Cytosolic Sensors for Pathogenic Viral and Bacterial Nucleic Acids in Fish

Abstract
:1. Introduction
2. Viral and Bacterial PAMPs and Their PRRs
3. DExD/H-Box RNA Helicases
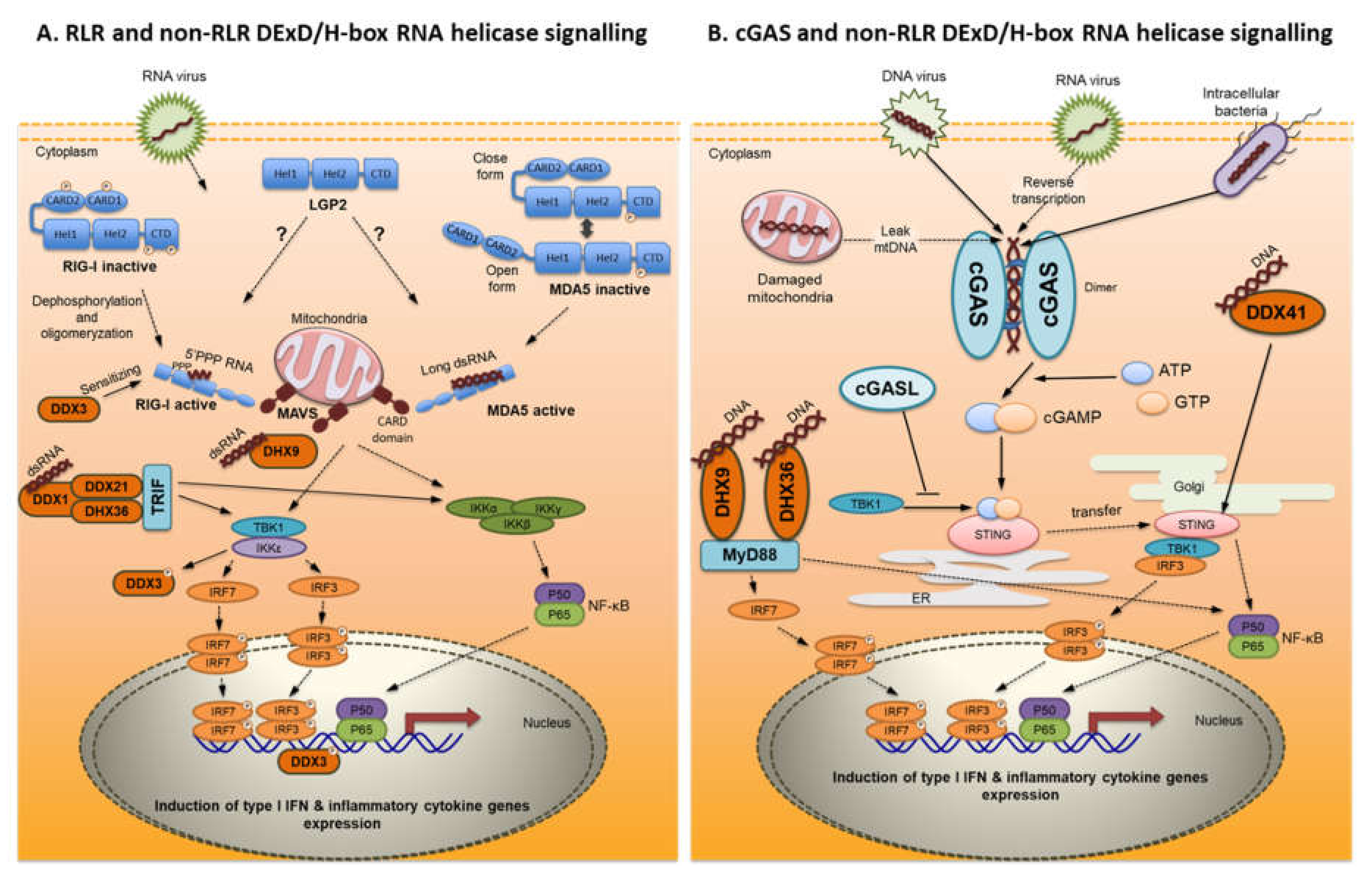
3.1. RIG-I-Like Receptors (RLR)
3.1.1. Receptor Structure
3.1.2. Ligands
3.1.3. Signal Transduction
4. RLRs in Fish
4.1. RIG-I (DDX58)
4.2. MDA5
4.3. LGP2
5. Non-RLR DExD/H-box RNA Helicases
6. MAVS
7. cGAS
8. LSm14A
9. PKR and PKZ
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, S.; Thomsen, A.R. Sensing of RNA viruses: A review of innate immune receptors involved in recognizing RNA virus invasion. J. Virol. 2012, 86, 2900–2910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, T.; Kawai, T.; Akira, S. Recognition of nucleic acids by pattern-recognition receptors and its relevance in autoimmunity. Immunol. Rev. 2011, 243, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Paludan, S.R.; Bowie, A.G. Immune sensing of DNA. Immunity 2013, 38, 870–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, D.; McCarthy, J.; O’Driscoll, C.; Melgar, S. Pattern recognition receptors-molecular orchestrators of inflammation in inflammatory bowel disease. Cytokine Growth Factor Rev. 2013, 24, 91–104. [Google Scholar] [CrossRef]
- Wilkins, C.; Gale, M. Recognition of viruses by cytoplasmic sensors. Curr. Opin. Immunol. 2011, 22, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Yoneyama, M.; Fujita, T. Recognition of viral nucleic acids in innate immunity. Rev. Med. Virol. 2010, 20, 4–22. [Google Scholar] [CrossRef]
- Medzhitov, R.; Janeway, C.A. Innate immunity: Impact on the adaptive immune response. Curr. Opin. Immunol. 1997, 9, 4–9. [Google Scholar] [CrossRef]
- Beg, A.A. Endogenous ligands of Toll-like receptors: Implications for regulating inflammatory and immune responses. Trends Immunol. 2002, 23, 509–512. [Google Scholar] [CrossRef]
- Matzinger, P. The danger model: A renewed sense of self. Science 2002, 296, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, T.H.; Paludan, S.R. Reading the viral signature by Toll-like receptors and other pattern recognition receptors. J. Mol. Med. 2005, 83, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Bieback, K.; Lien, E.; Klagge, I.M.; Avota, E.; Schneider-Schaulies, J.; Duprex, W.P.; Wagner, H.; Kirschning, C.J.; ter Meulen, V.; Schneider-Schaulies, S. Hemagglutinin protein of wild-type measles virus activates Toll-like receptor 2 signaling. J. Virol. 2002, 76, 8729–8736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burzyn, D.; Rassa, J.C.; Kim, D.; Nepomnaschy, I.; Ross, S.R.; Piazzon, I. Toll-Like receptor 4-dependent activation of dendritic cells by a retrovirus. J. Virol. 2004, 78, 576–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rassa, J.C.; Meyers, J.L.; Zhang, Y.; Kudaravalli, R.; Ross, S.R. Murine retroviruses activate B cells via interaction with toll-like receptor 4. Proc. Natl. Acad. Sci. USA 2002, 99, 2281–2286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbah, A.; Chang, T.H.; Harnack, R.; Frohlich, V.; Tominaga, K.; Dube, P.H.; Xiang, Y.; Bose, S. Activation of innate immune antiviral responses by Nod2. Nat. Immunol. 2009, 10, 1073–1080. [Google Scholar] [CrossRef]
- Albiger, B.; Dahlberg, S.; Henriques-Normark, B.; Normark, S. Role of the innate immune system in host defence against bacterial infections: Focus on the Toll-like receptors. J. Intern. Med. 2007, 261, 511–528. [Google Scholar] [CrossRef]
- Schwandner, R.; Dziarski, R.; Wesche, H.; Rothe, M.; Kirschning, C.J. Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated by Toll-like receptor 2. J. Biol. Chem. 1999, 274, 17406–17409. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Miao, E.A.; Ernst, R.K.; Dors, M.; Mao, D.P.; Aderem, A. Pseudomonas aeruginosa activates caspase 1 through Ipaf. Proc. Natl. Acad. Sci. USA 2008, 105, 2562–2567. [Google Scholar] [CrossRef] [Green Version]
- Kanwal, Z.; Wiegertjes, G.F.; Veneman, W.J.; Meijer, A.H.; Spaink, H.P. Comparative studies of Toll-like receptor signalling using zebrafish. Dev. Comp. Immunol. 2014, 46, 35–52. [Google Scholar] [CrossRef]
- Quiniou, S.M.A.; Boudinot, P.; Bengtén, E. Comprehensive survey and genomic characterization of Toll-like receptors (TLRs) in channel catfish, Ictalurus punctatus: Identification of novel fish TLRs. Immunogenetics 2013, 65, 511–530. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Hikima, J.; Hwang, S.D.; Jung, T.S. Innate immunity of finfish: Primordial conservation and function of viral RNA sensors in teleosts. Fish Shellfish Immunol. 2013, 35, 1689–1702. [Google Scholar] [CrossRef] [PubMed]
- Palti, Y. Toll-like receptors in bony fish: From genomics to function. Dev. Comp. Immunol. 2011, 35, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Rebl, A.; Goldammer, T.; Seyfert, H.M. Toll-like receptor signaling in bony fish. Vet. Immunol. Immunopathol. 2010, 134, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Ranji, A.; Boris-Lawrie, K. RNA helicases: Emerging roles in viral replication and the host innate response. RNA Biol. 2010, 7, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Cordin, O.; Banroques, J.; Tanner, N.K.; Linder, P. The DEAD-box protein family of RNA helicases. Gene 2006, 367, 17–37. [Google Scholar] [CrossRef]
- Fullam, A.; Schröder, M. DExD/H-box RNA helicases as mediators of anti-viral innate immunity and essential host factors for viral replication. Biochim. Biophys. Acta Gene Regul. Mech. 2013, 1829, 854–865. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.; Pelletier, J. The biology of DHX9 and its potential as a therapeutic target. Oncotarget 2016, 7, 42716–42739. [Google Scholar] [CrossRef] [Green Version]
- Story, R.M.; Li, H.; Abelson, J.N. Crystal structure of a DEAD box protein from the hyperthermophile Methanococcus jannaschii. Proc. Natl. Acad. Sci. USA 2001, 98, 1465–1470. [Google Scholar] [CrossRef] [Green Version]
- Ariumi, Y. Multiple functions of DDX3 RNA helicase in gene regulation, tumorigenesis, and viral infection. Front. Genet. 2014, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fuller-Pace, F.V. DExD/H box RNA helicases: Multifunctional proteins with important roles in transcriptional regulation. Nucleic Acids Res. 2006, 34, 4206–4215. [Google Scholar] [CrossRef] [PubMed]
- Linder, P. Dead-box proteins: A family affair-active and passive players in RNP-remodeling. Nucleic Acids Res. 2006, 34, 4168–4180. [Google Scholar] [CrossRef] [PubMed]
- Schröder, M. Human DEAD-box protein 3 has multiple functions in gene regulation and cell cycle control and is a prime target for viral manipulation. Biochem. Pharmacol. 2010, 79, 297–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, T.; Poeck, H. Apoptosis induction by cytosolic RNA helicases. J. Med. Microb. Diagn. 2012, 1, 4–9. [Google Scholar] [CrossRef]
- Kolokoltsova, O.A.; Grant, A.M.; Huang, C.; Smith, J.K.; Poussard, A.L.; Tian, B.; Brasier, A.R.; Peters, C.J.; Tseng, C.K.; Torre, J.C.; et al. RIG-I enhanced interferon independent apoptosis upon junin virus infection. PLoS ONE 2014, 9, e99610. [Google Scholar] [CrossRef] [PubMed]
- Abdelhaleem, M. Over-expression of RNA helicases in cancer. Anticancer Res. 2004, 3954, 3951–3953. [Google Scholar]
- Fuller-Pace, F.V. DEAD box RNA helicase functions in cancer DEAD. RNA Biol. 2013, 10, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, O.; Akira, S. MDA5/RIG-I and virus recognition. Curr. Opin. Immunol. 2008, 20, 17–22. [Google Scholar] [CrossRef]
- Yoneyama, M.; Onomoto, K.; Jogi, M.; Akaboshi, T.; Fujita, T. Viral RNA detection by RIG-I-like receptors. Curr. Opin. Immunol. 2015, 32, 48–53. [Google Scholar] [CrossRef]
- Brisse, M.; Ly, H. Comparative structure and function analysis of the RIG-I-like receptors: RIG-I and MDA5. Front. Immunol. 2019, 10, 1–27. [Google Scholar] [CrossRef]
- Loo, Y.; Gale, M. Immune signaling by RIG-I-like receptors. Immunity 2011, 34, 680–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.J.; Lee, H.; Kim, J.; Park, S.Y.; Kim, T.; Lee, W.; Jang, D.; Yoon, J.; Choi, Y.; Kim, S.; et al. HDAC 6 regulates cellular viral RNA sensing by deacetylation of RIG. EMBO J. 2016, 35, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.N.; Zou, P.F.; Nie, P. Retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) in fish: Current knowledge and future perspectives. Immunology 2017, 151, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Takahasi, K.; Yoneyama, M.; Nishihori, T.; Hirai, R.; Kumeta, H.; Narita, R.; Gale, M.; Inagaki, F.; Fujita, T. Nonself RNA-sensing mechanism of RIG-I helicase and activation of antiviral immune responses. Mol. Cell 2008, 29, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Deddouche, S.; Goubau, D.; Rehwinkel, J.; Chakravarty, P.; Begum, S.; Maillard, P.V.; Borg, A.; Matthews, N.; Feng, Q.; van Kuppeveld, F.J.M.; et al. Identification of an LGP2-associated MDA5 agonist in picornavirus-infected cells. eLife 2014, 3, 1–20. [Google Scholar] [CrossRef]
