Different Sensitivity of Control and MICU1- and MICU2-Ablated Trypanosoma cruzi Mitochondrial Calcium Uniporter Complex to Ruthenium-Based Inhibitors



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
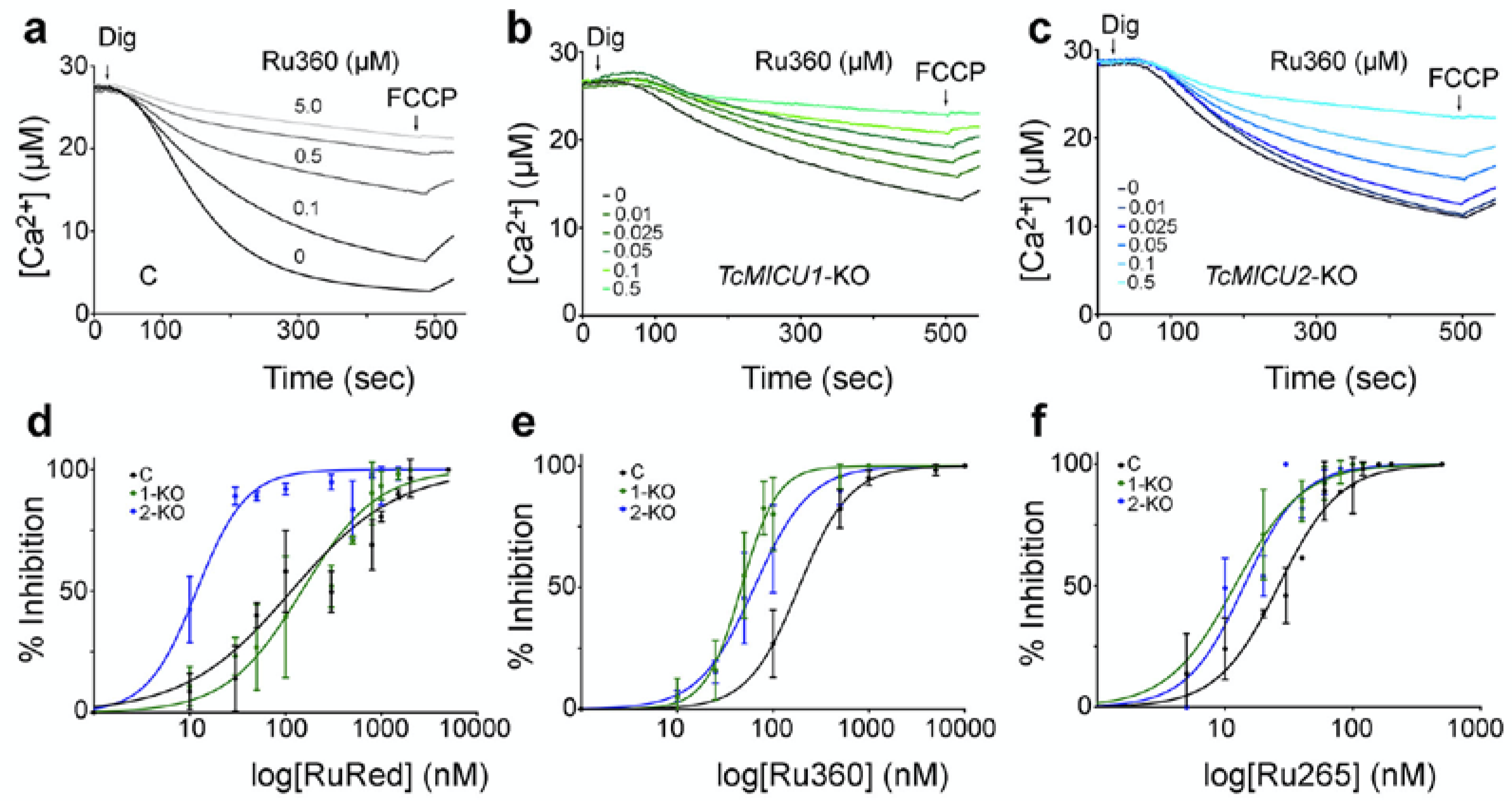
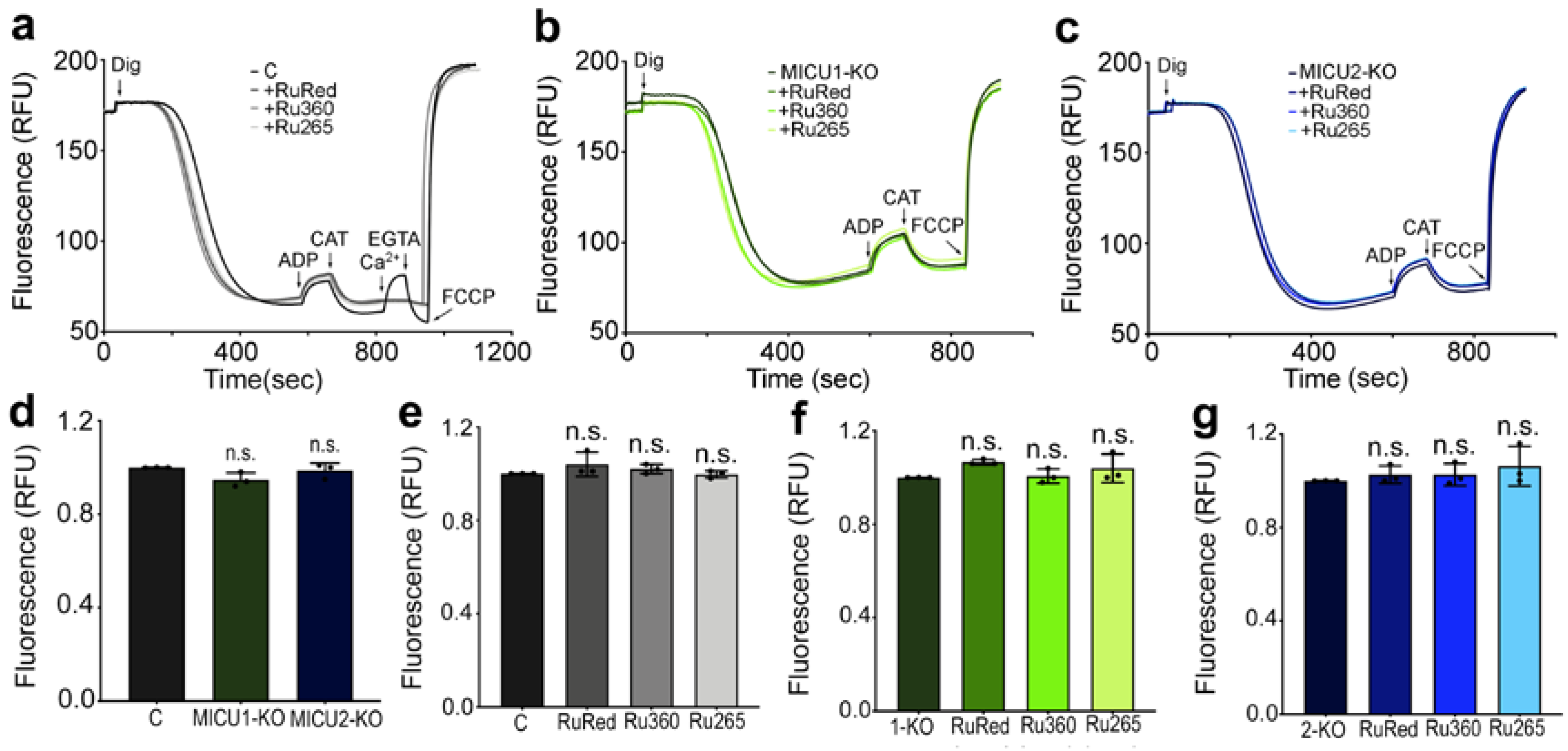
2.1. Inhibition of Mitochondrial Ca2+ Uptake by Ruthenium-Based Compounds
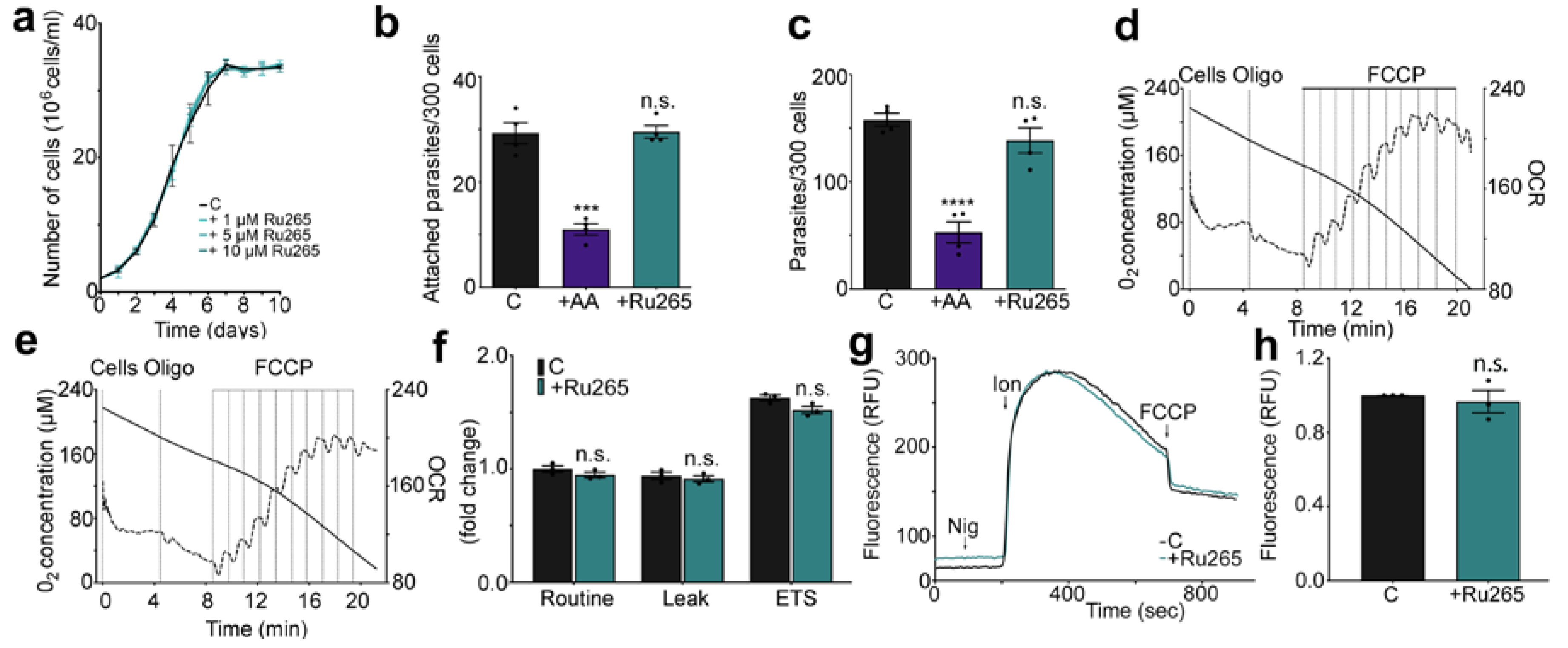
2.2. Effects of Ru265 on Trypanosoma cruzi
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Culture Methods
4.3. Ca2+ Uptake by Digitonin-Permeabilized T. cruzi Epimastigotes
4.4. Mitochondrial Membrane Potential
4.5. Cellular Respiration
4.6. Measurement of Mitochondrial Ca2+ Uptake in Intact Epimastigotes Using Rhod-2 AM
4.7. Attachment Assay
4.8. In Vitro Infection Assay
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MCU | Mitochondrial calcium uniporter |
| MICU | Mitochondrial calcium uptake |
| MCUR1 | MCU regulator 1 |
| RuRed | Ruthenium red |
References
- McCormack, J.G.; Denton, R.M. Ca2+ as a second messenger within mitochondria. Trends Biochem. Sci. 1986, 11, 258–262. [Google Scholar] [CrossRef]
- Denton, R.M. Regulation of mitochondrial dehydrogenases by calcium ions. Biochim. Biophys. Acta 2009, 1787, 1309–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Territo, P.R.; French, S.A.; Dunleavy, M.C.; Evans, F.J.; Balaban, R.S. Calcium activation of heart mitochondrial oxidative phosphorylation: Rapid kinetics of mVO2, NADH, AND light scattering. J. Biol. Chem. 2001, 276, 2586–2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Territo, P.R.; Mootha, V.K.; French, S.A.; Balaban, R.S. Ca2+ activation of heart mitochondrial oxidative phosphorylation: Role of the F0/F1-ATPase. Am. J. Physiol. Cell Physiol. 2000, 278, C423–C435. [Google Scholar] [CrossRef]
- Deluca, H.F.; Engstrom, G.W. Calcium uptake by rat kidney mitochondria. Proc. Natl. Acad. Sci. USA 1961, 47, 1744–1750. [Google Scholar] [CrossRef] [Green Version]
