The Fundamental Role of Bicarbonate Transporters and Associated Carbonic Anhydrase Enzymes in Maintaining Ion and pH Homeostasis in Non-Secretory Organs


Abstract
:1. Convergent Regulation of Ion Homeostasis
2. Carbonic Anhydrases: Regulation of HCO3−
3. Regulatory Factors of HCO3− Transporters
3.1. Regulatory Molecules
3.2. Cl− as A Signaling Ion
4. HCO3− and pH Regulation and Coordinated Transporters
4.1. Immune System
4.2. Tumor System
4.3. Tooth Developmental System
4.4. Vascular Smooth Muscle System
4.5. Cardiac System
4.6. Digestive System
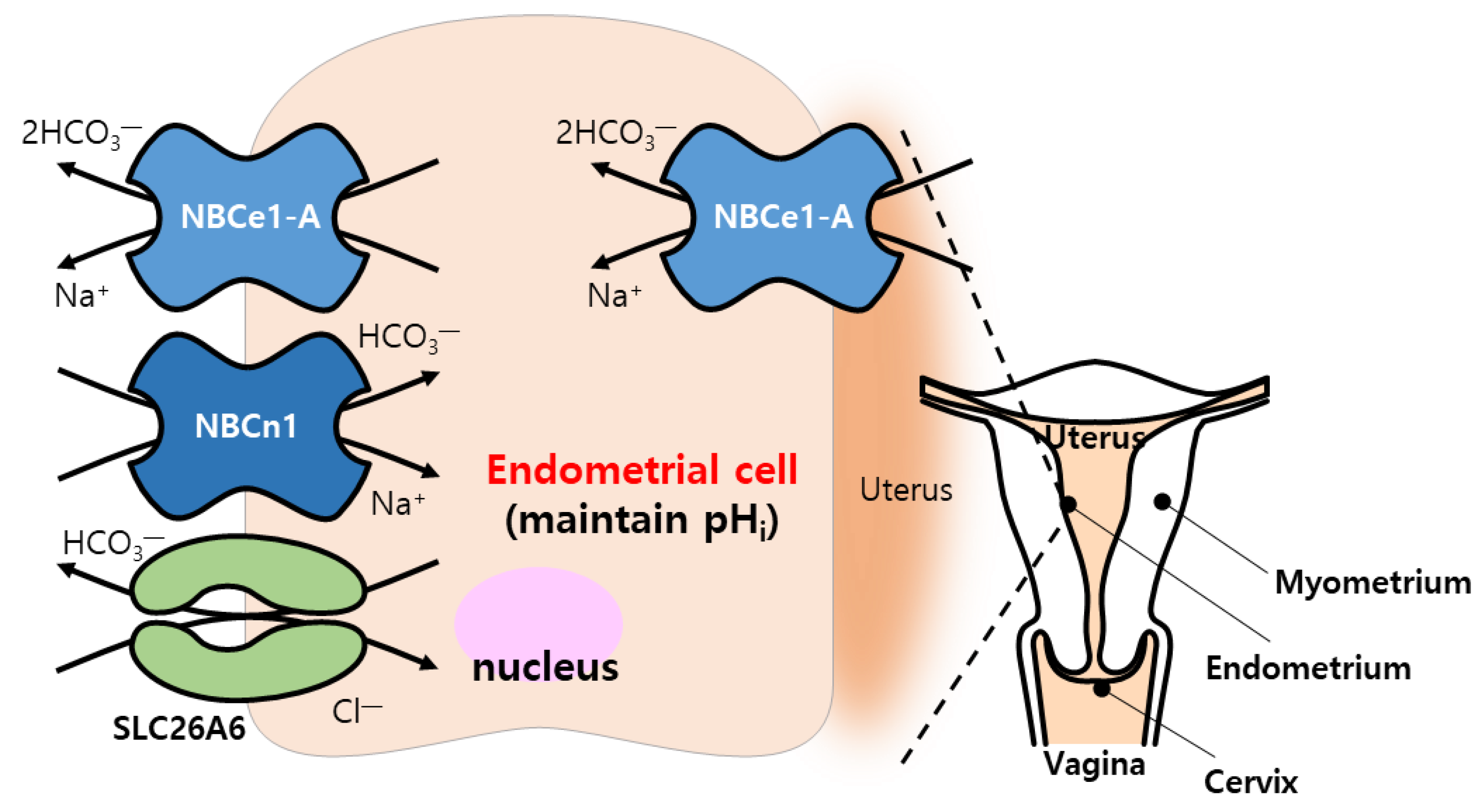
4.7. Reproduction System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological/Pathological System | Transporters | Localization and Function | References |
|---|---|---|---|
| Immune | NBCn1 | Macrophage differentiation | [76,77] |
| NBCe1, NBCn1 | Neutrophils, maintenance of intracellular pH | [78,79] | |
| AE2 | CD8+ T cells, controlling cell proliferation | [80] | |
| Tumorigenesis | NBCe1 | Colon/breast cancer, inducing cell proliferation | [31] |
| NBCn1 | Development and motility of breast cancer | [83,84,101] | |
| AE3, DRA | Pancreatic ductal adenocarcinoma | [94] | |
| AE2 | Colon cancer, promotion of cell proliferation | [96] | |
| Tooth development | NBCe1B–E, AE2 | Ameloblasts in maturation stage, enamel development | [104,105] |
| SLC26A1, SLC26A7 | Maturation-stage rodent ameloblasts, enamel formation | [106] | |
| NBC, AE | Human dental pulp stem cells, pH modulation | [108] | |
| Vascular smooth muscle | NBCn1 | Vascular smooth muscle cells, myogenic tone, regulation of blood pressure, migration, arterial remodeling | [109], [110], [111], [112,113,114] |
| Cardiac | NBCe1, NBCn1 | Cardiac hypertrophy | [120] |
| Digestive | NBCe1-B, NBCe1-C | Proximal colon | [129] |
| NBCn1-C, NBCn1-D | Proximal and distal colon | [129] | |
| AE2 | Parietal cells | [130] | |
| SLC26A9 | GI tract | [131] | |
| Reproduction | CFTR, AE2, NBCe1, NBCn1, NDCBE | Sertoli cells, spermatogenesis | [134,135] |
| CFTR, DRA, SLC26A6 | Male reproductive duct, maintenance of luminal pH | [133,142,143,144] | |
| NBCn1, SLC26A4, SLC26A6 | Endometrial cells | [145,147] | |
| NBCe1-A | Luminal and grandular epithelial cells of uteri | [146] |
5. Future Perspectives and Challenges
Author Contributions
Funding
Conflicts of Interest
References
- Noor, Z.N.; Deitmer, J.W.; Theparambil, S.M. Cytosolic sodium regulation in mouse cortical astrocytes and its dependence on potassium and bicarbonate. J. Cell. Physiol. 2018, 234, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowitz, D. CFTR, bicarbonate, and the pathophysiology of cystic fibrosis. Pediatr. Pulmonol. 2015, 50, S24–S30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Criq, V.; Gray, M.A. Role of CFTR in epithelial physiology. Cell. Mol. Life Sci. 2017, 74, 93–115. [Google Scholar] [CrossRef] [Green Version]
- Boron, W.F.; Fong, P.; Hediger, M.A.; Boulpaep, E.L.; Romero, M.F. The electrogenic Na/HCO3 cotransporter. Kidney Int. 1997, 109, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Parker, M.D.; Boron, W.F. The divergence, actions, roles, and relatives of sodium-coupled bicarbonate transporters. Physiol. Rev. 2013, 93, 803–959. [Google Scholar] [CrossRef] [Green Version]
- Kopito, R.R. Molecular biology of the anion exchanger gene family. Int. Rev. Cytol. 1990, 123, 177–199. [Google Scholar]
- Alper, S.L.; Sharma, A.K. The SLC26 gene family of anion transporters and channels. Mol. Asp. Med. 2013, 34, 494–515. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Shewan, A.M.; Thorn, P. HCO3− Transport through Anoctamin/Transmembrane Protein ANO1/TMEM16A in Pancreatic Acinar Cells Regulates Luminal pH. J. Biol. Chem. 2016, 291, 20345–20352. [Google Scholar] [CrossRef] [Green Version]
- Sondo, E.; Caci, E.; Galietta, L.J. The TMEM16A chloride channel as an alternative therapeutic target in cystic fibrosis. Int. J. Biochem. Cell Biol. 2014, 52, 73–76. [Google Scholar] [CrossRef]
- Puga Molina, L.C.; Pinto, N.A.; Torres, N.I.; Gonzalez-Cota, A.L.; Luque, G.M.; Balestrini, P.A.; Romarowski, A.; Krapf, D.; Santi, C.M.; Trevino, C.L.; et al. CFTR/ENaC-dependent regulation of membrane potential during human sperm capacitation is initiated by bicarbonate uptake through NBC. J. Biol. Chem. 2018, 293, 9924–9936. [Google Scholar] [CrossRef] [Green Version]
- Collawn, J.F.; Matalon, S. CFTR and lung homeostasis. Am. J. Physiol.-Lung Cell Mol. Physiol. 2014, 307, L917–L923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aalkjaer, C.; Boedtkjer, E.; Choi, I.; Lee, S. Cation-coupled bicarbonate transporters. Compr. Physiol. 2014, 4, 1605–1637. [Google Scholar] [PubMed] [Green Version]
- Felder, R.A.; Jose, P.A.; Xu, P.; Gildea, J.J. The Renal Sodium Bicarbonate Cotransporter NBCe2: Is It a Major Contributor to Sodium and pH Homeostasis? Curr. Hypertens. Rep. 2016, 18, 71. [Google Scholar] [CrossRef] [PubMed]
