Release of Cholecystokinin from Rat Intestinal Mucosal Cells and the Enteroendocrine Cell Line STC-1 in Response to Maleic and Succinic Acid, Fermentation Products of Alcoholic Beverages

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
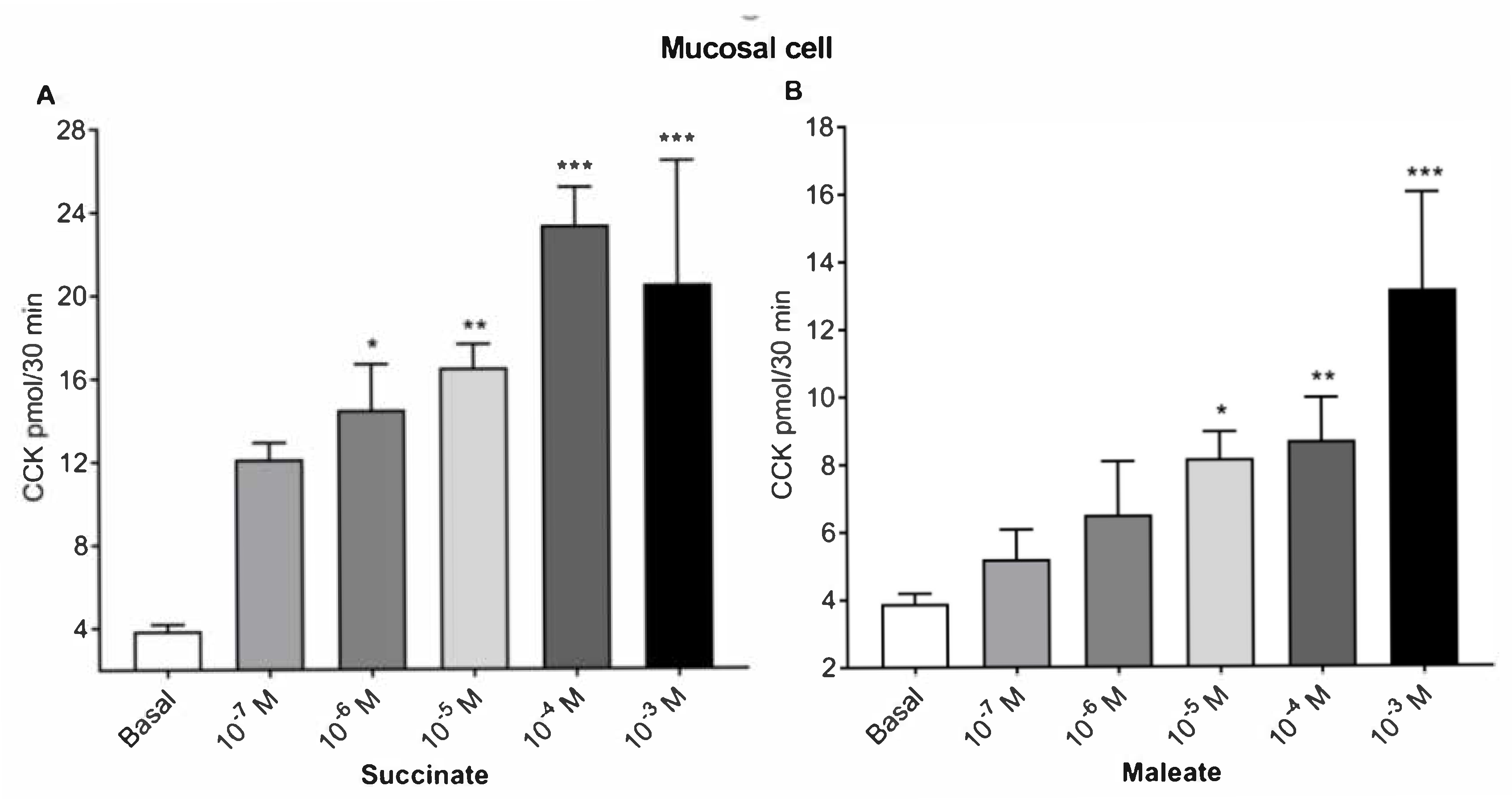
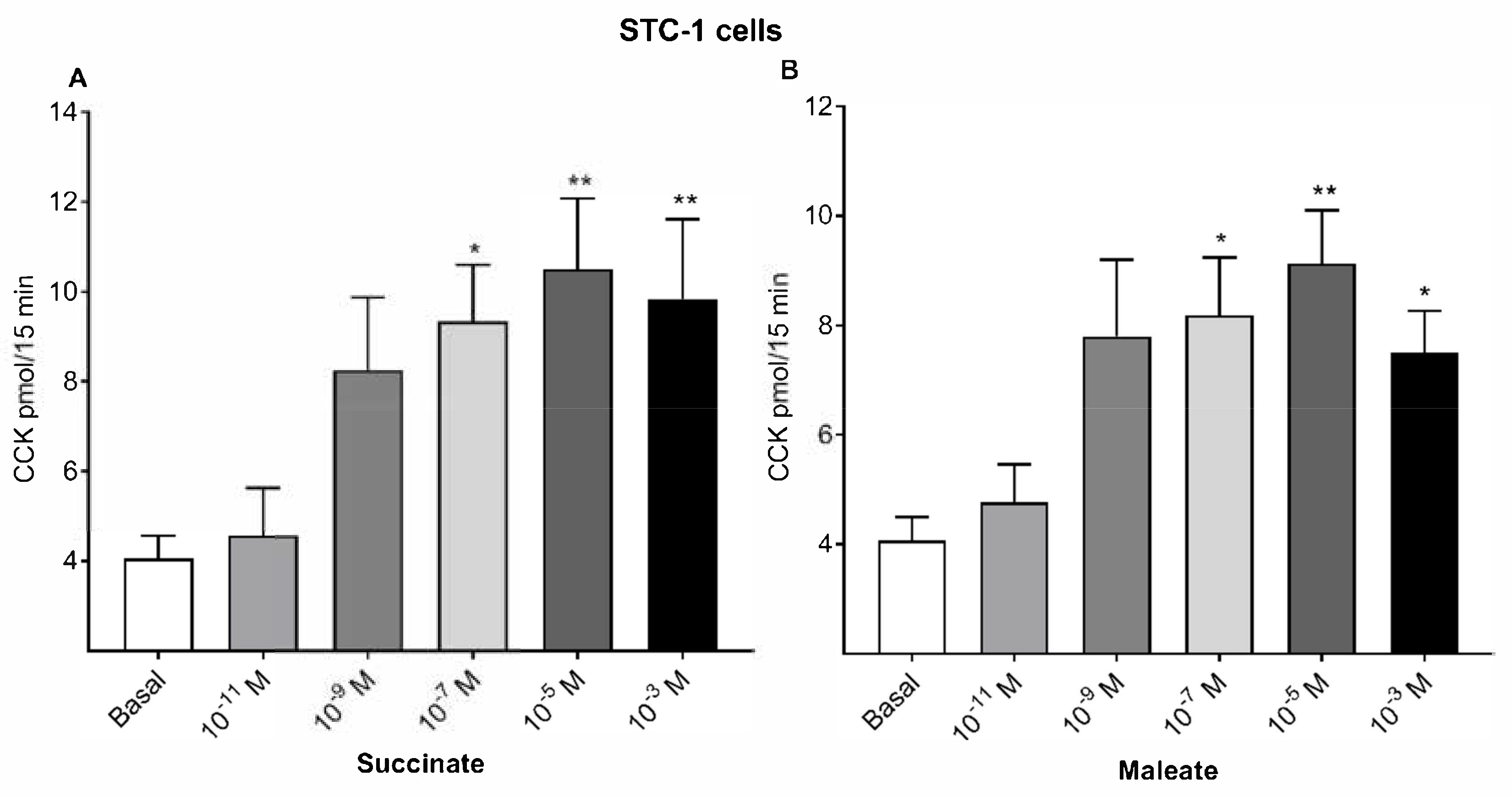
2.1. Succinate and Maleate Stimulate CCK Secretions from Mucosal Cells and STC-1 Cells
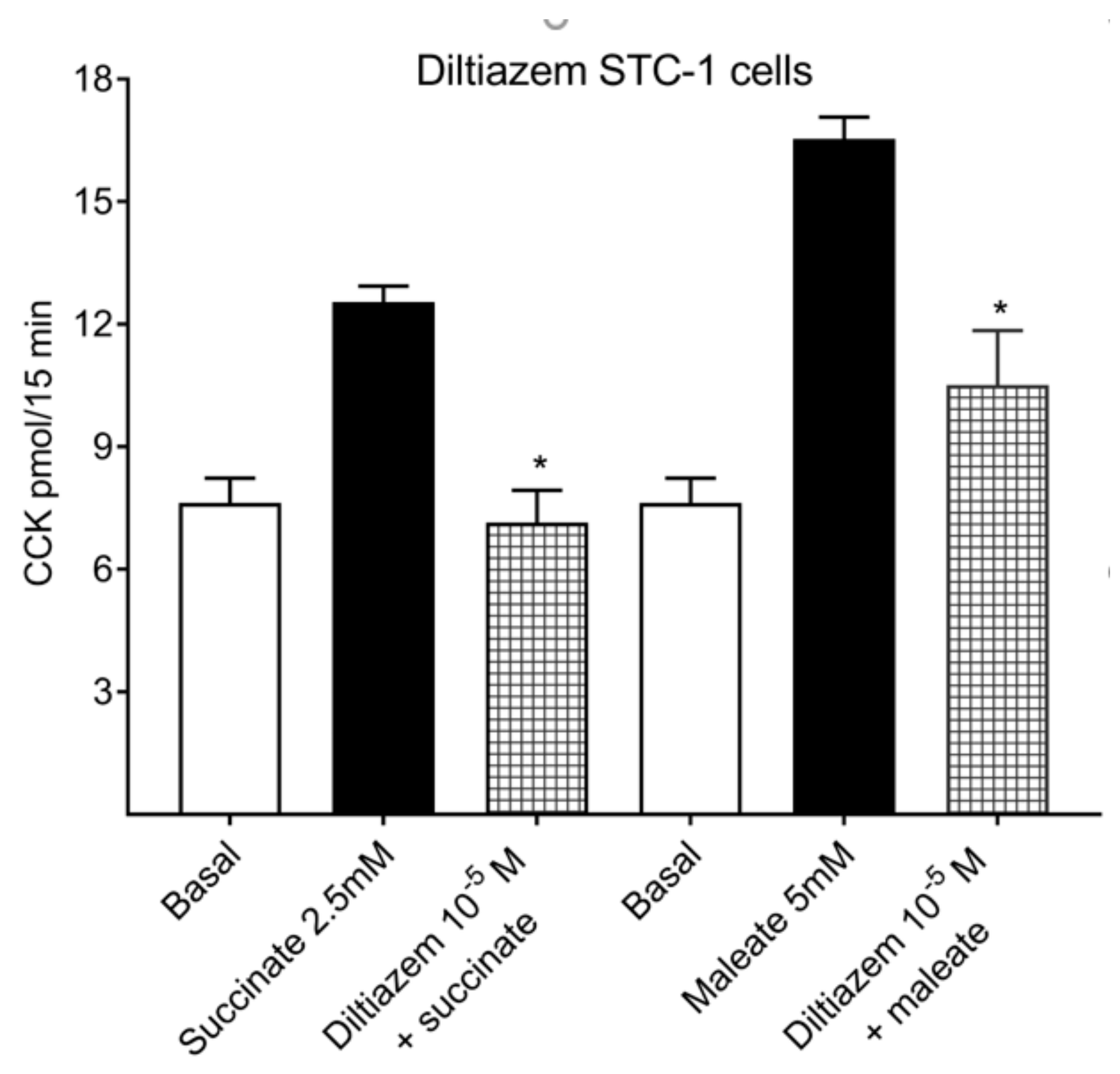
2.2. Calcium Channel Inhibitor: Diltiazem Inhibits Succinate and Maleate-Stimulated CCK Releases
2.3. Lactate Dehydrogenase (LDH) Release with Succinate and Maleate for Cytotoxicity
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Intestinal Mucosa Cell Perfusions
4.3. Cell Cultures
4.4. Rat Pancreatic Acini for CCK Bioassay
4.5. CCK Measurements by RIA
4.6. Lactate Dehydrogenase (LDH) Release from STC-1 Cells
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Choi, S.; Lee, M.; Shiu, A.L.; Yo, S.J.; Hallden, G.; Aponte, G.W. GPR93 Activation by Protein Hydrolysate Induces CCK Transcription and Secretion in STC-1 Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1366–G1375. [Google Scholar] [CrossRef]
- Herzig, K.H.; Brunke, G.; Schon, I.; Schaffer, M.; Folsch, U.R. Mechanism of Galanin’s Inhibitory Action on Pancreatic Enzyme Secretion: Modulation of Cholinergic Transmission—Studies in Vivo and in Vitro. Gut 1993, 34, 1616–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamakura, R.; Kovalainen, M.; Riikonen, J.; Nissinen, T.; Shere Raza, G.; Walkowiak, J.; Lehto, V.P.; Herzig, K.H. Inorganic Mesoporous Particles for Controlled Alpha-Linolenic Acid Delivery to Stimulate GLP-1 Secretion in Vitro. Eur. J. Pharm. Biopharm. 2019, 144, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Gukovsky, I.; Gukovskaya, A.S.; Blinman, T.A.; Zaninovic, V.; Pandol, S.J. Early NF-kappaB Activation is Associated with Hormone-Induced Pancreatitis. Am. J. Physiol. 1998, 275, G1402–G1414. [Google Scholar] [PubMed]
- Sarles, H. Chronic Pancreatitis and Diabetes. Baillieres Clin. Endocrinol. Metab. 1992, 6, 745–775. [Google Scholar] [CrossRef]