- Lazarte, J.M.S.; Thompson, K.D.; Jung, T.S. Pattern recognition by melanoma differentiation-associated gene 5 (Mda5) in teleost fish: A review. Front. Immunol. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Bruns, A.M.; Horvath, C.M. LGP2 synergy with MDA5 in RLR-mediated RNA recognition and antiviral signaling. Cytokine 2015, 74, 198–206. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Kinch, L.N.; Brautigam, C.A.; Chen, X.; Du, F.; Grishin, N.V.; Chen, Z.J. Ubiquitin-induced oligomerization of the RNA sensors RIG-I and MDA5 activates antiviral innate immune response. Immunity 2012, 36, 959–973. [Google Scholar] [CrossRef] [Green Version]
- Schoggins, J.W.; Rice, C.M. Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef]
- Yoneyama, M.; Kikuchi, M.; Matsumoto, K.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Foy, M.; Loo, Y.; Gale, M.; Akira, S.; et al. Shared and unique functions of the DExD/H-Box helicases RIG-I, MDA5, and LGP2 in antiviral innate immunity. J. Immunol. 2005, 175, 2851–2858. [Google Scholar] [CrossRef] [Green Version]
- Komuro, A.; Horvath, C.M. RNA- and virus-independent inhibition of antiviral signaling by RNA helicase LGP2. J. Virol. 2006, 80, 12332–12342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisien, J.; Lenoir, J.J.; Mandhana, R.; Rodriguez, K.R.; Qian, K.; Bruns, A.M.; Horvath, C.M. RNA sensor LGP 2 inhibits TRAF ubiquitin ligase to negatively regulate innate immune signaling. EMBO Rep. 2018, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, T.; Valdes, M.; Elsby, R.; Kakuta, S.; Caceres, G.; Saijo, S.; Iwakura, Y.; Barber, G.N. Loss of DExD/H Box RNA helicase LGP2 manifests disparate antiviral responses. J. Immunol. 2007, 178, 6444–6455. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Kato, H.; Kumagai, Y.; Yoneyama, M.; Sato, S.; Matsushita, K.; Tsujimura, T.; Fujita, T.; Akira, S.; Takeuchi, O. LGP2 is a positive regulator of RIG-I- and MDA5-mediated antiviral responses. Proc. Natl. Acad. Sci. USA 2010, 107, 1512–1517. [Google Scholar] [CrossRef] [Green Version]
- Liniger, M.; Summerfield, A.; Zimmer, G.; McCullough, K.C.; Ruggli, N. Chicken cells sense influenza A virus infection through MDA5 and CARDIF signaling involving LGP2. J. Virol. 2012, 86, 705–717. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Yu, D.; Fan, Y.; Peng, L.; Wu, Y.; Yao, Y.G. Loss of RIG-I leads to a functional replacement with MDA5 in the Chinese tree shrew. Proc. Natl. Acad. Sci. USA 2016, 113, 10950–10955. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, M.; Hikima, J.; Kondo, H.; Hirono, I.; Jung, T.; Aoki, T. Evolutional conservation of molecular structure and antiviral function of a viral RNA receptor, LGP2, in Japanese flounder, Paralichthys olivaceus. J. Immunol. 2010, 185, 7507–7517. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Su, J.; Heng, J.; Dong, J.; Zhang, R.; Zhu, H. Identifcation and expression profiling analysis of grass carp Ctenopharyngodon idella LGP2 cDNA. Fish Shellfish Immunol. 2010, 29, 349–355. [Google Scholar] [CrossRef]
- Xu, C.; Evensen, Ø.; Munang’andu, H.M. De Novo transcriptome analysis shows that SAV-3 infection upregulates pattern recognition receptors of the endosomal Toll-Like and RIG-I-like receptor signaling pathways in macrophage/dendritic like TO cells. Viruses 2016, 8, 114. [Google Scholar] [CrossRef]
- Nerbøvik, I.G.; Solheim, M.A.; Eggestøl, H.Ø.; Rønneseth, A.; Jakobsen, R.A.; Wergeland, H.I.; Haugland, G.T. Molecular cloning of MDA5, phylogenetic analysis of RIG-I-like receptors (RLRs) and differential gene expression of RLRs, interferons and proinflammatory cytokines after in vitro challenge with IPNV, ISAV and SAV in the salmonid cell line TO. J. Fish Dis. 2017, 40, 1529–1544. [Google Scholar] [CrossRef]
- Wessel, Ø.; Olsen, C.M.; Rimstad, E.; Dahle, M.K. Piscine orthoreovirus (PRV) replicates in Atlantic salmon (Salmo salar L.) erythrocytes ex vivo. Vet. Res. 2015, 26, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skjesol, A.; Skjæveland, I.; Elnæs, M.; Timmerhaus, G.; Fredriksen, B.N.; Jørgensen, S.M.; Krasnov, A.; Jørgensen, J.B. IPNV with high and low virulence: Host immune responses and viral mutations during infection. Virol. J. 2011, 8, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajendran, K.V.; Zhang, J.; Liu, S.; Peatman, E.; Kucuktas, H.; Wang, X.; Liu, H.; Wood, T.; Terhune, J.; Liu, Z. Pathogen recognition receptors in channel catfish: II. Identification, phylogeny and expression of retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs). Dev. Comp. Immunol. 2012, 37, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Liu, H.; Kong, R.; Wang, L.; Wang, Y.; Hu, W.; Guo, Q. Expression profiles of carp IRF-3/-7 correlate with the up-regulation of RIG-I/MAVS/TRAF3/TBK1, four pivotal molecules in RIG-I signaling pathway. Fish Shellfish Immunol. 2011, 30, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Mou, C.; Wang, Y.; Zhang, Q.; Gao, F.; Li, Z.; Tong, J.; Zhou, L.; Gui, J. Differential interferon system gene expression profiles in susceptible and resistant gynogenetic clones of gibel carp challenged with herpesvirus CaHV. Dev. Comp. Immunol. 2018, 86, 52–64. [Google Scholar] [CrossRef]
- Yang, C.; Su, J.; Huang, T.; Zhang, R.; Peng, L. Identification of a retinoic acid-inducible gene I from grass carp (Ctenopharyngodon idella) and expression analysis in vivo and in vitro. Fish Shellfish Immunol. 2011, 30, 936–943. [Google Scholar] [CrossRef]
- Chen, L.; Li, Q.; Su, J.; Yang, C.; Li, Y.; Rao, Y. Trunk kidney of grass carp (Ctenopharyngodon idella) mediates immune responses against GCRV and viral/bacterial PAMPs in vivo and in vitro. Fish Shellfish Immunol. 2013, 34, 909–919. [Google Scholar] [CrossRef]
- Liao, Z.; Wan, Q.; Su, H.; Wu, C.; Su, J. Pattern recognition receptors in grass carp Ctenopharyngodon idella: I. Organization and expression analysis of TLRs and RLRs. Dev. Comp. Immunol. 2017, 76, 93–104. [Google Scholar] [CrossRef]
- Feng, J.; Guo, S.; Lin, P.; Wang, Y.; Zhang, Z.; Zhang, Z.; Yu, L. Identification of a retinoic acid-inducible gene I from Japanese eel (Anguilla japonica) and expression analysis in vivo and in vitro. Fish Shellfish Immunol. 2016, 55, 249–256. [Google Scholar] [CrossRef]
- Huang, B.; Wang, Z.X.; Zhang, C.; Zhai, S.W.; Han, Y.S.; Huang, W.S.; Nie, P. Identification of a novel RIG-I isoform and its truncating variant in Japanese eel, Anguilla japonica. Fish Shellfish Immunol. 2019, 94, 373–380. [Google Scholar] [CrossRef]
- Chen, H.; Liu, W.; Wu, S.; Chiou, P.P.; Li, Y.; Chen, Y.; Lin, G.; Lu, M.; Wu, J. RIG-I specifically mediates group II type I IFN activation in nervous necrosis virus infected zebrafish cells. Fish Shellfish Immunol. 2015, 43, 427–435. [Google Scholar] [CrossRef]
- Wang, W.; Asim, M.; Yi, L.; Hegazy, A.M.; Hu, X.; Zhou, Y.; Ai, T.; Lin, L. Abortive infection of snakehead fish vesiculovirus in ZF4 cells was associated with the RLRs pathway activation by viral replicative intermediates. Int. J. Mol. Sci. 2015, 16, 6235–6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, P.F.; Chang, M.X.; Li, Y.; Zhang, S.H.; Fu, J.P.; Chen, S.N.; Nie, P. Higher antiviral response of RIG-I through enhancing RIG-I/MAVS-mediated signaling by its long insertion variant in zebrafish. Fish Shellfish Immunol. 2015, 43, 13–24. [Google Scholar] [CrossRef] [PubMed]
- López-Muñoz, A.; Roca, F.J.; Sepulcre, M.P.; Meseguer, J.; Mulero, V. Zebrafish larvae are unable to mount a protective antiviral response against waterborne infection by spring viremia of carp virus. Dev. Comp. Immunol. 2010, 34, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.; Zhang, Y.; Dong, W.; Xiang, L.; Shao, J. Involvement of zebrafish RIG-I in NF-κB and IFN signaling pathways: Insights into functional conservation of RIG-I in antiviral innate immunity. Dev. Comp. Immunol. 2015, 48, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Chen, S.; Liu, Y.; Lin, Y.; Liu, H.; Guo, L.; Lin, B.; Huang, S.; Xu, A. Zebrafish TRIF, a golgi-localized protein, participates in IFN induction and NF- B activation. J. Immunol. 2008, 180, 5373–5383. [Google Scholar] [CrossRef] [PubMed]
- Rakus, K.; Mojzesz, M.; Widziolek, M.; Pooranachandran, N.; Teitge, F.; Surachetpong, W.; Chadzinska, M.; Steinhagen, D.; Adamek, M. Antiviral response of adult zebrafish (Danio rerio) during tilapia lake virus (TiLV) infection. Fish Shellfish Immunol. 2020, 101, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Collet, B.; Nie, P.; Lester, K.; Campbell, S.; Secombes, C.J.; Zou, J. Expression and functional characterization of the RIG-I-Like receptors MDA5 and LGP2 in Rainbow trout (Oncorhynchus mykiss). J. Virol. 2011, 85, 8403–8412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Li, J.; Xiao, J.; Chen, H.; Lu, L.; Wang, X.; Tian, Y.; Feng, H. The antiviral signaling mediated by black carp MDA5 is positively regulated by LGP2. Fish Shellfish Immunol. 2017, 66, 360–371. [Google Scholar] [CrossRef]