- Gildea, J.J.; Xu, P.; Kemp, B.A.; Carlson, J.M.; Tran, H.T.; Bigler Wang, D.; Langouet-Astrie, C.J.; McGrath, H.E.; Carey, R.M.; Jose, P.A.; et al. Sodium bicarbonate cotransporter NBCe2 gene variants increase sodium and bicarbonate transport in human renal proximal tubule cells. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, F.L.; Boedtkjer, E.; Witkowska, K.; Ren, M.; Zhang, R.; Tucker, A.; Aalkjaer, C.; Caulfield, M.J.; Ye, S. Increased NBCn1 expression, Na+/HCO3− co-transport and intracellular pH in human vascular smooth muscle cells with a risk allele for hypertension. Hum. Mol. Genet. 2017, 26, 989–1002. [Google Scholar]
- Pena-Munzenmayer, G.; George, A.T.; Shull, G.E.; Melvin, J.E.; Catalan, M.A. Ae4 (Slc4a9) is an electroneutral monovalent cation-dependent Cl−/HCO3− exchanger. J. Gen. Physiol. 2016, 147, 423–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampik, N.B.; Gehring, N.; Schnitzbauer, U.; Hennings, J.C.; Hubner, C.A.; Wagner, C.A. The murine Cl−/HCO3− exchanger Ae3 (Slc4a3) is not required for acid-base balance but is involved in magnesium handling by the kidney. Cell. Physiol. Biochem. 2014, 34, 1566–1577. [Google Scholar] [CrossRef]
- Su, Y.; Al-Lamki, R.S.; Blake-Palmer, K.G.; Best, A.; Golder, Z.J.; Zhou, A.; Karet Frankl, F.E. Physical and functional links between anion exchanger-1 and sodium pump. J. Am. Soc. Nephrol. 2015, 26, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Satoh, N.; Nakamura, M.; Suzuki, M.; Suzuki, A.; Seki, G.; Horita, S. Roles of Akt and SGK1 in the Regulation of Renal Tubular Transport. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.G.; Ohana, E.; Park, H.W.; Yang, D.; Muallem, S. Molecular mechanism of pancreatic and salivary gland fluid and HCO3 secretion. Physiol. Rev. 2012, 92, 39–74. [Google Scholar] [CrossRef] [Green Version]
- Şentürk, M.; Çavdar, H.; Talaz, O.; Supuran, C.T. Carbonic Anhydrase Inhibitors and Activators: Small Organic Molecules as Drugs and Prodrugs. In Medicinal Chemistry and Drug Design; Intech Open: Rijeka, Croatia, 2012. [Google Scholar]
- Widdas, W.F.; Baker, G.F.; Baker, P. The acceleration of pH volume changes in human red cells by bicarbonate and the role of carbonic anhydrase. Cytobios 1994, 80, 7–24. [Google Scholar] [PubMed]
- Jacob, P.; Christiani, S.; Rossmann, H.; Lamprecht, G.; Vieillard-Baron, D.; Muller, R.; Gregor, M.; Seidler, U. Role of Na+HCO3− cotransporter NBC1, Na+/H+ exchanger NHE1, and carbonic anhydrase in rabbit duodenal bicarbonate secretion. Gastroenterology 2000, 119, 406–419. [Google Scholar] [CrossRef] [PubMed]
- Knutson, T.W.; Koss, M.A.; Hogan, D.L.; Isenberg, J.I.; Knutson, L. Acetazolamide inhibits basal and stimulated HCO3- secretion in the human proximal duodenum. Gastroenterology 1995, 108, 102–107. [Google Scholar] [CrossRef]
- Mizumori, M.; Meyerowitz, J.; Takeuchi, T.; Lim, S.; Lee, P.; Supuran, C.T.; Guth, P.H.; Engel, E.; Kaunitz, J.D.; Akiba, Y. Epithelial carbonic anhydrases facilitate PCO2 and pH regulation in rat duodenal mucosa. J. Physiol. 2006, 573, 827–842. [Google Scholar] [CrossRef]
- Chen, M.; Praetorius, J.; Zheng, W.; Xiao, F.; Riederer, B.; Singh, A.K.; Stieger, N.; Wang, J.; Shull, G.E.; Aalkjaer, C.; et al. The electroneutral Na+:HCO3− cotransporter NBCn1 is a major pHi regulator in murine duodenum. J. Physiol. 2012, 590, 3317–3333. [Google Scholar] [CrossRef]
- Sterling, D.; Brown, N.J.; Supuran, C.T.; Casey, J.R. The functional and physical relationship between the DRA bicarbonate transporter and carbonic anhydrase II. Am. J. Physiol.-Cell Physiol. 2002, 283, C1522–C1529. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.H.; Muhammad, E.; Zheng, C.; Hershkovitz, E.; Alkrinawi, S.; Loewenthal, N.; Parvari, R.; Muallem, S. Essential role of carbonic anhydrase XII in secretory gland fluid and HCO3− secretion revealed by disease causing human mutation. J. Physiol. 2015, 593, 5299–5312. [Google Scholar] [CrossRef] [Green Version]
- Rafajova, M.; Zatovicova, M.; Kettmann, R.; Pastorek, J.; Pastorekova, S. Induction by hypoxia combined with low glucose or low bicarbonate and high posttranslational stability upon reoxygenation contribute to carbonic anhydrase IX expression in cancer cells. Int. J. Oncol. 2004, 24, 995–1004. [Google Scholar] [CrossRef]
- Svastova, E.; Witarski, W.; Csaderova, L.; Kosik, I.; Skvarkova, L.; Hulikova, A.; Zatovicova, M.; Barathova, M.; Kopacek, J.; Pastorek, J.; et al. Carbonic anhydrase IX interacts with bicarbonate transporters in lamellipodia and increases cell migration via its catalytic domain. J. Biol. Chem. 2012, 287, 3392–3402. [Google Scholar] [CrossRef] [Green Version]
- Parks, S.K.; Pouyssegur, J. The Na+/HCO3− Co-Transporter SLC4A4 Plays a Role in Growth and Migration of Colon and Breast Cancer Cells. J. Cell. Physiol. 2015, 230, 1954–1963. [Google Scholar] [CrossRef]
- Svichar, N.; Waheed, A.; Sly, W.S.; Hennings, J.C.; Hubner, C.A.; Chesler, M. Carbonic anhydrases CA4 and CA14 both enhance AE3-mediated Cl−-HCO3− exchange in hippocampal neurons. J. Neurosci. 2009, 29, 3252–3258. [Google Scholar] [CrossRef] [Green Version]
