Molecular Mapping and Candidate Gene Analysis for GA3 Responsive Short Internode in Watermelon (Citrullus lanatus)

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
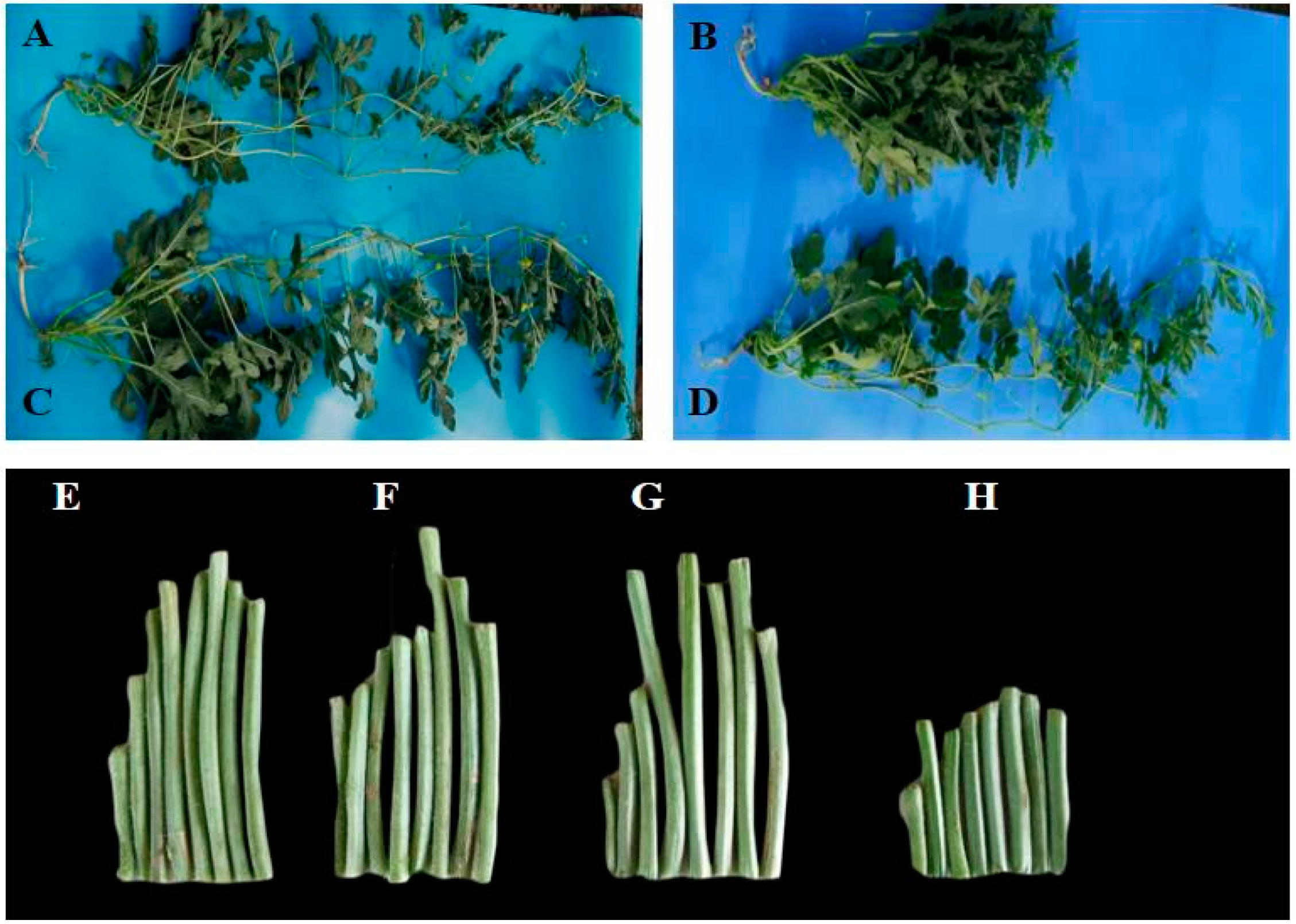
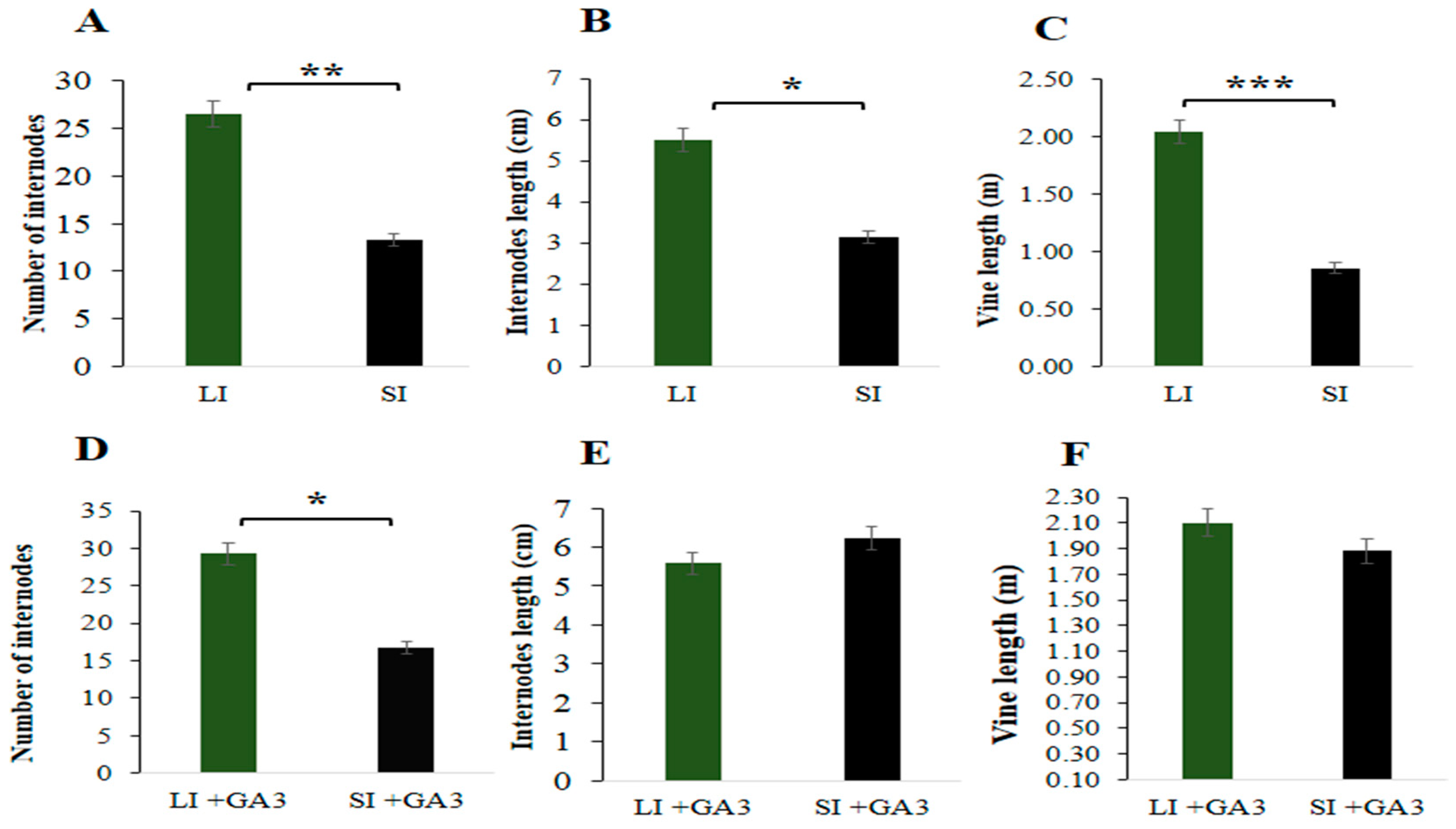
2.1. Agronomic Characteristics and Inheritance of the Short Internode
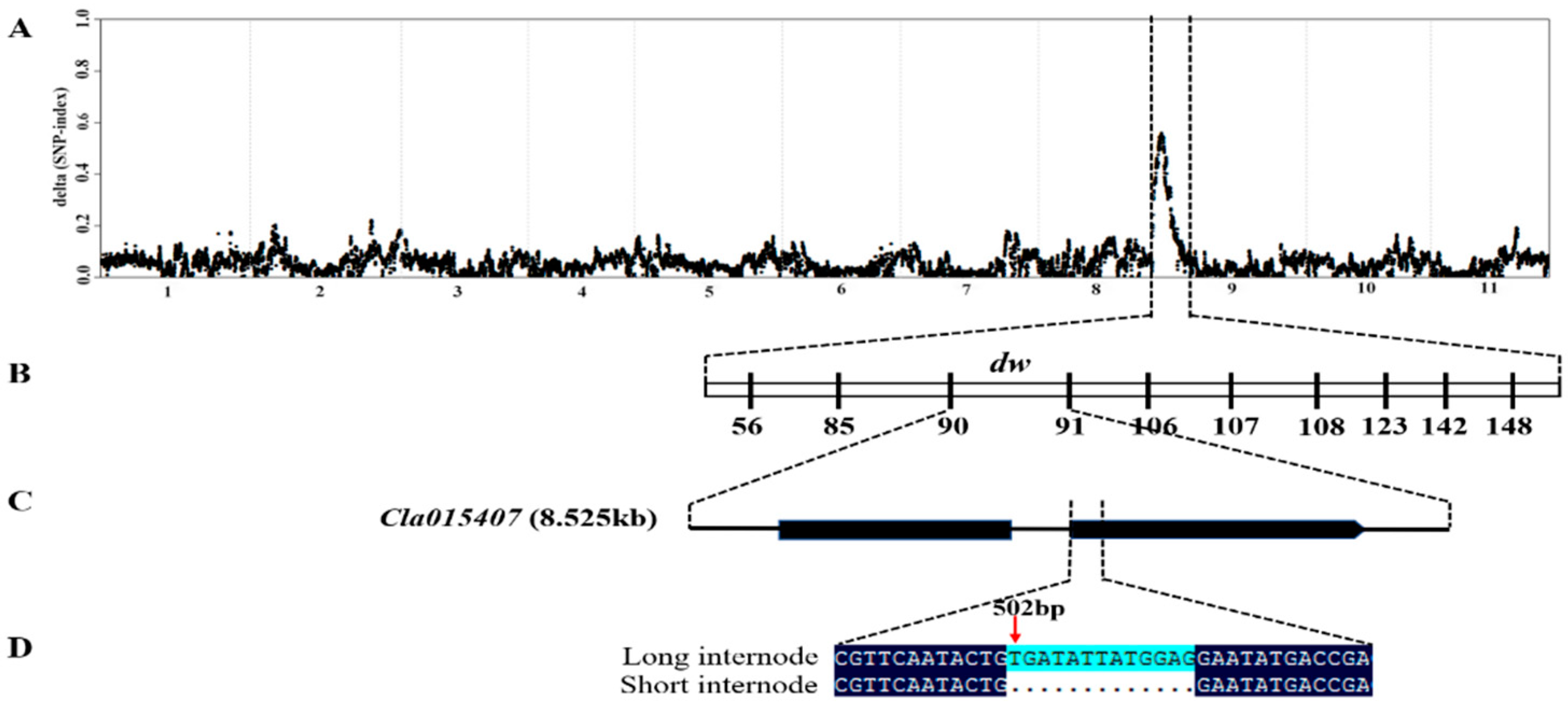
2.2. BSA-Seq Analysis Identified a Candidate Gene Located on Chromosome 9
2.3. Fine Mapping, Annotation and Candidate Gene Sequencing
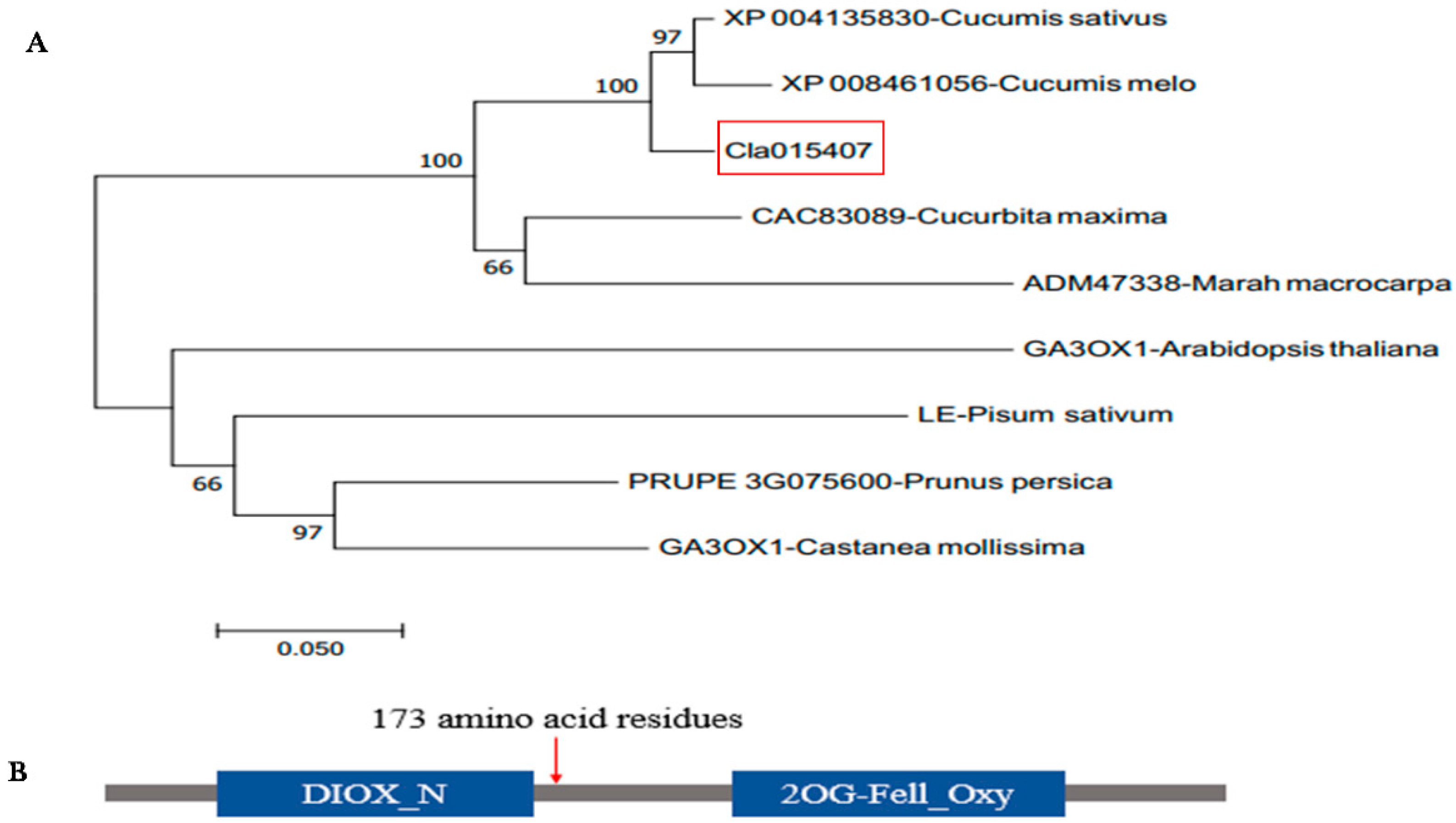
2.4. Homology, Phylogenic Tree and Conseved Domain Analysis
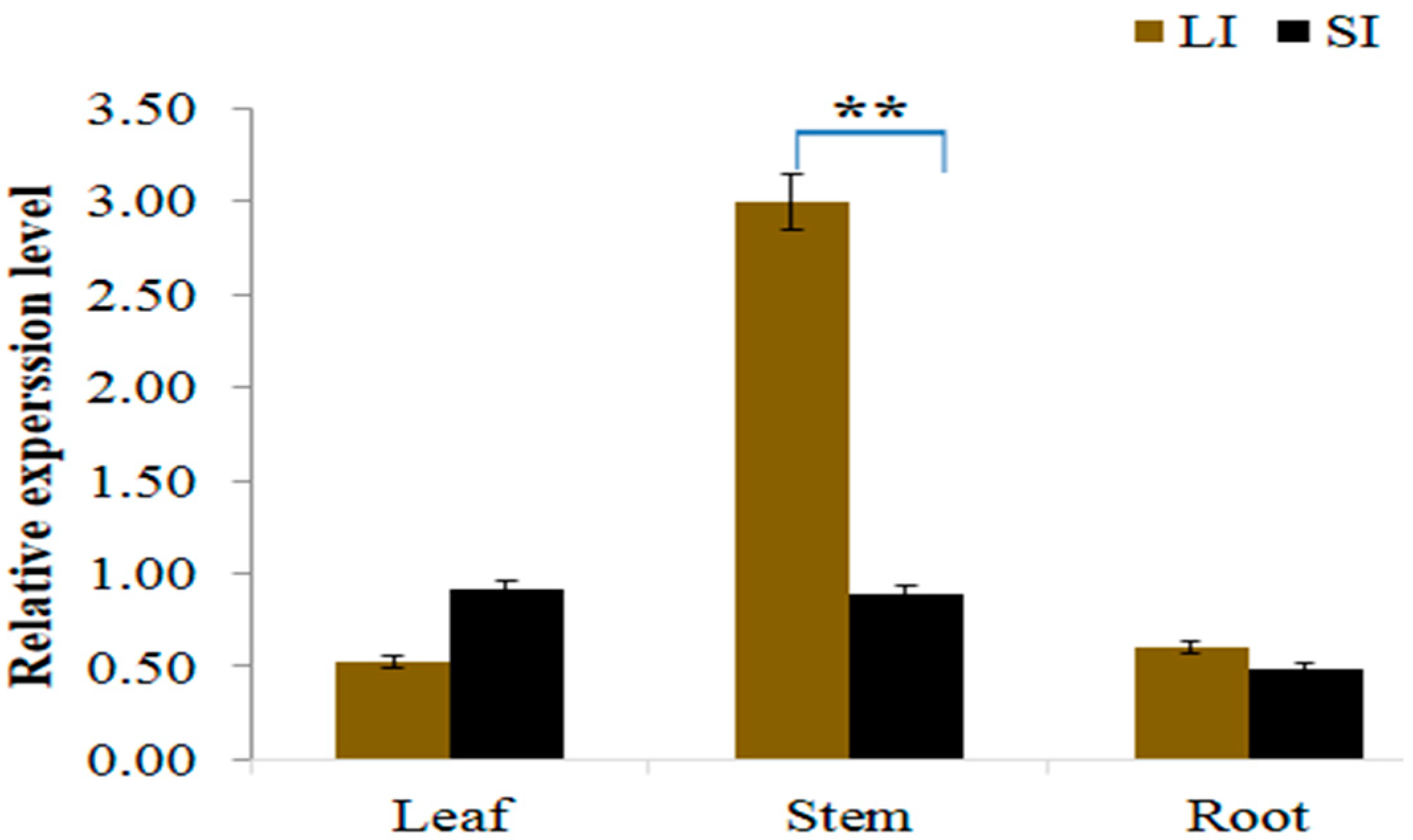
2.5. Gene Expression Analysis
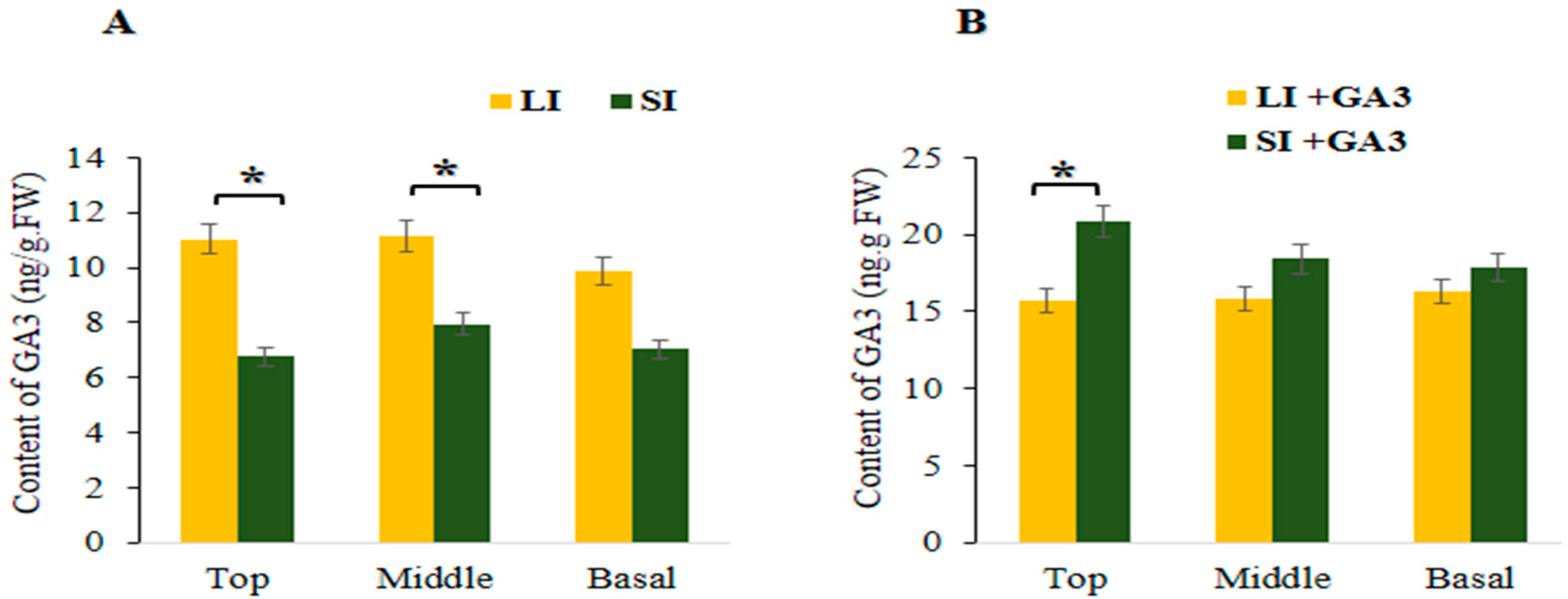
2.6. Determination of GA3 Hormone in the Short Internode
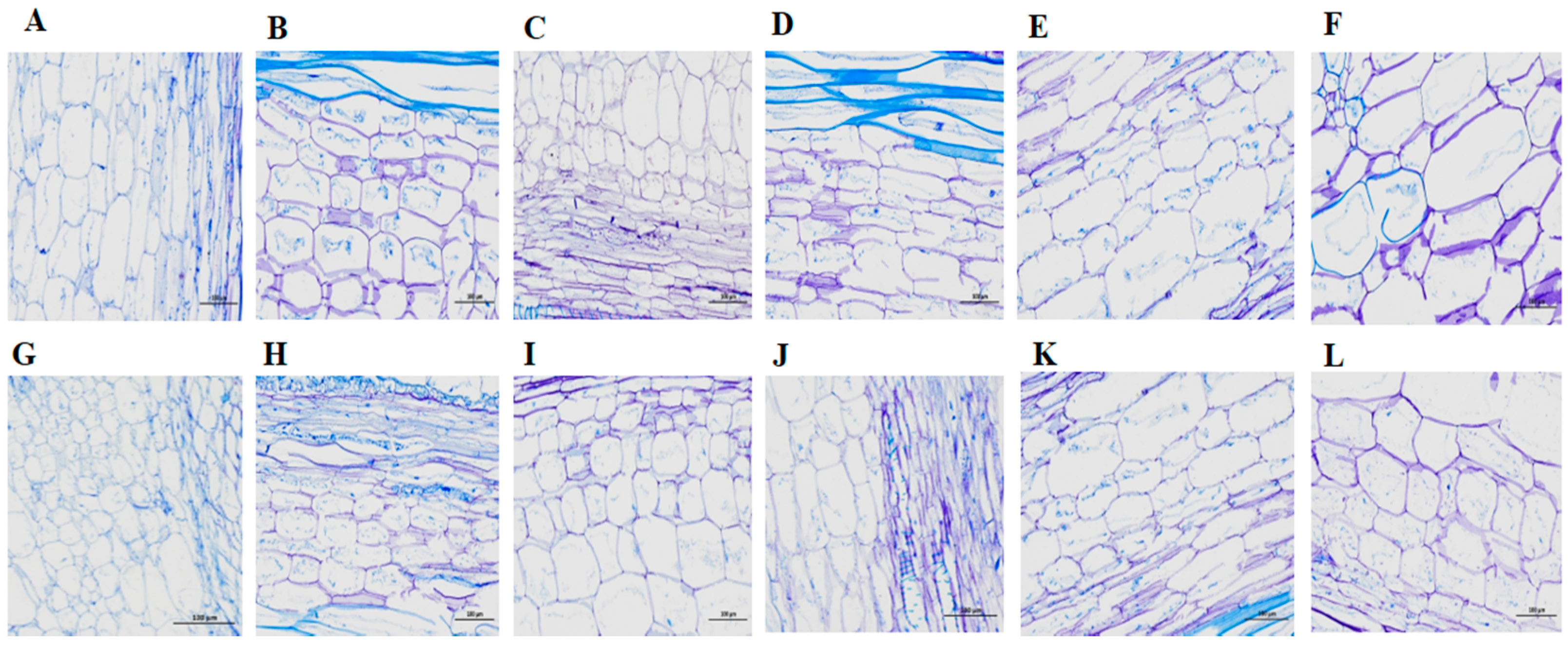
2.7. Microscopic Observation and Verification of the Short Internode
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Mapping Population
4.2. DNA Extraction and Bulked Segregant Analysis
4.3. Fine Mapping through CAPS Markers
4.4. Annotation, Cloning and Sequencing Analysis of Candidate Genes
4.5. Conserved Domain, Phylogenetic Analysis and Verification of the Short Internode Using an InDel Marker
4.6. Expression Analysis of Candidate Gene
4.7. Application of Exogenous GA3 Hormone
4.8. Cytological Analysis of the Short Internodes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dane, F.; Liu, J. Diversity and origin of cultivated and citron type watermelon (Citrullus lanatus). Genet. Resour. Crop Evol. 2006, 54, 1255–1265. [Google Scholar] [CrossRef]
- Levi, A.; Thomas, C.E.; Keinath, A.P.; Wehner, T.C. Genetic diversity among watermelon (Citrullus lanatus and Citrullus colocynthis) accessions. Genet. Resour. Crop Evol. 2001, 48, 559–566. [Google Scholar] [CrossRef]
- Levi, A.; Wechter, P.; Massey, L.; Carter, L.; Hopkins, D. An Extended Genetic Linkage Map for Watermelon Based on a Testcross and a BC2F2 Population. Am. J. Plant. Sci. 2011, 2, 93–110. [Google Scholar] [CrossRef] [Green Version]
- Paris, H.S. Origin and emergence of the sweet dessert watermelon. Citrullus lanatus. Ann. Bot. 2015, 116, 133–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xu, A.; Dong, W.; Li, Z.; Li, G. Genetic Analysis of a Dwarf Vine and Small Fruit Watermelon Mutant. Hortic. Plant J. 2016, 2, 224–228. [Google Scholar] [CrossRef]
- Arumuganathan, K.; Earle, E.D. Nuclear DNA content of some important plant species. Plant Mol. Biol. Rep. 1991, 9, 208–218. [Google Scholar] [CrossRef]
- Lou, L.; Wenhner, T.C. Qualitative Inheritance of External Fruit Traits in Watermelon. HortScience 2016, 51, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Zhao, S.; Sun, H.; Wang, X.; Wu, S.; Lin, T.; Ren, Y.; Gao, L.; Deng, Y.; Zhang, J.; et al. Resequencing of 414 cultivated and wild watermelon accessions identifies selection for fruit quality traits. Nat. Genet. 2019, 51, 1616–1623. [Google Scholar] [CrossRef]
