RNA-Binding Proteins HuB, HuC, and HuD are Distinctly Regulated in Dorsal Root Ganglia Neurons from STZ-Sensitive Compared to STZ-Resistant Diabetic Mice

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
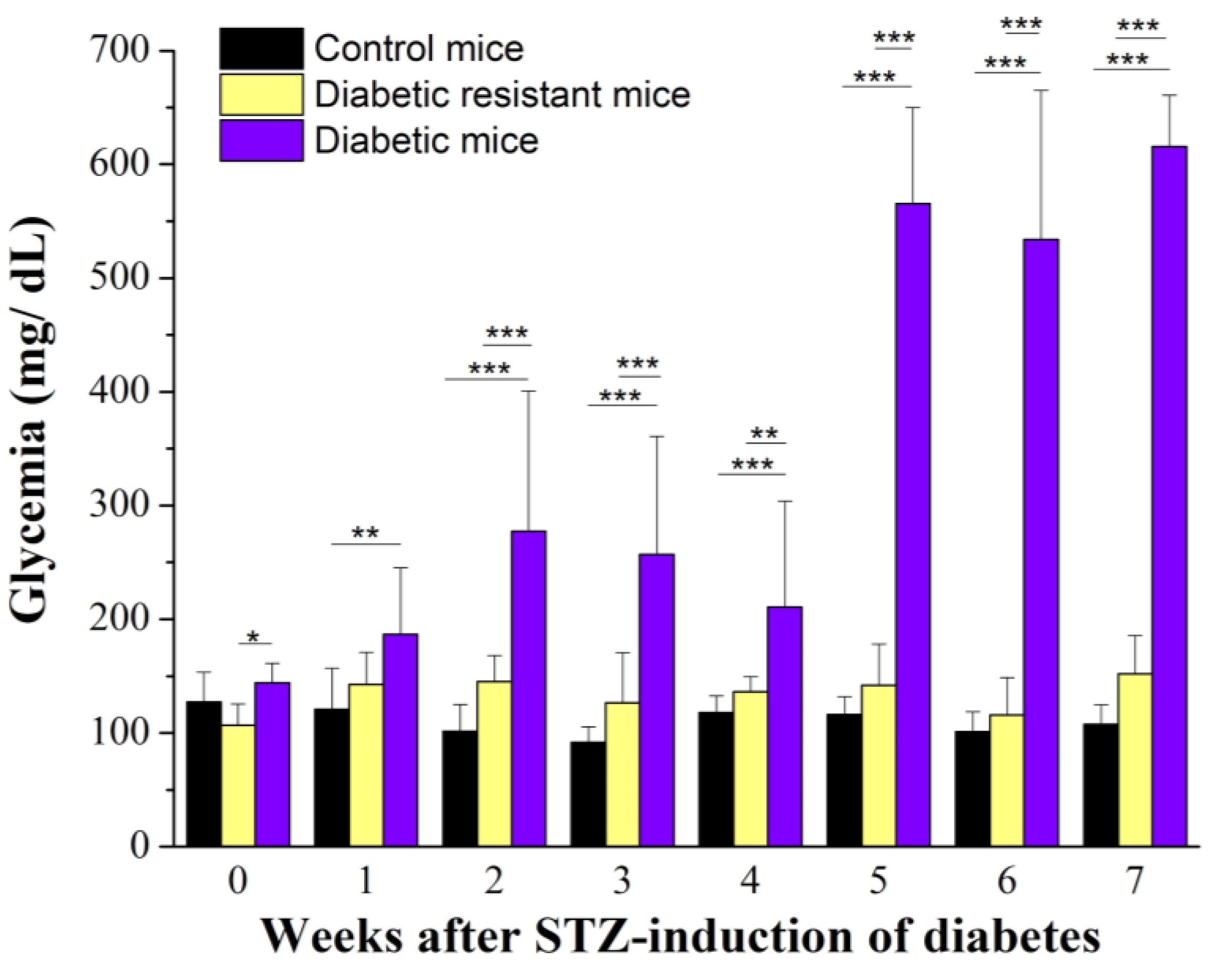
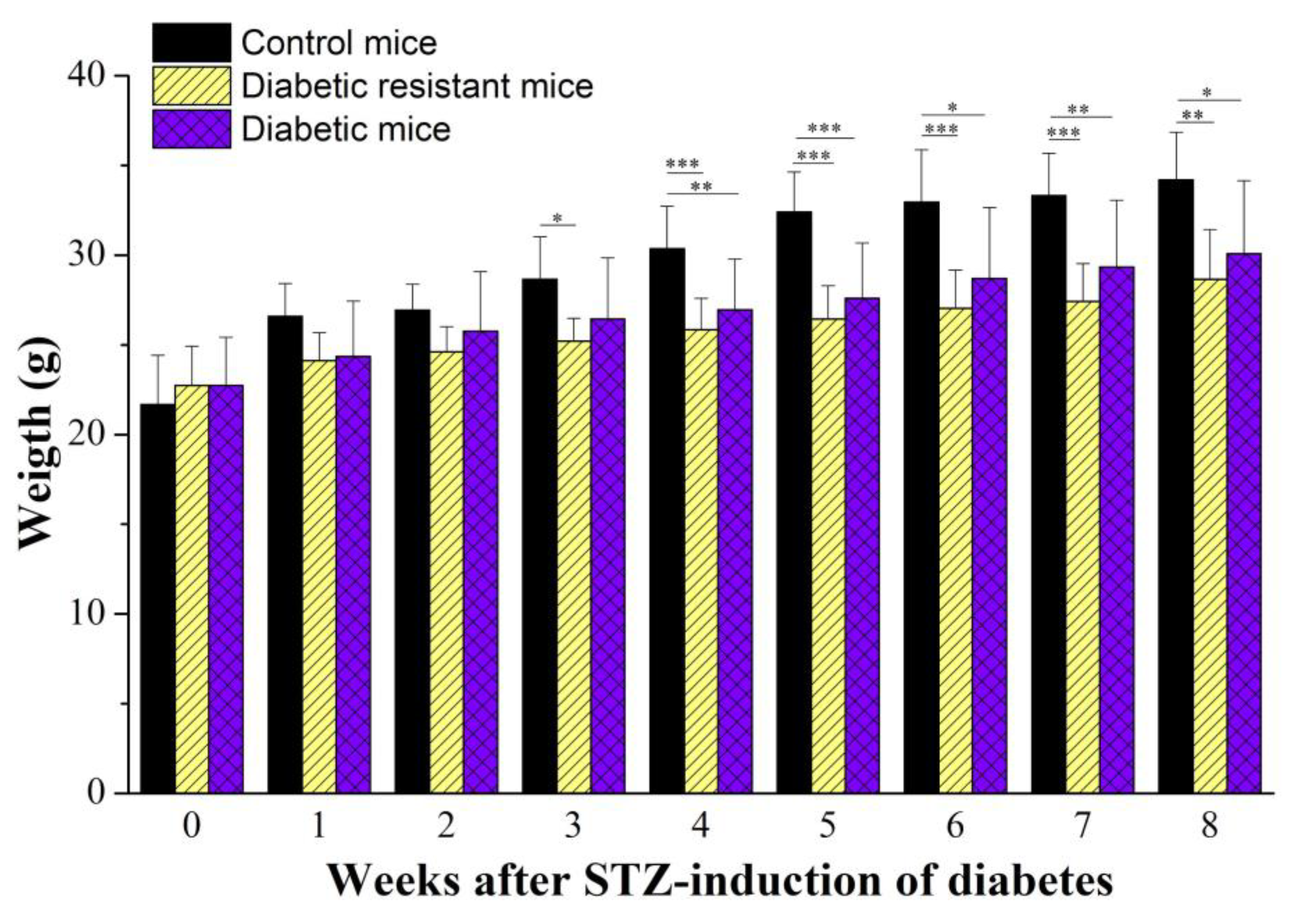
2.1. Diabetic Mice Have Changes in Glycemia and Body Weight Compared to Diabetic Resistant or Control Mice
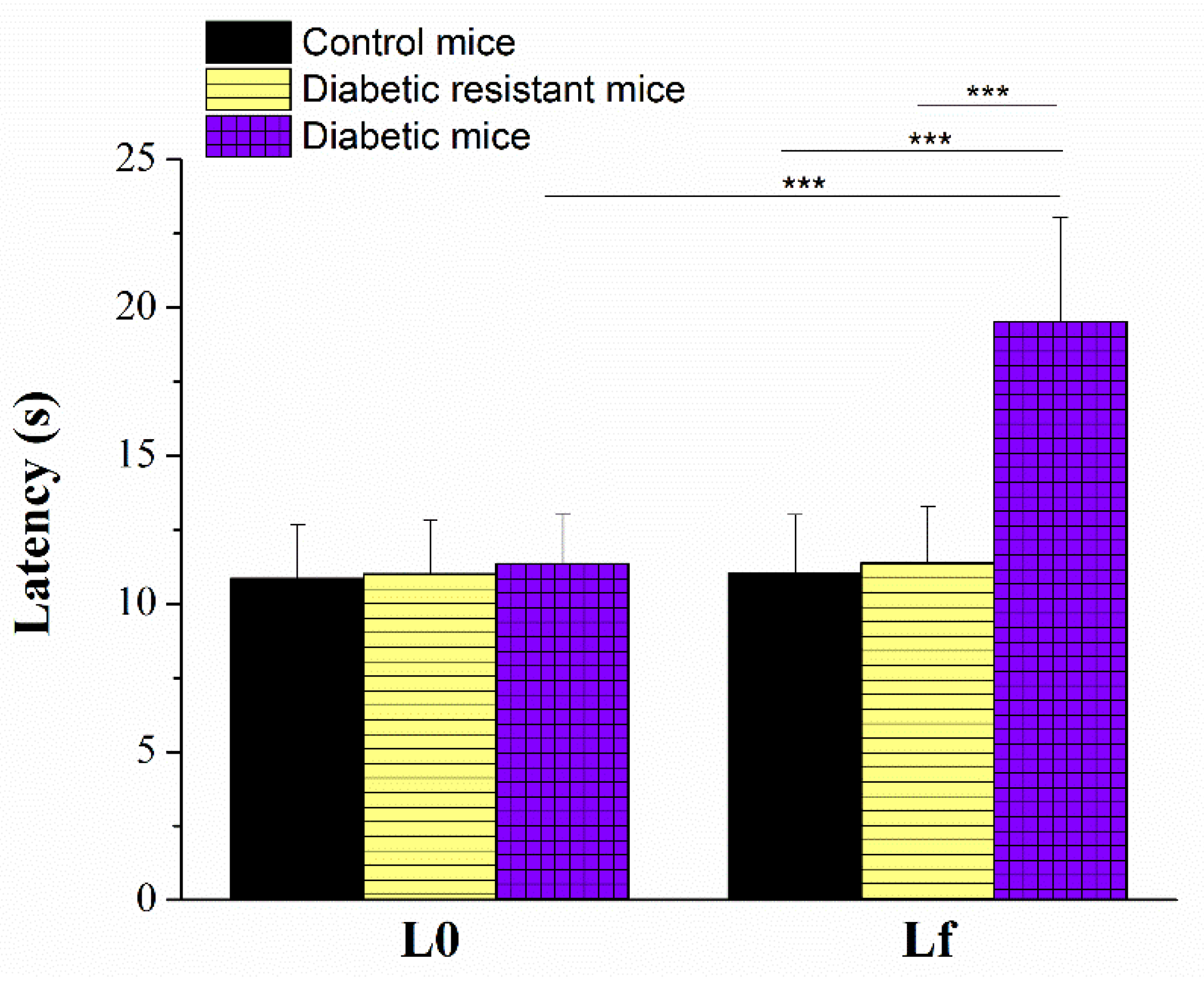
2.2. Diabetic Mice Have Longer Paw Withdrawal Latencies to Nociceptive Thermal Stimulation than Diabetic Resistant or Control Mice
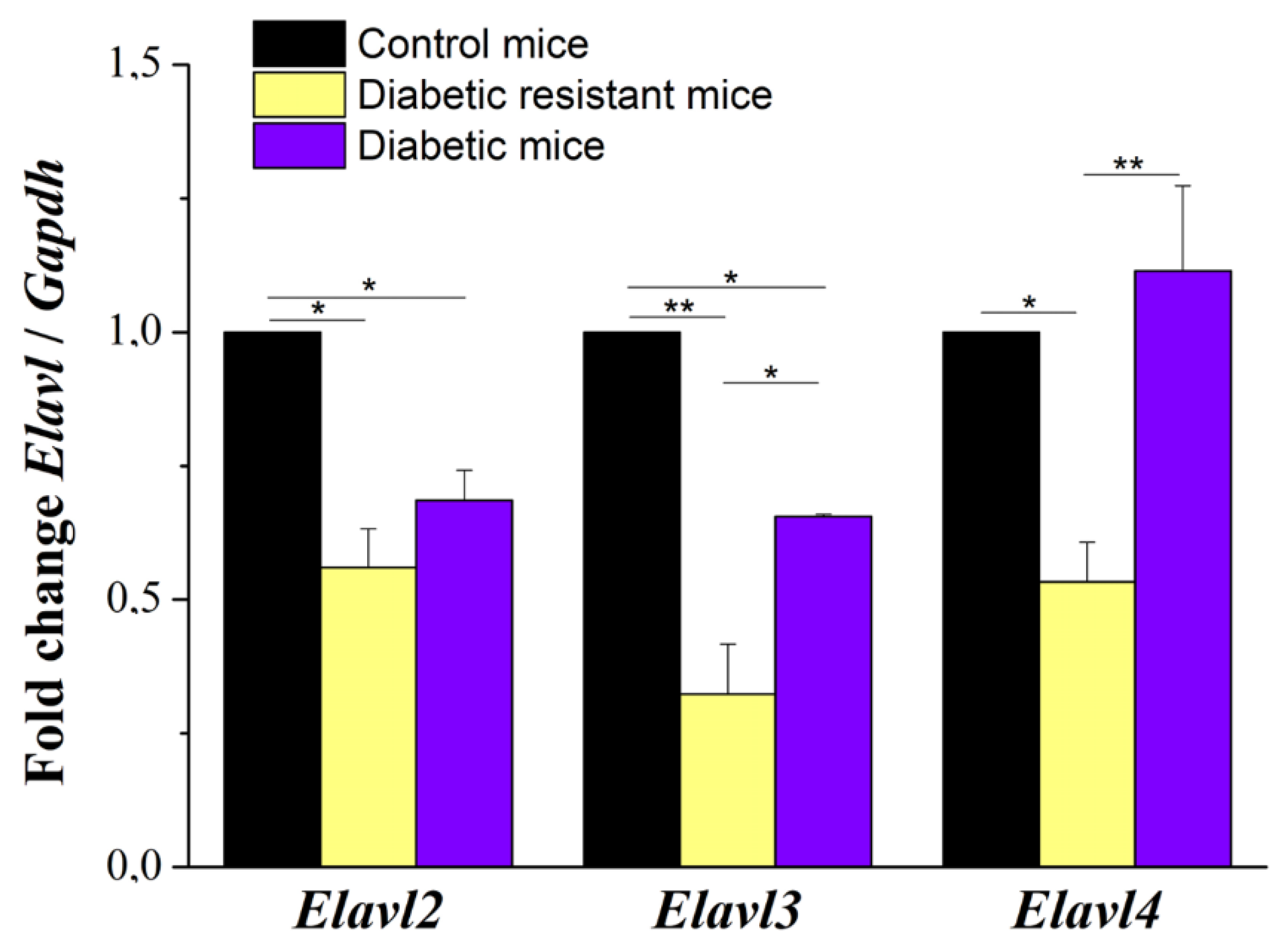
2.3. Elav-Like Gene Expression in Mouse DRG Neurons Is Decreased in Diabetic and Diabetic Resistant Mice Compared to Control Mice
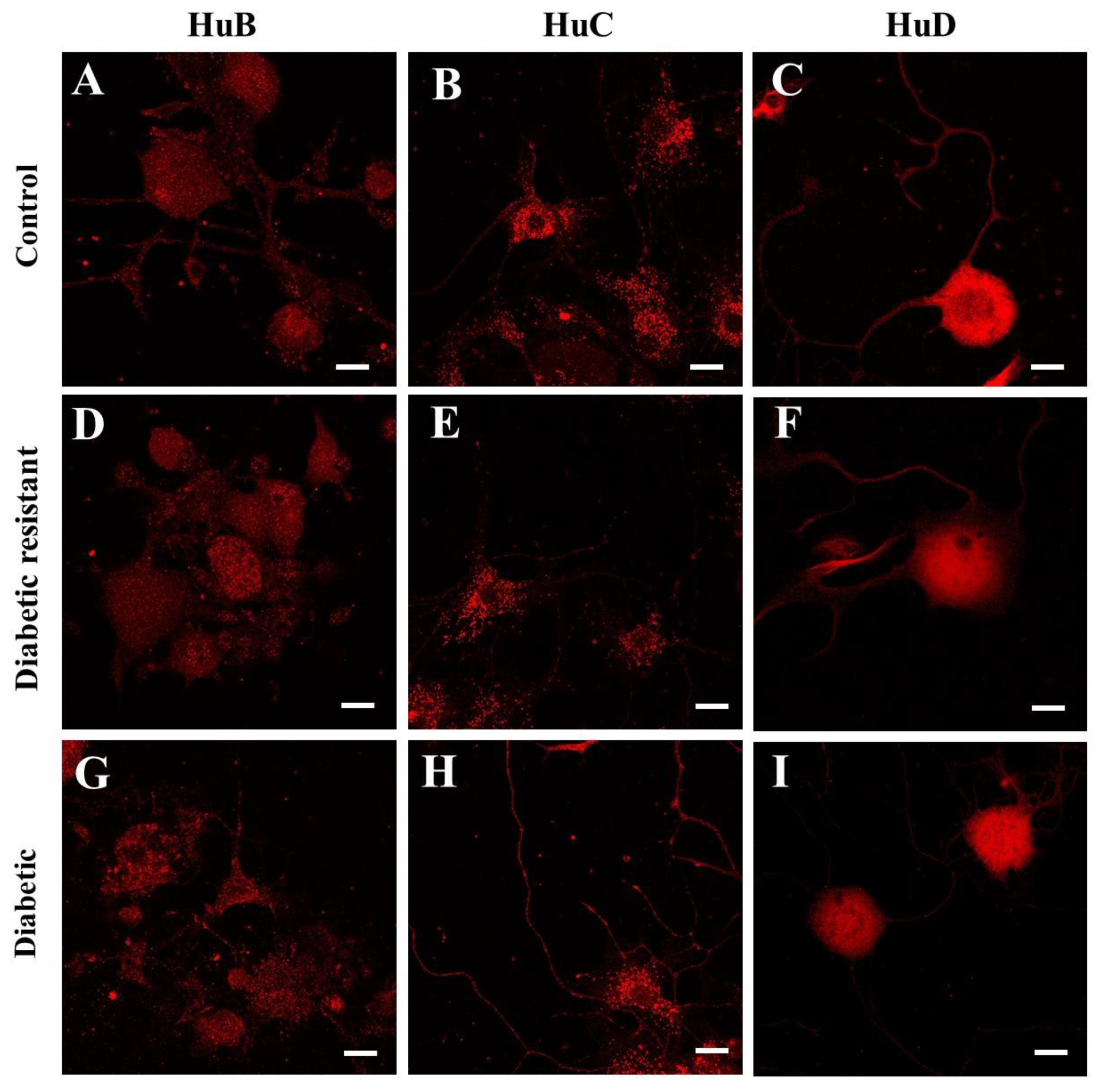
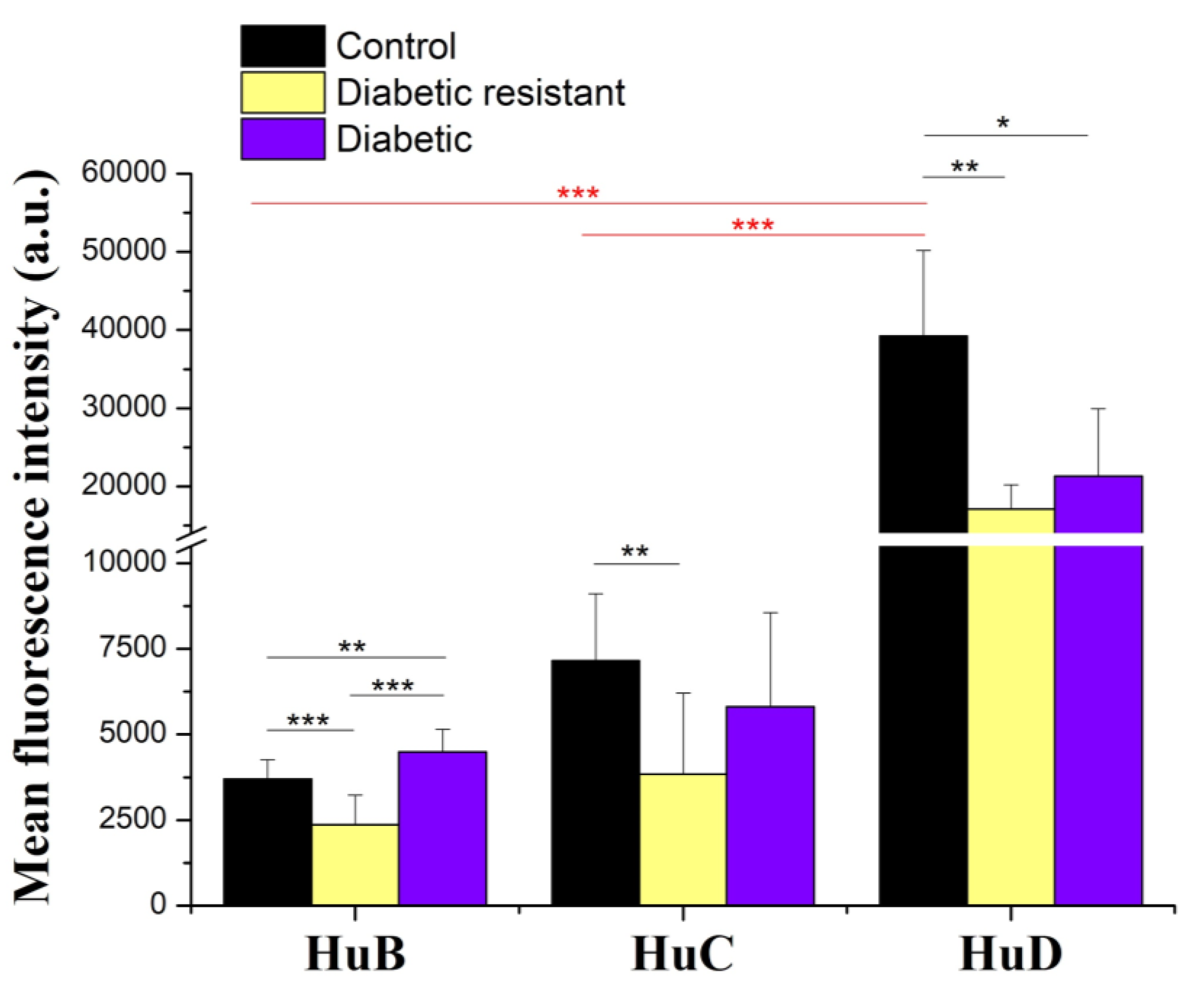
2.4. Hu Proteins Expression in Mouse DRG Neurons Is Decreased in Diabetic and Diabetic Resistant Mice Compared to Control Mice
3. Discussion
4. Materials and Methods
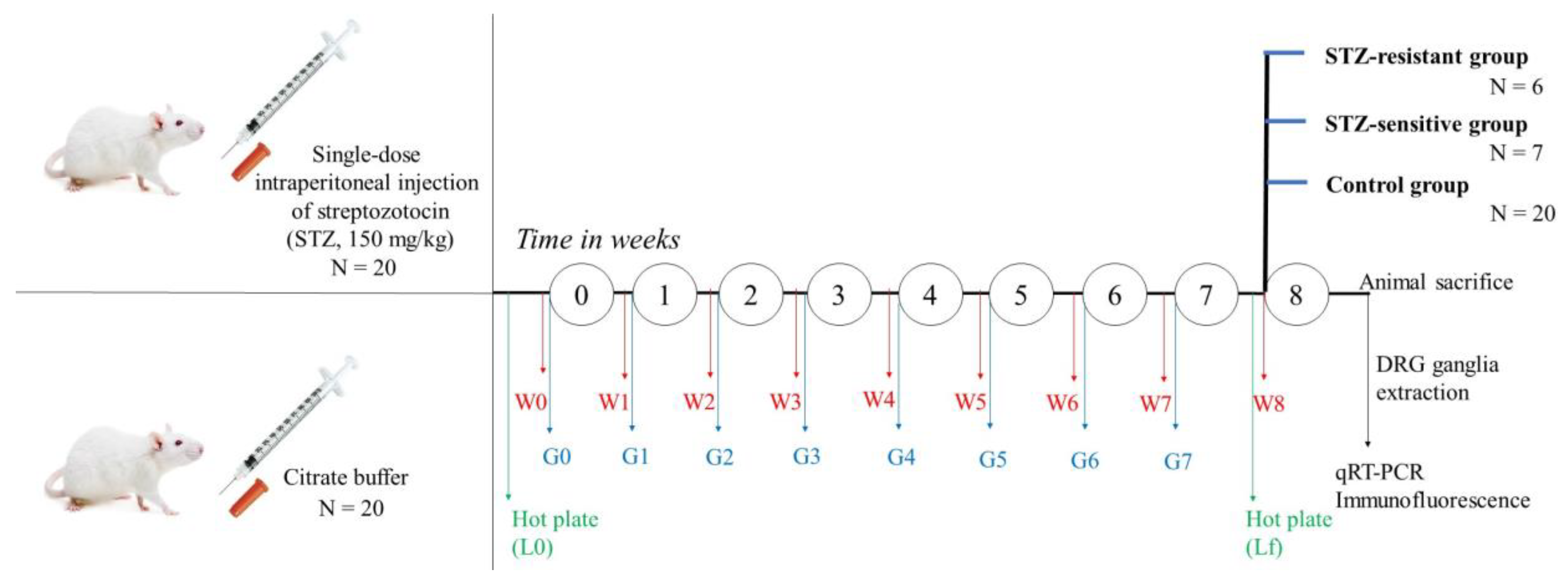
4.1. Animals
4.2. Streptozotocin-Induced Diabetes
4.3. Body Weight Measurements
4.4. Glycemia Measurements
4.5. Hot-Plate Test
4.6. Primary Cultures of Neurons from Dorsal Root Ganglia
4.7. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
4.8. Immunofluorescence
4.9. Digital Image Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Glisovic, T.; Bachorik, J.L.; Yong, J.; Dreyfuss, G. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 2008, 582, 1977–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.J.; Irizarry, K. Alternative splicing in the nervous system: An emerging source of diversity and regulation. Biol. Psychiatry. 2003, 54, 771–776. [Google Scholar] [CrossRef]
- Lipscombe, D. Neuronal proteins custom designed by alternative splicing. Curr. Opin. Neurobiol. 2005, 15, 358–363. [Google Scholar] [CrossRef]
- Sutton, M.A.; Schuman, E.M. Local translational control in dendrites and its role in long-term synaptic plasticity. J. Neurobiol. 2005, 64, 116–131. [Google Scholar] [CrossRef] [Green Version]