- Theparambil, S.M.; Naoshin, Z.; Thyssen, A.; Deitmer, J.W. Reversed electrogenic sodium bicarbonate cotransporter 1 is the major acid loader during recovery from cytosolic alkalosis in mouse cortical astrocytes. J. Physiol. 2015, 593, 3533–3547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villafuerte, F.C.; Swietach, P.; Youm, J.B.; Ford, K.; Cardenas, R.; Supuran, C.T.; Cobden, P.M.; Rohling, M.; Vaughan-Jones, R.D. Facilitation by intracellular carbonic anhydrase of Na+-HCO3− co-transport but not Na+/H+ exchange activity in the mammalian ventricular myocyte. J. Physiol. 2014, 592, 991–1007. [Google Scholar] [CrossRef] [PubMed]
- Peetz, J.; Barros, L.F.; San Martin, A.; Becker, H.M. Functional interaction between bicarbonate transporters and carbonic anhydrase modulates lactate uptake into mouse cardiomyocytes. Pflug. Arch.-Eur. J. Physiol. 2015, 467, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Lacruz, R.S.; Smith, C.E.; Moffatt, P.; Chang, E.H.; Bromage, T.G.; Bringas, P., Jr.; Nanci, A.; Baniwal, S.K.; Zabner, J.; Welsh, M.J.; et al. Requirements for ion and solute transport, and pH regulation during enamel maturation. J. Cell. Physiol. 2012, 227, 1776–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Shcheynikov, N.; Zeng, W.; Ohana, E.; So, I.; Ando, H.; Mizutani, A.; Mikoshiba, K.; Muallem, S. IRBIT coordinates epithelial fluid and HCO3− secretion by stimulating the transporters pNBC1 and CFTR in the murine pancreatic duct. J. Clin. Investig. 2009, 119, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Li, Q.; So, I.; Huang, C.L.; Ando, H.; Mizutani, A.; Seki, G.; Mikoshiba, K.; Thomas, P.J.; Muallem, S. IRBIT governs epithelial secretion in mice by antagonizing the WNK/SPAK kinase pathway. J. Clin. Investig. 2011, 121, 956–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.H.; Yang, D.; Shcheynikov, N.; Ohana, E.; Shin, D.M.; Muallem, S. Convergence of IRBIT, phosphatidylinositol (4,5) bisphosphate, and WNK/SPAK kinases in regulation of the Na+-HCO3− cotransporters family. Proc. Natl. Acad. Sci. USA 2013, 110, 4105–4110. [Google Scholar] [CrossRef] [Green Version]
- Park, H.W.; Nam, J.H.; Kim, J.Y.; Namkung, W.; Yoon, J.S.; Lee, J.S.; Kim, K.S.; Venglovecz, V.; Gray, M.A.; Kim, K.H.; et al. Dynamic regulation of CFTR bicarbonate permeability by [Cl−]i and its role in pancreatic bicarbonate secretion. Gastroenterology 2010, 139, 620–631. [Google Scholar] [CrossRef]
- Lee, D.; Lee, S.A.; Shin, D.M.; Hong, J.H. Chloride Influx of Anion Exchanger 2 Was Modulated by Calcium-Dependent Spinophilin in Submandibular Glands. Front. Physiol 2018, 9, 889. [Google Scholar] [CrossRef]
- Jeong, Y.S.; Hong, J.H. Governing effect of regulatory proteins for Cl−/HCO3− exchanger 2 activity. Channels 2016, 10, 214–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.H. Nanomaterials-Based Approaches for the Modulation of Sodium Bicarbonate Cotransporters. J. Nanomater. 2015. [Google Scholar] [CrossRef] [Green Version]
- Thornell, I.M.; Bevensee, M.O. Regulators of Slc4 bicarbonate transporter activity. Front. Physiol. 2015, 6, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, H.; Mizutani, A.; Kiefer, H.; Tsuzurugi, D.; Michikawa, T.; Mikoshiba, K. IRBIT suppresses IP3 receptor activity by competing with IP3 for the common binding site on the IP3 receptor. Mol. Cell 2006, 22, 795–806. [Google Scholar] [CrossRef]
- Shirakabe, K.; Priori, G.; Yamada, H.; Ando, H.; Horita, S.; Fujita, T.; Fujimoto, I.; Mizutani, A.; Seki, G.; Mikoshiba, K. IRBIT, an inositol 1,4,5-trisphosphate receptor-binding protein, specifically binds to and activates pancreas-type Na+/HCO3- cotransporter 1 (pNBC1). Proc. Natl. Acad. Sci. USA 2006, 103, 9542–9547. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.K.; Shcheynikov, N.; Muallem, S. IRBIT: It Is Everywhere. Neurochem. Res. 2011, 36, 1166–1174. [Google Scholar] [CrossRef]
- Kahle, K.T.; Gimenez, I.; Hassan, H.; Wilson, F.H.; Wong, R.D.; Forbush, B.; Aronson, P.S.; Lifton, R.P. WNK4 regulates apical and basolateral Cl− flux in extrarenal epithelia. Proc. Natl. Acad. Sci. USA 2004, 101, 2064–2069. [Google Scholar] [CrossRef] [Green Version]
- He, G.; Wang, H.R.; Huang, S.K.; Huang, C.L. Intersectin links WNK kinases to endocytosis of ROMK1. J. Clin. Investig. 2007, 117, 1078–1087. [Google Scholar] [CrossRef] [Green Version]
- Heise, C.J.; Xu, B.E.; Deaton, S.L.; Cha, S.K.; Cheng, C.J.; Earnest, S.; Sengupta, S.; Juang, Y.C.; Stippec, S.; Xu, Y.D.; et al. Serum and Glucocorticoid-induced Kinase (SGK) 1 and the Epithelial Sodium Channel Are Regulated by Multiple with No Lysine (WNK) Family Members. J. Biol. Chem. 2010, 285, 25161–25167. [Google Scholar] [CrossRef] [Green Version]
- Richardson, C.; Alessi, D.R. The regulation of salt transport and blood pressure by the WNK-SPAK/OSR1 signalling pathway. J. Cell Sci. 2008, 121, 3293–3304. [Google Scholar] [CrossRef] [Green Version]