- Vasington, F.D.; Murphy, J.V. Ca ion uptake by rat kidney mitochondria and its dependence on respiration and phosphorylation. J. Biol. Chem. 1962, 237, 2670–2677. [Google Scholar]
- De Stefani, D.; Raffaello, A.; Teardo, E.; Szabo, I.; Rizzuto, R. A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature 2011, 476, 336–340. [Google Scholar] [CrossRef]
- Baughman, J.M.; Perocchi, F.; Girgis, H.S.; Plovanich, M.; Belcher-Timme, C.A.; Sancak, Y.; Bao, X.R.; Strittmatter, L.; Goldberger, O.; Bogorad, R.L.; et al. Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature 2011, 476, 341–345. [Google Scholar] [CrossRef] [Green Version]
- Docampo, R.; Vercesi, A.E. Characteristics of Ca2+ transport by Trypanosoma cruzi mitochondria in situ. Arch Biochem. Biophys. 1989, 272, 122–129. [Google Scholar] [CrossRef]
- Docampo, R.; Vercesi, A.E. Ca2+ transport by coupled Trypanosoma cruzi mitochondria in situ. J. Biol. Chem. 1989, 264, 108–111. [Google Scholar]
- Carafoli, E.; Balcavage, W.X.; Lehninger, A.L.; Mattoon, J.R. Ca2+ metabolism in yeast cells and mitochondria. Biochim. Biophys. Acta 1970, 205, 18–26. [Google Scholar] [CrossRef]
- Perocchi, F.; Gohil, V.M.; Girgis, H.S.; Bao, X.R.; McCombs, J.E.; Palmer, A.E.; Mootha, V.K. MICU1 encodes a mitochondrial EF hand protein required for Ca2+ uptake. Nature 2010, 467, 291–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallilankaraman, K.; Cardenas, C.; Doonan, P.J.; Chandramoorthy, H.C.; Irrinki, K.M.; Golenar, T.; Csordas, G.; Madireddi, P.; Yang, J.; Muller, M.; et al. MCUR1 is an essential component of mitochondrial Ca2+ uptake that regulates cellular metabolism. Nat. Cell Biol. 2012, 14, 1336–1343. [Google Scholar] [CrossRef] [Green Version]
- Plovanich, M.; Bogorad, R.L.; Sancak, Y.; Kamer, K.J.; Strittmatter, L.; Li, A.A.; Girgis, H.S.; Kuchimanchi, S.; De Groot, J.; Speciner, L.; et al. MICU2, a paralog of MICU1, resides within the mitochondrial uniporter complex to regulate calcium handling. PLoS ONE 2013, 8, e55785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raffaello, A.; De