- Khush, G.S. Green revolution: The way forward. Nat. Rev. Genet. 2001, 1, 815–822. [Google Scholar] [CrossRef]
- Salamini, F. Hormones and the Green Revolution. Science 2003, 302, 71–72. [Google Scholar] [CrossRef] [PubMed]
- Kendall, S.L.; Holmes, H.; White, C.A.; Clarke, S.M.; Berry, P.M. Quantifying lodging-induced yield losses in oilseed rape. Field Crops Res. 2017, 211, 106–113. [Google Scholar] [CrossRef]
- Xiao, Z.; Fu, R.; Li, J.; Fan, Z.; Yin, H. Overexpression of the Gibberellin 2-Oxidase Gene from Camellia lipoensis Induces Dwarfism and Smaller Flowers in Nicotiana tabacum. Plant Mol. Biol. Rep. 2015, 34, 182–191. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, W.; Chu, P.; Wan, S.; Yang, M.; Wang, M.; Guan, R. Mapping a major QTL responsible for dwarf architecture in Brassica napus using a single-nucleotide polymorphism marker approach. BMC Plant Biol. 2016, 16, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, T.T.; Tomoaki, S.; Kazuko, O.; Tomonobu, T.; Hidemi, K.; Motoyuki, A.; Shigeyuki, I.; Makoto, M. A rice semi-dwarf gene, Tan-Ginbozu (D35), encodes the gibberellin biosynthesis enzyme, ent-kaurene oxidase. Plant Mol. Biol. 2004, 54, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, W.; Qin, Y.; Pan, Y.; Wang, X.; Weng, Y.; Chen, P.; Li, Y. The Cytochrome P450 Gene CsCYP85A1 Is a Putative Candidate for Super Compact-1 (Scp-1) Plant Architecture Mutation in Cucumber (Cucumis sativus L.). Front. Plant Sci. 2017, 8, 266–278. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Badigannavar, A.M.; D’Souza, S.F. Inheritance and molecular mapping of a gibberellin-insensitive dwarf mutant in groundnut (Arachis hypogaea L.). J. Appl. Genet. 2010, 52, 35–38. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Zhou, J.; Zhang, Y.; Cao, J. Characterization and inheritance of a bush-type in tropical pumpkin (Cucurbita moschata Duchesne). Sci. Hortic. 2007, 114, 1–4. [Google Scholar] [CrossRef]
- Dong, W.; Wu, D.; Li, G.; Wu, D.; Wang, Z. Next-generation sequencing from bulked segregant analysis identifies a dwarfism gene in watermelon. Sci. Rep. 2018, 8, 2908. [Google Scholar] [CrossRef] [Green Version]
- Holst, K.; Schmulling, T.; Werner, T. Enhanced cytokinin degradation in leaf primordia of transgenic Arabidopsis plants reduces leaf size and shoot organ primordia formation. J. Plant Physiol. 2011, 168, 1328–1334. [Google Scholar] [CrossRef]
- Sun, T.P. Gibberellin metabolism, perception and signaling pathways in Arabidopsis. Arab. Book 2008, 6, e0103. [Google Scholar] [CrossRef] [PubMed]