- Vitari, A.C.; Deak, M.; Morrice, N.A.; Alessi, D.R. The WNK1 and WNK4 protein kinases that are mutated in Gordon’s hypertension syndrome phosphorylate and activate SPAK and OSR1 protein kinases. Biochem. J. 2005, 391, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Hong, J.H.; Ohana, E.; Muallem, S. The WNK/SPAK and IRBIT/PP1 pathways in epithelial fluid and electrolyte transport. Physiology 2012, 27, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Piechotta, K.; Garbarini, N.; England, R.; Delpire, E. Characterization of the interaction of the stress kinase SPAK with the Na+-K+-2Cl− cotransporter in the nervous system: Evidence for a scaffolding role of the kinase. J. Biol. Chem. 2003, 278, 52848–52856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarrouilhe, D.; di Tommaso, A.; Metaye, T.; Ladeveze, V. Spinophilin: From partners to functions. Biochimie 2006, 88, 1099–1113. [Google Scholar] [CrossRef]
- Wu, J.P.; McNicholas, C.M.; Bevensee, M.O. Phosphatidylinositol 4,5-bisphosphate (PIP2) stimulates the electrogenic Na/HCO3 cotransporter NBCe1-A expressed in Xenopus oocytes. Proc. Natl. Acad. Sci. USA 2009, 106, 14150–14155. [Google Scholar] [CrossRef] [Green Version]
- Thornell, I.M.; Wu, J.; Liu, X.; Bevensee, M.O. PIP2 hydrolysis stimulates the electrogenic Na+-bicarbonate cotransporter NBCe1-B and -C variants expressed in Xenopus laevis oocytes. J. Physiol. 2012, 590, 5993–6011. [Google Scholar] [CrossRef]
- Hilgemann, D.W.; Ball, R. Regulation of cardiac Na+,Ca2+ exchange and K-ATP potassium channels by PIP2. Science 1996, 273, 956–959. [Google Scholar] [CrossRef]
- He, Z.P.; Feng, S.Y.; Tong, Q.S.; Hilgemann, D.W.; Philipson, K.D. Interaction of PIP2 with the XIP region of the cardiac Na/Ca exchanger. Am. J. Physiol.-Cell Physiol. 2000, 278, C661–C666. [Google Scholar] [CrossRef] [Green Version]
- Aharonovitz, O.; Zaun, H.C.; Balla, T.; York, J.D.; Orlowski, J.; Grinstein, S. Intracellular pH regulation by Na+/H+ exchange requires phosphatidylinositol 4,5-bisphosphate. J. Cell Biol. 2000, 150, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Soboloff, J.; Spassova, M.; Hewavitharana, T.; He, L.P.; Luncsford, P.; Xu, W.; Venkatachalam, K.; van Rossum, D.; Patterson, R.L.; Gill, D.L. TRPC channels: Integrators of multiple cellular signals. Handb. Exp. Pharmacol. 2007, 179, 575–591. [Google Scholar]
- Bernier, L.P.; Ase, A.R.; Seguela, P. Post-translational regulation of P2X receptor channels: Modulation by phospholipids. Front. Cell. Neurosci. 2013, 7, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier, L.P.; Blais, D.; Boue-Grabot, E.; Seguela, P. A dual polybasic motif determines phosphoinositide binding and regulation in the P2X channel family. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, E.; Crespo, P. The RAS-ERK pathway: A route for couples. Sci. Signal. 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Luscher, B.P.; Vachel, L.; Ohana, E.; Muallem, S. Cl− as a bona fide signalling ion. Am. J. Physiol.-Cell Physiol. 2019. [Google Scholar] [CrossRef]
- Dutzler, R.; Campbell, E.B.; Cadene, M.; Chait, B.T.; MacKinnon, R. X-ray structure of a ClC chloride channel at 3.0 A reveals the molecular basis of anion selectivity. Nature 2002, 415, 287–294. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Ishiguro, H.; Steward, M.; Sohma, Y.; Yamamoto, A.; Shimouchi, A.; Kondo, T. Apical Cl−/HCO3− exchanger stoichiometry in the modeling of HCO3− transport by pancreatic duct epithelium. J. Med. Investig. 2009, 56, 325–328. [Google Scholar] [CrossRef] [Green Version]
- Eladari, D.; Chambrey, R.; Picard, N.; Hadchouel, J. Electroneutral absorption of NaCl by the aldosterone-sensitive distal nephron: Implication for normal electrolytes homeostasis and blood pressure regulation. Cell. Mol. Life Sci. 2014, 71, 2879–2895. [Google Scholar] [CrossRef]
- Edwards, A.; Crambert, G. Versatility of NaCl transport mechanisms in the cortical collecting duct. Am. J. Physiol.-Renal Physiol. 2017, 313, F1254–F1263. [Google Scholar] [CrossRef]
- Shcheynikov, N.; Son, A.; Hong, J.H.; Yamazaki, O.; Ohana, E.; Kurtz, I.; Shin, D.M.; Muallem, S. Intracellular Cl− as a signaling ion that potently regulates Na+/HCO3− transporters. Proc. Natl. Acad. Sci. USA 2015, 112, E329–E337. [Google Scholar] [CrossRef] [Green Version]
- Kahle, K.T.; Ring, A.M.; Lifton, R.P. Molecular physiology of the WNK kinases. Annu. Rev. Physiol. 2008, 70, 329–355. [Google Scholar] [CrossRef]
- Alka, K.; Casey, J.R. Bicarbonate transport in health and disease. IUBMB Life 2014, 66, 596–615. [Google Scholar] [CrossRef] [PubMed]
- Levin, L.R.; Buck, J. Physiological roles of acid-base sensors. Annu. Rev. Physiol. 2015, 77, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, K.F.; Nauli, S.M. pH sensors and ion Transporters: Potential therapeutic targets for acid-base disorders. Int. J. Pharma Res. Rev. 2016, 5, 51–58. [Google Scholar] [PubMed]
- Tang, X.X.; Fok, K.L.; Chen, H.; Chan, K.S.; Tsang, L.L.; Rowlands, D.K.; Zhang, X.H.; Dong, J.D.; Ruan, Y.C.; Jiang, X.; et al. Lymphocyte CFTR promotes epithelial bicarbonate secretion for bacterial killing. J. Cell. Physiol. 2012, 227, 3887–3894. [Google Scholar] [CrossRef]
