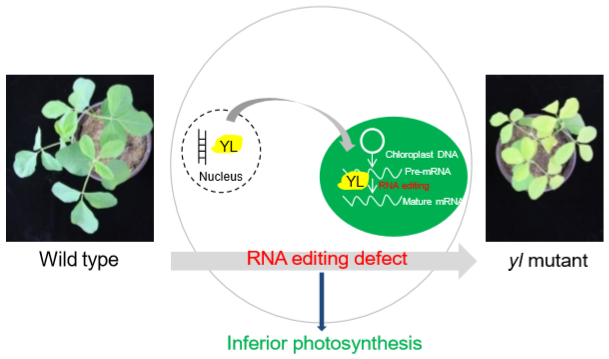
Mutation of YL Results in a Yellow Leaf with Chloroplast RNA Editing Defect in Soybean

Abstract
:
1. Introduction
2. Results
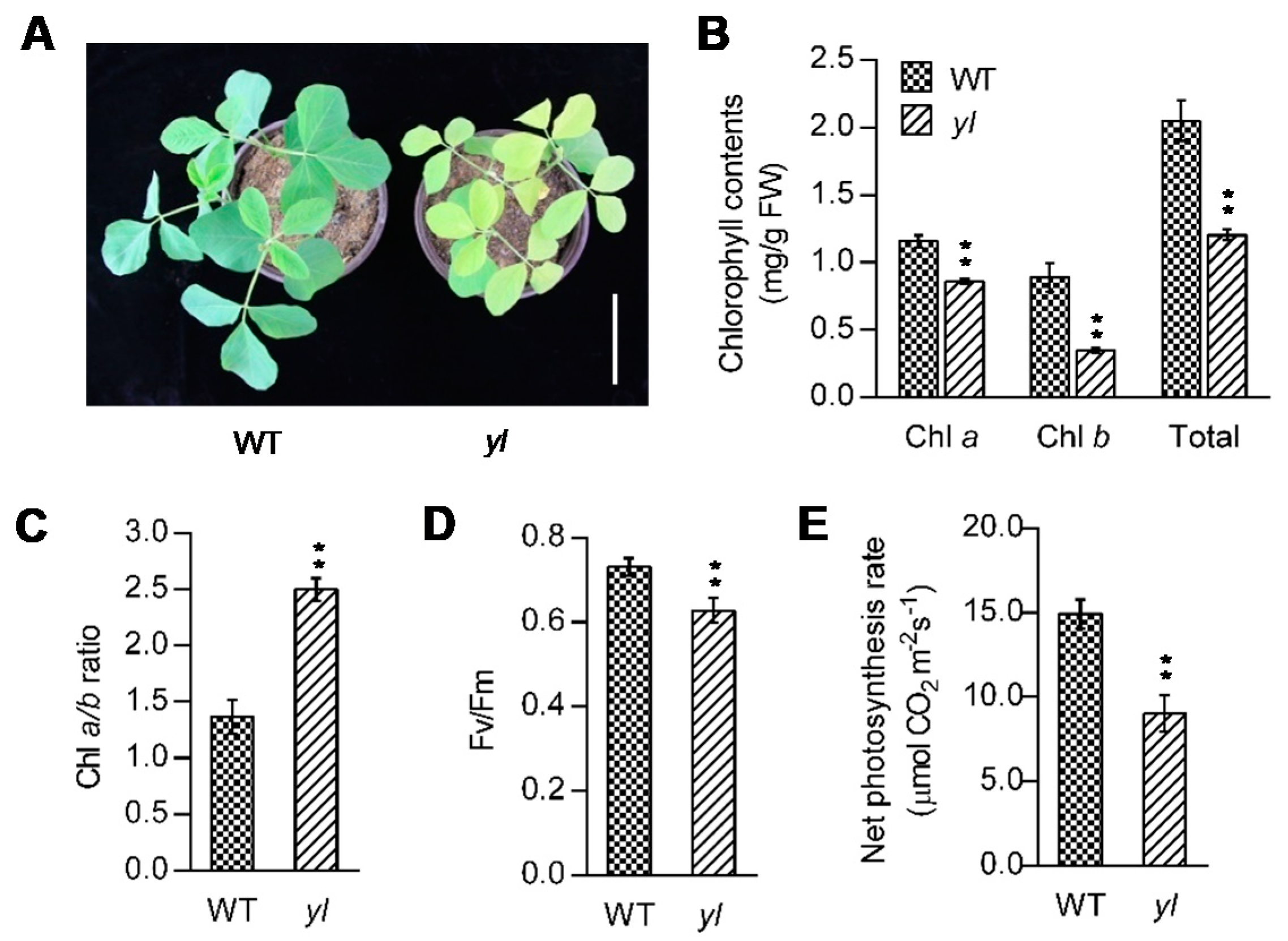
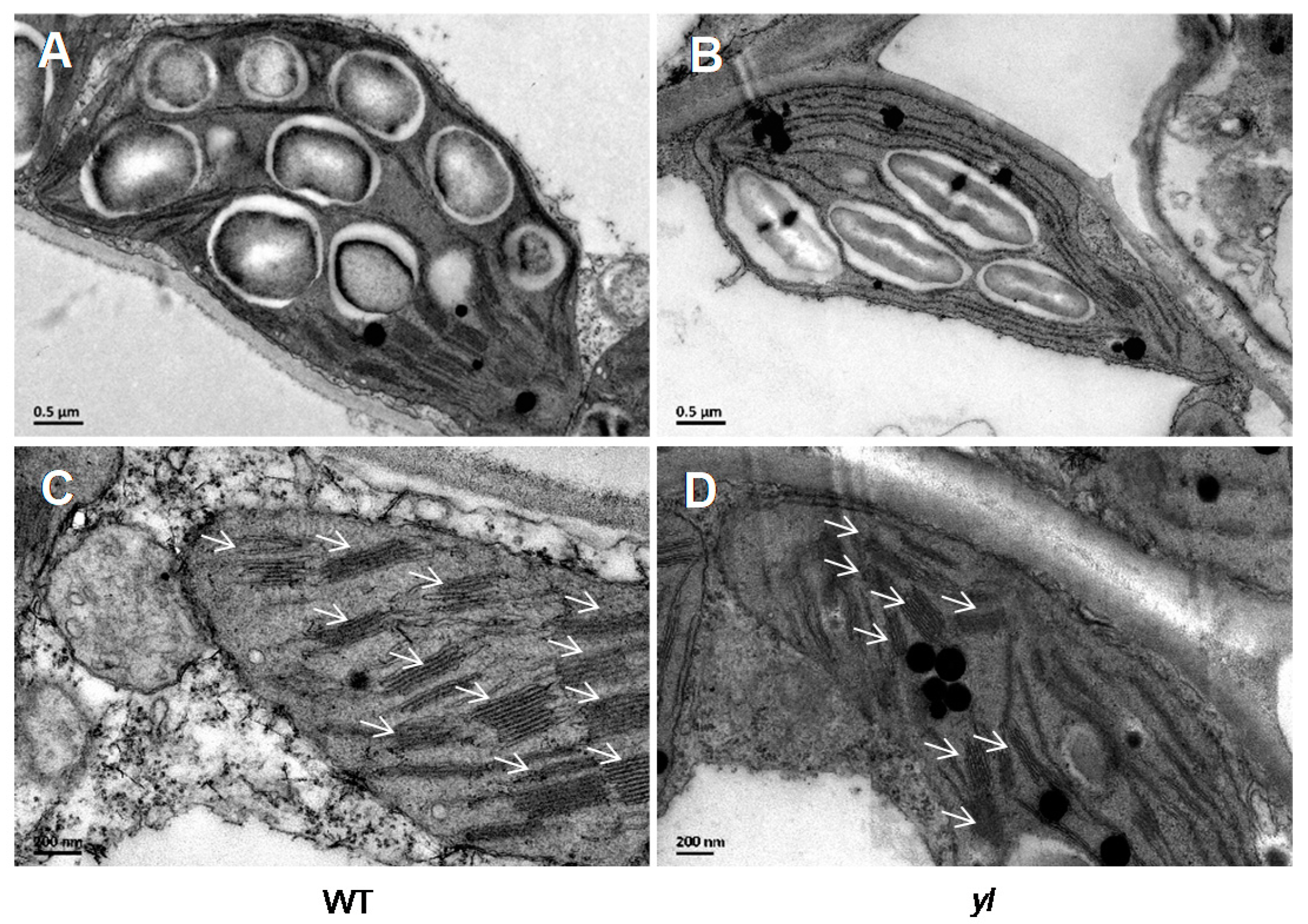
2.1. yl Plants Show Yellow Leaves and Abnormal Photosynthesis at the Seedling Stage
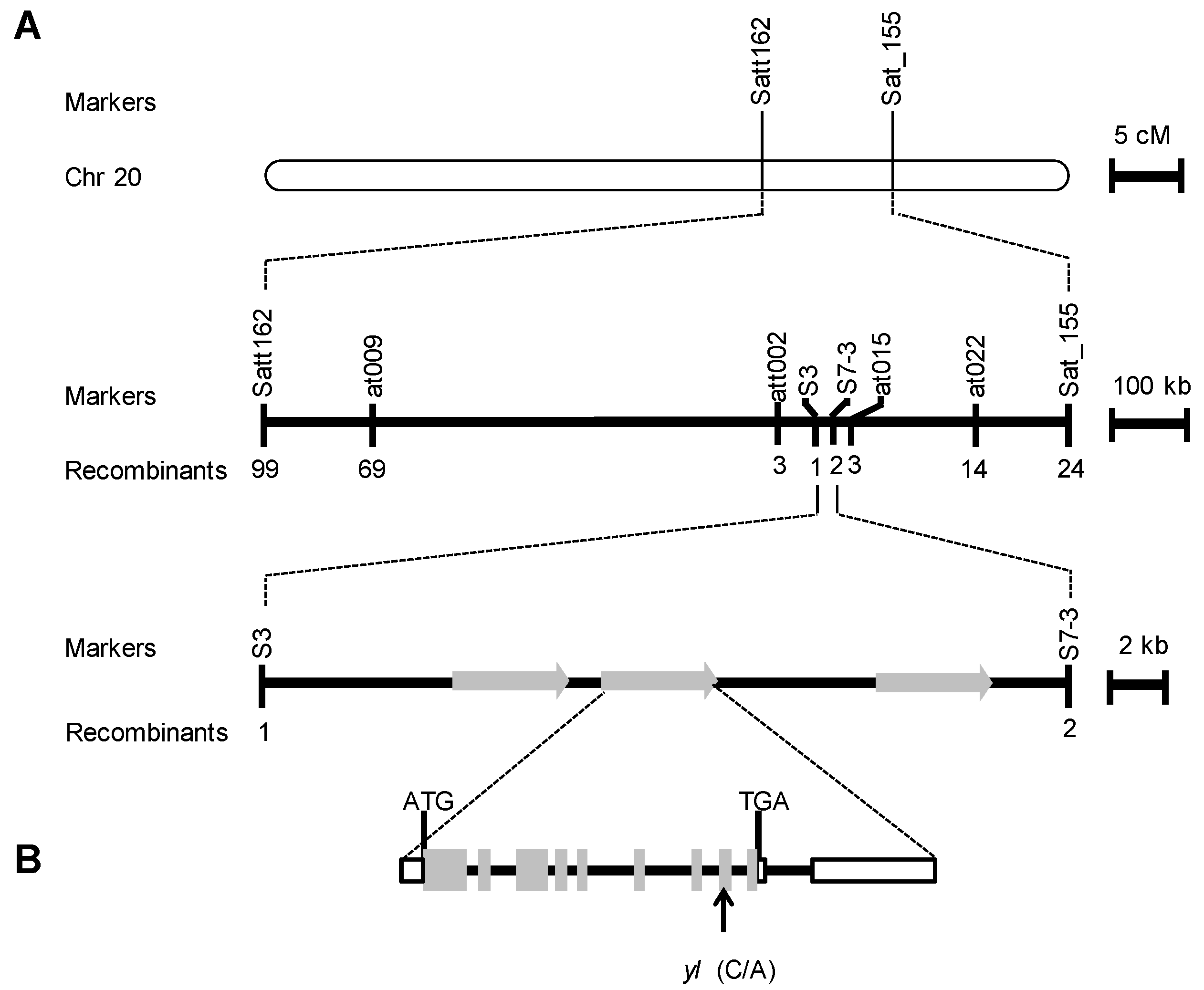
2.2. Fine Mapping of the YL Gene
2.3. YL Encodes an Organelle RNA Recognition Motif-Containing Protein 1 (GmORRM1)
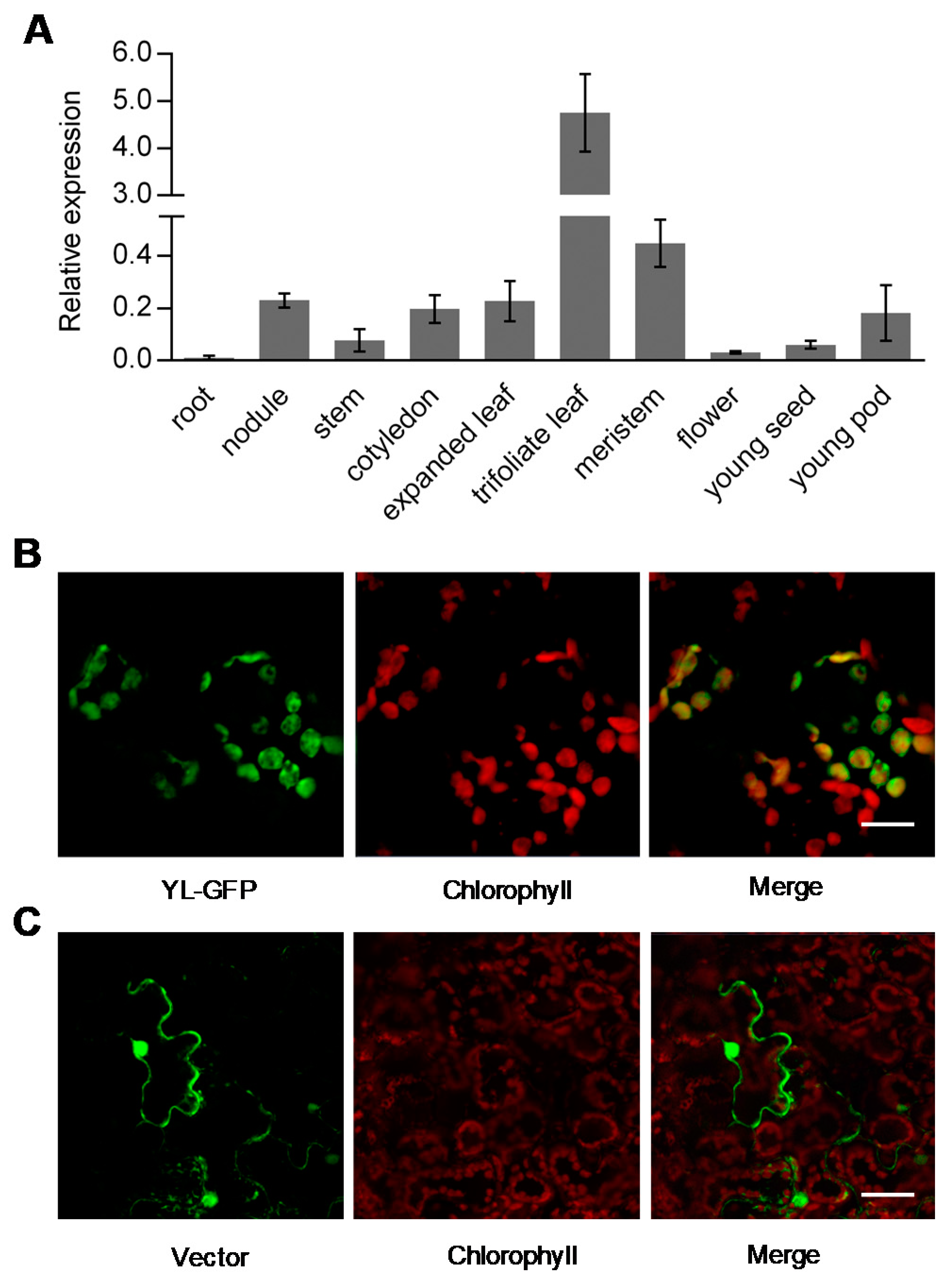
2.4. Expression Pattern and Protein Subcellular Localization of YL
2.5. Dramatic Defects of Chloroplast RNA Editing in yl
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Chl Contents, Net Photosynthetic Rate and Photochemical Efficiency Analysis
4.3. Transmission Electron Microscopy
4.4. Fine Mapping of the YL Gene
4.5. RNA Extraction and Quantitative Real-Time RT-PCR
4.6. Subcellular Localization
4.7. Chloroplast RNA Editing Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stirbet, A.; Lazar, D.; Guo, Y.; Govindjee, G. Photosynthesis: Basics, history, and modeling. Ann. Bot. 2019, XX, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.P. Photosynthesis. Essays Biochem. 