- Paria, A.; Makesh, M.; Chauhari, A.; Purushothaman, C.S.; Rajendran, K.V. Molecular characterisation, ontogeny and expression analysis of melanoma differentiation-associated factor 5 (MDA5) from Asian seabass, Lates calcarifer. Dev. Comp. Immunol. 2018, 78, 71–82. [Google Scholar] [CrossRef]
- Li, Y.; Jin, S.; Zhao, X.; Luo, H.; Li, R.; Li, D.; Xiao, T. Sequence and expression analysis of the cytoplasmic pattern recognition receptor melanoma differentiation-associated gene 5 from the barbel chub Squaliobarbus curriculus. Fish Shellfish Immunol. 2019, 94, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.Y.; Xing, W.X.; Shan, S.J.; Zhang, S.Q.; Li, Y.Q.; Li, T.; An, L.; Yang, G.W. Characterization and immune response expression of the Rig-I-like receptor mda5 in common carp Cyprinus carpio. J. Fish Biol. 2016, 88, 2188–2202. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Huang, T.; Dong, J.; Heng, J.; Zhang, R.; Peng, L. Molecular cloning and immune responsive expression of MDA5 gene, a pivotal member of the RLR gene family from grass carp Ctenopharyngodon idella. Fish Shellfish Immunol. 2010, 28, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Paria, A.; Deepika, A.; Sreedharan, K.; Makesh, M.; Bedekar, M.K.; Purushothaman, C.S.; Rajendran, K.V. Molecular cloning, characterisation and expression analysis of melanoma differentiation associated gene 5 (MDA5) of green chromide, Etroplus suratensis. Gene 2015, 557, 172–181. [Google Scholar] [CrossRef]
- Ohtani, M.; Hikima, J.; Kondo, H.; Hirono, I.; Jung, T.; Aoki, T. Characterization and antiviral function of a cytosolic sensor gene, MDA5, in Japanese flounder, Paralichthys olivaceus. Dev. Comp. Immunol. 2011, 35, 554–562. [Google Scholar] [CrossRef]
- Hikima, J.; Yi, M.K.; Ohtani, M.; Jung, C.Y.; Kim, Y.K.; Mun, J.Y.; Kim, Y.R.; Takeyama, H.; Aoki, T.; Jung, T.S. LGP2 expression is enhanced by interferon regulatory factor 3 in Olive flounder, Paralichthys olivaceus. PLoS ONE 2012, 7, e51522. [Google Scholar] [CrossRef] [Green Version]
- Gu, T.; Lu, L.; An, C.; Chen, B.; Wei, W.; Wu, X.; Xu, Q.; Chen, G. MDA5 and LGP2 acts as a key regulator though activating NF-κB and IRF3 in RLRs signaling of Mandarinfish. Fish Shellfish Immunol. 2019, 86, 1114–1122. [Google Scholar] [CrossRef]
- Huang, Y.; Yu, Y.; Yang, Y.; Yang, M.; Zhou, L.; Huang, X.; Qin, Q. Antiviral function of grouper MDA5 against iridovirus and nodavirus. Fish Shellfish Immunol. 2016, 54, 188–196. [Google Scholar] [CrossRef]
- Jia, P.; Jia, K.; Chen, L.; Le, Y.; Jin, Y.; Zhang, J.; Zhu, L.; Zhang, L.; Yi, M. Identification and characterization of the melanoma differentiation-associated gene 5 in sea perch, Lateolabrax japonicus. Dev. Comp. Immunol. 2016, 61, 161–168. [Google Scholar] [CrossRef]
- Mu, Y.; Li, M.; Ding, F.; Ding, Y.; Ao, J.; Hu, S.; Chen, X. De novo characterization of the spleen transcriptome of the large yellow croaker (Pseudosciaena crocea) and analysis of the immune relevant genes and pathways involved in the antiviral response. PLoS ONE 2014, 9, e101069. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Hu, Y.; Zhang, S.; Zheng, J.; Zeng, L.; Zhang, J.; Zhu, A.; Wu, C. Molecular characterization and expression analyses of three RIG-I-like receptor signaling pathway genes (MDA5, LGP2 and MAVS) in Larimichthys crocea. Fish Shellfish Immunol. 2016, 55, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.F.; Chang, M.X.; Xue, N.N.; Liu, X.Q.; Li, J.H.; Fu, J.P.; Chen, S.N.; Nie, P. Melanoma differentiation-associated gene 5 in zebrafish provoking higher interferon-promoter activity through signalling enhancing of its shorter splicing variant. Immunology 2013, 141, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Paria, A.; Deepika, A.; Sreedharan, K.; Makesh, M.; Chaudhari, A.; Purushothaman, C.S.; Rajendran, K.V. Identification, ontogeny and expression analysis of a novel laboratory of genetics and physiology 2 (LGP2) transcript in Asian seabass, Lates calcarifer. Fish Shellfish Immunol. 2017, 62, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Jensen, I.; Seppola, M.; Steiro, K.; Sandaker, E.; Mennen, S. Susceptibility of Atlantic cod Gadus morhua juveniles to different routes of experimental challenge with infectious pancreatic necrosis virus (IPNV). Dis. Aquat. Org. 2009, 85, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Yan, J.; Chen, H.; Li, J.; Tian, Y.; Feng, H. LGP2 of black carp plays an important role in the innate immune response against SVCV and GCRV. Fish Shellfish Immunol. 2016, 57, 127–135. [Google Scholar] [CrossRef]
- Cao, X.L.; Chen, J.J.; Cao, Y.; Nie, G.X.; Wan, Q.Y.; Wang, L.F.; Su, J.G. Identification and expression of the laboratory of genetics and physiology 2 gene in common carp Cyprinus carpio. J. Fish Biol. 2015, 86, 74–91. [Google Scholar] [CrossRef]
- Mohanty, A.; Sadangi, S.; Paichha, M.; Samanta, M. Molecular characterization and expressional quantification of lgp2, a modulatory co-receptor of RLR-signalling pathway in the Indian major carp Labeo rohita following pathogenic challenges and PAMP stimulations. J. Fish Biol. 2020, 96, 1399–1410. [Google Scholar] [CrossRef]
- Han, J.; Wang, Y.; Chu, Q.; Xu, T. The evolution and functional characterization of miiuy croaker cytosolic gene LGP2 involved in immune response. Fish Shellfish Immunol. 2016, 58, 193–202. [Google Scholar] [CrossRef]
- Yu, Y.; Huang, Y.; Yang, Y.; Wang, S.; Yang, M.; Huang, X.; Qin, Q. Negative regulation of the antiviral response by grouper LGP2 against fish viruses. Fish Shellfish Immunol. 2016, 56, 358–366. [Google Scholar] [CrossRef]
- Poynter, S.J.; Herrington-Krause, S.; Dewitte-orr, S.J. Two DExD/H-box helicases, DDX3 and DHX9, identified in rainbow trout are able to bind dsRNA. Fish Shellfish Immunol. 2019, 93, 1056–1066. [Google Scholar] [CrossRef]
- Jia, P.; Zhang, J.; Jin, Y.; Zeng, L.; Jia, K.; Yi, M. Characterization and expression analysis of laboratory of genetics and physiology 2 gene in sea perch, Lateolabrax japonicus. Fish Shellfish Immunol. 2015, 47, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, X.; Li, Z.; Wu, M.; Gui, J.; Zhang, Y. Alternative splicing transcripts of zebrafish LGP2 gene differentially contribute to IFN antiviral response. J. Immunol. 2018, 200, 688–703. [Google Scholar] [CrossRef] [PubMed]
- Mojzesz, M.; Klak, K.; Wojtal, P.; Adamek, M.; Podlasz, P.; Chmielewska-Krzesinska, M.; Matras, M.; Reichert, M.; Chadzinska, M.; Rakus, K. Viral infection-induced changes in the expression profile of non-RLR DExD/H-box RNA helicases (DDX1, DDX3, DHX9, DDX21 and DHX36) in zebrafish and common carp. Fish Shellfish Immunol. 2020, 104, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Huang, X.; Yu, Y.; Zhang, J.; Ni, S.; Hu, Y.; Huang, Y.; Qin, Q. Fish DDX3X exerts antiviral function against grouper nervous necrosis virus infection. Fish Shellfish Immunol. 2017, 71, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Morimoto, N.; Kono, T.; Sakai, M.; Hikima, J. Molecular characterization and expression of the teleost cytosolic DNA sensor genes cGAS, LSm14A, DHX9, and DHX36 in Japanese medaka, Oryzias latipes. Dev. Comp. Immunol. 2019, 99, 103402. [Google Scholar] [CrossRef] [PubMed]
- Quynh, N.T.; Hikima, J.; Kim, Y.; Fagutao, F.F.; Kim, M.S.; Aoki, T.; Jung, T.S. The cytosolic sensor, DDX41, activates antiviral and inflammatory immunity in response to stimulation with double-stranded DNA adherent cells of the olive flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2015, 44, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Huang, Y.; Huang, X.; Li, C.; Ni, S.; Yu, Y.; Qin, Q. Grouper DDX41 exerts antiviral activity against fish iridovirus and nodavirus infection. Fish Shellfish Immunol. 2019, 91, 40–49. [Google Scholar] [CrossRef]
- Chang, M.X.; Zhang, J. Alternative pre-mRNA splicing in mammals and teleost fish: A effective strategy for the regulation of immune responses against pathogen infection. Int. J. Mol. Sci. 2017, 18, 1530. [Google Scholar] [CrossRef]
- Gong, X.; Zhang, Q.; Gui, J.; Zhang, Y. SVCV infection triggers fish IFN response through RLR signaling pathway. Fish Shellfish Immunol. 2019, 86, 1058–1063. [Google Scholar] [CrossRef]