- Hengst, U.; Jaffrey, S.R. Function and translational regulation of mRNA in developing axons. Semin. Cell. Dev. Biol. 2007, 18, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Hinman, M.N.; Lou, H. Diverse molecular functions of Hu proteins. Cell. Mol. Life Sci. 2008, 65, 3168–3181. [Google Scholar] [CrossRef] [Green Version]
- Perrone-Bizzozero, N.; Bird, C.W. Role of HuD in nervous system function and pathology. Front. Biosci. (Schol. Ed.). 2013, 5, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.D.; Merhege, M.A.; Morin, M.; Bolognani, F.; Perrone-Bizzozero, N.I. Increased expression and localization of the RNA-binding protein HuD and GAP-43 mRNA to cytoplasmic granules in DRG neurons during nerve regeneration. Exp. Neurol. 2003, 183, 100–108. [Google Scholar] [CrossRef]
- Bolognani, F.; Tanner, D.C.; Merhege, M.; Deschênes-Furry, J.; Jasmin, B.; Perrone-Bizzozero, N.I. In vivo post-transcriptional regulation of GAP-43 mRNA by overexpression of the RNA-binding protein HuD. J. Neurochem. 2006, 96, 790–801. [Google Scholar] [CrossRef] [Green Version]
- King, P.H. RNA-binding analyses of HuC and HuD with the VEGF and c-myc 3′-untranslated regions using a novel ELISA-based assay. Nucleic Acids Res. 2000, 28, E20. [Google Scholar] [CrossRef]
- Lim, C.S.; Alkon, D.L. PKCε promotes HuD-mediated neprilysin mRNA stability and enhances neprilysin-induced Aβ degradation in brain neurons. PLoS ONE 2014, 9, e97756. [Google Scholar] [CrossRef]
- Borgeson, C.D.; Samson, M.L. Shared RNA-binding sites for interacting members of the Drosophila ELAV family of neuronal proteins. Nucleic Acids Res. 2005, 33, 6372–6383. [Google Scholar] [CrossRef] [Green Version]
- Ratti, A.; Fallini, C.; Cova, L.; Fantozzi, R.; Calzarossa, C.; Zennaro, E.; Pascale, A.; Quattrone, A.; Silani, V. A role for the ELAV RNA-binding proteins in neural stem cells: Stabilization of Msi1 mRNA. J. Cell Sci. 2006, 119, 1442–1452. [Google Scholar] [CrossRef]
- Ratti, A.; Fallini, C.; Colombrita, C.; Pascale, A.; Laforenza, U.; Quattrone, A.; Silani, V. Post-transcriptional regulation of neuro-oncological ventral antigen 1 by the neuronal RNA-binding proteins ELAV. J. Biol. Chem. 2008, 283, 7531–7541. [Google Scholar] [CrossRef]