2016, 60, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Von Wettstein, D.; Gough, S.; Kannangara, C.G. Chlorophyll biosynthesis. Plant Cell 1995, 7, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, D.; Coleman, Z.; Atkinson, T.; Rai, K.M.; Mendu, V. Genetics and physiology of the nuclearly inherited yellow foliar mutants in soybean. Front. Plant Sci. 2018, 9, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, K.K.; Palmer, R.G. Duplicate chlorophyll-deficient loci in soybean. Genome 2014, 47, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.W.; Mani, D.; Curtin, S.J.; Slattery, R.A.; Michno, J.M.; Ort, D.R.; Schaus, P.J.; Palmer, R.G.; Orf, J.H.; Stupar, R.M. Identical substitutions in magnesium chelatase paralogs result in chlorophyll-deficient soybean mutants. G3 Genes Genom. Genet. 2014, 5, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, S.; Atkinson, T.; Gorecki, C.; Espinosa, K.; Przybylski, S.; Goggi, A.S.; Palmer, R.G.; Sandhu, D. Candidate gene identification for a lethal chlorophyll-deficient mutant in soybean. Agronomy 2014, 4, 462–469. [Google Scholar] [CrossRef]
- Sandhu, D.; Atkinson, T.; Noll, A.; Johnson, C.; Espinosa, K.; Boelter, J.; Abel, S.; Dhatt, B.K.; Barta, T.; Singsaas, E.; et al. Soybean proteins GmTic110 and GmPsbP are crucial for chloroplast development and function. Plant Sci. 2016, 252, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Fang, C.; Duan, Z.B.; Liu, Y.C.; Qin, H.; Zhang, J.X.; Sun, P.; Li, W.B.; Wang, G.D.; Tian, Z.X. Functional conservation and divergence of GmCHLI genes in polyploid soybean. Plant J. 2016, 88, 584–596. [Google Scholar] [CrossRef]
- Weiss, M.G. Genetic linkage in soybeans: Linkage group I. Crop Sci. 1970, 10, 69–72. [Google Scholar] [CrossRef]
- Weiss, M.G. Genetic linkage in soybeans: Linkage group VII. Crop Sci. 1970, 10, 627–629. [Google Scholar] [CrossRef]
- Palmer, R.G.; Nelson, R.L.; Bernard, R.L.; Stelly, D.M. Genetics and linkage of three chlorophyll-deficient mutants in soybean: y19, y22, and y23. J. Hered. 1990, 81, 404–406. [Google Scholar]
- Zou, J.J.; Singh, R.J.; Hymowitz, T. Association of the yellow leaf (y10) mutant to soybean chromosome 3. J. Hered. 2003, 94, 352–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, R.G.; Xu, M. Positioning 3 qualitative trait loci on soybean molecular linkage group E. J. Hered. 2008, 99, 674–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, D.; Han, S.; Zhang, X.; Yu, D. Identification and gene mapping of a soybean chlorophyll-deficient mutant. Plant Breed. 2011, 130, 133–138. [Google Scholar] [CrossRef]