Stefani, D.; Sabbadin, D.; Teardo, E.; Merli, G.; Picard, A.; Checchetto, V.; Moro, S.; Szabo, I.; Rizzuto, R. The mitochondrial calcium uniporter is a multimer that can include a dominant-negative pore-forming subunit. Embo J. 2013, 32, 2362–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancak, Y.; Markhard, A.L.; Kitami, T.; Kovacs-Bogdan, E.; Kamer, K.J.; Udeshi, N.D.; Carr, S.A.; Chaudhuri, D.; Clapham, D.E.; Li, A.A.; et al. EMRE is an essential component of the mitochondrial calcium uniporter complex. Science 2013, 342, 1379–1382. [Google Scholar] [CrossRef] [Green Version]
- Chiurillo, M.A.; Lander, N.; Bertolini, M.S.; Vercesi, A.E.; Docampo, R. Functional analysis and importance for host cell infection of the Ca2+-conducting subunits of the mitochondrial calcium uniporter of Trypanosoma cruzi. Mol. Biol. Cell 2019, 30, 1676–1690. [Google Scholar] [CrossRef]
- Huang, G.; Docampo, R. The mitochondrial Ca2+ uniporter complex (MCUC) of Trypanosoma brucei is a hetero-oligomer that contains novel subunits essential for Ca2+ uptake. mBio 2018, 9, e01700-18. [Google Scholar] [CrossRef] [Green Version]
- Chiurillo, M.A.; Lander, N.; Bertolini, M.S.; Storey, M.; Vercesi, A.E.; Docampo, R. Different roles of mitochondrial calcium uniporter complex subunits in growth and infectivity of Trypanosoma cruzi. mBio 2017, 8, e00574-17. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, M.S.; Chiurillo, M.A.; Lander, N.; Vercesi, A.E.; Docampo, R. MICU1 and MICU2 play an essential role in mitochondrial Ca2+ uptake, growth, andinfectivity of the human pathogen Trypanosoma cruzi. MBio 2019, 10, e00348-19. [Google Scholar] [CrossRef] [Green Version]
- Mallilankaraman, K.; Doonan, P.; Cardenas, C.; Chandramoorthy, H.C.; Muller, M.; Miller, R.; Hoffman, N.E.; Gandhirajan, R.K.; Molgo, J.; Birnbaum, M.J.; et al. MICU1 is an essential gatekeeper for MCU-mediated mitochondrial Ca2+ uptake that regulates cell survival. Cell 2012, 151, 630–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csordas, G.; Golenar, T.; Seifert, E.L.; Kamer, K.J.; Sancak, Y.; Perocchi, F.; Moffat, C.; Weaver, D.; de la Fuente Perez, S.; Bogorad, R.; et al. MICU1 controls both the threshold and cooperative activation of the mitochondrial Ca2+ uniporter. Cell Metab. 