- Ince-Dunn, G.; Okano, H.J.; Jensen, K.B.; Park, W.Y.; Zhong, R.; Ule, J.; Mele, A.; Fak, J.J.; Yang, C.; Zhang, C.; et al. Neuronal Elav-like (Hu) proteins regulate RNA splicing and abundance to control glutamate levels and neuronal excitability. Neuron. 2012, 75, 1067–1080. [Google Scholar] [CrossRef] [Green Version]
- Sosanya, N.M.; Huang, P.P.; Cacheaux, L.P.; Chen, C.J.; Nguyen, K.; Perrone-Bizzozero, N.I.; Raab-Graham, K.F. Degradation of high affinity HuD targets releases Kv1.1 mRNA from miR-129 repression by mTORC1. J. Cell Biol. 2013, 202, 53–69. [Google Scholar] [CrossRef] [Green Version]
- Fornaro, M.; Geuna, S. Confocal imaging of HuC/D RNA-binding proteins in adult rat primary sensory neurons. Ann. Anat. 2001, 183, 471–473. [Google Scholar] [CrossRef]
- Fornaro, M.; Raimondo, S.; Lee, J.M.; Giacobini-Robecchi, M.G. Neuron-specific Hu proteins sub-cellular localization in primary sensory neurons. Ann Anat. 2007, 189, 223–228. [Google Scholar] [CrossRef]
- Sanna, M.D.; Quattrone, A.; Mello, T.; Ghelardini, C.; Galeotti, N. The RNA-binding protein HuD promotes spinal GAP43 overexpression in antiretroviral-induced neuropathy. Exp. Neurol. 2014, 261, 343–353. [Google Scholar] [CrossRef]
- Gomes, C.; Lee, S.J.; Gardiner, A.S.; Smith, T.; Sahoo, P.K.; Patel, P.; Thames, E.; Rodriguez, R.; Taylor, R.; Yoo, S.; et al. Axonal localization of neuritin/CPG15 mRNA is limited by competition for HuD binding. J. Cell Sci. 2017, 130, 3650–3662. [Google Scholar] [CrossRef]
- Laedermann, C.J.; Pertin, M.; Suter, M.R.; Decosterd, I. Voltage-gated sodium channel expression in mouse DRG after SNI leads to re-evaluation of projections of injured fibers. Mol. Pain. 2014, 10, 19. [Google Scholar] [CrossRef]
- Sanna, M.D.; Ghelardini, C.; Galeotti, N. HuD-mediated distinct BDNF regulatory pathways promote regeneration after nerve injury. Brain Res. 2017, 1659, 55–63. [Google Scholar] [CrossRef]
- Nutter, C.A.; Kuyumcu-Martinez, M.N. Emerging roles of RNA-binding proteins in diabetes and their therapeutic potential in diabetic complications. Wiley Interdiscip. Rev. RNA 2018, 9. [Google Scholar] [CrossRef]