- Spanier, B.W.; Dijkgraaf, M.G.; Bruno, M.J. Epidemiology, Aetiology and Outcome of Acute and Chronic Pancreatitis: An Update. Best Pract. Res. Clin. Gastroenterol. 2008, 22, 45–63. [Google Scholar] [CrossRef]
- Chowdhury, P.; Gupta, P. Pathophysiology of Alcoholic Pancreatitis: An Overview. World J. Gastroenterol. 2006, 12, 7421–7427. [Google Scholar] [CrossRef]
- Strate, T.; Yekebas, E.; Knoefel, W.T.; Bloechle, C.; Izbicki, J.R. Pathogenesis and the Natural Course of Chronic Pancreatitis. Eur. J. Gastroenterol. Hepatol. 2002, 14, 929–934. [Google Scholar] [CrossRef]
- Chari, S.T.; Harder, H.; Teyssen, S.; Knodel, C.; Riepl, R.L.; Singer, M.V. Effect of Beer, Yeast-Fermented Glucose, and Ethanol on Pancreatic Enzyme Secretion in Healthy Human Subjects. Dig. Dis. Sci. 1996, 41, 1216–1224. [Google Scholar] [CrossRef]
- Sand, J.; Lankisch, P.G.; Nordback, I. Alcohol Consumption in Patients with Acute or Chronic Pancreatitis. Pancreatology 2007, 7, 147–156. [Google Scholar] [CrossRef]
- Bayer, M.; Rudick, J.; Lieber, C.S.; Janowitz, H.D. Inhibitory Effect of Ethanol on Canine Exocrine Pancreatic Secretion. Gastroenterology 1972, 63, 619–626. [Google Scholar] [CrossRef]
- Kristiansen, L.; Gronbaek, M.; Becker, U.; Tolstrup, J.S. Risk of Pancreatitis According to Alcohol Drinking Habits: A Population-Based Cohort Study. Am. J. Epidemiol. 2008, 168, 932–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajnal, F.; Flores, M.C.; Valenzuela, J.E. Effect of Alcohol and Alcoholic Beverages on Nonstimulated Pancreatic Secretion in Humans. Pancreas 1989, 4, 486–491. [Google Scholar] [CrossRef]
- Gerloff, A.; Singer, M.V.; Feick, P. Beer and its Non-Alcoholic Compounds: Role in Pancreatic Exocrine Secretion, Alcoholic Pancreatitis and Pancreatic Carcinoma. Int. J. Environ. Res. Public Health 2010, 7, 1093–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajnal, F.; Flores, M.C.; Radley, S.; Valenzuela, J.E. Effect of Alcohol and Alcoholic Beverages on Meal-Stimulated Pancreatic Secretion in Humans. Gastroenterology 1990, 98, 191–196. [Google Scholar] [CrossRef]