- Sedlyarov, V.; Eichner, R.; Girardi, E.; Essletzbichler, P.; Goldmann, U.; Nunes-Hasler, P.; Srndic, I.; Moskovskich, A.; Heinz, L.X.; Kartnig, F.; et al. The Bicarbonate Transporter SLC4A7 Plays a Key Role in Macrophage Phagosome Acidification. Cell Host Microbe 2018, 23, 766–774. [Google Scholar] [CrossRef] [Green Version]
- de la Rosa, L.A.; Cabado, A.G.; Botana, M.A.; Vieytes, M.R.; Vidal, J.I.; Botana, L.M. Evidence for an electrogenic, negatively protein-kinase-A-modulated, Na+-dependent HCO3− transporter in human lymphocytes. Pflug. Arch.-Eur. J. Physiol. 1999, 437, 935–943. [Google Scholar] [CrossRef]
- Giambelluca, M.S.; Ciancio, M.C.; Orlowski, A.; Gende, O.A.; Pouliot, M.; Aiello, E.A. Characterization of the Na/HCO3− cotransport in human neutrophils. Cell. Physiol. Biochem. 2014, 33, 982–990. [Google Scholar] [CrossRef]
- Giambelluca, M.S.; Gende, O.A. Cl−/HCO3− exchange activity in fMLP-stimulated human neutrophils. Biochem. Biophys. Res. Commun. 2011, 409, 567–571. [Google Scholar] [CrossRef]
- Concepcion, A.R.; Salas, J.T.; Sarvide, S.; Saez, E.; Ferrer, A.; Lopez, M.; Portu, A.; Banales, J.M.; Hervas-Stubbs, S.; Oude Elferink, R.P.; et al. Anion exchanger 2 is critical for CD8(+) T cells to maintain pHi homeostasis and modulate immune responses. Eur. J. Immunol. 2014, 44, 1341–1351. [Google Scholar] [CrossRef]
- Concepcion, A.R.; Salas, J.T.; Saez, E.; Sarvide, S.; Ferrer, A.; Portu, A.; Uriarte, I.; Hervas-Stubbs, S.; Oude Elferink, R.P.; Prieto, J.; et al. CD8+ T cells undergo activation and programmed death-1 repression in the liver of aged Ae2a,b-/-mice favoring autoimmune cholangitis. Oncotarget 2015, 6, 28588–28606. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.P.; Samsoe-Petersen, J.; Oernbo, E.K.; Boedtkjer, E.; Moreira, J.M.A.; Kveiborg, M.; Pedersen, S.F. The net acid extruders NHE1, NBCn1 and MCT4 promote mammary tumor growth through distinct but overlapping mechanisms. Int. J. Cancer 2018, 142, 2529–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Axelsen, T.V.; Andersen, A.P.; Vahl, P.; Pedersen, S.F.; Boedtkjer, E. Disrupting Na+, HCO3−-cotransporter NBCn1 (Slc4a7) delays murine breast cancer development. Oncogene 2016, 35, 2112–2122. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Mele, M.; Vahl, P.; Christiansen, P.M.; Jensen, V.E.; Boedtkjer, E. Na+, HCO3−-cotransport is functionally upregulated during human breast carcinogenesis and required for the inverted pH gradient across the plasma membrane. Pflug. Arch.-Eur. J. Physiol. 2015, 467, 367–377. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, A.; Hulikova, A.; Ledaki, I.; Snell, C.; Singleton, D.; Steers, G.; Seden, P.; Jones, D.; Bridges, E.; Wigfield, S.; et al. Disrupting Hypoxia-Induced Bicarbonate Transport Acidifies Tumor Cells and Suppresses Tumor Growth. Cancer Res. 2016, 76, 3744–3755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wykoff, C.C.; Beasley, N.J.; Watson, P.H.; Turner, K.J.; Pastorek, J.; Sibtain, A.; Wilson, G.D.; Turley, H.; Talks, K.L.; Maxwell, P.H.; et al. Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res. 2000, 60, 7075–7083. [Google Scholar]
- Watson, P.H.; Chia, S.K.; Wykoff, C.C.; Han, C.; Leek, R.D.; Sly, W.S.; Gatter, K.C.; Ratcliffe, P.; Harris, A.L. Carbonic anhydrase XII is a marker of good prognosis in invasive breast carcinoma. Br. J. Cancer 2003, 88, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Chiche, J.; Ilc, K.; Laferriere, J.; Trottier, E.; Dayan, F.; Mazure, N.M.; Brahimi-Horn, M.C.; Pouyssegur, J. Hypoxia-inducible carbonic anhydrase IX and XII promote tumor cell growth by counteracting acidosis through the regulation of the intracellular pH. Cancer Res. 2009, 69, 358–368. [Google Scholar] [CrossRef] [Green Version]
- Ambrosio, M.R.; Di Serio, C.; Danza, G.; Rocca, B.J.; Ginori, A.; Prudovsky, I.; Marchionni, N.; Del Vecchio, M.T.; Tarantini, F. Carbonic anhydrase IX is a marker of hypoxia and correlates with higher Gleason scores and ISUP grading in prostate cancer. Diagn. Pathol. 2016, 11, 45. [Google Scholar] [CrossRef] [Green Version]
- Finkelmeier, F.; Canli, O.; Peiffer, K.H.; Walter, D.; Tal, A.; Koch, C.; Pession, U.; Vermehren, J.; Trojan, J.; Zeuzem, S.; et al. Circulating hypoxia marker carbonic anhydrase IX (CA9) in patients with hepatocellular carcinoma and patients with cirrhosis. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Becker, H.M. Carbonic anhydrase IX and acid transport in cancer. Br. J. Cancer 2019. [Google Scholar] [CrossRef]
- Becker, H.M.; Klier, M.; Deitmer, J.W. Carbonic anhydrases and their interplay with acid/base-coupled membrane transporters. Subcell. Biochem. 2014, 75, 105–134. [Google Scholar] [PubMed]
- Gorbatenko, A.; Olesen, C.W.; Boedtkjer, E.; Pedersen, S.F. Regulation and roles of bicarbonate transporters in cancer. Front. Physiol. 2014, 5, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, S.C.; Giannuzzo, A.; Novak, I.; Pedersen, S.F. Acid-base transport in pancreatic cancer: Molecular mechanisms and clinical potential. Biochem. Cell Biol. 2014, 92, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Kant, S.; Kumar, A.; Singh, S.M. Bicarbonate transport inhibitor SITS modulates pH homeostasis triggering apoptosis of Dalton’s lymphoma: Implication of novel molecular mechanisms. Mol. Cell. Biochem. 2014, 397, 167–178. [Google Scholar] [CrossRef]