- Magro, M.G.; Solimena, M. Regulation of β-cell function by RNA-binding proteins. Mol. Metab. 2013, 2, 348–355. [Google Scholar] [CrossRef]
- Juan-Mateu, J.; Rech, T.H.; Villate, O.; Lizarraga-Mollinedo, E.; Wendt, A.; Turatsinze, J.V.; Brondani, L.A.; Nardelli, T.R.; Nogueira, T.C.; Esguerra, J.L.; et al. Neuron-enriched RNA-binding Proteins Regulate Pancreatic Beta Cell Function and Survival. J. Biol. Chem. 2017, 292, 3466–3480. [Google Scholar] [CrossRef]
- Paukku, K.; Backlund, M.; De Boer, R.A.; Kalkkinen, N.; Kontula, K.K.; Lehtonen, J.Y. Regulation of AT1R expression through HuR by insulin. Nucleic Acids Res. 2012, 40, 5250–5261. [Google Scholar] [CrossRef] [Green Version]
- Amadio, M.; Pascale, A.; Cupri, S.; Pignatello, R.; Osera, C.; D Agata, V.; D Amico, A.G.; Leggio, G.M.; Ruozi, B.; Govoni, S.; et al. Nanosystems based on siRNA silencing HuR expression counteract diabetic retinopathy in rat. Pharmacol. Res. 2016, 111, 713–720. [Google Scholar] [CrossRef]
- Li, X.; Zeng, L.; Cao, C.; Lu, C.; Lian, W.; Han, J.; Zhang, X.; Zhang, J.; Tang, T.; Li, M. Long noncoding RNA MALAT1 regulates renal tubular epithelial pyroptosis by modulated miR-23c targeting of ELAVL1 in diabetic nephropathy. Exp. Cell. Res. 2017, 350, 327–335. [Google Scholar] [CrossRef]
- Ishihara, E.; Nagahama, M.; Naruse, S.; Semba, R.; Miura, T.; Usami, M.; Narita, M. Neuropathological alteration of aquaporin 1 immunoreactive enteric neurons in the streptozotocin-induced diabetic rats. Auton. Neurosci. 2008, 138, 31–40. [Google Scholar] [CrossRef]
- De Mello, S.T.; de Miranda Neto, M.H.; Zanoni, J.N.; Furlan, M.M. Effects of insulin treatment on HuC/HuD, NADH diaphorase, and nNOS-positive myoenteric neurons of the duodenum of adult rats with acute diabetes. Dig. Dis. Sci. 2009, 54, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Lee, H.; Kang, H.; Shin, J.J.; Tak, H.; Kim, W.; Gorospe, M.; Lee, E.K. RNA-binding protein HuD reduces triglyceride production in pancreatic β cells by enhancing the expression of insulin-induced gene 1. Biochim. Biophys. Acta. 2016, 1859, 675–685. [Google Scholar] [CrossRef]
