Epidemiologic, Phenotypic, and Structural Characterization of Aminoglycoside-Resistance Gene aac(3)-IV


Abstract
:1. Introduction
2. Results
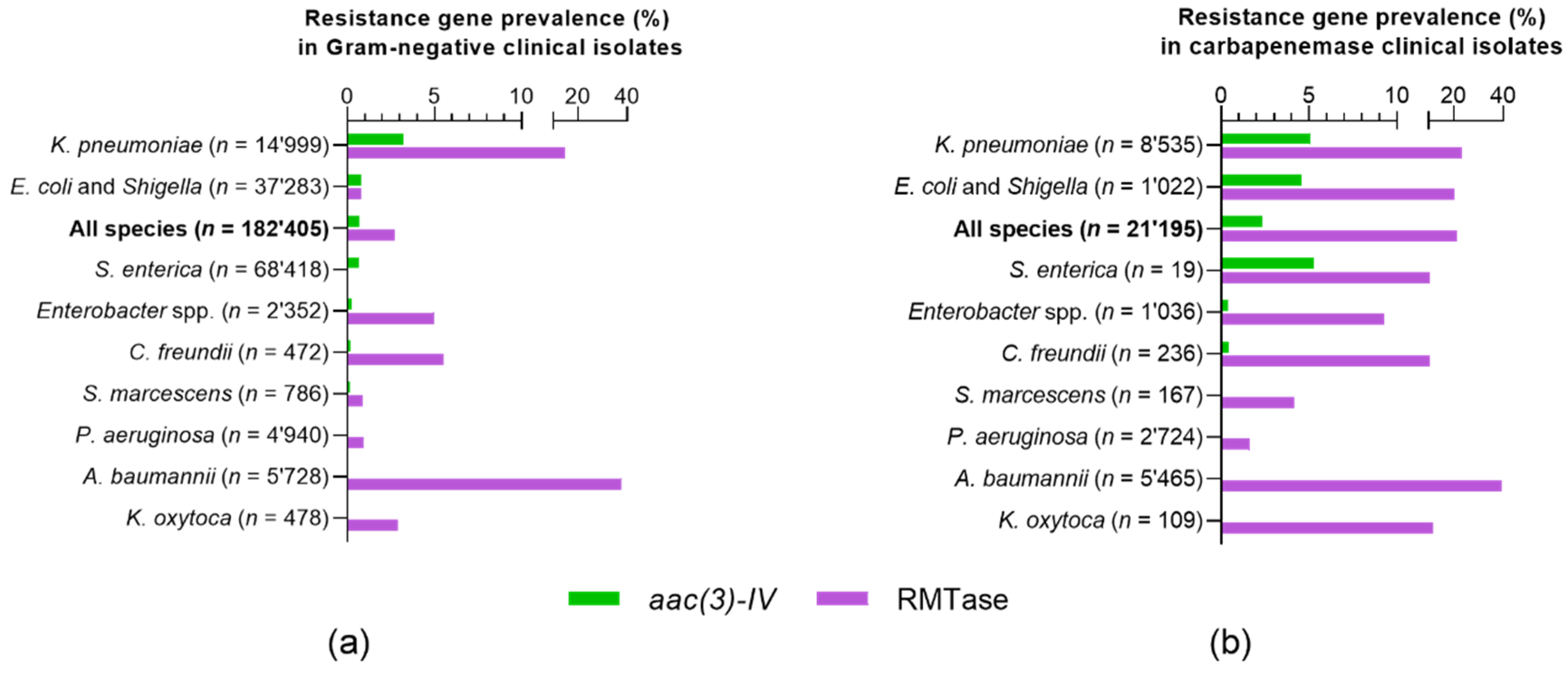
2.1. Genotypic Epidemiology of Apramycin Resistance
2.2. Phenotypic Characterization of aac(3)-IV
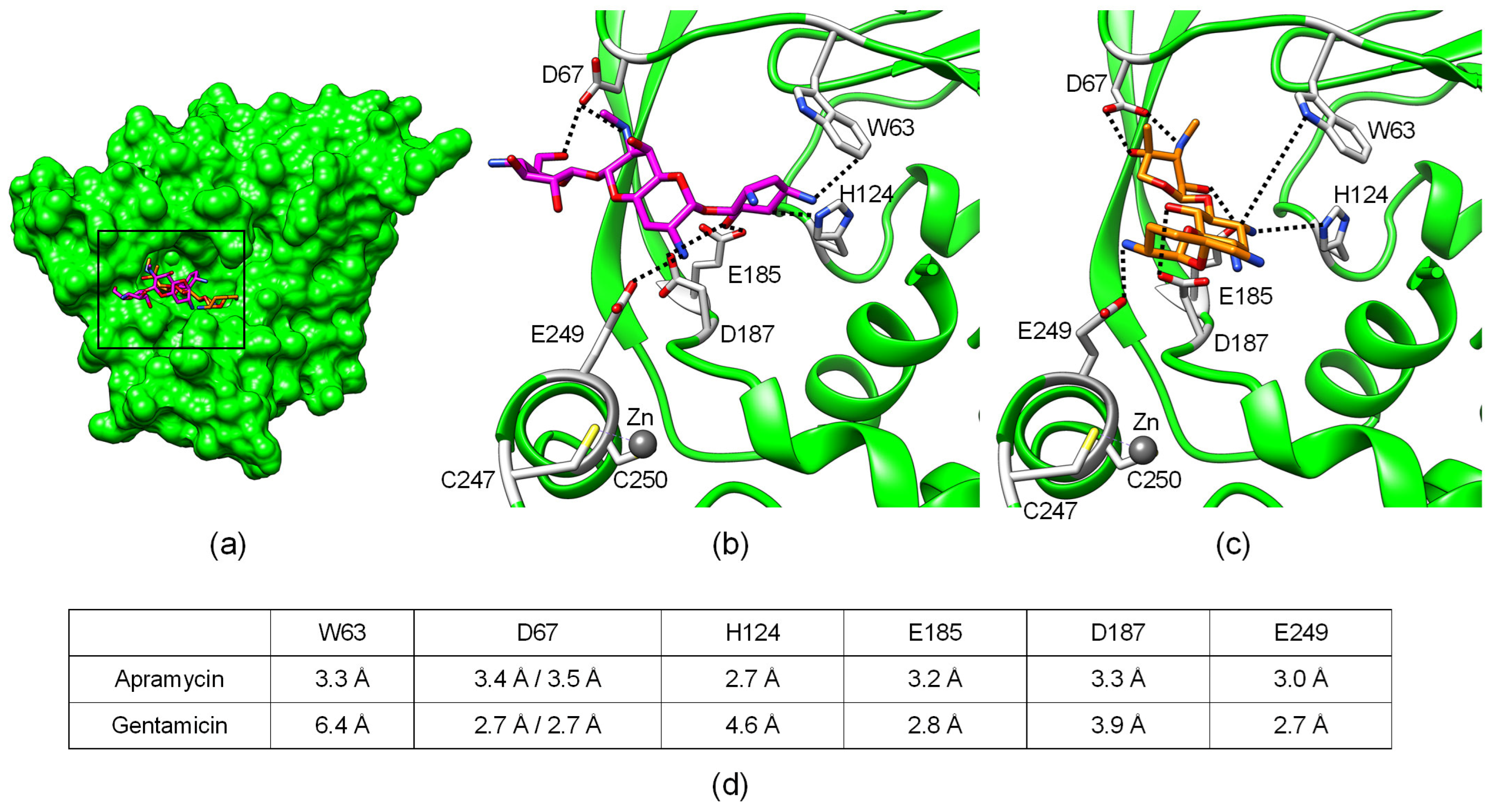
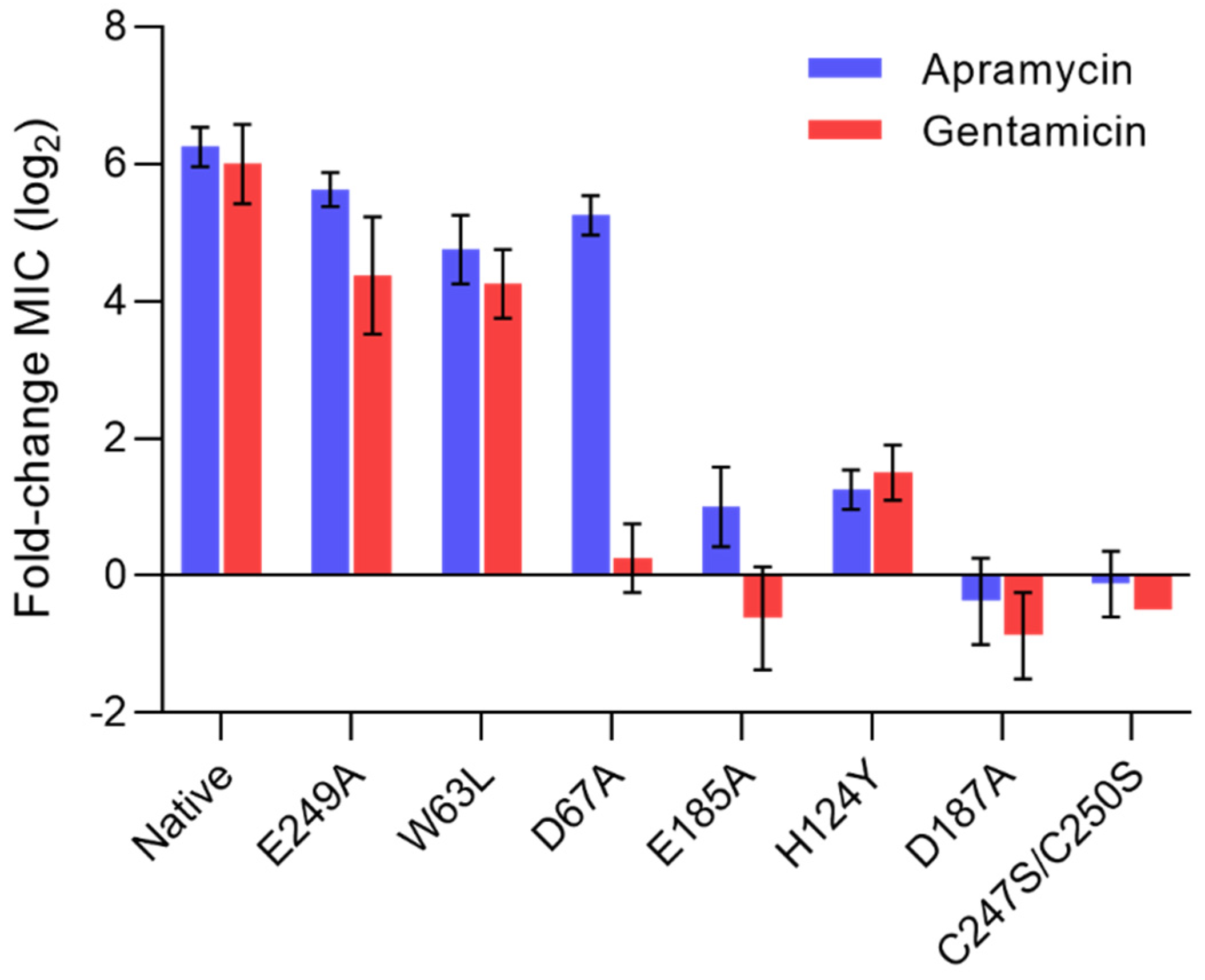
2.3. Structural Analysis of Aminoglycoside Binding to AAC(3)-IV
3. Discussion
4. Materials and Methods
4.1. Genomic Database Mining
4.2. Bacterial Strains
4.3. Antimicrobial-Susceptibility Testing