- Song, L.J.; Liu, R.J.; Zeng, Z.; Alper, S.L.; Cui, H.J.; Lu, Y.; Zheng, L.; Yan, Z.W.; Fu, G.H. Gastrin inhibits a novel, pathological colon cancer signaling pathway involving EGR1, AE2, and P-ERK. J. Mol. Med. 2012, 90, 707–718. [Google Scholar] [CrossRef]
- Chapman, J.M.; Knoepp, S.M.; Byeon, M.K.; Henderson, K.W.; Schweinfest, C.W. The colon anion transporter, down-regulated in adenoma, induces growth suppression that is abrogated by E1A. Cancer Res. 2002, 62, 5083–5088. [Google Scholar]
- Schweinfest, C.W.; Spyropoulos, D.D.; Henderson, K.W.; Kim, J.H.; Chapman, J.M.; Barone, S.; Worrell, R.T.; Wang, Z.; Soleimani, M. slc26a3 (dra)-deficient mice display chloride-losing diarrhea, enhanced colonic proliferation, and distinct up-regulation of ion transporters in the colon. J. Biol. Chem. 2006, 281, 37962–37971. [Google Scholar] [CrossRef] [Green Version]
- Boedtkjer, E.; Moreira, J.M.; Mele, M.; Vahl, P.; Wielenga, V.T.; Christiansen, P.M.; Jensen, V.E.; Pedersen, S.F.; Aalkjaer, C. Contribution of Na+, HCO3−-cotransport to cellular pH control in human breast cancer: A role for the breast cancer susceptibility locus NBCn1 (SLC4A7). Int. J. Cancer 2013, 132, 1288–1299. [Google Scholar] [CrossRef]
- Lauritzen, G.; Jensen, M.B.; Boedtkjer, E.; Dybboe, R.; Aalkjaer, C.; Nylandsted, J.; Pedersen, S.F. NBCn1 and NHE1 expression and activity in DeltaNErbB2 receptor-expressing MCF-7 breast cancer cells: Contributions to pHi regulation and chemotherapy resistance. Exp. Cell Res. 2010, 316, 2538–2553. [Google Scholar] [CrossRef]
- Lauritzen, G.; Stock, C.M.; Lemaire, J.; Lund, S.F.; Jensen, M.F.; Damsgaard, B.; Petersen, K.S.; Wiwel, M.; Ronnov-Jessen, L.; Schwab, A.; et al. The Na+/H+ exchanger NHE1, but not the Na+, HCO3− cotransporter NBCn1, regulates motility of MCF7 breast cancer cells expressing constitutively active ErbB2. Cancer Lett. 2012, 317, 172–183. [Google Scholar] [CrossRef]
- Yin, K.; Paine, M.L. Bicarbonate Transport During Enamel Maturation. Calcif. Tissue Int. 2017, 101, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Bronckers, A.L. Ion Transport by Ameloblasts during Amelogenesis. J. Dent. Res. 2017, 96, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Jalali, R.; Guo, J.; Zandieh-Doulabi, B.; Bervoets, T.J.; Paine, M.L.; Boron, W.F.; Parker, M.D.; Bijvelds, M.J.; Medina, J.F.; DenBesten, P.K.; et al. NBCe1 (SLC4A4) a potential pH regulator in enamel organ cells during enamel development in the mouse. Cell Tissue Res. 2014, 358, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalali, R.; Zandieh-Doulabi, B.; DenBesten, P.K.; Seidler, U.; Riederer, B.; Wedenoja, S.; Micha, D.; Bronckers, A.L. Slc26a3/Dra and Slc26a6 in Murine Ameloblasts. J. Dent. Res. 2015, 94, 1732–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, K.; Lei, Y.; Wen, X.; Lacruz, R.S.; Soleimani, M.; Kurtz, I.; Snead, M.L.; White, S.N.; Paine, M.L. SLC26A Gene Family Participate in pH Regulation during Enamel Maturation. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Bori, E.; Guo, J.; Racz, R.; Burghardt, B.; Foldes, A.; Keremi, B.; Harada, H.; Steward, M.C.; Den Besten, P.; Bronckers, A.L.; et al. Evidence for Bicarbonate Secretion by Ameloblasts in a Novel Cellular Model. J. Dent. Res. 2016, 95, 588–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.S.; Lee, S.P.; Huang, S.F.; Chao, S.C.; Chang, C.Y.; Wu, G.J.; Li, C.H.; Loh, S.H. Functional and molecular characterization of transmembrane intracellular pH regulators in human dental pulp stem cells. Arch. Oral Biol. 2018, 90, 19–26. [Google Scholar] [CrossRef]
- Danielsen, A.A.; Parker, M.D.; Lee, S.; Boron, W.F.; Aalkjaer, C.; Boedtkjer, E. Splice cassette II of Na+, HCO3− cotransporter NBCn1 (slc4a7) interacts with calcineurin A: Implications for transporter activity and intracellular pH control during rat artery contractions. J. Biol. Chem. 2013, 288, 8146–8155. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, A.B.; Kim, S.; Aalbaek, F.; Aalkjaer, C.; Boedtkjer, E. Intracellular acidification alters myogenic responsiveness and vasomotion of mouse middle cerebral arteries. J. Cereb. Blood Flow Metab. 2014, 34, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Boedtkjer, E.; Praetorius, J.; Matchkov, V.V.; Stankevicius, E.; Mogensen, S.; Fuchtbauer, A.C.; Simonsen, U.; Fuchtbauer, E.M.; Aalkjaer, C. Disruption of Na+, HCO3− cotransporter NBCn1 (slc4a7) inhibits NO-mediated vasorelaxation, smooth muscle Ca2+ sensitivity, and hypertension development in mice. Circulation 2011, 124, 1819–1829. [Google Scholar] [CrossRef] [Green Version]
- Loh, S.H.; Lee, C.Y.; Chen, G.S.; Wu, C.H.; Tsao, C.J.; Shih, S.J.; Chou, C.C.; Tsai, C.S.; Tsai, Y.T. The Effect and Underlying Mechanism of Ethanol on Intracellular H+-Equivalent Membrane Transporters in Human Aorta Smooth Muscle Cells. Alcohol. Clin. Exp. Res. 2015, 39, 2302–2312. [Google Scholar] [CrossRef] [PubMed]