- Sandhu, D.; Ghosh, J.; Johnson, C.; Baumbach, J.; Baumert, E.; Cina, T.; Grant, D.; Palmer, R.G.; Bhattacharyya, M.K. The endogenous transposable element Tgm9 is suitable for generating knockout mutants for functional analyses of soybean genes and genetic improvement in soybean. PLoS ONE 2017, 12, e0180732. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, M.; Zehrmann, A.; Verbitskiy, D.; Hartel, B.; Brennicke, A. RNA editing in plants and its evolution. Annu. Rev. Genet. 2013, 47, 335–352. [Google Scholar] [CrossRef]
- Ichinose, M.; Sugita, M. RNA editing and its molecular mechanism in plant organelles. Genes 2016, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Bentolila, S.; Hanson, M.R. The unexpected diversity of plant organelle RNA editosomes. Trends Plant Sci. 2016, 21, 962–973. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Germain, A.; Giloteaux, L.; Hammani, K.; Barkan, A.; Hanson, M.R.; Bentolila, S. An RNA recognition motif-containing protein is required for plastid RNA editing in Arabidopsis and maize. Proc. Natl. Acad. Sci. USA 2013, 110, E1169–E1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, J.B.; Shi, X.; Kobylarz, A.T.; Lucas, M.K.; Wessendorf, R.L.; Hines, K.M.; Bentolila, S.; Hanson, M.R.; Lu, Y. An organelle RNA recognition motif protein is required for photosystem II subunit psbF transcript editing. Plant Physiol. 