- Fleet, C.M.; Sun, T.P. A DELLAcate balance: The role of gibberellin in plant morphogenesis. Curr. Opin. Plant Biol. 2005, 8, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Spielmeyer, W.; Marc HEa Peter, M.C. Semidwarf (sd-1), green revolution rice, contains a defective gibberellin 20-oxidase gene. Proc. Natl. Acad. Sci. USA 2002, 99, 9043–9048. [Google Scholar] [CrossRef] [Green Version]
- Monna, L.; Kitazawa, N.; Yoshino, R.; Suzuki, J.; Masuda, H.; Maehara, Y.; Tanji, M.; Sato, M.; Nasu, S.; Minobe, Y. Positional Cloning of Rice Semi-dwarfing Gene, sd-1: Rice Green Revolution Gene Encodes a Mutant Enzyme Involved in Gibberellin Synthesis. DNA Res. 2002, 9, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, B.A.; Foo, E.; Sharwood, R.; Karafiatova, M.; Vrana, J.; MacMillan, C.; Nichols, D.S.; Steuernagel, B.; Uauy, C.; Dolezel, J.; et al. Rht18 Semidwarfism in Wheat Is Due to Increased GA 2-oxidaseA9 Expression and Reduced GA Content. Plant Physiol. 2018, 177, 168–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverstone, A.A.; Sun, T.P. Gibberellins and the Green Revolution. Trends Plant Sci. 2000, 5, 1–2. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, S.; Gui, J.; Fu, C.; Yu, H.; Song, D.; Shen, J.; Qin, P.; Liu, X.; Han, B.; et al. Shortened Basal Internodes Encodes a Gibberellin 2-Oxidase and Contributes to Lodging Resistance in Rice. Mol. Plant 2018, 11, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Sun, D.; Guan, W.; Sun, G.; Li, C. Inheritance and identification of molecular markers associated with a novel dwarfing gene in barley. BMC Genet. 2010, 11, 1471–2156. [Google Scholar] [CrossRef] [Green Version]
- Hilley, J.; Truong, S.; Olson, S.; Morishige, D.; Mullet, J. Identification of Dw1, a Regulator of Sorghum Stem Internode Length. PLoS ONE 2016, 11, e0151271. [Google Scholar] [CrossRef] [Green Version]
- Azhaguvel, P. Mapping the d1 and d2 Dwarfing Genes and the Purple Foliage Color Locus P in Pearl Millet. J. Hered. 2003, 94, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Donald, N.M.; Gary, W.E.; Stephen, T.T.A.; Carle, R.B. ‘El Dorado’ and ‘La Estrella’: Compact Plant Tropical Pumpkin Hybrids. HortScience 2002, 37, 831–833. [Google Scholar]
- Wu, T.; Cao, J. Differential gene expression of tropical pumpkin (Cucurbita moschata Duchesne) bush mutant during internode development. Sci. Hortic. 2008, 117, 219–224. [Google Scholar] [CrossRef]
- Li, Y.; Yang, L.; Pathak, M.; Li, D.; He, X.; Weng, Y. Fine genetic mapping of cp: A recessive gene for compact (dwarf) plant architecture in cucumber, Cucumis sativus L. Theor. Appl. Genet. 2011, 123, 973–983. [Google Scholar] [CrossRef]
- Lin, T.; Wang, S.; Zhong, Y.; Gao, D.; Cui, Q.; Chen, H.; Zhang, Z.; Shen, H.; Weng, Y.; Huang, S. A Truncated F-Box Protein Confers the Dwarfism in Cucumber. J. Genet. Genom. 2016, 43, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Fukino, N.; Ohara, T.; Sugiyama, M.; Kubo, N.; Hirai, M.; Sakata, Y.; Matsumoto, S. Mapping of a gene that confers short lateral branching (slb) in melon (Cucumis melo L.). Euphytica 2012, 187, 133–143. [Google Scholar] [CrossRef]