- Boedtkjer, E.; Bentzon, J.F.; Dam, V.S.; Aalkjaer, C. Na+, HCO3−-cotransporter NBCn1 increases pH gradients, filopodia, and migration of smooth muscle cells and promotes arterial remodelling. Cardiovasc. Res. 2016, 111, 227–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boedtkjer, E.; Praetorius, J.; Aalkjaer, C. NBCn1 (slc4a7) mediates the Na+-dependent bicarbonate transport important for regulation of intracellular pH in mouse vascular smooth muscle cells. Circ. Res. 2006, 98, 515–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonde, L.; Boedtkjer, E. Extracellular acidosis and very low [Na(+) ] inhibit NBCn1- and NHE1-mediated net acid extrusion from mouse vascular smooth muscle cells. Acta Physiol. 2017, 221, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Garciarena, C.D.; Ma, Y.L.; Swietach, P.; Huc, L.; Vaughan-Jones, R.D. Sarcolemmal localisation of Na+/H+ exchange and Na+-HCO3− co-transport influences the spatial regulation of intracellular pH in rat ventricular myocytes. J. Physiol. 2013, 591, 2287–2306. [Google Scholar] [CrossRef] [PubMed]
- Karmazyn, M.; Moffat, M.P. Role of Na+/H+ exchange in cardiac physiology and pathophysiology: Mediation of myocardial reperfusion injury by the pH paradox. Cardiovasc. Res. 1993, 27, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, B.V.; Kieller, D.M.; Quon, A.L.; Markovich, D.; Casey, J.R. Slc26a6: A cardiac chloride-hydroxyl exchanger and predominant chloride-bicarbonate exchanger of the mouse heart. J. Physiol. 2004, 561, 721–734. [Google Scholar] [CrossRef]
- Kim, H.J.; Myers, R.; Sihn, C.R.; Rafizadeh, S.; Zhang, X.D. Slc26a6 functions as an electrogenic Cl−/HCO3− exchanger in cardiac myocytes. Cardiovasc. Res. 2013, 100, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Fantinelli, J.C.; Orlowski, A.; Aiello, E.A.; Mosca, S.M. The electrogenic cardiac sodium bicarbonate co-transporter (NBCe1) contributes to the reperfusion injury. Cardiovasc. Pathol. 2014, 23, 224–230. [Google Scholar] [CrossRef]
- Alvarez, B.V.; Loiselle, F.B.; Supuran, C.T.; Schwartz, G.J.; Casey, J.R. Direct extracellular interaction between carbonic anhydrase IV and the human NBC1 sodium/bicarbonate co-transporter. Biochemistry 2003, 42, 12321–12329. [Google Scholar] [CrossRef]
- Casey, J.R.; Sly, W.S.; Shah, G.N.; Alvarez, B.V. Bicarbonate homeostasis in excitable tissues: Role of AE3 Cl−/HCO3− exchanger and carbonic anhydrase XIV interaction. Am. J. Physiol.-Cell Physiol. 2009, 297, 1091–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, P.E.; Pastorekova, S.; Stuart-Tilley, A.K.; Alper, S.L.; Casey, J.R. Interactions of transmembrane carbonic anhydrase, CAIX, with bicarbonate transporters. Am. J. Physiol.-Cell Physiol. 2007, 293, 738–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Giusti, V.C.; Orlowski, A.; Ciancio, M.C.; Espejo, M.S.; Gonano, L.A.; Caldiz, C.I.; Vila Petroff, M.G.; Villa-Abrille, M.C.; Aiello, E.A. Aldosterone stimulates the cardiac sodium/bicarbonate cotransporter via activation of the g protein-coupled receptor gpr30. J. Mol. Cell. Cardiol. 2015, 89, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Safsten, B. Duodenal bicarbonate secretion and mucosal protection. Neurohumoral influence and transport mechanisms. Acta Physiol. Scand. Suppl. 1993, 613, 1–43. [Google Scholar] [PubMed]
- Rossmann, H.; Bachmann, O.; Vieillard-Baron, D.; Gregor, M.; Seidler, U. Na+/HCO3− cotransport and expression of NBC1 and NBC2 in rabbit gastric parietal and mucous cells. Gastroenterology 1999, 116, 1389–1398. [Google Scholar] [CrossRef]
- Seidler, U.; Rossmann, H.; Jacob, P.; Bachmann, O.; Christiani, S.; Lamprecht, G.; Gregor, M. Expression and function of Na+HCO3− cotransporters in the gastrointestinal tract. Ann. N. Y. Acad. Sci. 2000, 915, 1–14. [Google Scholar] [CrossRef]
- Singh, A.K.; Xia, W.; Riederer, B.; Juric, M.; Li, J.; Zheng, W.; Cinar, A.; Xiao, F.; Bachmann, O.; Song, P.; et al. Essential role of the electroneutral Na+-HCO3− cotransporter NBCn1 in murine duodenal acid-base balance and colonic mucus layer build-up in vivo. J. Physiol. 2013, 591, 2189–2204. [Google Scholar] [CrossRef] [Green Version]
- Barmeyer, C.; Ye, J.H.; Soroka, C.; Geibel, P.; Hingsammer, L.M.; Weitgasser, L.; Atway, D.; Geibel, J.P.; Binder, H.J.; Rajendran, V.M. Identification of functionally distinct Na-HCO3 co-transporters in colon. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Stuart-Tilley, A.; Sardet, C.; Pouyssegur, J.; Schwartz, M.A.; Brown, D.; Alper, S.L. Immunolocalization of anion exchanger AE2 and cation exchanger NHE-1 in distinct adjacent cells of gastric mucosa. Am. J. Physiol. 1994, 266, C559–C568. [Google Scholar] [CrossRef]
- Liu, X.; Li, T.; Riederer, B.; Lenzen, H.; Ludolph, L.; Yeruva, S.; Tuo, B.; Soleimani, M.; Seidler, U. Loss of Slc26a9 anion transporter alters intestinal electrolyte and HCO3− transport and reduces survival in CFTR-deficient mice. Pflug. Arch. Eur. J. Physiol. 2015, 467, 1261–1275. [Google Scholar] [CrossRef] [Green Version]