- Biacchesi, S.; Leberre, M.; Lamoureux, A.; Louise, Y.; Lauret, E.; Boudinot, P.; Bremont, M. Mitochondrial antiviral signaling protein plays a major role in induction of the fish innate immune response against RNA and DNA viruses. J. Immunol. 2009, 83, 7815–7827. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Zhang, Y.; Liu, T.; Shi, J.; Wang, B.; Gui, J. Fish MITA serves as a mediator for distinct fish IFN gene activation dependent on IRF3 or IRF7. J. Immunol. 2011, 187, 2531–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Su, J.; Yang, C.; Peng, L.; Wan, Q.; Wang, L. Functional characterizations of RIG-I to GCRV and viral/bacterial PAMPs in Grass carp Ctenopharyngodon idella. PLoS ONE 2012, 7, e42182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabor, K.A.; Charette, J.R.; Pietraszewski, M.J.; Wingfield, D.J.; Shim, J.S.; Millard, P.J.; Kim, C.H. A DN-mda5 transgenic zebrafish model demonstrates that Mda5 plays an important role in snakehead rhabdovirus resistance. Dev. Comp. Immunol. 2015, 51, 298–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Zhang, B.; Sun, L. Poly(I:C) induces antiviral immune responses in Japanese flounder (Paralichthys olivaceus) that require TLR3 and MDA5 and is negatively regulated by Myd88. PLoS ONE 2014, 9, e112918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Q.; Yang, C.; Rao, Y.; Liao, Z.; Su, J. MDA5 induces a stronger interferon response than RIG-I to GCRV infection through a mechanism involving the phosphorylation and dimerization of IRF3 and IRF7 in CIK cells. Front. Immunol. 2017, 8, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, T.; Rao, Y.; Su, J.; Yang, C.; Chen, X.; Chen, L.; Yan, N. Functions of MDA5 and its domains in response to GCRV or bacterial PAMPs. Fish Shellfish Immunol. 2015, 46, 693–702. [Google Scholar] [CrossRef]
- Gao, F.; Lu, M.; Wang, M.; Liu, Z.; Ke, X.; Zhang, D.; Cao, J. Molecular characterization and function analysis of three RIG-I-like receptor signaling pathway genes (MDA5, LGP2 and MAVS) in Oreochromis niloticus. Fish Shellfish Immunol. 2018, 82, 101–114. [Google Scholar] [CrossRef]
- Chen, X.; Yang, C.; Su, J.; Rao, Y.; Gu, T. LGP2 plays extensive roles in modulating innate immune responses in Ctenopharyngodon idella kidney (CIK) cells. Dev. Comp. Immunol. 2015, 49, 138–148. [Google Scholar] [CrossRef]
- Rao, Y.; Wan, Q.; Yang, C.; Su, J. Grass carp laboratory of genetics and physiology 2 serves as a negative regulator in retinoic acid-inducible gene I- and melanoma differentiation-associated gene 5-mediated antiviral signaling in resting state and early stage of grass carp reovirus infection. Front. Immunol. 2017, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Kim, T.; Bao, M.; Facchinetti, V.; Jung, S.Y.; Ghaffari, A.A.; Qin, J.; Cheng, G.; Liu, Y. DDX1, DDX21, and DHX36 helicases form a complex with the adaptor molecule TRIF to sense dsRNA in dendritic cells. Immunity 2011, 34, 866–878. [Google Scholar] [CrossRef] [Green Version]
- Schröder, M.; Baran, M.; Bowie, A.G. Viral targeting of DEAD box protein 3 reveals its role in TBK1/IKKε-mediated IRF activation. EMBO J. 2008, 27, 2147–2157. [Google Scholar] [CrossRef] [PubMed]
- Soulat, D.; Bürckstümmer, T.; Westermayer, S.; Goncalves, A.; Bauch, A.; Stefanovic, A.; Hantschel, O.; Bennett, K.L.; Decker, T.; Superti-Furga, G. The DEAD-box helicase DDX3X is a critical component of the TANK-binding kinase 1-dependent innate immune response. EMBO J. 2008, 27, 2135–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yuan, B.; Lu, N.; Facchinetti, V.; Liu, Y. DHX9 pairs with IPS-1 to sense double-stranded RNA in myeloid dendritic cells. J. Immunol. 2011, 187, 4501–4508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosallanejad, K.; Sekine, Y.; Ishikura-Kinoshita, S.; Kumagai, K.; Nagano, T.; Matsuzawa, A.; Takeda, K.; Naguro, I.; Ichijo, H. The DEAH-box RNA helicase DHX15 activates NF-kB and MAPK signaling downstream of MAVS during antiviral eesponses. Immunology 2014, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Moy, R.H.; Cole, B.S.; Yasunaga, A.; Gold, B.; Shankarling, G.; Varble, A.; Molleston, J.M.; ten Oever, B.R.; Lynch, K.W.; Cherry, S. Stem-loop recognition by DDX17 facilitates miRNA processing and antiviral defense. Cell 2014, 158, 764–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, J.; Cao, Y.; Ling, T.; Li, P.; Wu, S.; Peng, D.; Wang, Y.; Jia, X.; Chen, S.; Xu, A.; et al. DDX23, an evolutionary conserved dsRNA sensor, participates in innate antiviral responses by pairing with TRIF or MAVS. Front. Immunol. 2019, 10, 2202. [Google Scholar] [CrossRef]
- Sugimoto, N.; Mitoma, H.; Kim, T.; Hanabuchi, S.; Liu, Y. Helicase proteins DHX29 and RIG-I cosense cytosolic nucleic acids in the human airway system. Proc. Natl. Acad. Sci. USA 2014, 111, 7747–7752. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yuan, B.; Bao, M.; Lu, N.; Kim, T.; Liu, Y. The helicase DDX41 senses intracellular DNA mediated by the adaptor STING in dendritic cells. Nat. Immunol. 2011, 12, 959–965. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, M.; Oshiumi, H.; Matsumoto, M.; Seya, T. DDX60, a DExD/H-box helicase, is a novel antiviral factor promoting RIG-I-Like receptor-mediated signaling. Mol. Cell. Biol. 2011, 31, 3802–3819. [Google Scholar] [CrossRef] [Green Version]
- Mitoma, H.; Hanabuchi, S.; Kim, T.; Bao, M.; Zhang, Z.; Sugimoto, N.; Liu, Y. The DHX33 RNA helicase senses cytosolic RNA and activates the NLRP3 inflammasome. Immunity 2013, 39, 123–135. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Pazhoor, S.; Bao, M.; Zhang, Z.; Hanabuchi, S.; Facchinetti, V.; Bover, L.; Plumas, J.; Chaperot, L.; Qin, J.; et al. Aspartate-glutamate-alanine-histidine box motif (DEAH)/RNA helicase A helicases sense microbial DNA in human plasmacytoid dendritic cells. Proc. Natl. Acad. Sci. USA 2010, 107, 15181–15186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshiumi, H.; Sakai, K.; Matsumoto, M.; Seya, T. DEAD/H BOX 3 (DDX3) helicase binds the RIG-I adaptor IPS-1 to up-regulate IFN-β-inducing potential. Eur. J. Immunol. 2010, 40, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Parvatiyar, K.; Zhang, Z.; Teles, R.M.; Ouyang, S.; Jiang, Y.; Iyer, S.S.; Zaver, S.A.; Schenk, M.; Zeng, S.; Zhong, W.; et al. The helicase DDX41 recognizes the bacterial secondary messengers cyclic di-GMP and cyclic di-AMP to activate a type I interferon immune response. Nat. Immunol. 2012, 13, 1155–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, C.; Tan, S.; Bao, L.; Zeng, Q.; Liu, S.; Yang, Y.; Zhong, X.; Liu, Z. DExD/H-box RNA helicase genes are differentially expressed between males and females during the critical period of male sex differentiation in channel catfish. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, W.; Song, H.; Wang, H.; Liu, J.; Zhao, H.; Du, X.; Zhang, Q. Comparative evolution of duplicated Ddx3 genes in teleosts: Insights from Japanese flounder, Paralichthys olivaceus. G3 Genes Genomes Genet. 2015, 5, 1765–1773. [Google Scholar] [CrossRef] [Green Version]
- Gan, Z.; Cheng, J.; Hou, J.; Xia, H.; Chen, W.; Xia, L.; Nie, P.; Lu, Y. Molecular and functional characterization of tilapia DDX41 in IFN regulation. Fish Shellfish Immunol. 2020, 99, 386–391. [Google Scholar] [CrossRef]
- Lauksund, S.; Svingerud, T.; Bergan, V.; Robertsen, B. Atlantic salmon IPS-1 mediates induction of IFNa1 and activation of NF-κB and localizes to mitochondria. Dev. Comp. Immunol. 2009, 33, 1196–1204. [Google Scholar] [CrossRef]
- Simora, R.M.C.; Ohtani, M.; Hikima, J.; Kondo, H.; Hirono, I.; Jung, T.S.; Aoki, T. Molecular cloning and antiviral activity of IFN-β promoter stimulator-1 (IPS-1) gene in Japanese flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2010, 29, 979–986. [Google Scholar] [CrossRef]
- Xiang, Z.; Qi, L.; Chen, W.; Dong, C.; Liu, Z.; Liu, D.; Huang, M.; Li, W.; Yang, G.; Weng, S.; et al. Characterization of a TnMAVS protein from Tetraodon nigroviridis. Dev. Comp. Immunol. 2011, 35, 1103–1115. [Google Scholar] [CrossRef]
- Kasthuri, S.R.; Wan, Q.; Whang, I.; Lim, B.; Yeo, S.; Choi, C.Y.; Lee, J. Functional characterization of the evolutionarily preserved mitochondrial antiviral signaling protein (MAVS) from rock bream, Oplegnathus fasciatus. Fish Shellfish Immunol. 2014, 40, 399–406. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, Y.; Yang, C.; Liao, L.; Wang, Y.; Su, J. Functional characterizations of IPS-1 in CIK cells: Potential roles in regulating IFN-I response dependent on IRF7 but not IRF3. Dev. Comp. Immunol. 2015, 53, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.F.; Li, S.; Lu, X.; Zhang, Y. Functions of the two zebrafish MAVS variants are opposite in the induction of IFN1 by targeting IRF7. Fish Shellfish Immunol. 2015, 45, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Xiao, T.; Jin, S.; Wang, J.; Wang, J.; Luo, H.; Li, R.; Sun, T.; Zou, J.; Li, Y. Characterization and immune function of the interferon-β promoter stimulator-1 in the barbel chub, Squaliobarbus curriculus. Dev. Comp. Immunol. 2020, 104. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, R.; Girish Babu, P.; Jeena, K.; Tripathi, G.; Pani Prasad, K. Molecular characterization, ontogeny and expression profiling of mitochondrial antiviral signaling adapter, MAVS from Asian seabass Lates calcarifer, Bloch (1790). Dev. Comp. Immunol. 2018, 79, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, R.; Jeena, K.; Mushtaq, Z.; Shyam, K.U.; Kurcheti, P.P. Antiviral activity of transiently expressed mitochondrial antiviral signaling adapter, MAVS orthologue from Asian seabass. Fish Shellfish Immunol. 2018, 76, 183–186. [Google Scholar] [CrossRef]