- Hwang, J.; Oh, J.; Kim, Z.; Staub, J.E.; Chung, S.-M.; Park, Y. Fine genetic mapping of a locus controlling short internode length in melon (Cucumis melo L.). Mol. Breed. 2014, 34, 949–961. [Google Scholar] [CrossRef]
- Paris, H.S.; Brown, R.N. The Genes of Pumpkin and Squash. HortScience 2005, 40, 1620–1630. [Google Scholar] [CrossRef] [Green Version]
- Itoh, H.; Ueguchi-Tanaka, M.; Sentoku, N.; Kitano, H.; Matsuoka Ma Kobayashi, M. Cloning and functional analysis of two gibberellin 3b-hydroxylase genes that are differently expressed during the growth of rice. Proc. Natl. Acad. Sci. USA 1999, 98, 8909–8914. [Google Scholar] [CrossRef] [Green Version]
- Tien, D.N.; Oo, M.M.; Soh, M.-S.; Park, S.K. Bioengineering of Male Sterility in Rice (Oryza sativa L.). Plant Breed. Biotechnol. 2013, 1, 218–235. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.G.; Phillips, A.L.; Hedden, P. Molecular cloning and functional expression of gibberellin 2-oxidases, multifunctional enzymes involved in gibberellin deactivation. Proc. Natl. Acad. Sci. USA 1999, 96, 4698–4704. [Google Scholar] [CrossRef] [Green Version]
- Hedden Pa Philips, A.L. Gibberellin metabolism: New insights revealed by the genes. Trends Plant Sci. 2000, 5, 553–563. [Google Scholar]
- Spielmeyer, W.; Ellis, M.; Robertson, M.; Ali, S.; Lenton, J.R.; Chandler, P.M. Isolation of gibberellin metabolic pathway genes from barley and comparative mapping in barley, wheat and rice. Theor. Appl. Genet. 2004, 109, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Kiyota, S.; Hanada, A.; Yamaguchi, S.; Takano, M. The multiple contributions of phytochromes to the control of internode elongation in rice. Plant Physiol. 2011, 157, 1187–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.; Andy, L.P.; Paul, G.; Peter, H. Function and Substrate Specificity of the Gibberellin 3b-Hydroxylase Encoded by the Arabidopsis GA4 Gene. Plant Physiol. 1998, 117, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Lange, T.; Robatzek, S.; Frisse, A. Cloning and expression of a gibberellin 2 beta,3 beta-hydroxylase cDNA from pumpkin endosperm. Plant Cell 1997, 9, 1459–1467. [Google Scholar] [CrossRef]
- Spray, C.R.; Masatomo, K.; Yoshihito, S.; Bernard, P.; Paul Ga Jake, M. The dwarf-i (dl) mutant of Zea mays blocks three steps in the gibberellin-biosynthetic pathway. Proc. Natl. Acad. Sci. USA 1996, 99, 10515–10518. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Liu, X.; Shen, D.; Miao, H.; Xie, B.; Li, X.; Zeng, P.; Wang, S.; Shang, Y.; Gu, X.; et al. A genomic variation map provides insights into the genetic basis of cucumber domestication and diversity. Nat. Genet. 2013, 45, 1510–1515. [Google Scholar] [CrossRef]