- Niv, Y.; Fraser, G.M. The alkaline tide phenomenon. J. Clin. Gastroenterol. 2002, 35, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.C.; Sun, X. SLC26 anion exchangers in uterine epithelial cells and spermatozoa: Clues from the past and hints to the future. Cell Biol. Int. 2014, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bernardino, R.L.; Martins, A.D.; Jesus, T.T.; Sa, R.; Sousa, M.; Alves, M.G.; Oliveira, P.F. Estrogenic regulation of bicarbonate transporters from SLC4 family in rat Sertoli cells. Mol. Cell. Biochem. 2015, 408, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Bernardino, R.L.; Costa, A.R.; Martins, A.D.; Silva, J.; Barros, A.; Sousa, M.; Sa, R.; Alves, M.G.; Oliveira, P.F. Estradiol modulates Na+ -dependent HCO3− transporters altering intracellular pH and ion transport in human Sertoli cells: A role on male fertility? Biol. Cell 2016, 108, 179–188. [Google Scholar] [CrossRef]
- Bi, R.Y.; Meng, Z.; Zhang, P.; Wang, X.D.; Ding, Y.; Gan, Y.H. Estradiol upregulates voltage-gated sodium channel 1.7 in trigeminal ganglion contributing to hyperalgesia of inflamed TMJ. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Ren, P.; Wang, W.B.; Pan, H.H.; Qiu, C.Y.; Hu, W.P. Up-regulation of ASIC3 expression by beta-estradiol. Neurosci. Lett. 2018, 684, 200–204. [Google Scholar] [CrossRef]
- Yang, X.; Mao, X.; Xu, G.; Xing, S.; Chattopadhyay, A.; Jin, S.; Salama, G. Estradiol up-regulates L-type Ca2+ channels via membrane-bound estrogen receptor/phosphoinositide-3-kinase/Akt/cAMP response element-binding protein signaling pathway. Heart Rhythm 2018, 15, 741–749. [Google Scholar] [CrossRef]
- Hill, B.J.F.; Dalton, R.J.; Joseph, B.K.; Thakali, K.M.; Rusch, N.J. 17beta-estradiol reduces Cav 1.2 channel abundance and attenuates Ca2+-dependent contractions in coronary arteries. Pharmacol. Res. Perspect. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Deng, J.; Wang, D.X.; He, J.; Deng, W. Regulation of epithelial sodium channel expression by oestradiol and progestogen in alveolar epithelial cells. Respir. Physiol. Neurobiol. 2015, 216, 52–62. [Google Scholar] [CrossRef]
- Diao, R.; Fok, K.L.; Zhao, L.; Chen, H.; Tang, H.; Chen, J.; Zheng, A.; Zhang, X.; Gui, Y.; Chan, H.C.; et al. Decreased expression of cystic fibrosis transmembrane conductance regulator impairs sperm quality in aged men. Reproduction 2013, 146, 637–645. [Google Scholar] [CrossRef] [Green Version]
- Pierucci-Alves, F.; Akoyev, V.; Stewart, J.C., 3rd; Wang, L.H.; Janardhan, K.S.; Schultz, B.D. Swine models of cystic fibrosis reveal male reproductive tract phenotype at birth. Biol. Reprod. 2011, 85, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Hihnala, S.; Kujala, M.; Toppari, J.; Kere, J.; Holmberg, C.; Hoglund, P. Expression of SLC26A3, CFTR and NHE3 in the human male reproductive tract: Role in male subfertility caused by congenital chloride diarrhoea. Mol. Hum. Reprod. 2006, 12, 107–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Clarke, L.; Nie, R.; Carnes, K.; Lai, L.W.; Lien, Y.H.; Verkman, A.; Lubahn, D.; Fisher, J.S.; Katzenellenbogen, B.S.; et al. Estrogen action and male fertility: Roles of the sodium/hydrogen exchanger-3 and fluid reabsorption in reproductive tract function. Proc. Natl. Acad. Sci. USA 2001, 98, 14132–14137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Royaux, I.E.; Everett, L.A.; Mori-Aoki, A.; Suzuki, S.; Nakamura, K.; Sakai, T.; Katoh, R.; Toda, S.; Green, E.D.; et al. Expression of PDS/Pds, the Pendred syndrome gene, in endometrium. J. Clin. Endocrinol. Metab. 2002, 87, 938–941. [Google Scholar] [CrossRef]
- Gholami, K.; Muniandy, S.; Salleh, N. Modulation of sodium-bicarbonate co-transporter (SLC4A4/NBCe1) protein and mRNA expression in rat’s uteri by sex-steroids and at different phases of the oestrous cycle. Res. Vet. Sci. 2014, 96, 164–170. [Google Scholar] [CrossRef]
- Xie, Z.D.; Guo, Y.M.; Ren, M.J.; Yang, J.; Wang, S.F.; Xu, T.H.; Chen, L.M.; Liu, Y. The Balance of HCO3− Secretion vs. Reabsorption in the Endometrial Epithelium Regulates Uterine Fluid pH. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Hong, J.H. The Fundamental Role of Bicarbonate Transporters and Associated Carbonic Anhydrase Enzymes in Maintaining Ion and pH Homeostasis in Non-Secretory Organs. Int. J. Mol. Sci. 2020, 21, 339. https://doi.org/10.3390/ijms21010339
Lee D, Hong JH. The Fundamental Role of Bicarbonate Transporters and Associated Carbonic Anhydrase Enzymes in Maintaining Ion and pH Homeostasis in Non-Secretory Organs. International Journal of Molecular Sciences. 2020; 21(1):339. https://doi.org/10.3390/ijms21010339
Chicago/Turabian StyleLee, Dongun, and Jeong Hee Hong. 2020. "The Fundamental Role of Bicarbonate Transporters and Associated Carbonic Anhydrase Enzymes in Maintaining Ion and pH Homeostasis in Non-Secretory Organs" International Journal of Molecular Sciences 21, no. 1: 339. https://doi.org/10.3390/ijms21010339