- Radu, B.M.; Iancu, A.D.; Dumitrescu, D.I.; Flonta, M.L.; Radu, M. TRPV1 properties in thoracic dorsal root ganglia neurons are modulated by intraperitoneal capsaicin administration in the late phase of type 1 autoimmune diabetes. Cell. Mol. Neurobiol. 2013, 33, 187–196. [Google Scholar] [CrossRef]
- Radu, B.M.; Dumitrescu, D.I.; Marin, A.; Banciu, D.D.; Iancu, A.D.; Selescu, T.; Radu, M. Advanced type 1 diabetes is associated with ASIC alterations in mouse lower thoracic dorsal root ganglia neurons. Cell Biochem. Biophys. 2014, 68, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Kojima, R.; Ito, M. Strain differences in the diabetogenic activity of streptozotocin in mice. Biol. Pharm. Bull. 2006, 29, 1110–1119. [Google Scholar] [CrossRef] [PubMed]
- Jolivalt, C.G.; Frizzi, K.E.; Guernsey, L.; Marquez, A.; Ochoa, J.; Rodriguez, M.; Calcutt, N.A. Peripheral Neuropathy in Mouse Models of Diabetes. Curr. Protoc. Mouse Biol. 2016, 6, 223–255. [Google Scholar]
- Rossini, A.A.; Appel, M.C.; Williams, R.M.; Like, A.A. Genetic influence of the streptozotocin-induced insulitis and hyperglycemia. Diabetes 1977, 26, 916–920. [Google Scholar] [CrossRef]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015, 70, 5.47.1–5.47.20. [Google Scholar] [PubMed]
- Romanovsky, D.; Wang, J.; Al-Chaer, E.D.; Stimers, J.R.; Dobretsov, M. Comparison of metabolic and neuropathy profiles of rats with streptozotocin-induced overt and moderate insulinopenia. Neuroscience 2010, 170, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Kurlawalla-Martinez, C.; Stiles, B.; Wang, Y.; Devaskar, S.U.; Kahn, B.B.; Wu, H. Insulin hypersensitivity and resistance to streptozotocin-induced diabetes in mice lacking PTEN in adipose tissue. Mol. Cell. Biol. 2005, 25, 2498–2510. [Google Scholar] [CrossRef]
- Kahraman, S.; Aydin, C.; Elpek, G.O.; Dirice, E.; Sanlioglu, A.D. Diabetes-resistant NOR mice are more severely affected by streptozotocin compared to the diabetes-prone NOD mice: Correlations with liver and kidney GLUT2 expressions. J. Diabetes Res. 2015, 2015, 450128. [Google Scholar] [CrossRef]