4.4. Structural Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Chopra, I.; Hesse, L.; O’Neill, A.J. Exploiting current understanding of antibiotic action for discovery of new drugs. J. Appl. Microbiol. 2002, 92, 4S–15S. [Google Scholar] [CrossRef]
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, a027029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updates 2010, 13, 151–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, G.; Ejim, L.; Stogios, P.J.; Koteva, K.; Bordeleau, E.; Evdokimova, E.; Sieron, A.O.; Savchenko, A.; Serio, A.W.; Krause, K.M.; et al. Plazomicin Retains Antibiotic Activity against Most Aminoglycoside Modifying Enzymes. ACS Infect. Dis. 2018, 4, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Doi, Y.; Wachino, J.I.; Arakawa, Y. Aminoglycoside Resistance: The Emergence of Acquired 16S Ribosomal RNA Methyltransferases. Infect. Dis. Clin. North Am. 2016, 30, 523–537. [Google Scholar] [CrossRef]
- Takahashi, Y.; Igarashi, M. Destination of aminoglycoside antibiotics in the “post-antibiotic era”. J. Antibiot. 2018, 71, 4–14. [Google Scholar] [CrossRef]
- World Health Organization Model List of Essential Medicines, 21st List, 2019. Geneva: World Health Organization. 2019. Licence: CCBY-NC-SA3.0IGO. Available online: https://apps.who.int/iris/bitstream/handle/10665/325771/WHO-MVP-EMP-IAU-2019.06-eng.pdf?ua=1 (accessed on 25 August 2020).
