| ERP of right ventricle (ms) | 35.00 ± 10.00 | 40.00 ± 14.14 | 47.50 ± 5.00 | 48.00 ± 13.04 |
| ERP of left ventricle (ms) | 35.00 ± 10.00 | 38.00 ± 4.47 | 20.00 ± 10.00 | 60.00 ± 10.00 † |
| Induced VT/VF mice | 1 | 0 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jhuo, S.-J.; Liu, I.-H.; Tasi, W.-C.; Chou, T.-W.; Lin, Y.-H.; Wu, B.-N.; Lee, K.-T.; Lai, W.-T. Characteristics of Ventricular Electrophysiological Substrates in Metabolic Mice Treated with Empagliflozin. Int. J. Mol. Sci. 2021, 22, 6105. https://doi.org/10.3390/ijms22116105
Jhuo S-J, Liu I-H, Tasi W-C, Chou T-W, Lin Y-H, Wu B-N, Lee K-T, Lai W-T. Characteristics of Ventricular Electrophysiological Substrates in Metabolic Mice Treated with Empagliflozin. International Journal of Molecular Sciences. 2021; 22(11):6105. https://doi.org/10.3390/ijms22116105
Chicago/Turabian StyleJhuo, Shih-Jie, I-Hsin Liu, Wei-Chung Tasi, Te-Wu Chou, Yi-Hsiung Lin, Bin-Nan Wu, Kun-Tai Lee, and Wen-Ter Lai. 2021. "Characteristics of Ventricular Electrophysiological Substrates in Metabolic Mice Treated with Empagliflozin" International Journal of Molecular Sciences 22, no. 11: 6105. https://doi.org/10.3390/ijms22116105
APA StyleJhuo, S.-J., Liu, I.-H., Tasi, W.-C., Chou, T.-W., Lin, Y.-H., Wu, B.-N., Lee, K.-T., & Lai, W.-T. (2021). Characteristics of Ventricular Electrophysiological Substrates in Metabolic Mice Treated with Empagliflozin. International Journal of Molecular Sciences, 22(11), 6105. https://doi.org/10.3390/ijms22116105