2017, 173, 2278–2293. [Google Scholar] [PubMed] [Green Version]
- Shi, X.; Hanson, M.R.; Bentolila, S. Two RNA recognition motif-containing proteins are plant mitochondrial editing factors. Nucleic Acids Res. 2015, 43, 3814–3825. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Germain, A.; Hanson, M.R.; Bentolila, S. RNA recognition motif-containing protein ORRM4 broadly affects mitochondrial RNA editing and impacts plant development and flowering. Plant Physiol. 2016, 170, 294–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Castandet, B.; Germain, A.; Hanson, M.R.; Bentolila, S. ORRM5, an RNA recognition motif-containing protein, has a unique effect on mitochondrial RNA editing. J. Exp. Bot. 2017, 68, 2833–2847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentolila, S.; Heller, W.P.; Sun, T.; Babina, A.M.; Friso, G.; van Wijk, K.J.; Hanson, M.R. RIP1, a member of an Arabidopsis protein family, interacts with the protein RARE1 and broadly affects RNA editing. Proc. Natl. Acad. Sci. USA 2012, 109, E1453–E1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenaka, M.; Zehrmann, A.; Verbitskiy, D.; Kugelmann, M.; Hartel, B.; Brennicke, A. Multiple organellar RNA editing factor (MORF) family proteins are required for RNA editing in mitochondria and plastids of plants. Proc. Natl. Acad. Sci. USA 2012, 109, 5104–5109. [Google Scholar] [CrossRef] [Green Version]
- Barkan, A.; Small, I. Pentatricopeptide repeat proteins in plants. Annu. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef]
- Zhang, F.; Tang, W.J.; Hedtke, B.; Zhong, L.L.; Liu, L.; Peng, L.W.; Lu, C.M.; Grimm, B.; Lin, R.C. Tetrapyrrole biosynthetic enzyme protoporphyrinogen IX oxidase 1 is required for plastid RNA editing. Proc. Natl. Acad. Sci. USA 2014, 111, 2023–2028. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Shi, X.; Friso, G.; Van Wijk, K.; Bentolila, S.; Hanson, M.R. A zinc finger motif-containing protein is essential for chloroplast RNA editing. PLoS Genet. 2015, 11, e1005028. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.B.; Huang, J.Y.; Chory, J. GUN1 interacts with MORF2 to regulate plastid RNA editing during retrograde signaling. Proc. Natl. Acad. Sci. USA 2019, 116, 10162–10167. [Google Scholar] [CrossRef] [Green Version]
- Phytozome. Available online: www.phytozome.net (accessed on 14 June 2020).