- Matt, T.; Ng, C.L.; Lang, K.; Sha, S.-H.; Akbergenov, R.; Shcherbakov, D.; Meyer, M.; Duscha, S.; Xie, J.; Dubbaka, S.R.; et al. Dissociation of antibacterial activity and aminoglycoside ototoxicity in the 4-monosubstituted 2-deoxystreptamine apramycin. Proc. Natl. Acad. Sci. USA 2012, 109, 10984–10989. [Google Scholar] [CrossRef] [Green Version]
- Butler, M.S.; Paterson, D.L. Antibiotics in the clinical pipeline in October 2019. J. Antibiot. 2020, 73, 329–364. [Google Scholar] [CrossRef]
- Juhas, M.; Widlake, E.; Teo, J.; Huseby, D.L.; Tyrrell, J.M.; Polikanov, Y.S.; Ercan, O.; Petersson, A.; Cao, S.; Aboklaish, A.F.; et al. In vitro activity of apramycin against multidrug-, carbapenem- and aminoglycoside-resistant Enterobacteriaceae and Acinetobacter baumannii. J. Antimicrob. Chemother. 2019, 74, 944–952. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.; O’Connor, S. Enzymatic modification of aminoglycoside antibiotics: 3-N-acetyltransferase with broad specificity that determines resistance to the novel aminoglycoside apramycin. Antimicrob. Agents Chemother. 1978, 14, 69–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fessler, A.T.; Kadlec, K.; Schwarz, S. Novel apramycin resistance gene apmA in bovine and porcine methicillin-resistant Staphylococcus aureus ST398 isolates. Antimicrob. Agents Chemother. 2011, 55, 373–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovering, A.M.; White, L.O.; Reeves, D.S. AAC(1): A new aminoglycoside-acetylating enzyme modifying the Cl aminogroup of apramycin. J. Antimicrob. Chemother. 1987, 20, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Wachino, J.; Shibayama, K.; Kurokawa, H.; Kimura, K.; Yamane, K.; Suzuki, S.; Shibata, N.; Ike, Y.; Arakawa, Y. Novel plasmid-mediated 16S rRNA m1A1408 methyltransferase, NpmA, found in a clinically isolated Escherichia coli strain resistant to structurally diverse aminoglycosides. Antimicrob. Agents Chemother. 2007, 51, 4401–4409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skeggs, P.A.; Holmes, D.J.; Cundliffe, E. Cloning of Aminoglycoside-Resistance Determinants from Streptomyces Tenebrarius and Comparison with Related Genes from Other Actinomycetes. Microbiology 1987, 133, 915–923. [Google Scholar] [CrossRef] [Green Version]
- Consaul, S.A.; Pavelka, M.S. Use of a novel allele of the Escherichia coli aacC4 aminoglycoside resistance gene as a genetic marker in mycobacteria. FEMS Microbiol. Lett. 2004, 234, 297–301. [Google Scholar] [CrossRef]
- Livermore, D.M.; Mushtaq, S.; Warner, M.; Zhang, J.-C.; Maharjan, S.; Doumith, M.; Woodford, N. Activity of aminoglycosides, including ACHN-490, against carbapenem-resistant Enterobacteriaceae isolates. J. Antimicrob. Chemother. 2011, 66, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Liu, L.; Zhang, X.; Feng, Y.; Zong, Z. In Vitro Activity of Neomycin, Streptomycin, Paromomycin and Apramycin against Carbapenem-Resistant Enterobacteriaceae Clinical Strains. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Kang, A.D.; Smith, K.P.; Eliopoulos, G.M.; Berg, A.H.; McCoy, C.; Kirby, J.E. In vitro Apramycin Activity against multidrug-resistant Acinetobacter baumannii and Pseudomonas aeruginosa. Diagn. Microbiol. Infect. Dis. 2017, 88, 188–191. [Google Scholar] [CrossRef]