- Ramirez-Gonzalez, R.H.; Segovia, V.; Bird, N.; Fenwick, P.; Holdgate, S.; Berry, S.; Jack, P.; Caccamo, M.; Uauy, C. RNA-Seq bulked segregant analysis enables the identification of high-resolution genetic markers for breeding in hexaploid wheat. Plant Biotechnol. J. 2015, 13, 613–624. [Google Scholar] [CrossRef]
- Cheng, W.; Gao, J.-S.; Feng, X.-X.; Shao, Q.; Yang, S.-X.; Feng, X.-Z. Characterization of dwarf mutants and molecular mapping of a dwarf locus in soybean. J. Integr. Agric. 2016, 15, 2228–2236. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.C.; Obholzer, N.D.; Shah, A.N.; Megason, S.G.; Moens, C.B. RNA-seq-based mapping and candidate identification of mutations from forward genetic screens. Genome Res. 2013, 23, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Song, Q.; Cregan, P.B.; Nelson, R.L.; Wang, X.; Wu, J.; Jiang, G.L. Genome-wide association study for flowering time, maturity dates and plant height in early maturing soybean (Glycine max) germplasm. BMC Genom. 2015, 16, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, C.; Wang, P.; Xu, Y. Bulked sample analysis in genetics, genomics and crop improvement. Plant Biotechnol. J. 2016, 14, 1941–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Mena, C.; de Folter, S.; Costa, M.M.; Angenent, G.C.; Sablowski, R. Transcriptional program controlled by the floral homeotic gene AGAMOUS during early organogenesis. Development 2005, 132, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellmer, F.; Alves-Ferreira, M.; Dubois, A.; Riechmann, J.L.; Meyerowitz, E.M. Genome-wide analysis of gene expression during early Arabidopsis flower development. PLoS Genet. 2006, 2, 1012–1024. [Google Scholar] [CrossRef] [Green Version]
- Russo, C.A.D.M.; Selvatti, A.P.; Tamura, K. Bootstrap and Rogue Identification Tests for Phylogenetic Analyses. Mol. Biol. Evol. 2018, 35, 2327–2333. [Google Scholar] [CrossRef]
- Li, Z.F.; Guo, Y.; Ou, L.; Hong, H.; Wang, J.; Liu, Z.X.; Guo, B.; Zhang, L.; Qiu, L. Identification of the dwarf gene GmDW1 in soybean (Glycine max L.) by combining mapping-by-sequencing and linkage analysis. Theor. Appl. Genet. 2018, 131, 1001–1016. [Google Scholar] [CrossRef] [Green Version]
- Bidinger, F.R.; Raju, D.S. Effects of the d2 dwarfing gene in pearl millet. Theor. Appl. Genet. 1990, 79, 521–524. [Google Scholar] [CrossRef]
- Gillmor, C.S.; Roeder, A.H.; Sieber, P.; Somerville, C.; Lukowitz, W. A Genetic Screen for Mutations Affecting Cell Division in the Arabidopsis thaliana Embryo Identifies Seven Loci Required for Cytokinesis. PLoS ONE 2016, 11, e0146492. [Google Scholar] [CrossRef] [Green Version]
- Klein, H.; Xiao, Y.; Conklin, P.A.; Govindarajulu, R.; Kelly, J.A.; Scanlon, M.J.; Whipple, C.J.; Bartlett, M. Bulked-Segregant Analysis Coupled to Whole Genome Sequencing (BSA-Seq) for Rapid Gene Cloning in Maize. G3 Genes Genet. Genom. 2018, 8, 3583–3592. [Google Scholar] [CrossRef] [Green Version]