2013, 17, 976–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamer, K.J.; Grabarek, Z.; Mootha, V.K. High-affinity cooperative Ca2+ binding by MICU1-MICU2 serves as an on-off switch for the uniporter. Embo Rep. 2017, 18, 1397–1411. [Google Scholar] [CrossRef] [PubMed]
- Pittis, A.A.; Goh, V.; Cebrian-Serrano, A.; Wettmarshausen, J.; Perocchi, F.; Gabaldon, T. Discovery of EMRE in fungi resolves the true evolutionary history of the mitochondrial calcium uniporter. Nat. Commun. 2020, 11, 4031. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.L.; Emerson, J.; Clarke, M.J.; Sanadi, D.R. Inhibition of mitochondrial calcium ion transport by an oxo-bridged dinuclear ruthenium ammine complex. Biochemistry 1991, 30, 4949–4952. [Google Scholar] [CrossRef]
- Arduino, D.M.; Wettmarshausen, J.; Vais, H.; Navas-Navarro, P.; Cheng, Y.; Leimpek, A.; Ma, Z.; Delrio-Lorenzo, A.; Giordano, A.; Garcia-Perez, C.; et al. Systematic identification of MCU modulators by orthogonal interspecies chemical screening. Mol. Cell 2017, 67, 711–723.e7. [Google Scholar] [CrossRef]
- Cao, C.; Wang, S.; Cui, T.; Su, X.C.; Chou, J.J. Ion and inhibitor binding of the double-ring ion selectivity filter of the mitochondrial calcium uniporter. Proc. Natl. Acad. Sci. USA 2017, 114, E2846–E2851. [Google Scholar] [CrossRef] [Green Version]
- Baradaran, R.; Wang, C.; Siliciano, A.F.; Long, S.B. Cryo-EM structures of fungal and metazoan mitochondrial calcium uniporters. Nature 2018, 559, 580–584. [Google Scholar] [CrossRef]
- Paillard, M.; Csordas, G.; Huang, K.T.; Varnai, P.; Joseph, S.K.; Hajnoczky, G. MICU1 interacts with the D-Ring of the MCU pore to control its Ca2+ flux and sensitivity to Ru360. Mol. Cell 2018, 72, 778–785.e3. [Google Scholar] [CrossRef] [Green Version]