- Xiao, J.; Fu, Y.; Wu, H.; Chen, X.; Liu, S.; Feng, H. MAVS of triploid hybrid of red crucian carp and allotetraploid possesses the improved antiviral activity compared with the counterparts of its parents. Fish Shellfish Immunol. 2019, 89, 18–26. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, J.; Ouyang, Z.; Liu, J.; Zhang, Y.; Hu, Y.; Huang, X.; Qin, Q. Grouper MAVS functions as a crucial antiviral molecule against nervous necrosis virus infection. Fish Shellfish Immunol. 2018, 72, 14–22. [Google Scholar] [CrossRef]
- Seth, R.B.; Sun, L.; Ea, C.K.; Chen, Z.J. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-κB and IRF3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef] [Green Version]
- Jia, P.; Jin, Y.; Chen, L.; Zhang, J.; Jia, K.; Yi, M. Molecular characterization and expression analysis of mitochondrial antiviral signaling protein gene in sea perch, Lateolabrax japonicus. Dev. Comp. Immunol. 2016, 55, 188–193. [Google Scholar] [CrossRef]
- Chen, W.Q.; Hu, Y.W.; Zou, P.F.; Ren, S.S.; Nie, P.; Chang, M.X. MAVS splicing variants contribute to the induction of interferon and interferon-stimulated genes mediated by RIG-I-like receptors. Dev. Comp. Immunol. 2015, 49, 19–30. [Google Scholar] [CrossRef]
- Su, J.; Huang, T.; Yang, C.; Zhang, R. Molecular cloning, characterization and expression analysis of interferon-β promoter stimulator 1 (IPS-1) gene from grass carp Ctenopharyngodon idella. Fish Shellfish Immunol. 2011, 30, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Yan, C.; Zhou, W.; Li, J.; Wu, H.; Chen, T.; Feng, H. CARD and TM of MAVS of black carp play the key role in its self-association and antiviral ability. Fish Shellfish Immunol. 2017, 63, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zhou, J.; Lv, Y.; Qu, Y.; Chi, M.; Li, J.; Feng, H. Identification and characterization of MAVS from black carp Mylopharyngodon piceus. Fish Shellfish Immunol. 2015, 43, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.F.; Chang, M.X.; Li, Y.; Xue, N.N.; Li, J.H.; Chen, S.N.; Nie, P. NOD2 in zebrafish functions in antibacterial and also antiviral responses via NF-κB, and also MDA5, RIG-I and MAVS. Fish Shellfish Immunol. 2016, 55, 173–185. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.B.; Wu, M.; Wang, B.; Chen, C.; Gui, J.F. Fish MAVS is involved in RLR pathway-mediated IFN response. Fish Shellfish Immunol. 2014, 41, 222–230. [Google Scholar] [CrossRef]
- Kranzusch, P.J. cGAS and CD-NTase enzymes: Structure, mechanism, and evolution. Curr. Opin. Struct. Biol. 2019, 59, 178–187. [Google Scholar] [CrossRef]
- Civril, F.; Deimling, T.; de Oliveira Mann, C.C.; Ablasser, A.; Moldt, M.; Witte, G.; Hornung, V.; Hopfner, K.P. Structural mechanism of cytosolic DNA sensing by cGAS. Nature 2013, 498, 332–337. [Google Scholar] [CrossRef] [Green Version]
- Kranzusch, P.J.; Lee, A.S.; Berger, J.; Doudna, J. Structure of human cGAS reveals a conserved family of second-messenger enzymes in innate immunity. Cell Rep. 2013, 3, 1362–1368. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Zhang, X.; Jin, J.; Du, X.; Lian, T.; Yang, J.; Zhou, X.; Jiang, Z.; Su, X. Nonspecific DNA binding of cGAS N terminus promotes cGAS activation. J. Immunol. 2017, 198, 3627–3636. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ji, J.; Jiang, X.; Shao, T.; Fan, D.; Jiang, X.; Lin, A.; Xiang, L.; Shao, J. Characterization of cGAS homologs in innate and adaptive mucosal immunities in zebrafish gives evolutionary insights into cGAS-STING pathway. FASEB J. 2020, 34, 7786–7809. [Google Scholar] [CrossRef] [Green Version]
- Ablasser, A.; Chen, Z.J. cGAS in action: Expanding roles in immunity and inflammation. Science 2019, 363, eaat8657. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, Z.J. The cGAS-cGAMP-STING pathway connects DNA damage to inflammation, senescence, and cancer. J. Exp. Med. 2018, 215, 1287–1299. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.; Jiang, W.; Hao, J. Research Advances in How the cGAS-STING pathway controls the cellular inflammatory response. Front. Immunol. 2020, 11, 615. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Whiteley, A.T.; de Oliveira Mann, C.C.; Morehouse, B.R.; Nowak, R.P.; Fischer, E.S.; Gray, N.S.; Mekalanos, J.J.; Kranzusch, P.J. Structure of the human cGAS-DNA complex reveals enhanced control of immune surveillance. Cell 2018, 174, 300–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luecke, S.; Holleufer, A.; Christensen, M.H.; Jønsson, K.L.; Boni, G.A.; Sørensen, L.K.; Johannsen, M.; Jakobsen, M.R.; Hartmann, R.; Paludan, S.R. cGAS is activated by DNA in a length-dependent manner. EMBO Rep. 2017, 18, 1707–1715. [Google Scholar] [CrossRef]
- García-Belmonte, R.; Perez-Nunez, D.; Pittau, M.; Richt, J.A.; Revilla, Y. African swine fever virus armenia/07 virulent strain controls interferon beta production through the cGAS-STING pathway. J. Virol. 2019, 93, e02298-18. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wu, J.; Du, F.; Chen, X.; Chen, Z.J. Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 2013, 339, 786–791. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; You, H.; Su, C.; Li, Y.; Chen, S.; Zheng, C. Herpes simplex virus 1 tegument protein VP22 abrogates cGAS/STING-mediated antiviral innate immunity. J. Virol. 2018, 92, e00841-18. [Google Scholar] [CrossRef] [Green Version]
- Verrier, E.R.; Yim, S.A.; Heydmann, L.; El-Saghire, H.; Bach, C.; Turon-Lagot, V.; Mailly, L.; Durand, S.C.; Lucifora, J.; Durantel, D.; et al. Hepatitis B virus evasion from cyclic guanosine monophosphate-adenosine monophosphate synthase sensing in human hepatocytes. Hepatology 2018, 68, 1695–1709. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, S.; Fernandez-Sesma, A. Collateral damage during dengue virus Infection: Making sense of DNA by cGAS. J. Virol. 2017, 91, e01081-16. [Google Scholar] [CrossRef] [Green Version]
- Marinho, F.V.; Benmerzoug, S.; Rose, S.; Campos, P.C.; Marques, J.T.; Báfica, A.; Barber, G.; Ryffel, B.; Oliveira, S.C.; Quesniaux, V.F.J. The cGAS/STING pathway is important for dendritic cell activation but is not essential to induce protective immunity against Mycobacterium tuberculosis infection. J. Innate Immun. 2018, 10, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.C.; Cai, H.; Li, T.; Franco, L.H.; Li, X.D.; Nair, V.R.; Scharn, C.R.; Stamm, C.E.; Levine, B.; Chen, Z.J.; et al. Cyclic GMP-AMP synthase is an innate immune DNA sensor for Mycobacterium tuberculosis. Cell Host Microbe 2015, 17, 820–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, R.O.; Bell, S.L.; MacDuff, D.A.; Kimmey, J.M.; Diner, E.J.; Olivas, J.; Vance, R.E.; Stallings, C.L.; Virgin, H.W.; Cox, J.S. The cytosolic sensor cGAS detects Mycobacterium tuberculosis DNA to induce type I interferons and activate autophagy. Cell Host Microbe 2015, 17, 811–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandakumar, R.; Tschismarov, R.; Meissner, F.; Prabakaran, T.; Krissanaprasit, A.; Farahani, E.; Zhang, B.C.; Assil, S.; Martin, A.; Bertrams, W.; et al. Intracellular bacteria engage a STING-TBK1-MVB12b pathway to enable paracrine cGAS-STING signaling. Nat. Microbiol. 2019, 4, 701–713. [Google Scholar] [CrossRef]
- Ruiz-Moreno, J.S.; Hamann, L.; Jin, L.; Sander, L.E.; Puzianowska-Kuznicka, M.; Cambier, J.; Witzenrath, M.; Schumann, R.R.; Suttorp, N.; Opitz, B. The cGAS/STING pathway detects Streptococcus pneumoniae but appears dispensable for antipneumococcal defense in mice and humans. Infect. Immun. 2017, 86, e00849-17. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Marin, C.; Schrum, J.E.; Andrade, W.A.; Shaffer, S.A.; Giraldo, L.F.; Lasso, A.M.; Kurt-Jones, E.A.; Fitzgerald, K.A.; Golenbock, D.T. Cyclic GMP–AMP synthase is the cytosolic sensor of Plasmodium falciparum genomic DNA and activates type I IFN in malaria. J. Immunol. 2018, 200, 768–774. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Kumar, A.; Mandal, A.; Abhishek, K.; Verma, S.; Kumar, A.; Das, P. Nucleic acid sensing activates the innate cytosolic surveillance pathway and promotes parasite survival in visceral leishmaniasis. Sci. Rep. 2019, 9, 9825. [Google Scholar] [CrossRef] [Green Version]
- Ge, R.; Zhou, Y.; Peng, R.; Wang, R.; Li, M.; Zhang, Y.; Zheng, C.; Wang, C. Conservation of the STING-mediated cytosolic DNA sensing pathway in zebrafish. J. Virol. 2015, 89, 7696–7706. [Google Scholar] [CrossRef] [Green Version]
- Rolf, J.; Siedentop, R.; Lütz, S.; Rosenthal, K. Screening and identification of novel cGAS homologues using a combination of in vitro and in vivo protein synthesis. Int. J. Mol. Sci. 2019, 21, 105. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Lu, L.; Lu, X.; Li, S.; Zhang, Y. Grass carp cGASL negatively regulates fish IFN response by targeting MITA. Fish Shellfish Immunol. 2019, 94, 871–879. [Google Scholar] [CrossRef]