- Gerloff, A.; Singer, M.V.; Feick, P. Beer but Not Wine, Hard Liquors, or Pure Ethanol Stimulates Amylase Secretion of Rat Pancreatic Acinar Cells in Vitro. Alcohol. Clin. Exp. Res. 2009, 33, 1545–1554. [Google Scholar] [CrossRef]
- Teyssen, S.; Lenzing, T.; Gonzalez-Calero, G.; Korn, A.; Riepl, R.L.; Singer, M.V. Alcoholic Beverages Produced by Alcoholic Fermentation but Not by Distillation are Powerful Stimulants of Gastric Acid Secretion in Humans. Gut 1997, 40, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Teyssen, S.; Gonzalez-Calero, G.; Schimiczek, M.; Singer, M.V. Maleic Acid and Succinic Acid in Fermented Alcoholic Beverages are the Stimulants of Gastric Acid Secretion. J. Clin. Investig. 1999, 103, 707–713. [Google Scholar] [CrossRef]
- Sadagopan, N.; Li, W.; Roberds, S.L.; Major, T.; Preston, G.M.; Yu, Y.; Tones, M.A. Circulating Succinate is Elevated in Rodent Models of Hypertension and Metabolic Disease. Am. J. Hypertens. 2007, 20, 1209–1215. [Google Scholar]
- Kushnir, M.M.; Komaromy-Hiller, G.; Shushan, B.; Urry, F.M.; Roberts, W.L. Analysis of Dicarboxylic Acids by Tandem Mass Spectrometry. High-Throughput Quantitative Measurement of Methylmalonic Acid in Serum, Plasma, and Urine. Clin. Chem. 2001, 47, 1993–2002. [Google Scholar]
- He, W.; Miao, F.J.; Lin, D.C.; Schwandner, R.T.; Wang, Z.; Gao, J.; Chen, J.L.; Tian, H.; Ling, L. Citric Acid Cycle Intermediates as Ligands for Orphan G-Protein-Coupled Receptors. Nature 2004, 429, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Mangel, A.W. Electrophysiology of Intestinal Cholecystokinin Secretion. Regul. Pept. 1995, 56, 121–129. [Google Scholar] [CrossRef]
- Larsson, K.P.; Akerman, K.E.; Magga, J.; Uotila, S.; Kukkonen, J.P.; Nasman, J.; Herzig, K.H. The STC-1 Cells Express Functional Orexin-A Receptors Coupled to CCK Release. Biochem. Biophys. Res. Commun. 2003, 309, 209–216. [Google Scholar] [CrossRef]