- Nawy, T.; Bayer, M.; Mravec, J.; Friml, J.; Birnbaum, K.D.; Lukowitz, W. The GATA factor HANABA TARANU is required to position the proembryo boundary in the early Arabidopsis embryo. Dev. Cell 2010, 19, 103–113. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, X.; Wei, Z.; Li, Q.; Li, X. Inheritance of male-sterility and dwarfism inwatermelon Citrullus lanatus (Thunb.) Matsum. and Nakaix. Sci. Hortic. 1998, 74, 175–181. [Google Scholar]
- Hou, S.; Niu, H.; Tao, Q.; Wang, S.; Gong, Z.; Li, S.; Weng, Y.; Li, Z. A mutant in the CsDET2 gene leads to a systemic brassinosteriod deficiency and super compact phenotype in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2017, 130, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, Y.; Zhang, W.H. Brassinosteroids are involved in response of cucumber (Cucumis sativus) to iron deficiency. Ann. Bot. 2012, 110, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, M.H.; Bonnett, D.G.; Rebetzke, G.J. A 192bp allele at the Xgwm261 locus is not always associated with the Rht8 dwarfing gene in wheat (Triticum aestivum L.). Euphytica 2007, 157, 209–214. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Bonnett, D.J.A.; Ellis, M.H. Combining gibberellic acid-sensitive and insensitive dwarfing genes in breeding of higher-yielding, sesqui-dwarf wheats. Field Crops Res. 2012, 127, 17–25. [Google Scholar] [CrossRef]
- Wang, Y.; Du, Y.; Yang, Z.; Chen, L.; Condon, A.G.; Hu, Y.-G. Comparing the effects of GA-responsive dwarfing genes Rht13 and Rht8 on plant height and some agronomic traits in common wheat. Field Crops Res. 2015, 179, 35–43. [Google Scholar] [CrossRef]
- Na, T.; Jiang, Y.; He, B.R.; Hu, Y.G. The Effects of Dwarfing Genes (Rht-B1b, Rht-D1b, and Rht8) with Different Sensitivity to GA3 on the Coleoptile Length and Plant Height of Wheat. Agric. Sci. China 2009, 8, 1028–1038. [Google Scholar] [CrossRef]
- Chen, L.; Phillips, A.L.; Condon, A.G.; Parry, M.A.; Hu, Y.G. GA-responsive dwarfing gene Rht12 affects the developmental and agronomic traits in common bread wheat. PLoS ONE 2013, 8, e62285. [Google Scholar] [CrossRef] [Green Version]
- Barratt, N.M.; Peter, J.D. Developmental changes in the gibberellin-induced growth response in stem segments of light-grown pea genotypes. Plant Growth Regul. 1997, 21, 127–134. [Google Scholar] [CrossRef]
- Chen, L.; Hao, L.; Condon, A.G.; Hu, Y.G. Exogenous GA3 application can compensate the morphogenetic effects of the GA-responsive dwarfing gene Rht12 in bread wheat. PLoS ONE 2014, 9, e86431. [Google Scholar] [CrossRef] [Green Version]