- Woods, J.J.; Nemani, N.; Shanmughapriya, S.; Kumar, A.; Zhang, M.; Nathan, S.R.; Thomas, M.; Carvalho, E.; Ramachandran, K.; Srikantan, S.; et al. A Selective and cell-permeable mitochondrial calcium uniporter (MCU) inhibitor preserves mitochondrial bioenergetics after hypoxia/reoxygenation injury. ACS Cent. Sci. 2019, 5, 153–166. [Google Scholar] [CrossRef]
- Schenkman, S.; Robbins, E.S.; Nussenzweig, V. Attachment of Trypanosoma cruzi to mammalian cells requires parasite energy, and invasion can be independent of the target cell cytoskeleton. Infect. Immun. 1991, 59, 645–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnaiger, E. Polarographic oxygen sensors, the oxygraph, and high-resolution respirometry to asses mitochondrial function. In Drug-Induced Mitochondrial Disfunction; Dykens, J., Will, Y., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2008; pp. 327–352. [Google Scholar]
- Docampo, R.; Scott, D.A.; Vercesi, A.E.; Moreno, S.N. Intracellular Ca2+ storage in acidocalcisomes of Trypanosoma cruzi. Biochem. J. 1995, 310, 1005–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, C.L. Specific inhibition of mitochondrial Ca++ transport by ruthenium red. Biochem. Biophys. Res. Commun. 1971, 42, 298–305. [Google Scholar] [CrossRef]
- Chamberlain, B.K.; Volpe, P.; Fleischer, S. Inhibition of calcium-induced calcium release from purified cardiac sarcoplasmic reticulum vesicles. J. Biol. Chem. 1984, 259, 7547–7553. [Google Scholar] [PubMed]
- Novorolsky, R.J.; Nichols, M.; Kim, J.S.; Pavlov, E.V.; Woods, J.W.; Wilson, J.J.; Robertson, G.S. The cell-permeable mitochondrial calcium uniporter inhibitor Ru265 preserves cortical neuron respiration after lethal oxygen glucose deprivation and reduces hypoxic/ischemic brain injury. J. Cereb. Blood Flow Metab. 2020, 40, 1172–1181. [Google Scholar] [CrossRef]
- Franco da Silveira, J.; Colli, W. Chemical composition of the plasma membrane from epimastigote forms of Trypanosoma cruzi. Biochim. Biophys. Acta 1981, 644, 341–350. [Google Scholar] [CrossRef]