- Purhonen, A.K.; Louhivuori, L.M.; Kiehne, K.; Kerman, K.E.; Herzig, K.H. TRPA1 Channel Activation Induces Cholecystokinin Release via Extracellular Calcium. FEBS Lett. 2008, 582, 229–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, O.H. Calcium Signal Compartmentalization. Biol. Res. 2002, 35, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Herzig, K.H. Cholecystokinin- and Secretin-Releasing Peptides in the Intestine—A New Regulatory Interendocrine Mechanism in the Gastrointestinal Tract. Regul. Pept. 1998, 73, 89–94. [Google Scholar] [CrossRef]
- Rehfeld, J.F.; Johnsen, A.H.; Odum, L.; Bardram, L.; Schifter, S.; Scopsi, L. Non-Sulphated Cholecystokinin in Human Medullary Thyroid Carcinomas. J. Endocrinol. 1990, 124, 501–506. [Google Scholar] [CrossRef]
- Larsson, L.I.; Rehfeld, J.F. Localization and Molecular Heterogeneity of Cholecystokinin in the Central and Peripheral Nervous System. Brain Res. 1979, 165, 201–218. [Google Scholar] [CrossRef]
- Beinfeld, M.C. An Introduction to Neuronal Cholecystokinin. Peptides 2001, 22, 1197–1200. [Google Scholar] [CrossRef]
- Rehfeld, J.F. Cholecystokinin-from Local Gut Hormone to Ubiquitous Messenger. Front. Endocrinol. (Lausanne) 2017, 8, 47. [Google Scholar] [CrossRef]
- Dufresne, M.; Seva, C.; Fourmy, D. Cholecystokinin and Gastrin Receptors. Physiol. Rev. 2006, 86, 805–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, T.; Dolai, S.; Xie, L.; Winter, E.; Orabi, A.I.; Karimian, N.; Cosen-Binker, L.I.; Huang, Y.C.; Thorn, P.; Cattral, M.S.; et al. Ex Vivo Human Pancreatic Slice Preparations Offer a Valuable Model for Studying Pancreatic Exocrine Biology. J. Biol. Chem. 2017, 292, 5957–5969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galindo, J.; Jones, N.; Powell, G.L.; Hollingsworth, S.J.; Shankley, N. Advanced qRT-PCR Technology Allows Detection of the Cholecystokinin 1 Receptor (CCK1R) Expression in Human Pancreas. Pancreas 2005, 31, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.T.; Lemp, G.F.; Gardner, J.D. Interaction of Cholecystokinin with Specific Membrane Receptors on Pancreatic Acinar Cells. Proc. Natl. Acad. Sci. USA 1980, 77, 2079–2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.A. Receptor-Mediated Signal Transduction Pathways and the Regulation of Pancreatic Acinar Cell Function. Curr. Opin. Gastroenterol. 