- Magome, H.; Shinjiro, S.; Atsushi, H.; Yuji, K.; Kenji, O. Dwarf and delayed flowering 1, a novel Arabidopsis mutant deficient in Gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J. 2004, 37, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-P.; Ali, S.A.; Xiao, G.; Chen, F.-J.; Yuan, L.-X.; Gu, R.-L. Phenotypic characterization and genetic mapping of the dwarf mutant m34 in maize. J. Integr. Agric. 2019, 18, 948–957. [Google Scholar] [CrossRef]
- Noguchi, H. Effects of red light on gibberellin 3β-hydroxylase gene expression in dark-grown seedlings of dwarf and tall cultivars of Pisum sativum. J. Plant Physiol. 2002, 159, 607–612. [Google Scholar] [CrossRef]
- Sun, X.; Shu, J.; Ali Mohamed, A.M.; Deng, X.; Zhi, X.; Bai, J.; Cui, Y.; Lu, X.; Du, Y.; Wang, X.; et al. Identification and Characterization of EI (Elongated Internode) Gene in Tomato (Solanum lycopersicum). Int. J. Mol. Sci. 2019, 20, 2204. [Google Scholar] [CrossRef] [Green Version]
- Ayano, M.; Kani, T.; Kojima, M.; Sakakibara, H.; Kitaoka, T.; Kuroha, T.; Angeles-Shim, R.B.; Kitano, H.; Nagai, K.; Ashikari, M. Gibberellin biosynthesis and signal transduction is essential for internode elongation in deepwater rice. Plant Cell Environ. 2014, 37, 2313–2324. [Google Scholar] [CrossRef]
- Liang, Y.C.; Reid, M.S.; Jiang, C.Z. Controlling plant architecture by manipulation of gibberellic acid signalling in petunia. Hortic. Res. 2014, 1, 14061. [Google Scholar] [CrossRef] [Green Version]
- Takatsuka, H.; Umeda, M. Hormonal control of cell division and elongation along differentiation trajectories in roots. J. Exp. Bot. 2014, 65, 2633–2643. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Xing, Y.; Lou, Q.; Feng, P.; Liu, S.; Zhu, M.; Yin, W.; Fang, S.; Lin, Y.; Zhang, T.; et al. Dwarf and short grain 1, encoding a putative U-box protein regulates cell division and elongation in rice. J. Plant Physiol. 2017, 209, 84–94. [Google Scholar] [CrossRef]
- Zhang, S.; Hu, W.; Wang, L.; Lin, C.; Cong, B.; Sun, C.; Luo, D. TFL1/CEN-like genes control intercalary meristem activity and phase transition in rice. Plant Sci. 2005, 168, 1393–1408. [Google Scholar] [CrossRef]
- Wang, L.; Mu, C.; Du, M.; Chen, Y.; Tian, X.; Zhang, M.; Li, Z. The effect of mepiquat chloride on elongation of cotton (Gossypium hirsutum L.) internode is associated with low concentration of gibberellic acid. Plant Sci. 2014, 225, 15–23. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, J.; Lu, W.; Deng, D. Gibberellin in plant height control: Old player, new story. Plant Cell Rep. 2017, 36, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.K.; Roddic, J.G. Effect of steroidal responses and gibberellic acid on stem elongation on tall and dwarf cultivars of Pisum sativum. New Phytol. 1977, 79, 493–499. [Google Scholar] [CrossRef]