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, I.; Peeters, N.; Legeai, F.; Lurin, C. Predotar: A tool for rapidly screening proteomes for N-terminal targeting sequences. Proteomics 2004, 4, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Shikanai, T.; Shimizu, K.; Ueda, K.; Nishimura, Y.; Kuroiwa, T.; Hashimoto, T. The chloroplast clpP gene, encoding a proteolytic subunit of ATP-dependent protease, is indispensable for chloroplast development in tobacco. Plant Cell Physiol. 2001, 42, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Burrows, P.A.; Sazanov, L.A.; Svab, Z.; Maliga, P.; Nixon, P.J. Identification of a functional respiratory complex in chloroplasts through analysis of tobacco mutants containing disrupted plastid ndh genes. EMBO J. 1998, 17, 868–876. [Google Scholar] [CrossRef]
- Voelker, R.; Barkan, A. Nuclear genes required for post-translational steps in the biogenesis of the chloroplast cytochrome b6f complex in maize. Mol. Gen. Genet. 1995, 249, 507–514. [Google Scholar] [CrossRef]
- Zito, F.; Kuras, R.; Choquet, Y.; Kossel, H.; Wollman, F.A. Mutations of cytochrome b6 in Chlamydomonas reinhardtii disclose the functional significance for a proline to leucine conversion by petB editing in maize and tobacco. Plant Mol. Biol. 1997, 33, 79–86. [Google Scholar] [CrossRef]
- Monde, R.A.; Zito, F.; Olive, J.; Wollman, F.A.; Stern, D.B. Post-transcriptional defects in tobacco chloroplast mutants lacking the cytochrome b6/f complex. Plant J. 2000, 21, 61–72. [Google Scholar] [CrossRef]
- Szczepaniak, A.; Black, M.T.; Cramer, W.A. Topography of the chloroplast cytochrome b6: Orientation of the cytochrome and accessibility of the lumen-side interhelix loops. Z. Naturforsch. C J. Biosci. 1989, 44, 453–461. [Google Scholar] [CrossRef]
- Hankamer, B.; Morris, E.; Nield, J.; Carne, A.; Barber, J. Subunit positioning and transmembrane helix organisation in the core dimer of photosystem II. FEBS Lett. 2001, 504, 142–151. [Google Scholar] [CrossRef]
- Swiatek, M.; Regel, R.E.; Meurer, J.; Wanner, G.; Pakrasi, H.B.; Ohad, I.; Herrmann, R.G. Effects of selective inactivation of individual genes for low-molecular-mass subunits on the assembly of photosystem II, as revealed by chloroplast transformation: The psbEFLJ operon in Nicotiana tabacum. Mol. Genet. Genom. 2003, 268, 699–710. [Google Scholar] [CrossRef]
- Suorsa, M.; Regel, R.E.; Paakkarinen, V.; Battchikova, N.; Herrmann, R.G.; Aro, E.M. Protein assembly of photosystem II and accumulation of subcomplexes in the absence of low molecular mass subunits PsbL and PsbJ. Eur. J. Biochem. 2004, 271, 96–107. [Google Scholar] [CrossRef] [Green Version]
- Krieger-Liszkay, A. Singlet oxygen production in photosynthesis. J. Exp. Bot. 2005, 56, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eva-Mari, A.; Ivar, V.; Bertil, A. Photoinhibition of Photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta 1993, 1143, 113–134. [Google Scholar]
- Liu, M.; Wang, Y.; Nie, Z.; Gai, J.; Bhat Akhter, J.; Kong, J.; Zhao, T. Double mutation of two homologous genes YL1 and YL2 results in a leaf yellowing phenotype in soybean [Glycine max (L.) Merr]. Plant Mol. Biol. 2020. online. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method Enzymol. 1987, 148, 350–382. [Google Scholar]
- SoyBase. Available online: http://soybase.org/ (accessed on 14 March 2014).
- FastQC. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 September 2015).
- Glycine Max Cultivar PI 437654 Chloroplast, Complete Genome. Available online: ncbi.nlm.nih.gov/nuccore/DQ312375.1 (accessed on 12 January 2016).