2008, 24, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Karhunen, L.J.; Juvonen, K.R.; Huotari, A.; Purhonen, A.K.; Herzig, K.H. Effect of Protein, Fat, Carbohydrate and Fibre on Gastrointestinal Peptide Release in Humans. Regul. Pept. 2008, 149, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, S.S.; Thompson, D.G.; Warhurst, G.; Case, R.M.; Benson, R.S. Fatty Acid-Induced Cholecystokinin Secretion and Changes in Intracellular Ca2+ in Two Enteroendocrine Cell Lines, STC-1 and GLUTag. J. Physiol. 2000, 528, 165–176. [Google Scholar] [CrossRef]
- Tanaka, T.; Katsuma, S.; Adachi, T.; Koshimizu, T.A.; Hirasawa, A.; Tsujimoto, G. Free Fatty Acids Induce Cholecystokinin Secretion through GPR120. Naunyn Schmiedebergs Arch. Pharmacol. 2008, 377, 523–527. [Google Scholar] [CrossRef]
- Chen, M.C.; Wu, S.V.; Reeve, J.R., Jr.; Rozengurt, E. Bitter Stimuli Induce Ca2+ Signaling and CCK Release in Enteroendocrine STC-1 Cells: Role of L-Type Voltage-Sensitive Ca2+ Channels. Am. J. Physiol. Cell. Physiol. 2006, 291, C726–C739. [Google Scholar] [CrossRef] [Green Version]
- Jeon, T.I.; Seo, Y.K.; Osborne, T.F. Gut Bitter Taste Receptor Signalling Induces ABCB1 through a Mechanism Involving CCK. Biochem. J. 2011, 438, 33–37. [Google Scholar] [CrossRef]
- Yamazaki, T.; Morimoto-Kobayashi, Y.; Koizumi, K.; Takahashi, C.; Nakajima, S.; Kitao, S.; Taniguchi, Y.; Katayama, M.; Ogawa, Y. Secretion of a Gastrointestinal Hormone, Cholecystokinin, by Hop-Derived Bitter Components Activates Sympathetic Nerves in Brown Adipose Tissue. J. Nutr. Biochem. 2019, 64, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Owyang, C. Physiological Mechanisms of Cholecystokinin Action on Pancreatic Secretion. Am. J. Physiol. 1996, 271, G1–G7. [Google Scholar] [CrossRef] [PubMed]