- Su, C.; Zheng, C. Herpes Simplex Virus 1 abrogates the cGAS/STING-mediated cytosolic DNA sensing pathway via its virion host Shutoff Protein, UL41. J. Virol. 2017, 91, e02414-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, P.; Wang, W.; Yang, N.; Serna-Tamayo, C.; Ricca, J.M.; Zamarin, D.; Shuman, S.; Merghoub, T.; Wolchok, J.D.; Deng, L. Intratumoral delivery of inactivated modified vaccinia virus ankara (iMVA) induces systemic antitumor immunity via STING and batf3-dependent dendritic cells. Sci. Immunol. 2017, 2, eaal1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, M.K.; Nandakumar, R.; Stadler, D.; Malo, A.; Marin-Valls, R.; Wang, F.; Reinert, L.S.; Dagnæs-Hansen, F.; Hollensen, A.K.; Mikkelsen, J.G.; et al. Lack of immunological DNA sensing in hepatocytes facilitates hepatitis B virus infection. Hepatology 2016, 64, 746–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, E.B.; Montoya, B.; Ferez, M.; Stotesbury, C.; Sigal, L.J. Resistance to ectromelia virus infection requires cGAS in bone marrow-derived cells which can be bypassed with cGAMP therapy. PLoS Pathog. 2019, 15, e1008239. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; He, X.; Jia, H.; Chen, G.; Jin, Q.; Long, Z.; Jing, Z. The cGAS-STING signaling pathway is required for the innate immune response against ectromelia virus. Front. Immunol. 2018, 9, 1297. [Google Scholar] [CrossRef] [Green Version]
- Paijo, J.; Döring, M.; Spanier, J.; Grabski, E.; Nooruzzaman, M.; Schmidt, T.; Witte, G.; Messerle, M.; Hornung, V.; Kaever, V.; et al. cGAS senses human cytomegalovirus and induces type I interferon responses in human monocyte-derived cells. PLoS Pathog. 2016, 12, e1005546. [Google Scholar] [CrossRef]
- Gao, D.; Wu, J.; Wu, Y.T.; Du, F.; Aroh, C.; Yan, N.; Sun, L.; Chen, Z.J. Cyclic GMP-AMP synthase is an innate immune sensor of HIV and other retroviruses. Science 2013, 341, 903–906. [Google Scholar] [CrossRef] [Green Version]
- Lieb, B.; Carl, M.; Hock, R.; Gebauer, D.; Scheer, U. Identification of a novel mRNA-associated protein in oocytes of Pleurodeles waltl and Xenopus laevis. Exp. Cell Res. 1998, 245, 272–281. [Google Scholar] [CrossRef]
- Tanaka, K.J.; Ogawa, K.; Takagi, M.; Imamoto, N.; Matsumoto, K.; Tsujimoto, M. RAP55, a cytoplasmic mRNP component, represses translation in Xenopus oocytes. J. Biol. Chem. 2006, 281, 40096–40106. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Yu, J.H.; Gulick, T.; Bloch, K.D.; Bloch, D.B. RNA-associated protein 55 (RAP55) localizes to mRNA processing bodies and stress granules. RNA 2006, 12, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, R.; Zhou, Q.; Xu, Z.; Li, C.; Wang, S.; Mao, A.; Zhang, X.; He, W.; Shu, H.B. LSm14A is a processing body-associated sensor of viral nucleic acids that initiates cellular antiviral response in the early phase of viral infection. Proc. Natl. Acad. Sci. USA 2012, 109, 11770–11775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, G.; Zhang, Y.; Zhang, X.; Hu, H.; Zhang, H.; Cheng, C.; Wang, X.; Li, X.; Fang, W. Functional characterization of porcine LSm14A in IFN-β induction. Vet. Immunol. Immunopathol. 2013, 155, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, R.; Zhao, J.; Qi, Z.; Ji, L.; Zhen, Y.; Liu, B. LSM14A inhibits porcine reproductive and respiratory syndrome virus (PRRSV) replication by activating IFN-β signaling pathway in Marc-145. Mol. Cell. Biochem. 2015, 399, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yang, Q.; Li, M.; Zhong, B.; Ran, Y.; Liu, L.; Yang, Y.; Wang, Y.; Shu, H. LSm14A plays a critical role in antiviral immune responses by regulating MITA level in a cell-specific manner. J. Immunol. 2016, 196, 5101–5111. [Google Scholar] [CrossRef] [PubMed]
- Hua, K.; Li, H.; Chen, H.; Foda, M.F.; Luo, R.; Jin, H. Functional characterization of duck LSm14A in IFN-β induction. Dev. Comp. Immunol. 2017, 76, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Wu, C.; Wen, G.; Li, C. Transcriptional response of LSm14A after infection of blue eggshell layers with Newcastle disease viruses. J. Vet. Med. Sci. 2019, 81, 1468–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.H.; Asano, A.; Kon, Y.; Watanabe, T.; Agui, T. Characterization of the chicken PKR: Polymorphism of the gene and antiviral activity against vesicular stomatitis virus. Jpn. J. Vet. Res. 2004, 51, 123–133. [Google Scholar] [CrossRef]
- Meurs, E.; Chong, K.; Galabru, J.; Thomas, N.S.B.; Kerr, I.M.; Williams, R.G.; Hovanessian, A. G Molecular cloning and characterization of the human double-stranded RNA-activated protein kinase induced by interferon. Cell 1990, 62, 379–390. [Google Scholar] [CrossRef]
- Rothenburg, S.; Deigendesch, N.; Dey, M.; Dever, T.E.; Tazi, L. Double-stranded RNA-activated protein kinase PKR of fishes and amphibians: Varying the number of double-stranded RNA binding domains and lineage-specific duplications. BMC Biol. 2008, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Zhu, R.; Zhang, Y.B.; Zhang, Q.Y.; Gui, J.F. Functional domains and the antiviral effect of the double-stranded RNA-dependent protein kinase PKR from Paralichthys olivaceus. J. Virol. 2008, 82, 6889–6901. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.A.; Gil, J.; Ventoso, I.; Guerra, S.; Domingo, E.; Rivas, C.; Esteban, M. Impact of protein kinase PKR in cell biology: From antiviral to antiproliferative action. Microbiol. Mol. Biol. Rev. 2006, 70, 1032–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.Y.; Zhang, Y.B.; Huang, G.P.; Zhang, Q.Y.; Gui, J.F. Molecular cloning and characterisation of a fish PKR-like gene from cultured CAB cells induced by UV inactivated virus. Fish Shellfish Immunol. 2004, 17, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Rothenburg, S.; Deigendesch, N.; Dittmar, K.; Koch-Nolte, F.; Haag, F.; Lowenhaupt, K.; Rich, A. A PKR-like eukaryotic initiation factor 2a kinase from zebrafish contains Z-DNA binding domains instead of dsRNA binding domains. Proc. Natl. Acad. Sci. USA 2005, 102, 1602–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.B.; Gui, J.F. Molecular regulation of interferon antiviral response in fish. Dev. Comp. Immunol. 2012, 38, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, T.; Rould, M.A.; Lowenhaupt, K.; Herbert, A.; Rich, A. Crystal structure of the Zα domain of the human editing enzyme ADAR1 bound to left-handed Z-DNA. Science 1999, 284, 1841–1845. [Google Scholar] [CrossRef] [PubMed]
- Bergan, V.; Jagus, R.; Lauksund, S.; Kileng, Ø.; Robertsen, B. The Atlantic salmon Z-DNA binding protein kinase phosphorylates translation initiation factor 2 alpha and constitutes a unique orthologue to the mammalian dsRNA-activated protein kinase R. FEBS J. 2007, 275, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.K.; Zhang, Y.B.; Liu, Y.; Sun, F.; Gui, J.F. Cooperative roles of fish protein kinase containing Z-DNA binding domains and double-stranded RNA-dependent protein kinase in interferon-mediated antiviral response. J. Virol. 2011, 85, 12769–12780. [Google Scholar] [CrossRef] [Green Version]
- Herbert, A.; Alfken, J.; Kim, Y.G.; Mian, I.S.; Nishikura, K.; Rich, A. A Z-DNA binding domain present in the human editing enzyme, double-stranded RNA adenosine deaminase. Proc. Natl. Acad. Sci. USA 1997, 94, 8421–8426. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, T.; Behlke, J.; Lowenhaupt, K.; Heinemann, U.; Rich, A. Structure of the DLM-1-Z-DNA complex reveals a conserved family of Z-DNA-binding proteins. Nat. Struct. Biol. 2001, 8, 761–765. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, Y.; Hu, C. PKZ, a Fish-Unique eIF2α Kinase Involved in Innate Immune Response. Front. Immunol. 2020, 11, 585. [Google Scholar] [CrossRef]
- Hu, Y.S.; Li, W.; Li, D.M.; Liu, Y.; Fan, L.H.; Rao, Z.C.; Lin, G.; Hu, C.Y. Cloning, expression and functional analysis of PKR from grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2013, 35, 1874–1881. [Google Scholar] [CrossRef] [PubMed]
- Del Castillo, C.S.; Hikima, J.; Ohtani, M.; Jung, T.S.; Aoki, T. Characterization and functional analysis of two PKR genes in fugu (Takifugu rubripes). Fish Shellfish Immunol. 2012, 32, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Zenke, K.; Nam, Y.K.; Kim, K.H. Molecular cloning and expression analysis of double-stranded RNA-dependent protein kinase (PKR) in rock bream (Oplegnathus fasciatus). Vet. Immunol. Immunopathol. 2010, 133, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Perelygin, A.A.; Lear, T.L.; Zharkikh, A.A.; Brinton, M.A. Comparative analysis of vertebrate EIF2AK2 (PKR) genes and assignment of the equine gene to ECA15q24-q25 and the bovine gene to BTA11q12-q15. Genet. Sel. Evol. 2006, 38, 551–563. [Google Scholar] [CrossRef]
- Su, J.; Zhu, Z.; Wang, Y. Molecular cloning, characterization and expression analysis of the PKZ gene in rare minnow Gobiocypris rarus. Fish Shellfish Immunol. 2008, 25, 106–113. [Google Scholar] [CrossRef]
- Yang, P.J.; Wu, C.X.; Li, W.; Fan, L.H.; Lin, G.; Hu, C.Y. Cloning and functional analysis of PKZ (PKR-like) from grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2011, 31, 1173–1178. [Google Scholar] [CrossRef]