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crosses | F1 | Number of F2 Plants | χ2 (3:1) | P | ||
|---|---|---|---|---|---|---|
| Wild Type | Mutant | Total | ||||
| yl × Williams 82 | Normal | 679 | 227 | 906 | 0 | >0.9 |
| yl × Zhonghuang13 | Normal | 440 | 163 | 603 | 1.221 | 0.50~0.25 |
| Zhonghuang13 × yl | Normal | 448 | 132 | 580 | 1.437 | 0.25~0.10 |
| Gene | Position a | Codon Site b | Conversion |
|---|---|---|---|
| accD | 617 | 2 | S(uCg)→L(uUg) |
| atpF | 92 | 2 | P(cCa)→L(cUa) |
| clpP | 559 | 1 | H(Cau)→Y(Uau) |
| ndhA | 341 | 2 | S(uCa)→L(uUa) |
| 1073 | 2 | S(uCu)→F(uUu) | |
| ndhB | 9 | 3 | W(ugG)→stop codon (ugA) |
| 149 | 2 | S(uCa)→L(uUa) | |
| 542 | 2 | T(aCg)→M(aUg) | |
| 586 | 1 | H(Cau)→Y(Uau) | |
| 737 | 2 | P(cCa)→L(cUa) | |
| 746 | 2 | S(uCu)→F(uUu) | |
| 830 | 2 | S(uCa)→L(uUa) | |
| 836 | 2 | S(uCa)→L(uUa) | |
| 1112 | 2 | S(uCa)→L(uUa) | |
| 1255 | 1 | H(Cau)→Y(Uau) | |
| 1481 | 2 | P(cCa)→L(cUa) | |
| ndhC | 323 | 2 | S(uCa)→L(uUa) |
| ndhD | 2 | 2 | T(aCg)→M(aUg) |
| 383 | 2 | T(aCa)→I(aUa) | |
| 674 | 2 | S(uCa)→L(uUa) | |
| 878 | 2 | S(uCa)→L(uUa) | |
| 1298 | 2 | S(uCa)→L(uUa) | |
| ndhE | 233 | 2 | P(cCg)→L(cUg) |
| ndhF | 290 | 2 | S(uCa)→L(uUa) |
| petB | 611 | 2 | S(uCa)→L(uUa) |
| psaI | 79 | 1 | H(Cau)→Y(Uau) |
| psbE | 214 | 2 | S(uCc)→F(uUc) |
| psbF | 6 | 3 | T(acC)→T(acU) |
| 77 | 2 | S(uCu)→F(uUu) | |
| psbL | 2 | 2 | T(aCg)→M(aUg) |
| rpl23 | 89 | 2 | S(uCa)→L(uUa) |
| rpoA | 200 | 2 | S(uCa)→L(uUa) |
| rpoB | 338 | 2 | S(uCu)→F(uUu) |
| 551 | 2 | S(uCa)→L(uUa) | |
| 566 | 2 | S(uCg)→L(uUg) | |
| 2000 | 2 | S(uCu)→F(uUu) | |
| rpoC1 | 41 | 2 | S(uCa)→L(uUa) |
| 488 | 2 | S(uCa)→L(uUa) | |
| rps2 | 134 | 2 | T(aCa)→I(aUa) |
| 248 | 2 | S(uCa)→L(uUa) | |
| rps12 | 554 c | ||
| rps14 | 80 | 2 | S(uCa)→L(uUa) |
| rps16 | 499 c | ||
| 212 | 2 | S(uCa)→L(uUa) | |
| rps18 | 221 | 2 | S(uCg)→L(uUg) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Pan, Y.; Liu, Z.; Liu, Y.; Zhong, D.; Duan, Z.; Tian, Z.; Zhu, B.; Zhou, G. Mutation of YL Results in a Yellow Leaf with Chloroplast RNA Editing Defect in Soybean. Int. J. Mol. Sci. 2020, 21, 4275. https://doi.org/10.3390/ijms21124275
Zhu X, Pan Y, Liu Z, Liu Y, Zhong D, Duan Z, Tian Z, Zhu B, Zhou G. Mutation of YL Results in a Yellow Leaf with Chloroplast RNA Editing Defect in Soybean. International Journal of Molecular Sciences. 2020; 21(12):4275. https://doi.org/10.3390/ijms21124275
Chicago/Turabian StyleZhu, Xiaowei, Yi Pan, Zhi Liu, Yucheng Liu, Deyi Zhong, Zongbiao Duan, Zhixi Tian, Baoge Zhu, and Guoan Zhou. 2020. "Mutation of YL Results in a Yellow Leaf with Chloroplast RNA Editing Defect in Soybean" International Journal of Molecular Sciences 21, no. 12: 4275. https://doi.org/10.3390/ijms21124275