- Lugea, A.; Gong, J.; Nguyen, J.; Nieto, J.; French, S.W.; Pandol, S.J. Cholinergic Mediation of Alcohol-Induced Experimental Pancreatitis. Alcohol. Clin. Exp. Res. 2010, 34, 1768–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widdison, A.L.; Alvarez, C.; Schwarz, M.; Reber, H.A. The Influence of Ethanol on Pancreatic Blood Flow in Cats with Chronic Pancreatitis. Surgery 1992, 112, 202–208. [Google Scholar] [PubMed]
- Han, B.; Logsdon, C.D. CCK Stimulates Mob-1 Expression and NF-kappaB Activation Via Protein Kinase C and Intracellular Ca (2+). Am. J. Physiol. Cell. Physiol. 2000, 278, C344–C351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evander, A.; Ihse, I.; Lundquist, I. Influence of Hormonal Stimulation by Caerulein on Acute Experimental Pancreatitis in the Rat. Eur. Surg. Res. 1981, 13, 257–268. [Google Scholar] [CrossRef]
- Barrett, T.D.; Yan, W.; Freedman, J.M.; Lagaud, G.J.; Breitenbucher, J.G.; Shankley, N.P. Role of CCK and Potential Utility of CCK1 Receptor Antagonism in the Treatment of Pancreatitis Induced by Biliary Tract Obstruction. Br. J. Pharmacol. 2008, 153, 1650–1658. [Google Scholar] [CrossRef] [Green Version]
- Saluja, A.K.; Lerch, M.M.; Phillips, P.A.; Dudeja, V. Why does Pancreatic Overstimulation Cause Pancreatitis? Annu. Rev. Physiol. 2007, 69, 249–269. [Google Scholar] [CrossRef]
- Herzig, K.H.; Schon, I.; Tatemoto, K.; Ohe, Y.; Li, Y.; Folsch, U.R.; Owyang, C. Diazepam Binding Inhibitor is a Potent Cholecystokinin-Releasing Peptide in the Intestine. Proc. Natl. Acad. Sci. USA. 1996, 93, 7927–7932. [Google Scholar] [CrossRef] [Green Version]
- Herzig, K.H.; Wilgus, C.; Schon, I.; Tatemoto, K.; Folsch, U.R. Regulation of the Action of the Novel Cholecystokinin-Releasing Peptide Diazepam Binding Inhibitor by Inhibitory Hormones and Taurocholate. Regul. Pept. 1998, 74, 193–198. [Google Scholar] [CrossRef]
- Rindi, G.; Grant, S.G.; Yiangou, Y.; Ghatei, M.A.; Bloom, S.R.; Bautch, V.L.; Solcia, E.; Polak, J.M. Development of Neuroendocrine Tumors in the Gastrointestinal Tract of Transgenic Mice. Heterogeneity of Hormone Expression. Am. J. Pathol. 1990, 136, 1349–1363. [Google Scholar] [PubMed]