- Rodrigues, C.O.; Catisti, R.; Uyemura, S.A.; Vercesi, A.E.; Lira, R.; Rodriguez, C.; Urbina, J.A.; Docampo, R. The sterol composition of Trypanosoma cruzi changes after growth in different culture media and results in different sensitivity to digitonin-permeabilization. J. Eukaryot. Microbiol. 2001, 48, 588–594. [Google Scholar] [CrossRef]
- Bone, G.J.; Steinert, M. Isotopes incorporated in the nucleic acids of Trypanosoma mega. Nature 1956, 178, 308–309. [Google Scholar] [CrossRef]
- Schmatz, D.M.; Murray, P.K. Cultivation of Trypanosoma cruzi in irradiated muscle cells: Improved synchronization and enhanced trypomastigote production. Parasitology 1982, 85, 115–125. [Google Scholar] [CrossRef]
- Chweih, H.; Castilho, R.F.; Figueira, T.R. Tissue and sex specificities in Ca2+ handling by isolated mitochondria in conditions avoiding the permeability transition. Exp. Physiol. 2015, 100, 1073–1092. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertolini, M.S.; Docampo, R. Different Sensitivity of Control and MICU1- and MICU2-Ablated Trypanosoma cruzi Mitochondrial Calcium Uniporter Complex to Ruthenium-Based Inhibitors. Int. J. Mol. Sci. 2020, 21, 9316. https://doi.org/10.3390/ijms21239316
Bertolini MS, Docampo R. Different Sensitivity of Control and MICU1- and MICU2-Ablated Trypanosoma cruzi Mitochondrial Calcium Uniporter Complex to Ruthenium-Based Inhibitors. International Journal of Molecular Sciences. 2020; 21(23):9316. https://doi.org/10.3390/ijms21239316
Chicago/Turabian StyleBertolini, Mayara S., and Roberto Docampo. 2020. "Different Sensitivity of Control and MICU1- and MICU2-Ablated Trypanosoma cruzi Mitochondrial Calcium Uniporter Complex to Ruthenium-Based Inhibitors" International Journal of Molecular Sciences 21, no. 23: 9316. https://doi.org/10.3390/ijms21239316
APA StyleBertolini, M. S., & Docampo, R. (2020). Different Sensitivity of Control and MICU1- and MICU2-Ablated Trypanosoma cruzi Mitochondrial Calcium Uniporter Complex to Ruthenium-Based Inhibitors. International Journal of Molecular Sciences, 21(23), 9316. https://doi.org/10.3390/ijms21239316