{kind=link}
| Species | Cell Line/Tissue | Infection/Treatment | References |
|---|---|---|---|
| rig-I | |||
| Atlantic salmon (Salmo salar) | In vitro: TO cell line | salmon alphavirus subtype 3 (SAV-3), infectious pancreatic necrosis virus (IPNV), infectious salmon anaemia virus (ISAV) | [59,60] |
| Ex vivo: erythrocytes | piscine orthoreovirus (PRV) | [61] | |
| In vivo: head kidney | infectious pancreatic necrosis virus (IPNV) | [62] | |
| Channel catfish (Ictalurus punctatus) | In vitro: CCO cells line | channel catfish virus (CCV) | [63] |
| In vivo: liver | Edwardsiella ictaluri | [63] | |
| Common carp (Cyprinus carpio) | In vivo: spleen, head kidney, intestine | spring viremia of carp virus (SVCV) | [64] |
| Gibel carp (Carassius gibelio) | In vivo: head kidney, spleen, liver | Carassius auratus herpesvirus (CaHV) | [65] |
| Grass carp (Ctenopharyngodon idella) | In vitro: CIK cell line | grass carp reovirus (GCRV), poly(I:C) | [66] |
| In vitro: primary trunk kidney cells | grass carp reovirus (GCRV), poly(I:C), LPS, PGN | [67] | |
| In vivo: spleen, liver, trunk kidney, hepatopancreas | grass carp reovirus (GCRV) | [66,67,68] | |
| Japanese eel (Anguilla japonica) | In vivo: liver, spleen, kidney | poly(I:C), LPS, Aeromonas hydrophila | [69] |
| In vivo: liver, spleen, head kidney, skin, intestine, and gills | poly(I:C) | [70] | |
| Zebrafish (Danio rerio) | In vitro: ZF4 cell line | nervous necrosis virus (NNV), snakehead fish vesiculovirus (SHVV), spring viremia of carp virus (SVCV), Edwardsiella tarda | [71,72,73] |
| In vivo: larvae | spring viremia of carp virus (SVCV), poly(I:C) | [74,75] | |
| In vivo: visceral tissues | poly(I:C) | [76] | |
| In vivo: spleen, kidney | tilapia lake virus (TiLV) | [77] | |
| mda5 | |||
| Atlantic salmon (Salmo salar) | In vitro: TO cell line | salmon alphavirus subtype 3 (SAV-3), infectious pancreatic necrosis virus (IPNV), infectious salmon anaemia virus (ISAV), salmonid alphavirus | [59,60,78] |
| In vivo: head kidney | infectious pancreatic necrosis virus (IPNV) | [62] | |
| Black carp (Mylopharyngodon piceus) | In vitro: MPF cell line | spring viremia of carp virus (SVCV), grass carp Reovirus (GCRV), poly(I:C) | [79] |
| In vivo: heart, liver, spleen, kidney, intestine, skin, gill | grass carp reovirus (GCRV), spring viremia of carp virus SVCV | [79] | |
| Asian seabass (Lates calcarifer) | In vitro: SISK cell line | poli (I:C), LPS, PGN | [80] |
| In vivo: spleen, kidney, gills, heart, liver, intestine | poli (I:C) | ||
| In vivo: gills, heart, kidney, liver, intestine | Vibrio alginolyticus | ||
| In vivo: gills, heart, kidney, liver, intestine | Staphylococcus aureus | ||
| Barbel chub (Squaliobarbus curriculus) | In vivo: liver | grass carp reovirus (GCRV) | [81] |
| Channel catfish (Ictalurus punctatus) | In vitro: CCO cell line | channel catfish virus (CCV) | [63] |
| In vivo: liver | Edwardsiella ictaluri. | [63] | |
| Common carp (Cyprinus carpio) | In vitro: peripheral blood leukocytes | poly(I:C) | [82] |
| In vivo: liver, spleen, head kidney, foregut, hindgut, gills, skin | poly(I:C), Aeromonas hydrophila | [82] | |
| Grass carp (Ctenopharyngodon idella) | In vitro: primarly trunk kidney cells | grass carp reovirus (GCRV), poly(I:C), LPS, PGN | [67] |
| In vivo: trunk kidney, spleen, liver, hepatopancreas | grass carp reovirus (GCRV) | [67,68,83] | |
| Green chromide (Etroplus suratensis) | In vivo: spleen, kidney, liver, intestine, gills, heart | poly(I:C) | [84] |
| Japanese flounder (Olive flounder) (Paralichthys olivaceus) | In vitro: peripheral blood leukocytes, kidney leukocytes | poly(I:C), LPS | [85] |
| In vivo: kidney | viral hemorrhagic septicemia virus (VHSV) | [85] | |
| In vitro: HINEA cell line | poly(I:C) | [86] | |
| Mandrinfish (Siniperca chuatsi) | In vivo: spleen, gills, head kidney | poly(I:C), LPS | [87] |
| Orange-spotted groupers (Epinephelus coioides) | In vivo: spleen | Singapore grouper iridovirus (SGIV), poly(I:C) | [88] |
| Rainbow trout (Oncorhynchus mykiss) | In vitro: RTG-2 cell line, RTS-11 cell lines | poly(I:C), recombinant trout IFN2 protein | [78] |
| In vivo: head kidney | viral hemorrhagic septicemia virus (VHSV) | [78] | |
| Sea perch (Lateolabrax japonicas) | In vitro: LJB cell line, LJF cell line | Poly(I:C), nervous necrosis virus (NNV) | [89] |
| In vivo: Spleen, kidney, eye, thymus, brain, intestine, muscle, gill, liver, heart | nervous necrosis virus (NNV) | ||
| Large yellow croaker (Pseudosciaena crocea) | In vivo: peripheral blood, liver, spleen and head kidney | poly(I:C) | [90,91] |
| Zebrafish (Danio rerio) | In vitro: ZF4 cell line | spring viremia of carp virus (SVCV), nervous necrosis virus (NNV), snakehead fish vesiculovirus (SHVV), Edwardsiella tarda | [71,72,92] |
| lgp2 | |||
| Asian seabass (Lates calcarifer) | In vitro: SISK cell line | poly(I:C), LPS | [93] |
| In vivo: liver, spleen, kidney, gill, heart, intestine | poly(I:C), Vibrio alginolyticus, Staphylococcus aureus | [93] | |
| Atlantic salmon (Salmo salar) | In vitro: TO cell line | infectious pancreatic necrosis virus (IPNV), infectious salmon anaemia virus (ISAV), salmonid alphavirus (SAV) | [60,78] |
| Atlantic cod (Gadus morhua) | In vivo: head kidney | infectious pancreatic necrosis virus (IPNV) | [94] |
| Black carp (Mylopharyngodon piceus) | In vitro: MPF cell line | spring viremia of carp virus (SVCV), grass carp reovirus (GCRV), poly(I:C) | [95] |
| In vivo: liver, spleen, kidney, intestine, heart, muscle, skin | spring viremia of carp virus (SVCV), grass carp reovirus (GCRV) | [95] | |
| Common carp (Cyprinus carpio) | In vivo: muscle, spleen, gill, brain, skin, heart, intestine, liver, head kidney | koi herpes virus (KHV) | [96] |
| Channel catfish (Ictalurus punctatus) | In vitro: CCO cell line | channel catfish virus (CCV) | [63] |
| In vivo: liver | Edwardsiella ictaluri | [63] | |
| Fathead minnow (Pimephales promelas) | In vitro: EPC cell line | koi herpes virus, (KHV), poly(I:C) | [96] |
| Gibel carp (Carassius gibelio) | In vivo: head kidney, spleen, liver | Carassius auratus herpesvirus (CaHV) | [65] |
| Grass carp (Ctenopharyngodon idella) | In vitro: primary trunk kidney cells | grass carp reovirus (GCRV), poly(I:C), LPS, PGN | [67] |
| In vivo: trunk kidney, spleen, liver, hepatopancreas | grass carp reovirus (GCRV) | [58,67,68] | |
| Indian major carp (Labeo rohita) | In vitro: LRG cell line | poly(I:C), iE-DAP, MDP | [97] |
| In vivo: liver, spleen, kidney, blood, gill | poly (I:C), Aeromonas hydrophila, Bacillus subtilis | [97] | |
| Japanese flounder (Olive flounder) (Paralichthys olivaceus) | In vitro: leukocytes isolated from kidney | poly(I:C), LPS | [57] |
| In vivo: kidney | viral hemorrhagic septicemia virus (VHSV) | [57] | |
| In vitro: HINEA cell line | poly(I:C) | [86] | |
| Large yellow croaker (Pseudosciaena crocea) | In vivo: peripheral blood, liver, spleen, head kidney | poly(I:C) | [91] |
| Mandrinfish (Siniperca chuatsi) | In vivo: spleen, gills, head kidney | poly(I:C), LPS | [87] |
| Miiuy croaker (Miichthys miiuy) | In vitro: macrophages In vivo: liver, spleen, kidney | poly(I:C) | [98] |
| Orange-spotted groupers (Epinephelus coioides) | In vivo: spleen | Singapore grouper iridovirus (SGIV), poly(I:C) | [99] |
| Rainbow trout (Oncorhynchus mykiss) | In vitro: RTG-2 cell line, RTS-11 cell lines | poly(I:C), recombinant trout IFN2 protein | [78,100] |
| In vivo: head kidney | viral hemorrhagic septicemia virus (VHSV) | [78] | |
| Sea perch (Lateolabrax japonicas) | In vitro: LJF cell line, LJH cell line, LJB cell line | poly(I:C), nervous necrosis virus NNV | [101] |
| In vivo: liver, heart, intestines, gill, spleen, muscle, brain, kidney, thymus, eye | nervous necrosis virus (NNV) | [101] | |
| Zebrafish (Danio rerio) | In vitro: ZF4 and ZFL cell lines | poly(I:C), nervous necrosis virus (NNV), snakehead fish vesiculovirus (SHVV) | [71,72,102] |
| In vivo: gill, liver, spleen, head kidney, and body kidney | spring viremia of carp virus (SVCV) | [102] | |
| ddx1 | |||
| Common carp (Cyprinus carpio) | In vivo: head kidney | spring viremia of carp virus (SVCV) | [103] |
| Zebrafish (Danio rerio) | In vivo: kidney, spleen | spring viremia of carp virus (SVCV), chum salmon reovirus (CSV) | [103] |
| ddx3 | |||
| Orange-spotted grouper (Epinephelus coioides) | In vitro: GS cell line | poly(I:C), red-spotted grouper nervous necrosis virus RGNNV) | [104] |
| Rainbow trout (Oncorhynchus mykiss) | In vitro: RTG-2 cell line | poly(I:C) | [100] |
| Zebrafish (Danio rerio) | In vitro: ZF4 cell line | chum salmon reovirus (CSV) | [103] |
| dhx9 | |||
| Common carp (Cyprinus carpio) | In vivo: head kidney | spring viremia of carp virus (SVCV) | [103] |
| Japanese medeka (Oryzias latipes) | In vivo: spleen, kidney, intestine | CpG mixture, FKC of Edwardsiella tarda | [105] |