- Truelson, K.A.; Brennan-Krohn, T.; Smith, K.P.; Kirby, J.E. Evaluation of apramycin activity against methicillin-resistant, methicillin-sensitive, and vancomycin-intermediate Staphylococcus aureus clinical isolates. Diagn. Microbiol. Infect. Dis. 2018, 92, 168–171. [Google Scholar] [CrossRef]
- Kang, A.D.; Smith, K.P.; Berg, A.H.; Truelson, K.A.; Eliopoulos, G.M.; McCoy, C.; Kirby, J.E. Efficacy of Apramycin against Multidrug-Resistant Acinetobacter baumannii in the Murine Neutropenic Thigh Model. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedel, S.; Vijayakumar, D.; Berg, G.; Kang, A.D.; Smith, K.P.; Kirby, J.E. Evaluation of apramycin against spectinomycin-resistant and -susceptible strains of Neisseria gonorrhoeae. J. Antimicrob. Chemother. 2019, 74, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Brennan-Krohn, T.; Kirby, J.E. Synergistic Combinations and Repurposed Antibiotics Active against the Pandrug-Resistant Klebsiella pneumoniae Nevada Strain. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galani, I.; Nafplioti, K.; Adamou, P.; Karaiskos, I.; Giamarellou, H.; Souli, M. Nationwide epidemiology of carbapenem resistant Klebsiella pneumoniae isolates from Greek hospitals, with regards to plazomicin and aminoglycoside resistance. BMC Infect. Dis. 2019, 19, 167. [Google Scholar] [CrossRef]
- Nafplioti, K.; Galani, I.; Angelidis, E.; Adamou, P.; Moraitou, E.; Giannopoulou, P.; Chra, P.; Damala, M.; Vogiatzakis, E.; Trikka-Graphakos, E.; et al. Dissemination of International Clone II Acinetobacter baumannii Strains Coproducing OXA-23 Carbapenemase and 16S rRNA Methylase ArmA in Athens, Greece. Microb. Drug Resist. 2020, 26, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Shi, X.; Lv, J.; Niu, S.; Cheng, S.; Du, H.; Yu, F.; Tang, Y.-W.; Kreiswirth, B.N.; Zhang, H.; et al. In vitro Activity of Apramycin Against Carbapenem-Resistant and Hypervirulent Klebsiella pneumoniae Isolates. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.P.; Kirby, J.E. Evaluation of apramycin activity against carbapenem-resistant and -susceptible strains of Enterobacteriaceae. Diagn. Microbiol. Infect. Dis. 2016, 86, 439–441. [Google Scholar] [CrossRef]
- Costello, S.E.; Deshpande, L.M.; Davis, A.P.; Mendes, R.E.; Castanheira, M. Aminoglycoside-modifying enzyme and 16S ribosomal RNA methyltransferase genes among a global collection of Gram-negative isolates. J. Glob. Antimicrob. Resist. 2019, 16, 278–285. [Google Scholar] [CrossRef]
- Rahman, M.; Prasad, K.N.; Gupta, S.; Singh, S.; Singh, A.; Pathak, A.; Gupta, K.K.; Ahmad, S.; Gonzalez-Zorn, B. Prevalence and Molecular Characterization of New Delhi Metallo-Beta-Lactamases in Multidrug-Resistant Pseudomonas aeruginosa and Acinetobacter baumannii from India. Microb. Drug Resist. 2018, 24, 792–798. [Google Scholar] [CrossRef]
- Taylor, E.; Sriskandan, S.; Woodford, N.; Hopkins, K.L. High prevalence of 16S rRNA methyltransferases among carbapenemase-producing Enterobacteriaceae in the UK and Ireland. Int. J. Antimicrob. Agents 2018, 52, 278–282. [Google Scholar] [CrossRef]