- Ueguchi-Tanaka, M.; Ashikari, M.; Nakajima, M.; Itoh, H.; Katoh, E.; Kobayashi, M.; Chow, T.Y.; Hsing, Y.I.; Kitano, H.; Yamaguchi, I.; et al. GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor for gibberellin. Nature 2005, 437, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, L.; Wang, X.; Liang, L.; Yue, J.; Li, L. Comparative Analyses of Anatomical Structure, Phytohormone Levels, and Gene Expression Profiles Reveal Potential Dwarfing Mechanisms in Shengyin Bamboo (Phyllostachys edulis f. tubaeformis). Int. J. Mol. Sci. 2018, 19, 1697. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.; Xiao, Y.; Liu, D.; Gao, S.; Liu, L.; Yin, Y.; Jin, Y.; Qian, Q.; Chu, C. Brassinosteroid regulates cell elongation by modulating gibberellin metabolism in rice. Plant Cell 2014, 26, 4376–4393. [Google Scholar] [CrossRef] [Green Version]
- Luan, F.; Delannay, I.; Staub, J.E. Chinese melon (Cucumis melo L.) diversity analyses provide strategies for germplasm curation, genetic improvement, and evidentiary support of domestication patterns. Euphytica 2008, 164, 445–461. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. Genome Project Data Processing S: The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Mitsuoka, C.; Tamiru, M.; et al. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 2012, 30, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S.; et al. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef]
- Das, S.; Upadhyaya, H.D.; Bajaj, D.; Kujur, A.; Badoni, S.; Laxmi Kumar, V.; Tripathi, S.; Gowda, C.L.; Sharma, S.; Singh, S.; et al. Deploying QTL-seq for rapid delineation of a potential candidate gene underlying major trait-associated QTL in chickpea. DNA Res. 2015, 22, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Khan, A.W.; Saxena, R.K.; Kumar, V.; Kale, S.M.; Sinha, P.; Chitikineni, A.; Pazhamala, L.T.; Garg, V.; Sharma, M.; et al. Next-generation sequencing for identification of candidate genes for Fusarium wilt and sterility mosaic disease in pigeonpea (Cajanus cajan). Plant Biotechnol. J. 2016, 14, 1183–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebremeskel, H.; Dou, J.; Li, B.; Zhao, S.; Muhammad, U.; Lu, X.; He, N.; Liu, W. Molecular Mapping and Candidate Gene Analysis for GA3 Responsive Short Internode in Watermelon (Citrullus lanatus). Int. J. Mol. Sci. 2020, 21, 290. https://doi.org/10.3390/ijms21010290
Gebremeskel H, Dou J, Li B, Zhao S, Muhammad U, Lu X, He N, Liu W. Molecular Mapping and Candidate Gene Analysis for GA3 Responsive Short Internode in Watermelon (Citrullus lanatus). International Journal of Molecular Sciences. 2020; 21(1):290. https://doi.org/10.3390/ijms21010290
Chicago/Turabian StyleGebremeskel, Haileslassie, Junling Dou, Bingbing Li, Shengjie Zhao, Umer Muhammad, Xuqiang Lu, Nan He, and Wenge Liu. 2020. "Molecular Mapping and Candidate Gene Analysis for GA3 Responsive Short Internode in Watermelon (Citrullus lanatus)" International Journal of Molecular Sciences 21, no. 1: 290. https://doi.org/10.3390/ijms21010290