- Tozzo, E.; Gnudi, L.; Kahn, B.B. Amelioration of insulin resistance in streptozotocin diabetic mice by transgenic overexpression of GLUT4 driven by an adipose-specific promoter. Endocrinology 1997, 138, 1604–1611. [Google Scholar] [CrossRef]
- Ventura-Sobrevilla, J.; Boone-Villa, V.D.; Aguilar, C.N.; Román-Ramos, R.; Vega-Avila, E.; Campos-Sepúlveda, E.; Alarcón-Aguilar, F. Effect of varying dose and administration of streptozotocin on blood sugar in male CD1 mice. Proc. West Pharmacol. Soc. 2011, 54, 5–9. [Google Scholar]
- Callaghan, B.C.; Cheng, H.T.; Stables, C.L.; Smith, A.L.; Feldman, E.L. Diabetic neuropathy: Clinical manifestations and current treatments. Lancet Neurol. 2012, 11, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Calcutt, N.A.; Freshwater, J.D.; Mizisin, A.P. Prevention of sensory disorders in diabetic Sprague-Dawley rats by aldose reductase inhibition or treatment with ciliary neurotrophic factor. Diabetologia 2004, 47, 718–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, E.P.; Coppey, L.J.; Dake, B.; Yorek, M.A. Treatment of streptozotocin-induced diabetic rats with alogliptin: Effect on vascular and neural complications. Exp Diabetes Res. 2011, 2011, 810469. [Google Scholar] [CrossRef]
- Murakami, T.; Iwanaga, T.; Ogawa, Y.; Fujita, Y.; Sato, E.; Yoshitomi, H.; Sunada, Y.; Nakamura, A. Development of sensory neuropathy in streptozotocin-induced diabetic mice. Brain Behav. 2013, 3, 35–41. [Google Scholar] [CrossRef]
- Beiswenger, K.K.; Calcutt, N.A.; Mizisin, A.P. Dissociation of thermal hypoalgesia and epidermal denervation in streptozotocin-diabetic mice. Neurosci. Lett. 2008, 442, 267–272. [Google Scholar] [CrossRef]
- Sanna, M.D.; Quattrone, A.; Galeotti, N. Silencing of the RNA-binding protein HuR attenuates hyperalgesia and motor disability in experimental autoimmune encephalomyelitis. Neuropharmacology 2017, 123, 116–125. [Google Scholar] [CrossRef]
- Ohnishi, M.; Yukawa, R.; Akagi, M.; Ohsugi, Y.; Inoue, A. Bradykinin and interleukin-1β synergistically increase the expression of cyclooxygenase-2 through the RNA-binding protein HuR in rat dorsal root ganglion cells. Neurosci. Lett. 2019, 694, 215–219. [Google Scholar] [CrossRef]