- Glassmeier, G.; Herzig, K.H.; Hopfner, M.; Lemmer, K.; Jansen, A.; Scherubl, H. Expression of Functional GABAA Receptors in Cholecystokinin-Secreting Gut Neuroendocrine Murine STC-1 Cells. J. Physiol. 1998, 510, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Herzig, K.H.; Bardocz, S.; Grant, G.; Nustede, R.; Folsch, U.R.; Pusztai, A. Red Kidney Bean Lectin is a Potent Cholecystokinin Releasing Stimulus in the Rat Inducing Pancreatic Growth. Gut 1997, 41, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Flemstrom, G.; Makela, K.; Purhonen, A.K.; Sjoblom, M.; Jedstedt, G.; Walkowiak, J.; Herzig, K.H. Apelin Stimulation of Duodenal Bicarbonate Secretion: Feeding-Dependent and Mediated Via Apelin-Induced Release of Enteric Cholecystokinin. Acta Physiol. (Oxf.) 2011, 201, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Arita, Y.; Kimura, T.; Yazu, T.; Ogami, Y.; Nawata, H. Effects of Calcium-Channel Blockers on Cytosolic Free Calcium and Amylase Secretion in Rat Pancreatic Acini. Pharmacol. Toxicol. 1991, 68, 83–87. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egberts, J.-H.; Raza, G.S.; Wilgus, C.; Teyssen, S.; Kiehne, K.; Herzig, K.-H. Release of Cholecystokinin from Rat Intestinal Mucosal Cells and the Enteroendocrine Cell Line STC-1 in Response to Maleic and Succinic Acid, Fermentation Products of Alcoholic Beverages. Int. J. Mol. Sci. 2020, 21, 589. https://doi.org/10.3390/ijms21020589
Egberts J-H, Raza GS, Wilgus C, Teyssen S, Kiehne K, Herzig K-H. Release of Cholecystokinin from Rat Intestinal Mucosal Cells and the Enteroendocrine Cell Line STC-1 in Response to Maleic and Succinic Acid, Fermentation Products of Alcoholic Beverages. International Journal of Molecular Sciences. 2020; 21(2):589. https://doi.org/10.3390/ijms21020589
Chicago/Turabian StyleEgberts, Jan-Hendrik, Ghulam Shere Raza, Cornelia Wilgus, Stefan Teyssen, Karlheinz Kiehne, and Karl-Heinz Herzig. 2020. "Release of Cholecystokinin from Rat Intestinal Mucosal Cells and the Enteroendocrine Cell Line STC-1 in Response to Maleic and Succinic Acid, Fermentation Products of Alcoholic Beverages" International Journal of Molecular Sciences 21, no. 2: 589. https://doi.org/10.3390/ijms21020589