| Rainbow trout (Oncorhynchus mykiss) | In vitro: RTG-2 cell line | poly(I:C) | [100] |
| Zebrafish (Danio rerio) | In vitro: ZF4 cell line | chum salmon reovirus (CSV) | [103] |
| In vivo: spleen | chum salmon reovirus (CSV) | [103] | |
| ddx21 | |||
| Common carp (Cyprinus carpio) | In vivo: head kidney | spring viremia of carp virus (SVCV) | [103] |
| Zebrafish (Danio rerio) | In vitro: ZF4 cell line | chum salmon reovirus (CSV) | [103] |
| dhx36 | |||
| Japanese medeka (Oryzias latipes) | In vivo: spleen, kidney, intestine | CpG-mixture, FKC of Edwardsiella tarda | [105] |
| Zebrafish (Danio rerio) | In vitro: ZF4 cell line | chum salmon reovirus (CSV) | [103] |
| In vivo: kidney | spring viremia of carp virus (SVCV) | ||
| ddx41 | |||
| Japanese flounder (Olive flounder) (Paralichthys olivaceus) | In vitro: adherent (monocyte-like) and non-adherent (lymphocyte- enriched) cells | ranavirus | [106] |
| In vivo: spleen, kidney, liver, heart, gill | lymphocystis disease virus (LCDV) | [106] | |
| Orange-spotted grouper (Epinephelus coioides) | In vitro: GS cell line | Singapore grouper iridovirus (SGIV), red-spotted grouper nervous necrosis virus (RGNNV) | [107] |
| Pathogens | Species | Function/Mechanism | References |
|---|---|---|---|
| DNA viruses | |||
| African swine fever virus (ASFV) | Pig (Wild boar) (Sus scrofa domesticus) | ASFV occurs in attenuated and virulent forms. When avian alveolar macrophages are infected with attenuated ASFV, cGAS senses viral DNA and induces IFNβ. On the other hand, the virulent strain of ASFV strongly inhibits IFNβ via the cGAS-STING pathway by suppressing the downstream cascade of cGAS. | [166] |
| Herpes simplex virus 1 (HSV-1) | Mouse (Mus musculus) | Knockdown of cGAS by shRNA in murine fibrosarcoma cell line L929 strongly inhibits IRF3 dimerization induced by HSV-1 infection. | [167] |
| Human (Homo sapiens) | HSV-1 tegument protein UL41 prevents cGAS DNA sensing by degrading cGAS mRNA. | [181] | |
| HSV-1 segment protein VP22 binds to cGAS protein and directly acts on the cGAS protein to degrade it. | [168] | ||
| Zebrafish (Danio rerio) | The knockdown of cGAS did not cause an obvious effect on the induction of IFN-φ1, ISG15, and viperin in zebrafish infected with HSV-1. | [178] | |
| Vaccinia virus Ankara (MVA) | Human, Mouse | Infection of human and mouse dendritic cells (DCs) with heat- or UV-inactivated MVA has been shown to induce higher levels of IFNs through the cGAS-STING pathway other than wild type-MVA. | [182] |
| MVA DNAs in the cytosol are sensed by cGAS, what leads to activation of STING and downstream transcription factors, IRF3 and IRF7, resulting in the activation of type I IFN gene expression. | [182] | ||
| Chronic hepatitis B virus (HBV) | Human, Mouse | HBV rcDNA (rcDNA; a precursor of ccc) is sensed by cGAS in the cytoplasm of hepatocytes, but may form a viral capsid that covers the DNA and escapes from cGAS sensing. HBV cccDNA was increased in cGAS knockout cells and decreased in cGAS overexpressing cells. | [169] |
| Constitutive low expression of cGAS-STING in the liver may explain liver-specific HBV infection and a weak capacity of hepatocyte cells to clear HBV infection. | [183] | ||
| Ectromelia virus (ECTV) | Mouse | Cells originated from bone marrow are the main type I IFN producers, required for ISG expression. | [184] |
| In addition to TLR9, type I IFN production stimulated by the cGAS-STING pathway is also important for survival of mice after ECTV infection. | [185] | ||
| Human cytomegalovirus (HCMV) | Human | cGAS recognizes HCMV DNA and induces type I IFN in human monocyte-derived plasmacytoid dendritic cells and macrophages. | [186] |
| RNA viruses (ssRNA) | |||
| Human immunodeficiency virus (HIV) | Human, Mouse | cGAS senses reverse transcribed HIV DNA in the cytoplasm and induces cGAMP-STING-dependent IFNβ production. Knockout or knockdown of cGAS in mouse or human cell lines blocks cytokine induction by HIV, MLV and SIV. | [187] |
| Murine leukemia virus (MLV) | |||
| Simian immunodeficiency virus (SIV) | |||
| Dengue virus (DENV) | Human | DENV infects cells and localizes to endoplasmic reticulum (ER) and mitochondrial membrane. Mitochondria are disrupted by infection stress, and cGAS senses mtDNA leaked into the cytoplasm. | [170] |
| Intracellular bacteria | |||
| Mycobacterium tuberculosis | Human, mouse | The cGAS-STING pathway appears to activate dendritic cells by sensing Mycobacterium tuberculosis (Mtb) DNA in the cytoplasm, but does not contribute to host protection in vivo (lung cells). | [171] |
| cGAS induces production of type I IFN and promotes early regulation of intracellular replication by inducing autophagy. | [172] | ||
| Sensing Mtb DNA via the cGAS-STING pathway induces type I IFN and autophagy. | [173] | ||
| Lysteria monocytogenes | Human | Cell infected with L. monocytogenes DNA is carried to the extracellular vesicles (EV) of infected cells and delivered to bystander cells to stimulate the cGAS-STING pathway. | [174] |
| Edwardsiella tarda (E. piscicida) | Japanese medaka (Oryzias latipes) | E. tarda infection in medaka in vivo upregulated cgas gene expression. | [105] |
| Zebrafish (Danio rerio) | DrcGASa contributed to IgZ/IgZ2 induction in response to E tarda infect/ion by upregulating ifnφ1 expression in zebrafish gill γδ T cells. | [160] | |
| Extracellular bacteria | |||
| Streptococcus pneumoniae | Human, Mouse | The type I IFN response induced by mouse S. pneumoniae is highly dependent on the cGAS-STING pathway. | [175] |
| Parasites | |||
| Malaria, Plasmodium falciparum | Human | cGAS is the cytosolic sensor of P. falciparum DNA and is required for the induction of IFNβ by malaria hemozoin (Hz) as carrier of P. falciparum gDNA. | [176] |
| Leishmania donovani | Human | cGAS dependent targeting of L. donovani DNA induces IFN-β over-production that contributes to antimony resistance in L. donovani infection. | [177] |
| Fish Species | Superorder/Order | Features/Function | References |
|---|---|---|---|
| PKR | |||
| Goldfish (Carassius auratus) | Ostariophysi/Cypriniformes | Tandem arrangement of PKR and PKZ genes Induction of PKR mRNA by IFN-stimulation Phosphorylation of eIF2α Inhibition of translation Interaction with poly(I:C) | [207] |
| Grass carp (Ctenopharyngodon idella) | Ostariophysi/Cypriniformes | Induction of PKR mRNA by GCHV injection Inhibition of translation | [211] |
| Japanese flounder (Olive flounder) (Paralichthys olivaceus) | Acanthopterygii/Pleuronectiformes | Induction of PKR mRNA by SMRV injection Inhibition of translation Phosphorylation of eIF2α in response to SMRV infection | [200] |
| Japanese pufferfish (torafugu) (Takifugu rubripes) | Acanthopterygii/Tetraodontiformes | Tandem duplication of two PKR genes; Induction of PKR1 mRNA by poly(I:C) stimulation Inhibition of translation and NF-κB activation by PKR1 and PKR2 | [199,212] |
| Rock bream (Barred knifejaw) (Oplegnathus fasciatus) | Acanthopterygii/Perciformes | Induction of PKR mRNA by poly(I:C) | [213] |
| Spotted green pufferfish (Tetraodon nigroviridis) | Acanthopterygii/Tetraodontiformes | Tandem duplication of three PKR genes | [199] |
| Zebrafish (Danio rerio) | Ostariophysi/Cypriniformes | Tandem arrangement of PKR and PKZ genes Phosphorylation of eIF2α | [199,207,214] |
| PKZ | |||
| Atlantic salmon (Salmo salar) | Protacanthopterygii/Salmoniformes | Induction of PKZ mRNA by IFN-stimulation Phosphorylation of eIF2α by PKZ-stimulated with Z-DNA in vitro | [206] |
| Chinese rare minnow (Gobiocypris rarus) | Ostariophysi/Cypriniformes | Induction of PKZ mRNA by GCRV and Aeromonas hydrophila infection | [215] |
| Goldfish (Carassius auratus) | OstariophysiCypriniformes | Induction of PKZ mRNA by IFN-stimulation; | [202,207] |
| Grass carp (Ctenopharyngodon idella) | Ostariophysi/Cypriniformes | Phosphorylation of eIF2α by PKZ in vitro Inhibition of translation | [216] |
| Zebrafish (Danio rerio) | Ostariophysi/Cypriniformes | Specific binding of Zα domains to Z-DNA Inhibition of translation Phosphorylation of eIF2α by both PKR and PKZ | [199,203] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mojzesz, M.; Rakus, K.; Chadzinska, M.; Nakagami, K.; Biswas, G.; Sakai, M.; Hikima, J.-i. Cytosolic Sensors for Pathogenic Viral and Bacterial Nucleic Acids in Fish. Int. J. Mol. Sci. 2020, 21, 7289. https://doi.org/10.3390/ijms21197289
Mojzesz M, Rakus K, Chadzinska M, Nakagami K, Biswas G, Sakai M, Hikima J-i. Cytosolic Sensors for Pathogenic Viral and Bacterial Nucleic Acids in Fish. International Journal of Molecular Sciences. 2020; 21(19):7289. https://doi.org/10.3390/ijms21197289
Chicago/Turabian StyleMojzesz, Miriam, Krzysztof Rakus, Magdalena Chadzinska, Kentaro Nakagami, Gouranga Biswas, Masahiro Sakai, and Jun-ichi Hikima. 2020. "Cytosolic Sensors for Pathogenic Viral and Bacterial Nucleic Acids in Fish" International Journal of Molecular Sciences 21, no. 19: 7289. https://doi.org/10.3390/ijms21197289