- Sanna, M.D.; Peroni, D.; Quattrone, A.; Ghelardini, C.; Galeotti, N. Spinal RyR2 pathway regulated by the RNA-binding protein HuD induces pain hypersensitivity in antiretroviral neuropathy. Exp. Neurol. 2015, 267, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Sanna, M.D.; Quattrone, A.; Ghelardini, C.; Galeotti, N. PKC-mediated HuD-GAP43 pathway activation in a mouse model of antiretroviral painful neuropathy. Pharmacol. Res. 2014, 81, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Sanna, M.D.; Ghelardini, C.; Galeotti, N. Altered Expression of Cytoskeletal and Axonal Proteins in Oxaliplatin-Induced Neuropathy. Pharmacology 2016, 97, 146–150. [Google Scholar] [CrossRef]
- Clayton, G.H.; Perez, G.M.; Smith, R.L.; Owens, G.C. Expression of mRNA for the elav-like neural-specific RNA binding protein, HuD, during nervous system development. Brain Res. Dev. Brain Res. 1998, 109, 271–280. [Google Scholar] [CrossRef]
- Okano, H.J.; Darnell, R.B. A hierarchy of Hu RNA binding proteins in developing and adult neurons. J. Neurosci. 1997, 17, 3024–3037. [Google Scholar] [CrossRef]
- Georgescu, A.; Popov, D.; Dragan, E.; Dragomir, E.; Badila, E. Protective effects of nebivolol and reversal of endothelial dysfunction in diabetes associated with hypertension. Eur. J. Pharmacol. 2007, 570, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Tuk, B.; Shang, P.; Hekking, I.M.; Fijneman, E.M.; Guijt, M.; Hovius, S.E.; van Neck, J.W. Diabetes-impaired wound healing is improved by matrix therapy with heparan sulfate glycosaminoglycan mimetic OTR4120 in rats. Diabetes 2012, 61, 2633–2641. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tallafuss, A.; Kelly, M.; Gay, L.; Gibson, D.; Batzel, P.; Karfilis, K.V.; Eisen, J.; Stankunas, K.; Postlethwait, J.H.; Washbourne, P. Transcriptomes of post-mitotic neurons identify the usage of alternative pathways during adult and embryonic neuronal differentiation. BMC Genom. 2015, 16, 1100. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.C.; Kim, D.S.; Yang, G.; Zaslavsky, K.; Ha, K.C.; Mok, R.S.; Ross, P.J.; Zhao, M.; Piekna, A.; Wei, W.; et al. MECP2 Is Post-transcriptionally Regulated during Human Neurodevelopment by Combinatorial Action of RNA-Binding Proteins and miRNAs. Cell Rep. 2016, 17, 720–734. [Google Scholar] [CrossRef] [PubMed]