- Bodendoerfer, E.; Marchesi, M.; Imkamp, F.; Courvalin, P.; Böttger, E.C.; Mancini, S. Co-occurrence of aminoglycoside and β-lactam resistance mechanisms in aminoglycoside- non-susceptible Escherichia coli isolated in the Zurich area, Switzerland. Int. J. Antimicrob. Agents 2020, 56, 106019. [Google Scholar] [CrossRef] [PubMed]
- Eiland, L.S.; Jenkins, L.S. Optimal treatment of campylobacter dysentery. J. Pediatr. Pharmacol. Ther. 2008, 13, 170–174. [Google Scholar] [PubMed]
- Bagdasarian, N.; Rao, K.; Malani, P.N. Diagnosis and Treatment of Clostridium difficile in Adults. JAMA 2015, 313, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Bräu, B.; Pilz, U.; Piepersberg, W. Genes for gentamicin-(3)-N-acetyltransferases III and IV: I. Nucleotide sequence of the AAC(3)-IV gene and possible involvement of an IS140 element in its expression. Mol. Gen. Genet. 1984, 193, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, M.L.B.; Blanchard, J.S. The Kinetic Mechanism of AAC(3)-IV Aminoglycoside Acetyltransferase from Escherichia coli †. Biochemistry 2005, 44, 16275–16283. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.H.; Rubin, A.J.; Sauer, R.T. Design, construction and characterization of a set of insulated bacterial promoters. Nucleic Acids Res. 2011, 39, 1131–1141. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Selvaraj, B.; Serpersu, E.H.; Cuneo, M.J. Encoding of Promiscuity in an Aminoglycoside Acetyltransferase. J. Med. Chem. 2018, 61, 10218–10227. [Google Scholar] [CrossRef]
- Kumar, P.; Serpersu, E.H.; Cuneo, M.J. A low-barrier hydrogen bond mediates antibiotic resistance in a noncanonical catalytic triad. Sci. Adv. 2018, 4, eaas8667. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| E. coli Clinical aac(3)-IV Isolates | E. coli Recombinant aac(3)-IV Strains | |||||||
|---|---|---|---|---|---|---|---|---|
| ATCC | AG173 | AG380 | AG381 | Promoter Strength | ||||
| Aminoglycoside | 25,922 | WT | + | ++ | +++ | |||
| Apramycin | 4 | 256–512 | >512 | >512 | 2 | 128 | >512 | >512 |
| Gentamicin | 0.5–1 | 8–16 | 32–64 | 64–128 | 0.25 | 8 | 64 | 128–256 |
| Tobramycin | 0.5–1 | 32 | 64–128 | 128 | 0.5 | 16 | 128–256 | 256–512 |
| Amikacin | 2 | 1 | 2–4 | 4 | 0.5–1 | 0.25–0.5 | 0.25 | 0.25–0.5 |
| Sisomicin | 0.5 | 8–16 | 16 | 32 | 0.125–0.25 | 2 | 16–32 | 128 |
| Netilmicin | 0.5 | 8–16 | 32–64 | 32 | 0.125–0.25 | 4–8 | 32–64 | 128 |
| Plazomicin | 0.5–1 | 0.5–1 | 1 | 1–2 | 0.25–0.5 | 0.125 | 0.125–0.25 | 0.125 |
| Paromomycin | 4 | 8 | 8 | 16 | 1 | 2 | 16 | 64–128 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plattner, M.; Gysin, M.; Haldimann, K.; Becker, K.; Hobbie, S.N. Epidemiologic, Phenotypic, and Structural Characterization of Aminoglycoside-Resistance Gene aac(3)-IV. Int. J. Mol. Sci. 2020, 21, 6133. https://doi.org/10.3390/ijms21176133
Plattner M, Gysin M, Haldimann K, Becker K, Hobbie SN. Epidemiologic, Phenotypic, and Structural Characterization of Aminoglycoside-Resistance Gene aac(3)-IV. International Journal of Molecular Sciences. 2020; 21(17):6133. https://doi.org/10.3390/ijms21176133
Chicago/Turabian StylePlattner, Michel, Marina Gysin, Klara Haldimann, Katja Becker, and Sven N. Hobbie. 2020. "Epidemiologic, Phenotypic, and Structural Characterization of Aminoglycoside-Resistance Gene aac(3)-IV" International Journal of Molecular Sciences 21, no. 17: 6133. https://doi.org/10.3390/ijms21176133