- Radu, B.M.; Osculati, A.M.M.; Suku, E.; Banciu, A.; Tsenov, G.; Merigo, F.; Di Chio, M.; Banciu, D.D.; Tognoli, C.; Kacer, P.; et al. All muscarinic acetylcholine receptors (M1-M5) are expressed in murine brain microvascular endothelium. Sci. Rep. 2017, 7, 5083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubový, P.; Jancálek, R.; Klusáková, I.; Svízenská, I.; Pejchalová, K. Intra- and extraneuronal changes of immunofluorescence staining for TNF-alpha and TNFR1 in the dorsal root ganglia of rat peripheral neuropathic pain models. Cell. Mol. Neurobiol. 2006, 26, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Tsunematsu, H.; Uyeda, A.; Yamamoto, N.; Sugo, N. Immunocytochemistry and fluorescence imaging efficiently identify individual neurons with CRISPR/Cas9-mediated gene disruption in primary cortical cultures. BMC Neurosci. 2017, 18, 55. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mustăciosu, C.C.; Banciu, A.; Rusu, C.M.; Banciu, D.D.; Savu, D.; Radu, M.; Radu, B.M. RNA-Binding Proteins HuB, HuC, and HuD are Distinctly Regulated in Dorsal Root Ganglia Neurons from STZ-Sensitive Compared to STZ-Resistant Diabetic Mice. Int. J. Mol. Sci. 2019, 20, 1965. https://doi.org/10.3390/ijms20081965
Mustăciosu CC, Banciu A, Rusu CM, Banciu DD, Savu D, Radu M, Radu BM. RNA-Binding Proteins HuB, HuC, and HuD are Distinctly Regulated in Dorsal Root Ganglia Neurons from STZ-Sensitive Compared to STZ-Resistant Diabetic Mice. International Journal of Molecular Sciences. 2019; 20(8):1965. https://doi.org/10.3390/ijms20081965
Chicago/Turabian StyleMustăciosu, Cosmin Cătălin, Adela Banciu, Călin Mircea Rusu, Daniel Dumitru Banciu, Diana Savu, Mihai Radu, and Beatrice Mihaela Radu. 2019. "RNA-Binding Proteins HuB, HuC, and HuD are Distinctly Regulated in Dorsal Root Ganglia Neurons from STZ-Sensitive Compared to STZ-Resistant Diabetic Mice" International Journal of Molecular Sciences 20, no. 8: 1965. https://doi.org/10.3390/ijms20081965